TINJAUAN PUSTAKA
Vermicomposting
Dominguez et al. (1997a) mendefinisikan vermicomposting sebagai proses dekomposisi bahan organik yang melibatkan kerjasama antara cacing tanah dan mikroorganisme. Mikroorganisme yang berperan dalam proses vermicomposting terutama bakteri, fungi, dan actinomycetes.
Selama proses vermicomposting, nutrisi pada tumbuhan yang penting seperti nitrogen, kalium, dan fosfor yang terdapat di dalam bahan makanan diubah melalui aktivitas mikroorganisme menjadi bentuk yang lebih mudah diserap oleh tumbuhan (Ndegwa & Thompson. 2001). Pada proses ini cacing tanah mengubah aktivitas mikroorganisme (Aira et al. 2002), sehingga laju mineralisasi bahan-bahan organik bertambah cepat (Albanell et al. 1988). Beberapa enzim yang terlibat di dalam dekomposisi bahan organik adalah dehidrogenase, protease, glukosidase, dan fosfatase (Lazcano et al. 2008). Vermicomposting menghasilkan dua manfaat utama, yaitu biomassa cacing tanah dan vermikompos (Sharma et al. 2005).
Vermikompos merupakan bahan organik seperti tanah yang memiliki struktur, porositas, aerasi, drainase, dan kapasitas menahan kelembaban yang sangat baik (Dominguez et al. 1997a). Vermikompos mengandung banyak aktivitas, populasi, dan keanekaragaman mikroorganisme. Vermikompos juga mengandung beberapa enzim seperti protease, amilase, lipase, selulase, dan kitinase (Subler et al. 1998), serta zat pengatur tumbuh seperti giberelin, sitokinin, dan auksin (Tomatti et al. 1988).
Syarat-syarat biologi cacing tanah yang digunakan dalam proses
vermicomposting terdiri atas: tingkat produksi kokon yang tinggi, waktu
perkembangan kokon yang pendek, keberhasilan penetasan kokon yang tinggi, dan memiliki laju reproduksi yang tinggi (Bhattacharjee & Chaudhuri 2002). Selain itu, tingkat konsumsi bahan organik yang tinggi pada cacing tanah dan toleran terhadap perubahan lingkungan yang luas juga merupakan sebagian syarat biologi cacing tanah yang dapat dimanfaatkan untuk mendekomposisi bahan organik (Edwards 1998; Dominguez et al. 2000).
Beberapa spesies cacing tanah yang memenuhi syarat biologi dan digunakan dalam proses vermicomposting adalah: E. fetida (Albanell 1988), (Reinecke et al. 1992), (Gunadi et al. 2003), (Garg et al. 2005), (Aira et al. 2006a); E. andrei (Dominguez et al. 2000); L. rubellus (Delgado et al. 1995); L. terrestris, Eudrilus
eugeniae (Banu et al. 2008); Perionyx excavatus, dan P. sansibaricus (Suthar
2007a; Suthar & Singh 2008).
Faktor-faktor yang mempengaruhi proses vermicomposting
Proses vermicomposting dalam kaitannya dengan pertumbuhan dan kelangsungan hidup cacing tanah dipengaruhi oleh beberapa faktor, misalnya: suhu, kelembaban, rasio C:N, pH, aerasi, dan makanan. Pengaruh faktor-faktor tersebut bervariasi pada setiap spesies cacing tanah.
Suhu pada substrat mempengaruhi pertumbuhan cacing tanah. Pada kisaran suhu 20-29 oC cacing tanah tumbuh dan berkembang dengan maksimal (Kaplan et
al. 1980). Hou et al. (2005) menemukan bahwa laju pertumbuhan cacing tanah
tercepat pada suhu 20 oC. Akan tetapi kebutuhan suhu pada masing-masing spesies cacing tanah berbeda-beda. Di awal proses vermicomposting terjadi peningkatan suhu, dan di akhir periode suhu menurun.
Berat tubuh cacing tanah terdiri atas 75-90% air (Edwards & Lofty 1972), maka kekurangan air merupakan masalah besar dalam kehidupan cacing tanah. Cacing tanah dapat berpindah ke tanah yang lebih dalam jika permukaan tanah terlalu kering. Kelembaban yang rendah dapat menyebabkan cacing tanah menjadi pasif atau mengalami fase diapause (Gerard 1967). Menurut Reinecke dan Venter (1987), kelembaban substrat yang lebih disukai oleh cacing tanah dewasa berkisar antara 65-75%, tetapi juvenil lebih dapat bertahan hidup pada kelembaban dengan kisaran 65-70%. Laju pertumbuhan cacing tanah tertinggi terdapat pada kelembaban 75% (Gunadi et al. 2003). Dominguez et al. (1997a) menemukan bahwa kisaran kelembaban yang terbaik adalah 80-90%, dengan kisaran optimum sebesar 85%. Kebutuhan cacing tanah akan kelembaban media bervariasi pada berbagai spesies dan daya adaptasi masing-masing spesies tersebut. Kelembaban media dapat dipertahankan dengan penambahan air pada media dan menyediakan bahan makanan yang mengandung banyak air.
Rasio C:N pada substrat (media biak yang terdiri atas kotoran ternak dan sampah organik) mencerminkan mineralisasi dan stabilisasi sampah organik selama proses vermicomposting (Suthar & Singh 2008). Laju dekomposisi tertinggi terdapat pada substrat dengan rasio C:N sebesar 20 (Hou et al. 2005). Ndegwa dan Thompson (2000) menemukan bahwa cacing tanah dapat tumbuh lebih baik ketika rasio C:N substrat 25. Cacing tanah lebih memilih sampah organik yang memiliki rasio C:N lebih rendah sebagai makanannya (Moody et al. 1995). Semakin rendah rasio C:N, maka semakin tinggi laju dekomposisi sampah organik (Pramanik et al. 2007).
Substrat yang dikonsumsi cacing tanah memliki kisaran nilai pH tertentu. Substrat yang terlalu asam atau basa tidak cocok untuk kelangsungan hidup cacing tanah. Kisaran nilai pH optimum bagi cacing tanah antara 6.5 dan 8.5 (Hou
et al. 2005). Pada umumnya spesies cacing tanah lebih memilih substrat yang
memiliki pH sekitar 7.0 (Arrhenius 1921). Singh et al. (2005) menemukan bahwa pH substrat awal yang mendekati netral mengoptimalkan stabilisasi bahan organik dalam waktu yang singkat. Substrat awal yang memiliki pH sangat asam tidak sesuai digunakan dalam proses vermicomposting.
Proses vermicomposting dapat berlangsung dengan baik dalam kondisi aerob. Cacing tanah memerlukan oksigen untuk bernafas dan sangat sensitif terhadap kondisi anaerob. Laju respirasinya melemah jika konsentrasi oksigen di dalam substrat rendah (Edwards & Bohlen 1996). Pergerakan cacing tanah dapat menciptakan aerasi pada medianya. Untuk meningkatkan aerasi, perlu dilakukan pembalikan substrat.
Kualitas dan kuantitas bahan makanan mempengaruhi pertumbuhan dan reproduksi cacing tanah dalam proses vermicomposting (Edwards et al. 1988). Kualitas bahan makanan pada substrat awal sangat mempengaruhi biomassa cacing tanah (Suthar 2007b). Berkurangnya biomassa cacing tanah dapat disebabkan oleh berkurangnya bahan makanan di dalam media biak (Garg et al. 2005). Pertumbuhan cacing tanah dibatasi oleh ketersediaan sumber karbon pada bahan makanannya (Tiunov & Scheu 2004). Cacing tanah yang mengkonsumsi bahan makan dengan rasio C:N rendah, lebih banyak menggunakan energinya untuk pertumbuhan daripada untuk reproduksi (Aira et al. 2006a).
Taksonomi Cacing Tanah
Cacing tanah adalah invertebrata darat yang termasuk ke dalam Filum Annelida, Kelas Clitellata, Sub Kelas Oligochaeta (Brusca & Brusca 2003). Cacing tanah terbagi ke dalam 5 famili, yaitu Moniligastridae, Megascolecidae, Eudrilidae, Glossoscolecidae, dan Lumbricidae.
Klasifikasi Cacing Tanah
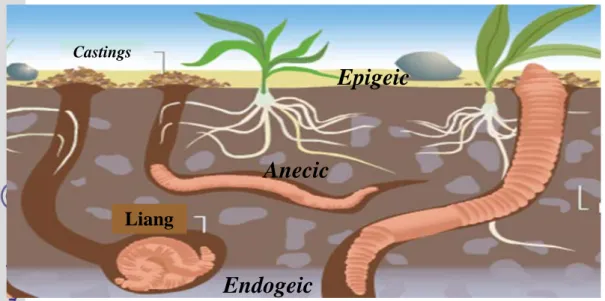
Spesies cacing tanah yang berbeda memiliki sejarah hidup yang berbeda dan menempati relung ekologi yang berbeda. Lee (1985) mengelompokkan spesies cacing tanah ke dalam tiga kategori ekologi berdasarkan strategi mencari makanan dan membuat liang, yaitu spesies epigeic, endogeic, dan anecic (Gambar 1).
Cacing tanah epigeic pada dasarnya merupakan cacing tanah penghuni sampah. Cacing tanah ini hidup di dalam atau dekat permukaan sampah dan memakan sampah organik yang kasar, serta sejumlah sampah yang belum terurai. Cacing tanah epigeic membuat liang ephemeral ke dalam tanah mineral selama periode diapause. Tubuhnya berukuran kecil dan berpigmen. Laju metabolisme dan reproduksinya tinggi. Hal itu menggambarkan daya adaptasinya tinggi terhadap perubahan kondisi lingkungan pada permukaan tanah. Beberapa spesies cacing tanah yang termasuk ke dalam kategori ini adalah L. rubellus, E. fetida,
E. andrei, Dendrobaena rubida, Eudrilus eugeniae, Perionyx excavatus, dan Eiseniella tetraedra (Bouche 1977; Lee 1985).
Cacing tanah endogeic hidup di dalam tanah yang lebih dalam dan memakan tanah serta kumpulan bahan-bahan organik (Gambar 1). Cacing tanah jenis ini tidak memiliki pigmen tubuh, dan membuat liang horizontal yang bercabang ke dalam. Cacing tanah endogeic tidak memiliki pengaruh yang besar dalam penguraian sampah karena cacing tanah ini memakan bahan-bahan di bawah permukaan tanah. Beberapa spesies cacing tanah yang termasuk ke dalam kategori ini adalah Allolobophora caliginosa, A. rosea, Octolasion cyaneum (Bouche 1977), Metaphire posthuma, dan Octochaetona thurstoni (Ismail 1997).
Cacing tanah jenis anecic hidup di dalam sistem liang vertikal yang lebih permanen, yang dapat meluas beberapa meter ke dalam tanah (Gambar 1). Cacing tanah jenis ini dapat di temukan di dalam liang yang dangkal atau dalam tergantung pada kondisi tanah yang baik sebagai habitatnya. Cacing tanah anecic mengeluarkan sisa pencernaannya (casting) pada permukaan tanah dan muncul di malam hari untuk memakan sampah pada permukaan tanah, kotoran, dan bahan organik lain yang diturunkan ke dalam liangnya. Laju reproduksinya relatif lambat, terbukti dari produksi kokonnya. Cacing tanah anecic memiliki peran yang sangat besar dalam dekomposisi bahan organik, siklus makanan, dan pembentukan tanah (Lavelle 1988). Lumbricus terrestris, Aporrectodea
trapezoids, dan Allolobophora longa termasuk dalam kelompok kategori ini.
Biologi Cacing Tanah
Karakteristik cacing tanah adalah tubuhnya bersegmen, dan memiliki sedikit seta pada seluruh segmen tubuh. Cacing tanah dewasa memiliki klitelum yang terletak di bagian anterior tubuh. Klitelum merupakan bagian kelenjar epidermis segmen tubuh yang mengalami perkembangan, terdiri atas kelenjar epidermis yang menebal, terutama di bagian dorsal dan lateral tubuh. (Edwards & Lofty 1972). Pada umumnya klitelum berwarna lebih cerah daripada segmen yang lain. Gambar 1 Klasifikasi cacing tanah berdasarkan kategori ekologi (Lee 1985)
Epigeic
Endogeic
Anecic
LiangSistem reproduksi
Cacing tanah bersifat hermafrodit, tetapi fertilisasi tidak dapat terjadi pada diri sendiri. Pada umumnya individu cacing tanah dewasa melakukan reproduksi silang sebelum menghasilkan kokon (Gambar 2a). Beberapa spesies cacing tanah melakukan reproduksi pada permukaan tanah, dan beberapa spesies melakukannya di bawah tanah (Edwards & Lofty 1972).
Metode kopulasi untuk seluruh spesies cacing tanah tidak sama. Pada cacing tanah yang tergolong famili Lumbricidae, ketika akan melakukan perkawinan dua spesies cacing tanah saling berdekatan dengan mendeteksi mukus yang dikeluarkan oleh bagian ventral tubuhnya bersama-sama. Ujung kepala cacing tanah terletak pada arah yang berlawanan. Keduanya saling mendekatkan diri pada daerah pembukaan spermateka dimana daerah klitelum salah satu cacing tanah menyentuh permukaan pembukaan spermateka yang lainnya. Pada saat kopulasi, kedua cacing tanah tidak sensitif dalam merespon rangsangan luar seperti sentuhan dan cahaya. Banyak mukus yang disekresikan sehingga masing-masing cacing tanah diselubungi oleh mukus antara segmen sembilan dan sisi posterior klitelum, mukus-mukus tersebut saling melekat (Edwards & Lofty 1972).
Sebuah celah semen terbentang dari gonofor jantan sampai klitelum dan nampak seperti benang. Tiap-tiap celah semen merupakan bagian dari dinding luar tubuh yang melekuk ke dalam akibat dari terbentuknya rangkaian pori-pori oleh kontraksi otot yang terbentang pada lapisan otot longitudinal. Kontraksi otot membawa cairan sperma dari gonofor jantan menuju daerah klitelum. Cairan sperma berkumpul di daerah klitelum, dan akhirnya memasuki spermateka cacing tanah lawannya (Edwards & Lofty 1972).
Cara pemindahan sel sperma pada seluruh spesies cacing tanah tidak sama. Spesies cacing tanah yang tidak termasuk ke dalam famili Lumbricidae memindahkan sel spermanya secara langsung tanpa membentuk selubung mukus. Tembe dan Dubash (1961) menjelaskan kopulasi Pheretima yang memiliki tiga atau empat pasang spermateka. Gonofor jantan saling bersentuhan dengan sepasang celah spermateka paling belakang dan menyalurkan cairan sperma ke dalamnya. Kemudian masing-masing cacing tanah bergerak ke arah belakang, dan
cairan sperma disalurkan ke dalam sepasang spermateka berikutnya sampai seluruhnya terisi.
Setelah kopulasi berlangsung selama satu jam, cacing tanah terpisah, dan masing-masing klitelum mengeluarkan getah mukus yang akhirnya mengeras di sekeliling permukaan luarnya. Ketika getah mukus mengeras, cacing tanah bergerak ke arah belakang, kemudian membuat selubung di sekeliling kepalanya, dan ketika cacing tanah terpisah sempurna, ujung selubung menutup untuk membentuk kokon (Gambar 2b). Kokon mengandung cairan albumin yang diproduksi oleh kelenjar klitelum, ovum, dan spermatozoa yang disalurkan ke dalamnya ketika melewati pembukaan spermateka. Kokon terus dibentuk sampai cairan sperma yang tersedia habis. Fertilisasi terjadi secara eksternal tubuh cacing tanah, di dalam kokon (Edwards & Lofty 1972).
Produksi kokon dapat dipengaruhi oleh beberapa faktor. Keadaan tanah dan jenis cacing tanah yang berbeda menyebabkan perbedaan jumlah kokon yang diproduksi. Cacing tanah memproduksi lebih sedikit kokon pada kondisi tanah yang terlalu kering dan terlalu basah (Gerard 1967). Jenis makanan yang dikonsumsi cacing tanah dewasa juga dapat mempengaruhi produksi kokon. Produksi kokon bergantung pada spesies cacing tanah dan kondisi lingkungan (Evans & Guild 1948). Jumlah kokon yang diproduksi cacing tanah epigeic lebih banyak daripada kokon cacing tanah anecic (Bhattacharjee & Chaudhuri 2002).
Warna kokon berubah sesuai dengan perkembangannya (Gambar 2c). Pada saat terbentuk kokon berwarna keputihan, kemudian berubah menjadi kuning, kehijauan dan kecoklat-coklatan (Edwards & Lofty 1972). Kokon yang berwarna kecoklatan mengindikasikan perkembangan yang matang, dan siap untuk menetas. Penetasan kokon dipengaruhi oleh suhu lingkungan (Gerard 1967; Bhattacharjee & Chaudhuri 2002). Suhu yang lebih tinggi dari 25 oC menurunkan masa inkubasi rata-rata kokon cacing tanah epigeic (Reinecke et al. 1992). Jumlah ovum yang dibuahi di dalam setiap kokon berkisar 1-20 untuk cacing tanah Lumbricidae, tapi sering kali hanya satu atau dua yang bertahan hidup dan menetas menjadi juvenil (Gambar 2d) (Stephenson 1930).
Sistem pencernaan
Sistem pencernaan cacing tanah terdiri atas rongga mulut, faring, esofagus, tembolok, lambung, dan usus. Cacing tanah memperoleh makanan dari bahan organik berupa organ tumbuhan, protozoa, rotifera, nematoda, bakteri, fungi, dan sisa-sisa pembusukan hewan (Edwards & Lofty 1972).
Bagian tubuh cacing tanah dikelompokkan ke dalam tiga daerah berdasarkan pada segmen tubuhnya (Gansen 1962). Daerah yang pertama adalah daerah penerimaan, yang terdapat pada segmen 1-14. Daerah ini terdiri atas mulut yang peka, esofagus, dan kelenjar faring tak berpembuluh yang mensekresi getah asam yang mengandung enzim amilase.
Daerah yang kedua adalah daerah sekresi, yang terdapat pada segmen 15-44. Daerah ini terdiri atas tembolok, yang menuju ke lambung dan usus. Cacing tanah mensekresi dua enzim protease dan satu enzim amilase terutama dari “sel piala”
b c Juvenil d a Kepala Klitelum Kepala Klitelum Liang jantan Reproduksi cacing tanah Perpindahan sperma Klitelum
Gambar 2 Reproduksi cacing tanah (a), pembentukan kokon (b), perkembangan kokon (c), dan penetasan kokon menjadi juvenil (d).
dinding usus yang mensekresi banyak getah. Beberapa enzim yang berbeda berasal dari daerah ini untuk spesies lain, seperti lipase dan protease (Arthur 1965) serta selulase dan kitinase (Tracey 1951). Makanan yang telah dicerna melintasi aliran darah di sepanjang epithelium usus dan dialirkan ke berbagai bagian tubuh dan jaringan untuk digunakan dalam proses metabolisme dan sebagai cadangan makanan.
Daerah yang ketiga terdapat pada segmen 44 sampai ke anus, disebut daerah absorpsi. Pada daerah ini, bahan-bahan makanan yang tidak tercerna di dalam usus diselubungi oleh membran peritrofik yang melapisi usus. Ketika bahan-bahan makanan diekskresi, membran tersebut akan membungkus casting (sisa pencernaan/kotoran cacing) (Gansen 1962).
Sistem ekskresi
Organ ekskresi yang terpenting pada cacing tanah adalah nefridia. Organ tersebut mengekstraksi bahan-bahan limbah dari cairan selom dan mengeluarkannya ke luar tubuh melalui nefridiofor sebagai urin yang mengandung amonia dan urea. Cairan selom yang mengandung material eksresi melintas melalui nefrostom dan dilanjutkan ke tabung nefridia oleh gerakan silia. (Edwards & Lofty 1972).
Nefridia berperan sebagai penyaring diferensial. Disebut demikian karena terdapat lebih banyak urea dan amonia, tetapi lebih sedikit kreatinin dan protein di dalam urin yang dihasilkan daripada di dalam cairan selom (Ramsay 1949). Nefridia memiliki tiga fungsi pada proses ekskresi, yaitu filtrasi, reabsorbsi, dan transformasi kimiawi (Bahl 1947).
Cacing tanah mengekskresikan zat-zat nitrogen dari dinding tubuh sebagai mukus. Mukus ini berperan sebagai pelumas, mengikat partikel tanah untuk membentuk dinding liang, dan membentuk lapisan pelindung yang melawan bahan-bahan beracun. Sekitar setengah nitrogen total yang diekskresikan per hari terdapat di dalam mukus ini (Edwards & Lofty 1972). Beberapa cacing tanah seperti Pheretima memiliki kelenjar limfa, tempat akumulasi sel-sel ameba. Sel-sel ameba juga terdapat di dalam darah, beberapa Sel-sel ameba tersimpan di dalam
dinding usus, dan selanjutnya masuk ke dalam usus untuk diekskresikan bersama feses (Edwards & Lofty 1972).
Sistem saraf
Sistem saraf cacing tanah terdiri atas sebuah ganglion serebral dorsal, sepasang konektif sirkumentrik dan satu atau lebih tali saraf longitudinal. Ganglion serebral dorsal menyuplai saraf bagian anterior tubuh dan saraf prostomial. Pengontrolan pergerakan tubuh cacing tanah diatur oleh ganglion subenterik. Sedangkan konektif sirkumenterik dan tali saraf longitudinal mengontrol saraf sensoris dan motoris keseluruhan dinding tubuh serta organ di setiap tubuh (Edwards & Lofty 1972).
Organ sensoris cacing tanah terdiri atas dua macam, yaitu organ fotoreseptor dan organ sensoris epitel. Di dalam setiap sel organ fotoreseptor terdapat organel optik yang berbentuk lonjong atau memanjang. Permukaan luar organel ini terdiri atas retina, dan permukaan dalam terdiri atas substansi hialin yang transparan. Organ sensoris epitel merupakan kumpulan dari 35-45 sel-sel yang memanjang dan besar di bagian dasarnya. Ujung distalnya berakhir pada penonjolan rambut-rambut sensori sepanjang daerah kutikula yang tipis. Cacing tanah tidak memiliki mata, tetapi spesies ini memiliki sel-sel sensori yang strukturnya seperti lensa di daerah epidermis dan dermis, terutama pada prostomium. Bagian tengah dan prosterior tubuhnya kurang sensitif terhadap cahaya (Laverack 1963).
Cacing tanah sangat respon terhadap rangsangan cahaya, terutama jika tiba-tiba terpapar cahaya setelah berdiam lama dalam kondisi gelap (Laverack 1963). Cacing tanah Lumbricus bersifat fotopositif terhadap sumber cahaya yang sangat lemah, dan fotonegatif terhadap sumber cahaya yang kuat. Akan tetapi cacing tanah tidak terlalu bereaksi terhadap peningkatan intensitas cahaya yang tiba-tiba jika telah teradaptasi dalam waktu yang lama dalam kondisi cahaya yang kuat. Hal itu disebabkan oleh saturasi reseptor cahaya (Hess 1924). Anggota spesies dari genus Pheretima seluruhnya bersifat fotonegatif terhadap intensitas cahaya (Howell 1939).
Organ sensoris yang bereaksi terhadap rangsangan kimia (kemoreseptor) terdapat pada prostomium (Laverack 1963). Kemoreseptor berperan penting
dalam kehidupan cacing tanah. Kemoreseptor dapat mendeteksi dan mengumpulkan bahan makanan. Kemoreseptor juga dapat digunakan untuk memberi informasi tentang kondisi lingkungan. Disamping itu, kemoreseptor berperan dalam proses perkawinan dengan mendeteksi getah mukus yang dihasilkan oleh cacing tanah yang lain (Smith 1902).
Distribusi Geografi Cacing Tanah
Distribusi cacing tanah sangat luas di seluruh dunia. Akan tetapi pada daerah gurun, kutub, pegunungan, dan daerah dengan sedikit tanah dan vegetasi cacing tanah jarang ditemukan. Beberapa spesies cacing tanah yang terdistribusi secara luas dikenal dengan istilah perigrin (kosmopolitan), sedangkan spesies yang hanya terdapat pada satu daerah tertentu dikenal dengan istilah endemik (Edwards & Lofty 1972).
Spesies perigrin dari famili Lumbricidae menyebar ke beberapa wilayah, terutama di Eropa. Wilayah-wilayah lain yang merupakan daerah penyebaran spesies cacing tanah dari famili ini adalah Chili, Selandia Baru, beberapa daerah di Amerika Serikat, Afrika Barat, India bagian barat laut, dan Australia. Penyebaran Eisenia terdapat di wilayah Eropa, Siberia, Rusia Selatan, Israel, dan Amerika Utara. Sedangkan Lumbricus tersebar ke wilayah Eropa, Siberia, Islandia, dan Amerika Utara. Beberapa spesies perigrin dari genus-genus tersebut terdistribusi di seluruh dunia (Edwards & Lofty 1972; Reynolds 1994).
Cacing tanah Megascolecidae genus Pheretima berasal dari Asia Tenggara. Namun cacing tanah ini juga bermigrasi ke beberapa daerah tropik, subtropik, dan daerah beriklim sedang. Beberapa spesies perigrin dari famili Megascolecidae tersebar di Amerika Selatan dan Amerika Pusat serta Hindia Barat. Genus
Pheretima juga terdistribusi di Indo-Malaya, Asia Timur, dan Australia (Edwards