www.elsevier.com/locate/jinsphys
Spectral correspondence between visual spectral sensitivity and
bioluminescence emission spectra in the click beetle Pyrophorus
punctatissimus (Coleoptera: Elateridae)
Abner B. Lall
a,*, Dora S.F. Ventura
b, Etelvino J.H. Bechara
c, John M. de Souza
b,
Pio Colepicolo-Neto
c, Vadim R. Viviani
caDepartment of Biology, Howard University, 415 College Street, N.W., Washington, DC 20059-0001, USA
bInstituto de Psicologia, Departamento de Psicologia Experimental, Av. Prof. Mello Moraes, 1721 Universidade de Sa˜o Paulo, Sa˜o Paulo, SP
Brazil
cInstituto de Quimica, Departamento de Bioquimica, Av. Prof. Lineu Prestes, 748 Universidade de Sa˜o Paulo, Sa˜o Paulo, SP Brazil
Accepted 1 December 1999
Abstract
The presence of two spectral mechanisms, near-ultraviolet and green (lmax=545 nm), is strongly suggested by electroretinographic
visual spectral sensitivity curves obtained under dark and red chromatic adaptation conditions in the compound eyes of the click beetle Pyrophorus punctatissimus. The bioluminescence emission of the dorsal prothoracic lanterns is deep green (lmax=543 nm)
and that of the ventral abdominal lantern is lime green (lmax=556 nm) in colour in P. punctatissimus. A broad green visual receptor
would detect both deep green and lime green bioluminescent optical signals.2000 Elsevier Science Ltd. All rights reserved.
Keywords: Spectral sensitivity; Click beetle; Bioluminescence; Compound eyes; Visual ecology
1. Introduction
A correspondence exists between the electroretino-graphic (ERG) visual spectral sensitivity [S(l)] and the
bioluminescent optical signal emission spectrum among 10 species of North American fireflies (Lall, 1981; Lall et al. 1980a,b, 1982; Cronin et al., 2000; Seliger et al., 1982a,b) adapted to different photic niches. Since the bioluminescent optical signals are spectrally narrow and limited in photon content, a spectral correspondence between the S(l) functions of the visual receptors and the bioluminescence emission spectrum provides a major selective advantage for the optical signalling for sexual communication among fireflies (Seliger et al., 1982a,b). Unlike lampyrid fireflies, which have only one lantern in the tail, the members of the family Elateridae have three: a pair of dorsal lateral prothoracic lanterns and a
* Corresponding author. Tel.:+1-202-806-6797/6933; fax:+ 1-202-806-4565.
E-mail address: [email protected] (A.B. Lall).
0022-1910/00/$ - see front matter2000 Elsevier Science Ltd. All rights reserved. PII: S 0 0 2 2 - 1 9 1 0 ( 9 9 ) 0 0 2 2 4 - 3
single ventral abdominal lantern. The bioluminescent click beetles or fire-beetles are also commonly called “automobile bugs” because they have a pair of green light (range: 525 to 559 nm) emitting prothoracic lan-terns resembling the headlamps of a car. Among differ-ent species of click beetles, the colour of biolumin-escence in the ventral abdominal lantern varies from lime green to bright yellow or orange (range: 560 to 584 nm) (Bechara, 1988; Colepicolo-Neto et al., 1986). While the spectral characteristics of bioluminescence emission in Elateridae are known (Bechara, 1988; Col-epicolo-Neto et al., 1986), the spectral sensitivity of the elaterid visual system for detection of the bioluminescent optical signal is not known. This paper is the first attempt to describe the visual spectral sensitivity of the com-pound eyes of one species of click beetle, Pyrophorus
punctatissimus, and to establish a correlation between
the species’ bioluminescence spectral emissions and the ERG S(l) function. A preliminary report of this work
2. Materials and methods
2.1. Animals
Pyrophorus punctatissimus Blancher 1843 adult males
were collected during the early hours of the night in the Fazenda Santa Cruz in Costa Rica Prafactue located in the state of Mato Grosso do Sul, Central Western Brazil, during the month of October in 1995 and in 1998. These specimens were maintained in the laboratory in plastic containers at room temperature (21–23°C). Periodically the insects were fed sugar water. The species were ident-ified by Professora C. Costa of the Universidade de Sa˜o Paulo, Museu de Zoologia.
2.2. Optical system
A double-beam optical stimulator was constructed. The test beam originated from a 150 W xenon arc (Quick Illumination Industries, Albany, NY, USA) and passed through a high-intensity grating monochromator (600 lines/mm; Bausch and Lomb Inc., Rochester, NY, USA) which isolated narrow wavelength bands (half bandwidth of 9.9 nm) from 320 to 680 nm. A series of coloured accessory filters (CS7-54, .40% transmission at 250–390 nm; CS4-97, .40% transmission from 355 to 605 nm; and CS3-72, 40% transmission at .450 nm; Corning Glass Co., Corning, NY, USA) was placed in the light beam to eliminate stray light and the second-order spectrum. The light beam was focused by a series of quartz lenses on to one arm of a Y-shaped quartz light guide (3×750 mm). A tungsten light source provided the adaptation beam which was focused on the second arm of the Y-shaped light guide. The colour of the adaptation beam was manipulated with appropriate coloured filters. The light coming out of the common end of the light guide illuminated the entire corneal surface of the com-pound eye. However, the emerging light beam from the light guide was on-axis for facets on the dorsal medial equatorial region. A series of quartz neutral-density fil-ters attenuated the intensity of the test beam over five log units. The stimulus beams were interrupted by an electronic shutter (Uniblitz; Vincent Associates, Roches-ter, NY, USA) which controlled the duration of the stimulus flash.
2.3. Intensity calibration
The quantum flux of the photic stimulus at different wavelengths across the spectrum was determined by placing a calibrated photodiode (SED 033; International Light, Inc., Newburyport, MA, USA) at the spot where the experimental eye was placed. The output of the pho-todiode was read on an electrometer (model IL 700; International Light, Inc., Newburyport, MA, USA).
2.4. Electrical recordings
The electroretinograms (ERGs) were recorded in dark-adapted intact beetles with a glass microelectrode (tip diameter|10µm) filled with insect Ringer’s (Muri and Jones, 1983) and inserted underneath the corneal surface of the compound eye. The reference electrode was another Ringer’s-filled microelectrode inserted in the antenna. An Ag–AgCl pellet connected the microelectrodes to the input grid of a P16 Grass DC/AC preamplifier (Astro Instruments, Providence, RI, USA), the output of which was monitored on an oscilloscope (model 5113; Tektronix, Inc., Beaverton, OR, USA) and displayed on an inkwriter (RS 3200; Gould, Inc., Cleve-land, OH, USA) for later evaluation and measurements. We presume that the responses we obtained arise prim-arily from medial ommatidia, since that was the direction of the stimulating illumination.
2.5. Experimental procedures
Intact insects were immobilized with a mixture of equal amounts of beeswax and rosin, on a recording stage, and dark-adapted for at least an hour prior to experimentation. Test flashes of 0.1 s were delivered at intervals varying from 0.5 to 2 min, permitting complete recovery of the sensitivity of the eye following the pre-vious flash. A standard flash of a selected wavelength and intensity was periodically given to check the overall changes in the sensitivity of the eye. Test flashes were delivered at 19 different wavelengths which covered the spectrum from 320 to 680 nm in 20 nm steps. The ampli-tude of the ERG provided an index of the sensitivity of the eye to quantum flux and to the wavelength compo-sition of the photic stimulus. In some experiments ERGs were recorded over the entire range of available inten-sities for selected stimulus wavelengths, and intensity– response curves (i.e., V/log I functions) were obtained. The spectral sensitivity of the eye was obtained by determining the number of photons needed to elicit a criterion amplitude (2 mV) of the ERG at different stimulus wavelengths. A plot of 1/Q as a function of wavelength gave the S(l) function. All recordings were obtained during the photophase. All experiments were conducted at room temperature (21–23°C). The data were obtained during the month of November 1995 in the Laboratory of Sensory Psychophysiology at the Uni-versity of Sa˜o Paulo and during in the months of Nov-ember and DecNov-ember of 1998 at Howard University.
2.6. Bioluminescence emission spectra
beetles were anaesthetized with chloroform vapour for 2 min, immobilized with tape on glass slides, and placed in front of the emission window. In addition, in some animals, 10µl of commercial epinephrine (1 mg/ml) was injected under the cuticle of the dorsal lantern to stimu-late a constant glow. The ventral lantern was directly exposed without epinephrine treatment. The emission slit was 5 mm (half bandwidth=9.42 nm) and the scan rate equal to 600 nm/min. The spectra were automatically corrected for the spectral photosensitivity of the equip-ment with an internal program identified as M-correct on the instrument by the manufacturer. The final emis-sion curve is the average of five independent traces on five separate individuals. These data were obtained in the Department of Biochemistry at the University of Sa˜o Paulo.
3. Results
3.1. Electroretinogram and visual spectral sensitivity of the compound eyes
The electroretinograms (ERGs) recorded from the compound eyes in P. punctatissimus were on-negative responses similar to those described for Photuris
versic-olor (Lall, 1981) and Photuris lucicrescens (Lall et al.,
1982). There was no off-transient.
The V/log I functions (Fig. 1) were obtained to deter-mine the spectral sensitivity curves (Fig. 2) and to exam-ine whether there were any wavelength-dependent changes in the slopes of curves obtained at different test wavelengths. No consistent change in the slopes of
V/log I curves as a function of wavelength was observed.
This fact alone cannot be taken as evidence for the pres-ence of only a single receptor type in P. punctatissimus compound eyes, since the slopes of V/log I functions
Fig. 1. V/log I curves for selected wavelengths from one
dark-adapted compound eye in P. punctatissimus. Abscissa: log10intensity
of the stimulus given as neutral density filters attenuation of the test stimulus; ordinate: peak mV responses to 100 ms test flash.
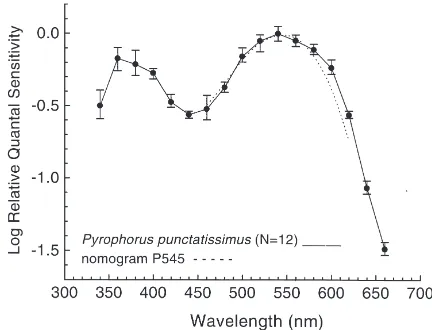
Fig. 2. Electroretinographic spectral sensitivity function in dark-adapted compound eyes in the male of P. punctatissimus (an average of 12 runs on 10 eyes from five individuals). Error bars indicate±one standard deviation (SD). The dotted line is the nomogram for P545 rhodopsin (Ebrey and Honig, 1977).
tend to be constant across the spectrum even when dif-ferent receptor types are present (Lall, 1981; Lall et al. 1980a, 1982).
The visual spectral sensitivity functions were obtained for a threshold criterion ERG response of 2 mV peak-to-peak amplitude in P. punctatissimus and the average
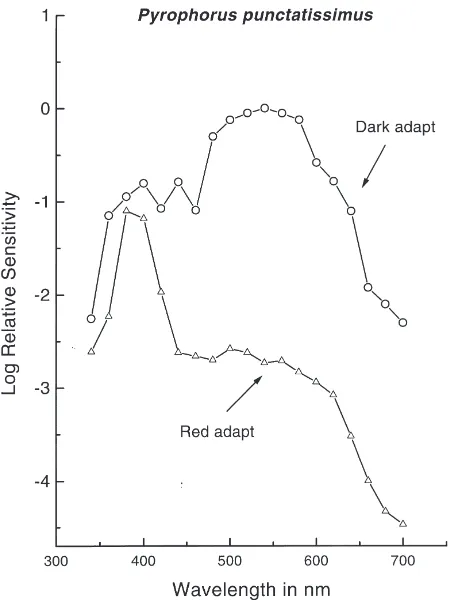
S(l) curve is presented in Fig. 2. The dark-adapted S(l) function has a broad sensitivity peak in the green with a shoulder in the near-ultraviolet (near-UV) (Fig. 2). Under red chromatic adaptation conditions visual sensi-tivity was reduced precipitously in the long-wavelength region leaving high sensitivity in the near-UV region of the spectrum (Fig. 3). The responses from the dorsal and the ventral sectors were not isolated. The S(l) curves
represent the average response of the dorsal medial equa-torial region of the compound eye. In vivo biolumin-escence emission spectra of the dorsal prothoracic (lmax=543 nm) and the ventral abdominal lanterns
(lmax=556 nm) in P. punctatissimus are presented in
Fig. 4.
4. Discussion
The S(l) functions in Figs. 2 and 3 strongly suggest the presence of two spectral mechanisms, green and near-UV, in the compound eyes in P. punctatissimus. Red chromatic adaptation had a differential effect on the dark-adapted curve such that a near-UV spectral mech-anism was isolated (Fig. 3). Similar S(l) functions have
Fig. 3. A comparison of electroretinograhic spectral sensitivity func-tion between dark and red chromatic adaptafunc-tion experiments for P.
punctatissimus (an average of three runs on three eyes).
Fig. 4. A comparison between the average spectral sensitivity func-tion from Fig. 2 plotted on a percentage scale (the curve is fitted to B-spline by the Microcal Origin 4.1 computer software) and the spec-ies’ in vivo bioluminescence emission spectra of the dorsal prothoracic and ventral abdominal lanterns of P. punctatissimus.
Honig, 1977) and the ERG S(l) function in Fig. 2
sug-gests that the broad green spectral mechanism is mediated by P545 rhodopsin.
The bioluminescence emission peaks from the dorsal (543 nm) and ventral (556 nm) lanterns in P.
punctatis-simus in Fig. 4 are similar to those obtained in 11 other
Brazilian species of elaterids (Colepicolo-Neto et al., 1986). In all cases the peak of the bioluminescence emis-sions from the abdominal light organ were shifted (13– 40 nm) towards long wavelengths relative to the peak for the prothorax lanterns which varied from 525 to 559 nm. Thus far the functional significance of optical signal-ling of the dorsal prothoracic and the ventral abdominal lantern is unclear. When the animal is disturbed, the dor-sal lanterns light up. When in flight, the insect usually emits a very tiny glow from the dorsal lanterns. One function attributed to dorsal lanterns is that of a warning signal for potential predators. In such a situation, the selection of a green optical signal would be favoured if it were detectable by the predators of the click beetles. According to the Sensitivity Hypothesis of visual ecol-ogy (Seliger et al., 1982a,b), the selection of P545 rho-dopsin would maximize absolute sensitivity in a green foliage habitat at night. Presumably the predators of the click beetles possess visual systems optimized for func-tioning under such conditions.
The light from the ventral lantern is emitted only when the male is flying in broad sweeps in search of a responsive female. The light emission of the ventral lan-tern is implicated in sexual communication. The details of the communication between the male and female have yet to be elucidated by a field biologist. The visual sys-tem of P. punctatissimus can detect green and lime green bioluminescence emissions from the dorsal and the ven-tral lanterns, respectively; the lime green biolumin-escence will be detected with almost 92% efficiency as compared with the green one. The behavioural signifi-cance of such a system at present eludes us. There exists now a growing body of evidence to suggest that bio-luminescence is an achromatic behaviour mediated exclusively by long-wavelength spectral mechanisms (Case, 1984; Lall and Worthy, 2000). The species speci-ficity with respect to the colour of bioluminescence emission is governed by the characteristics of the lucifer-ase enzyme (McElroy et al., 1974) that is selected for the detectability of the bioluminescent optical signal against interfering environmental light of varying intensity and wavelength composition in the species habitat (Seliger et al., 1982a,b).
Acknowledgements
References
Bechara, E.J.H., 1988. Luminescent elaterid beetles: biochemical, bio-logical and ecobio-logical aspects. In: Baumstark, A.L. (Ed.). Advances in Oxygenated Processes, vol. 1. JAI Press, London, pp. 123–178. Case, J.F., 1984. Vision and mating behavior in fireflies. In: Lewis, T. (Ed.) Insect Communication. Royal Entomological Society of London, London, pp. 195–222.
Colepicolo-Neto, P., Costa, D., Bechara, E.J.H., 1986. Brazilian spec-ies of luminescent elateridae: luciferase identification and biolumin-escence spectra. Insect Biochemistry 16, 803–810.
Cronin, T.W., Ja¨rvilehto, M., Weckstro¨m, M., Lall, A.B., 2000. Tuning of photoreceptor spectral sensitivity in fireflies (Coleoptera: Lampyridae). Journal of Comparative Physiology A 186, 1–12. Ebrey, T.W., Honig, B., 1977. New wavelength dependent visual
pig-ment nomograms. Vision Research 17, 147–151.
Lall, A.B., 1981. Electroretinogram and the spectral sensitivity of the compound eyes in the firefly Photuris versicolor (Coleoptera: Lampyridae): a correspondence between green sensitivity and spec-ies bioluminescence emission. Journal of Insect Physiology 27, 461–468.
Lall, A.B., Worthy, K.M., 2000. Action spectra of the female’s response in firefly Photinus pyralis (Coleoptera: Lampyridae): evi-dence for an achromatic detection of the optical signal. Journal of Insect Physiology 24, in press.
Lall, A.B., Chapman, R.M., Trouth, C.O., Holloway, J.A., 1980a. Spectral mechanisms of the compound eye in the firefly Photinus
pyralis (Coleoptera: Lampyridae). Journal of Comparative
Physi-ology 135, 21–27.
Lall, A.B., Seliger, H.H., Biggley, W.H., Lloyd, J.E., 1980b. Ecology of colors of firefly bioluminescence. Science 210, 560–562. Lall, A.B., Lord, E.T., Trouth, C.O., 1982. Vision in the firefly
Phot-uris lucicrescens (Coleoptera: Lampyridae): spectral sensitivity and
selective adaptation in the compound eyes. Journal of Comparative Physiology 147, 195–200.
Lall, A.B., Strother, G.K., Cronin, T.W., Seliger, H.H., 1988. Modifi-cation of spectral sensitivities by the screening pigments in the compound eyes of twilight-active fireflies (Coleoptera: Lampyridae). Journal of Comparative Physiology A 162, 23–33. Lall, A.B., Cronin, T.W., Bechara, E.J.H., Ventura, D.S.F., De Souza,
J., Colepicolo-Neto, P., Viviani, V.R., 1997. Tuning between visual spectral sensitivity and bioluminescence emission spectra in click beetle Pyrophorus punctatissimus male. Society for Neuroscience Abstracts 23, 1818# 707.11.
McElroy, W.D., Seliger, H.H., DeLuca, M., 1974. Insect biolumin-escence. In: Rockstein, M. (Ed.) The Insects. Academic Press, New York.
Menzel, R., Backhaus, W., 1991. Colour vision in insects. In: Gouras, P. (Ed.) The Perception of Colour. Vision and Visual Dysfunction, vol. 6. CRC Press/Ann Arbor, Boca Raton, FL/Boston, MA, pp. 262–293.
Muri, R.B., Jones, G.J., 1983. Microspectrophotometry of single rhab-doms in the retina of the honeybee drone (Apis mellifera). Journal of General Physiology 82, 469–496.
Seliger, H.H., Lall, A.B., Lloyd, J.E., Biggley, W.H., 1982a. On the colors of firefly bioluminescence. I. An optimization model. Photo-chemistry and Photobiology 36, 673–680.