4
2 TINJAUAN PUSTAKA
2.1 Probiotik, Prebiotik dan Sinbiotik
Probiotik merupakan mikroorganisme hidup yang ketika diberikan dalam jumlah cukup dapat memberikan manfaat kesehatan pada inang (FAO/WHO 2001). Menurut Lisal (2005), kriteria dalam pemilihan probiotik yaitu sebaiknya merupakan mikroflora normal usus, bersifat nonpatogenik dan nontoksik bagi inang, toleran terhadap asam lambung dan garam empedu, mampu menempel dan berkoloniasi dalam usus, bersifat antagonistik terhadap patogen, memiliki pengaruh yang menguntungkan bagi inang, dan memiliki jumlah serta viabilitas yang tinggi. Dalam akuakultur, jenis probiotik yang dievaluasi dan digunakan lebih luas dibandingkan hewan terestrial, baik dalam bentuk monospesies maupun multispesies. Jenis-jenis probiotik tersebut memiliki mekanisme aksi yang berbeda diantaranya mampu meningkatkan efisiensi pakan dan bobot tubuh, memberi proteksi dalam melawan patogen melalui kompetisi ruang, produksi asam organik (asam formik, asam asetat dan asam laktat), produksi hidrogen peroksida dan beberapa bahan lainnya seperti antibiotik, bakteriosin, siderophores, lisozim serta memodulasi respons fisiologis dan imunologis ikan. Beberapa genus bakteri yang telah diteliti sebagai probiotik yaitu Bacillus, Lactobacillus, Lactococcus, Aeromonas, Shewanella, Vibrio, Carnobacterium, dan Clostridium (Nayak 2010).
Prebiotik adalah bahan makanan yang tidak dapat dicerna yang bermanfaat untuk menstimulasi pertumbuhan dan aktivitas bakteri tertentu (bakteri menguntungkan) di dalam usus. Studi mengenai prebiotik pada hewan akuatik berkaitan dengan efek terhadap pertumbuhan, konversi pakan, mikrobiota usus, resistensi terhadap patogen dan parameter imunitas. Prebiotik umumnya merupakan karbohidrat yang tidak dapat dicerna dalam saluran pencernaan inang. Karbohidrat dikelompokkan berdasarkan berat molekul atau tingkat polimerasinya (jumlah unit monosakarida), menjadi monosakarida, oligosakarida dan polisakarida (Tabel 1). Oligosakarida tidak dapat dicerna (non-digestible oligosaccharide) memiliki konfigurasi atom C dalam unit monosakarida yang membuat ikatan glikosidiknya tidak dapat dicerna oleh aktivitas hidrolisis dari enzim pencernaan manusia atau hewan. Prebiotik yang umum digunakan di akuakutur sampai sekarang meliputi inulin, fructooligosaccarides (FOS), short-cain fructooligosaccharides (scFOS), mannanoligosaccharides (MOS), galactooligosaccharides (GOS), xylooligosaccharides (XOS), arabinoxylooligosaccharides (AXOS), isomaltooligosaccharides (IMO) dan GroBiotic –A (Ringo et al. 2010).
Sinbiotik merupakan kombinasi yang seimbang dari probiotik dan prebiotik. Aplikasi sinbiotik memberikan hasil yang lebih baik dibandingkan pemberian probiotik dan prebiotik secara terpisah. Efek sinergis dari Bacillus OJ dan IMO mampu meningkatkan populasi mikrobial, respons imunitas dan resistensi terhadap penyakit pada L. vanammei, lebih baik dibandingkan dengan aplikasi terpisah dari Bacillus OJ dan IMO (Li et al. 2009). Hasil serupa juga dilaporkan oleh Rodriguez-Estrada et al. (2009) yang menyebutkan bahwa aplikasi Enterococcus faecalis dan MOS pada ikan salmon mampu meningkatkan respons imun dan sintasan ikan terhadap infeksi V. (L.) anguillarum.
5 Tabel 1 Klasifikasi karbohidrat (Subandiyono dan Hastuti 2009)
No. Kelompok Jenis Contoh
1 Monosakarida a. Triosa (C3H6O3) Gliseraldehida; Dihidroksiaseton (1 unit b. Tetrosa (C4H8O4) Eritrosa
glukosa) c. Pentosa (C5H10O5) Rribosa; Arabinosa; Xilosa; Xilulosa d. Heksosa (C6H12O6) Glukosa; Galaktosa; Mannosa; Fruktosa 2 Oligosakarida a. Disakarida (C12H22O11) Sukrosa; Laktosa; Maltosa; Selobiosa
(2-10 unit b. Trisakarida (C18H32O16) Rafinosa glukosa) c. Tetrasakarida (C24H42O16) Stasilosa
d. Pentasakarida (C30H52O26) Verbaskosa
3 Polisakarida a. Homo-polisakarida 1. Pentosan (Araban; Xilan) ( >10 unit
glukosa)
(glukosa dengan jenis yang sama)
2. Heksosan (Glukan [Starch, Dekstrin, Glikogen, Selulosa]; Fruktan [Inulin, Levan]; Galaktan; Manan)
b. Hetero-polisakarida (glukosa dengan jenis
Pektin; Hemiselulosa; Gum; Mucilage; Mukopolisakarida
yang berbeda) 4 Persenyawaan a. Khitin
khusus b. Lignin
2.2 Sistem Imunitas Krustasea
Mekanisme pertahanan tubuh krustasea kurang berkembang dibandingkan ikan bersirip (finfish) dan vertebrata lainnya. Krustasea tidak memiliki memori adaptif dan hanya bergantung pada sistem pertahanan nonspesifik (Roch 1999). Sistem pertahanan tersebut meliputi pertahanan seluler berupa aktivitas sel-sel hemosit (fagositosis, enkapsulasi, dan pembentukkan nodul), serta pertahanan humoral berupa aktivasi dan pelepasan molekul-molekul penting yang tersimpan dalam hemosit (protein antikoagulan, aglutinin, enzim phenoloxidase [PO], peptida antimikrobial, protease inhibitor, dan sebagainya) (Jiravanichpaisal et al. 2006; Holmblad dan Soderhall 1999). Mekanisme pertahanan tubuh krustasea dijelaskan pada Gambar 1 (Smith et al. 2003).
Berdasarkan keberadaan granular pada sitoplasma, terdapat tiga tipe hemosit pada krustasea yaitu sel hialin (nongranular), semi granular dan sel granular (Gambar 2). Fungsi dari masing-masing hemosit seperti pada Tabel 2 (Soderhall dan Cerenius 1992). Fagositosis merupakan mekanisme pertahanan sel yang paling umum dengan cara menelan dan menghancurkan patogen dan partikel asing yang masuk ke dalam tubuh. Penghancuran material yang difagosit melibatkan produksi intraseluler berupa radikal bebas. Selama proses kontak dan pengenalan dengan patogen, enzim inang seperti NADPH-oksidase menjadi aktif yang ditandai dengan meningkatnya konsumsi oksigen dan menghasilkan radikal bebas diantaranya anion superoksida (O2-) dan hidrogen peroksida (H2O2) (Munoz et al. 2000; Rodriguez
dan Le Moullac 2000). Radikal bebas ini dapat langsung membunuh organisme yang menyerang, berkombinasi dengan senyawa-senyawa nitrogen (nitric oxide) atau bersinergi dengan lisozim (Roch 1999). Enkapsulasi dan pembentukan nodul
6
merupakan kumpulan hemosit yang saling bekerja sama untuk menangani partikel asing. Enkapsulasi terjadi bila inang diserang oleh partikel yang berukuran besar, sedangkan nodul terbentuk bila inang diserang oleh partikel kecil dalam jumlah banyak (Gambar 3) (Soderhall dan Cerenius 1992).
Gambar 1 Mekanisme pertahanan tubuh nonspesifik pada krustasea
Gambar 2 Klasifikasi sel hemosit: a) hialin, b) semigranular, c) granular (Giulianini et al. 2007) β-1,3-glucan Peptidoglican Live bacteria Bacterial antigen Phenolic
compounds Quinones Melanin
Cell adhesion Release of reactive O2 Opsonization Encapsulation Phenoloxidase (PO) Prophenoloxidase (proPO) Active serine proteinase (ppA) Antibacterial peptides Peroxinectin Phagocytosis Inactive serine proteinase (proppA) Granular
Semi granular hyaline
β-1,3-glucan binding protein (βGBP)
Degranulation
Degranulation
7 Tabel 2 Fungsi berbagai tipe sel hemosit pada sistem imunitas krustasea
Tipe hemosit
Fungsi
Fagositosis Enkapsulasi Sitotoksisitas Sistem aktivasi ProPO Hialin Semigranular Granular Ya Terbatas Tidak Tidak Ya Sangat terbatas -* Ya Ya Tidak Ya Ya
*Belum ada informasi
Gambar 3 Mekanisme pertahanan seluler: a) fagositosis, b) enkapsulasi, c) pembentukan nodul
Sistem prophenoloxidase (proPO) merupakan bagian yang dominan dari mekanisme pertahanan tubuh krustasea, disimpan dan diproduksi oleh sel semigranular dan granular yang diaktifkan oleh kehadiran sejumlah kecil mikroba. Komponen awal dari sistem proPO adalah PO yang ketika aktif akan mengoksidasi senyawa fenol menjadi kuinolon dan secara spontan membentuk melanin sebagai produk akhir. Melanin beserta produk antaranya merupakan senyawa yang sangat reaktif yang berperan diantaranya dalam inaktivasi dan mencegah penyebaran partikel asing dalam tubuh melalui penghambatan produk ekstraseluler mikroorganisme (proteinase dan khitinase). Sistem proPO berpengaruh terhadap tingkah laku sel, perbanyakan dan atau aktivasi molekul-molekul penting yang fungsional serta netralisasi agen penginfeksi (Smith et al. 2003).
2.3 Infectious Myonecrosis (IMN)
Wabah IMN pertama kali teridentifikasi pada budidaya udang vaname (Litopenaeus vanammei) di timur laut Brazil pada tahun 2004 dan ditemukan di Indonesia pada awal tahun 2006. Hasil analisis subsekuen membuktikan bahwa virus IMN (IMNV) yang menginfeksi udang vaname di Indonesia memiliki kemiripan sekuen asam nukleat 99,6% dengan IMNV yang menginfeksi di Brazil (Senapin et al. 2007). IMNV menginfeksi postlarva, juvenil dan subadult pada pembesaran udang vaname. Mewabahnya penyakit ini diduga berkaitan dengan stress fisik dan lingkungan seperti salinitas dan suhu ekstrim, penanganan udang,
8
serta penggunaan pakan kualitas rendah. Infeksi IMNV memperlihatkan gejala nekrosis dari mulai ringan sampai berat terutama di bagian abdomen dan ekor yang dapat berkembang menjadi kemerahan (Gambar 4) (Lightner et al. 2004).
Gambar 4 Gejala klinis udang vaname yang terinfeksi IMNV: a) Lightner et al. 2004, b) Poulos et al. 2006
Hasil histopatologi, udang yang terinfeksi IMNV memperlihatkan lesi di otot skeletal meliputi multifocal necrosis, kongesi hemosit, inflamasi fibrosis, fagositosis dan munculnya badan inklusi sitoplasmik. Hasil in situ hybridisation (ISH) pada L. vannamei menunjukkan bahwa otot skeletal merupakan organ target utama dari infeksi IMNV. Hal ini yang menyebabkan penyakit IMN bersifat kronis dan kematian terjadi secara perlahan. Selain otot skeletal, IMNV juga ditemukan di organ limfoid, usus bagian belakang, insang serta sel fagositik dalam hepatopankreas dan hati. L vanammei merupakan inang utama dari IMNV, meskipun mampu menginfeksi L. stylirostris dan Penaeus monodon, namun tidak menyebabkan kematian sampai dengan empat minggu pengamatan setelah infeksi (Tang et al. 2005).
Berdasarkan analisis imunologi, perubahan parameter imunologis yang signifikan hanya terjadi pada udang di tahap akhir infeksi IMNV, ketika pemulihan diri tidak dimungkinkan lagi. Infeksi IMNV menyebabkan peningkatan apoptosis pada hemosit (8 kali), titer aglutinasi (16 kali), produksi anion superoksida dari hemosit (50%), dan aktivitas antimikrobial dari hemolimph (21 kali melawan Micrococcus luteus). Infeksi IMNV juga menyebabkan penurunan total hemosit (30%) dan persentase granulosit sirkular (7%) (Costa et al. 2009).
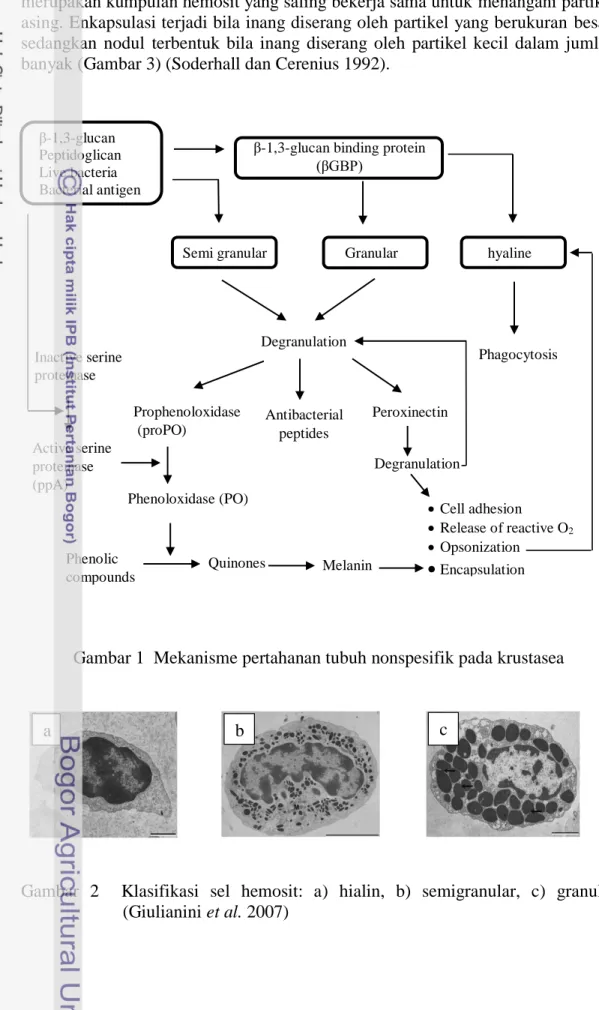
Agen penyebab penyakit IMN adalah virus berbentuk ikosahedral dengan diameter 40 nm, memiliki genom tunggal, double-stranded (dsRNA) dengan panjang molekul 7560 bp (Gambar 5). Hasil analisis filogenetik IMNV berdasarkan RDA-dependent dari gen RNA polimerase (RdRp), IMNV memiliki kemiripan dengan virus Giardia lamblia, yang merupakan anggota dari family Totiviridae. Berdasarkan hal ini, IMNV mungkin merupakan anggota unik dari Totiviridae atau mungkin merepresentasikan family virus dsRNA baru yang menginfeksi inang invertebrata (Poulos et al. 2006).
9
Gambar 5 Bentuk ikosahedral dari IMNV: a) transmisi elektron mikrograf, fraksi gradien calsium chloride, diwarnai dengan phosphotungstic acid 2%, garis menunjukkan 100 nm (Poulos et al. 2006), b) Rekonstruksi 3-dimensi virion IMNV dengan resolusi 8.0-Å (Tang et al. 2008)
2.4 Ubi Jalar (Ipomoea batatas)
Ubi jalar merupakan tanaman asli dari Amerika Tengah dan Amerika Selatan. Tanaman ini mulai menyebar ke seluruh dunia, terutama negara-negara beriklim tropis, pada abad ke 16. Orang-orang Spanyol menyebarkan ubi jalar ke kawasan Asia terutama Filipina, Jepang dan Indonesia. Data tahun 2009 menunjukkan produksi ubi jalar Indonesia menempati urutan ke 4 setelah China, Uganda dan Nigeria. Ubi jalar termasuk dalam ordo Solanaceae, family Convolvulaceae, genus Ipomoea dan spesies Ipomoea batatas serta memiliki nama binomial Ipomoea batatas (L) Lam. Tanaman ini memiliki warna kulit yang bervariasi antara kuning, oranye, merah, coklat, ungu dan abu-abu kecoklatan. Warna daging juga bervariasi mulai dari abu-abu kecoklatan, putih, merah, merah muda, ungu, kuning, sampai oranye, tergantung jenis dan banyaknya pigmen yang terdapat di dalamnya. Varietas umbi warna putih dan kuning pucat memiliki rasa manis dan kadar air lebih rendah dibandingkan dengan varietas berwarna merah, merah muda dan oranye (Wikipedia 2012). Menurut North Carolina Sweet Potato Commission (2013), terdapat ratusan varietas ubi jalar (Gambar 6), kebanyakan di produksi dalam jumlah kecil serta hanya dijual oleh petani lokal. Secara umum, ubi jalar mengandung karbohidrat yang tinggi (20,1%), terdiri dari pati (12,7%), gula (4,2%), dan serat (3,0%). Ubi jalar juga mengandung protein (1,6%) dan lemak (0,1%) yang rendah (Wikipedia 2012).
Komposisi kimia ubi jalar bervariasi tergantung pada waktu panen, varietas dan proses pengolahan. Kandungan gula yang terdapat pada ubi jalar varietas sukuh terdiri dari fruktosa, glukosa, sukrosa, maltose dan maltotriosa (Marlis 2008). Roxas et al. (1985) menyebutkan bahwa ubi jalar varietas Kinabakab, Tinipay, BNAS 51 dan G113-2b hanya tediri dari monosakarida dan sukrosa. Pengukusan dapat meningkatkan konsentrasi gula dalam ubi jalar dibandingkan dengan kondisi mentahnya. Hal ini disebabkan oleh pemecahan struktur pati karena pengaruh panas dan air, menjadi molekul glukosa yang lebih sederhana (Marlis 2008).
10
Gambar 6 Varietas ubi jalar yang banyak diminati di North Carolina Sweet Potato Commission