DERAJAT KETAHANAN GENOTIPE KEDELAI
TERHADAP HAMA ULAT GRAYAK
M. Muchlish Adie, Ayda Krisnawati1), dan Ayu Zuhrotul Mufidah2) 1)Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian Raya Kendalpayak Km 8 Kotak Pos 66 Malang 65101, Telp.(0341) 801468
2)Alumni Jurusan Biologi MIPA FKIP UMM E-mail: mm_adie@yahoo.com
ABSTRAK
Ulat grayak (Spodoptera litura) merupakan hama pemakan daun paling penting di Indo-nesia, dan tingkat kerugian akibat serangannya bergantung pada varietas, fase pertumbuhan dan waktu serangan. Uji ketahanan dari 10 genotipe kedelai (Shr/W-C-60, Aochi/Wil-60, 9837/K-D-8-185, 9837/K-D-3-185-95, W/9837-D-6-220, 9837/K-D-3-185-82, 9837/W-D-5-211, GI, G100H dan Wilis) terhadap hama ulat grayak dilakukan di kurungan kasa dan labo-ratorium pemuliaan Balitkabi pada tahun 2005. Penelitian di kurungan kasa menggunakan rancangan acak kelompok, 10 genotipe sebagai perlakuan dan setiap perlakuan diulang tiga kali. Setiap genotipe ditanam pada pot plastik (Ø 18 cm) berisi dua tanaman. Pada saat tanaman berumur 27 hari, 10 genotipe kedelai disungkup dengan kurungan kasa (2m x 2m x 2m) dan pada masing-masing pot diinfestasi larva ulat grayak instar I sebanyak 10 ekor. Penelitian di laboratorium menggunakan rancangan acak kelompok diulang lima kali. Daun kedelai trifoliat yang terdapat pada buku ketiga dari tanaman kedelai berumur 27 HST, dipindahkan pada cawan petri berdiamater 15 cm, yang sebelumnya telah dilapisi oleh kertas saring yang telah dibasahi dengan air suling. Pada setiap cawan diinfestasi lima larva ulat grayak yang baru menetas (neonate). Kajian biologi ulat grayak dilakukan dengan meng-gunakan rancangan acak kelompok diulang tujuh kali. Pada setiap cawan diinfestasi satu larva ulat grayak yang baru menetas (neonate). Intensitas kerusakan daun oleh hama ulat grayak berbeda antargenotipe kedelai. Rata-rata intensitas kerusakan daun pada 5 HSI adalah 12,9%, meningkat menjadi 63,5% setelah 20 HSI. Genotipe G100H, kerusakan daunnya konsisten terendah, yang mengindikasikan bahwa G100H lebih tahan terhadap ulat grayak dibandingkan dengan genotipe lainnya. Perbedaan bobot individu larva mulai terlihat setelah 6 HSI. Bobot larva pada umur 3 HSI adalah 0,026 mg/ekor, meningkat menjadi 0,462 mg/ekor pada umur 6 HSI dan 1,860 mg/ekor pada umur 9 HSI. Bobot larva terendah diperoleh dari pakan daun genotipe G100H, yang menunjukkan bahwa G100H kurang disukai sebagai pakan oleh larva ulat grayak, sebagaimana ditunjukkan oleh konsumsi daun dari G100H yang hanya 21,5 cm2/hari, lebih rendah dibandingkan dengan genotipe lainnya. Pada genotipe tahan ulat grayak (G100H), umur larvanya lima hari lebih lama dibandingkan dengan umur larva asal pakan genotipe peka (GI) yang mencapai rata-rata 13,1 hari. Faktor penentu ketahanan G100H terhadap ulat grayak adalah trikoma rapat dan panjang. G100H disarankan digunakan sebagai sumber gen untuk perbaikan ketahanan kedelai terhadap hama ulat grayak.
Kata kunci: genotipe tahan ulat grayak, trikoma, biologi ulat grayak
ABSTRACT
The resistance level of soybean genotypes to armyworm common cutworm. Armyworm (Spodoptera litura F.) is the most important leaf-feeding pest in Indonesia. The yield loss due to armyworm depends on the variety and crop growth stage when the pest attacks. The resistance evaluation of ten soybean genotypes was done in 2005. The experiment in the
screen cage was arranged in complete randomized design, three replicates. Each genotype was planted in a plastic pot with two plants per pot. At 27 days after planting (DAP), all pots were placed in a 2 x 2 x 2 m screen cage, where each pot was invested by ten larvae (first instar). The experiment in the laboratory was arranged in randomized block design with five replications. A 27 days of trifoliate leaf from each line was transferred into a 15 cm-diameter petri dish, which had previously been covered by moistened filter paper. Each plate was then invested with five neonate larvae. The biological study of armyworm was carried out using randomized block design, seven replications. Each plate was invested with one neonate larvae. The intensity of leaf damage was different among genotypes, with the average was 12.86% and 63.52% at 5 and 20 DAP, respectively. Genotype G100H was more resistant to armyworm than others, based on its consistently lowest leaf damage. The different weight of individual larvae was started after 6 DAP. The weight of larvae at 3 DAP was 0.026 mg, increased to 0.462 mg and 1,860 mg at 6 and 9 DAP, respectively. The lowest weight of larvae was obtained from those fed by G100H leaves. This showed that G100H was less favored by armyworm larvae, as indicated by 21.51 cm2/day of consumption only. The determinant factor this resistance was the dense and long trichomes. Genotype G100H was suggested to be used as gene source in the improvement of soybean resistance to armyworms.
Keywords: resistant genotype to armyworm, trichome, biology of armyworm
PENDAHULUAN
Di Indonesia, ulat grayak, Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) merupakan hama penting pemakan daun kedelai dibandingkan dengan hama daun lainnya, seperti ulat jengkal (Chrysodeixis chalcites), ulat helicoverpa (Heliothis armigera), dan ulat penggulung daun (Lamprosema indicata). Kerugian hasil kedelai akibat serangan ulat grayak dilaporkan lebih dari 80% di Jepang (Komatsu et al. 2010) dan 90% di Amerika (Wier dan Boethel 1996). Di Indonesia, intensitas serangan ulat grayak berfluktuasi, pada tahun 1997 luas serangannya mencapai 842 ha dengan intensitas serangan 23% (BPS 1999). Tingkat kerugian hasil biji bergantung pada varietas yang digunakan, fase pertumbuhan, dan waktu serangan.
Sejarah pengelolaan ketahanan kedelai terhadap hama relatif belum terlalu lama. Sebelum tahun 1960, proses perbaikan ketahanannya difokuskan pada penggunaan trikoma pada tanaman untuk mengatasi serangan hama Empoasca, hasilnya diperoleh varietas kedelai yang ketahanannya bersifat stabil tetapi hanya untuk satu jenis hama. Pada akhir 1960an, perakitan varietas kedelai dilakukan dengan mengidentifikasi aksesi plasma nutfah dan diperoleh tiga aksesi (PI 229358, PI 227687 dan PI 171451) yang memiliki ketahanan tinggi terhadap kompleks hama pemakan daun. Sejak periode tersebut, target hama sasaran menjadi lebih luas, khususnya untuk kompleks hama pemakan daun. Teridentifikasinya tiga aksesi kedelai tersebut, maka program peningkatan ketahanan kedelai terhadap hama terus berkembang. Beberapa peneliti mengungkapkan bahwa ketahanan terhadap hama pemakan daun Epilachna varivestis dan Anticarsia
gemmatalis disebabkan oleh mekanisme antibiosis (van Duyn et al. 1971; Clarck et al.
1972; Lambert dan Kilen 1984; Kilen et al. 1977). Ketahanan antisenosis, khususnya keta-hanan morfologi, merupakan potensi ketaketa-hanan yang perlu ditelaah dan dimanfaatkan dalam pembentukan varietas kedelai.
Selama ini, program perakitan varietas kedelai di Indonesia difokuskan pada peningkatan potensi hasil dan perbaikan ketahanan terhadap berbagai cekaman abiotik
seperti lahan masam, penaungan dan kekeringan. Tantangan perakitan varietas kedelai ke depan adalah bagaimana mengalokasikan dan meningkatkan ketahanan kedelai terhadap hama utama di Indonesia, termasuk ulat grayak. Strategi perbaikannya dapat ditempuh melalui pemahaman terhadap mekanisme ketahanan hama. Trikoma merupakan bentuk mekanisme ketahanan antisenosis dan menjadi karakter pertahanan potensial bagi tanaman untuk hama tertentu, termasuk hama perusak daun ulat grayak. Bentuk ketahanan demikian penting untuk kondisi Indonesia, karena sejak awal telah diupayakan untuk memutus atau mengurangi terjadinya interaksi antara tanaman inang dengan serangga hama. Apalagi dilaporkan bahwa penggunaan varietas tahan hama dinilai mampu mempertahanan keseimbangan ekosistem, sejalan dengan konsep pengelolaan hama terpadu (PHT). Bahkan menurut Teetes (1996), penggunaan varietas tahan hama lebih baik dibandingkan dengan komponen pengendalian langsung lainnya.
Tujuan penelitian ini adalah mengidentifikasi ketahanan beberapa genotipe kedelai terhadap hama ulat grayak.
BAHAN DAN METODE
Bahan penelitian adalah 10 genotipe kedelai yaitu Shr/W-C-60, Aochi/Wil-60, 9837/K-D-8-185, 9837/K-D-3-185-95, W/9837-D-6-220, 9837/K-D-3-185-82, 9837/W-D-5-211, GI, G100H dan Wilis. Penelitian dilaksanakan pada tahun 2005 di Balitkabi, Malang. Uji ketahanan genotipe kedelai terhadap ulat grayak dilakukan di kurungan kasa dan laboratorium pemuliaan.
Penelitian di Kurungan Kasa
Penelitian menggunakan rancangan acak kelompok, 10 genotipe sebagai perlakuan dan setiap perlakuan diulang tiga kali. Setiap genotipe ditanam pada pot plastik (Ø 18 cm) berisi dua tanaman. Pada saat tanaman berumur 27 hari, 10 genotipe kedelai disungkup dengan kurungan kasa (2 m x 2 m x 2 m) dan pada masing-masing pot diinfestasi larva ulat grayak instar I sebanyak 10 ekor. Satu sungkup merupakan satu ulangan, sehingga seluruhnya terdapat tiga kurungan kasa. Pada setiap kurungan kasa, daun antargenotipe kedelai diupayakan saling bersentuhan sehingga memungkinkan bagi larva untuk memilih daun genotipe yang disukai secara bebas. Pengamatan dilakukan terhadap tingkat kerusakan daun dari 10 genotipe kedelai pada 5, 10, 15 dan 20 HSI (hari setelah infestasi) dengan rumus sebagai berikut.
∑ (n x v)
I =x 100%
Z x N
Di mana, I = intensitas serangan
n = jumlah daun dalam tiap kategori serangan v = nilai skala dari tiap kategori serangan (1–4) Z = nilai skala dari kategori serangan tertinggi V = jumlah daun yang diamati
Skala serangan
0 = tidak ada serangan
1 = luas daun yang dimakan mencapai 1–25% 2 = luas daun yang dimakan mencapai 26–50% 3 = luas daun yang dimakan mencapai 51–75%
PENELITIAN DI LABORATORIUM
Uji antibiosis menggunakan rancangan acak kelompok diulang lima kali. Bahan yang digunakan adalah daun kedelai trifoliat yang terdapat pada buku ketiga dari tanaman kedelai berumur 27 HST, kemudian dipindahkan ke cawan petri berdiamater 15 cm, yang sebelumnya telah dilapisi dengan kertas saring yang telah dibasahi dengan air suling. Pada setiap cawan diinfestasi lima larva ulat grayak yang baru menetas (neonate). Penggantian daun dilakukan pada umur 3, 6, dan 9 HSI. Kertas saring dipertahankan lembab. Peneli-tian dihentikan hingga hari kesembilan setelah infestasi.
Kajian biologi ulat grayak menggunakan rancangan acak kelompok diulang tujuh kali. Daun trifoliat yang berada pada buku ketiga dari tanaman kedelai umur 27 HST dipindah-kan pada cawan petri (petridish) berdiamater 15 cm, yang sebelumnya telah dilapisi dengan kertas saring yang telah dibasahi air suling. Pada setiap cawan diinfestasi satu larva ulat grayak yang baru menetas (neonate). Penggantian daun dilakukan sesuai kebu-tuhan dan secara bersamaan untuk setiap perlakuan. Kertas saring dipertahankan lembab secara bebas. Pengamatan dilakukan pada umur larva dan pupa, bobot, diameter dan panjang pupa, bobot larva pada umur 3, 6, dan 9 hari. Di samping itu juga diamati mor-fologi daun yaitu luas daun, tebal daun, kerapatan trikoma daun (4 mm2), panjang trikoma
pada daun, dan luas daun. Pengamatan dilakukan pada saat tanaman berumur 27 HST.
HASIL DAN PEMBAHASAN
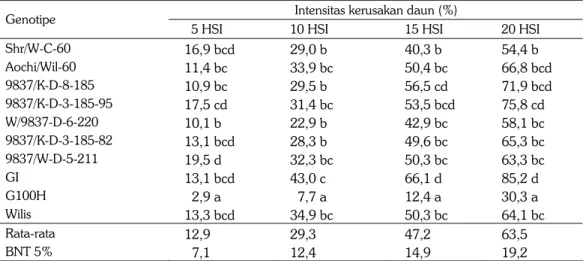
Intensitas kerusakan daun kedelai berbeda antargenotipe (Tabel 1). Intensitas kerusakan daun sejak umur 5 HSI (hari setelah infestasi) memiliki hubungan linear dengan umur larva. Rata-rata intensitas kerusakan daun 12,9% setelah 5 HSI, meningkat menjadi 63,5% setelah 20 HSI. Setelah 5 HSI, kerusakan daun genotipe 9837/W-D-5-211 tertinggi (19,5%) dan terendah pada G100H (2,89%). Dari 10 genotipe yang diuji terdapat dua genotipe yang kerusakan daunnya konsisten rendah (G100H) dan yang tertinggi adalah pada genotipe GI sejak 10 HSI. Hal ini mengindikasikan bahwa G100H lebih tahan dan GI lebih peka terhadap ulat grayak dibandingkan dengan genotipe lainnya.
Tabel 1. Intensitas kerusakan daun oleh larva ulat grayak pada 10 genotipe kedelai. Laboratorium, MK, 2005.
Intensitas kerusakan daun (%) Genotipe
5 HSI 10 HSI 15 HSI 20 HSI
Shr/W-C-60 16,9 bcd 29,0 b 40,3 b 54,4 b Aochi/Wil-60 11,4 bc 33,9 bc 50,4 bc 66,8 bcd 9837/K-D-8-185 10,9 bc 29,5 b 56,5 cd 71,9 bcd 9837/K-D-3-185-95 17,5 cd 31,4 bc 53,5 bcd 75,8 cd W/9837-D-6-220 10,1 b 22,9 b 42,9 bc 58,1 bc 9837/K-D-3-185-82 13,1 bcd 28,3 b 49,6 bc 65,3 bc 9837/W-D-5-211 19,5 d 32,3 bc 50,3 bc 63,3 bc GI 13,1 bcd 43,0 c 66,1 d 85,2 d G100H 2,9 a 7,7 a 12,4 a 30,3 a Wilis 13,3 bcd 34,9 bc 50,3 bc 64,1 bc Rata-rata 12,9 29,3 47,2 63,5 BNT 5% 7,1 12,4 14,9 19,2
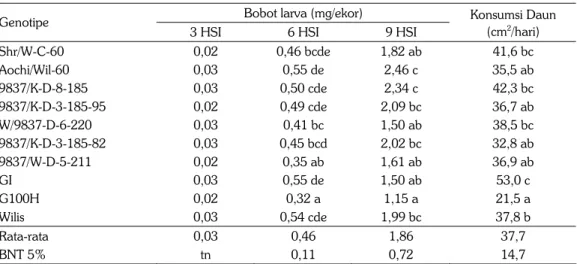
Bobot larva ulat grayak tidak dipengaruhi oleh asal pakan dari 10 genotipe kedelai pada umur 3 HSI, namun pakan mulai berpengaruh secara nyata pada bobot larva setelah umur 6 dan 9 HSI. Hal ini menunjukkan daya makan larva hingga umur 3 hari relatif sedikit. Bobot larva pada umur 3 HSI adalah 0,026 mg/ekor, meningkat menjadi 0,462 mg/ekor pada umur 6 HSI dan 1,860 mg/ ekor pada umur 9 HSI (Tabel 2). Bobot larva terendah diperoleh dari pakan daun genotipe G100H (0,323 mg/ekor) pada umur 6 HSI dan tetap konsisten terendah menjadi 1,147 mg/ekor pada umur 9 HSI. Hal ini menunjukkan bahwa G100H kurang disukai sebagai pakan oleh larva ulat grayak, sebagaimana ditunjukkan oleh konsumsi daun dari G100H yang hanya 21,5 cm2/hari,
lebih rendah dibandingkan dengan genotipe lainnya. Hambatan pakan sebagaimana terjadi pada G100H, juga dilaporkan oleh Al Ayedh (1997), yakni efektif digunakan sebagai pertahanan tanaman terhadap beberapa hama daun karena akan menghambat pergerakan larva dan hambatan terhadap konsumsi daun. Perilaku serangga dalam mencari inang yang tepat sangat penting karena digunakan sebagai penentuan kisaran inang. Artinya, G100H kurang tepat digunakan sebagai inang hama ulat grayak dibandingkan dengan sembilan genotipe kedelai lainnya.
Tabel 2. Bobot larva ulat grayak asal pakan pada 10 genotipe kedelai. Laboratorium, MK, 2005. Bobot larva (mg/ekor)
Genotipe
3 HSI 6 HSI 9 HSI
Konsumsi Daun (cm2/hari) Shr/W-C-60 0,02 0,46 bcde 1,82 ab 41,6 bc Aochi/Wil-60 0,03 0,55 de 2,46 c 35,5 ab 9837/K-D-8-185 0,03 0,50 cde 2,34 c 42,3 bc 9837/K-D-3-185-95 0,02 0,49 cde 2,09 bc 36,7 ab W/9837-D-6-220 0,03 0,41 bc 1,50 ab 38,5 bc 9837/K-D-3-185-82 0,03 0,45 bcd 2,02 bc 32,8 ab 9837/W-D-5-211 0,02 0,35 ab 1,61 ab 36,9 ab GI 0,03 0,55 de 1,50 ab 53,0 c G100H 0,02 0,32 a 1,15 a 21,5 a Wilis 0,03 0,54 cde 1,99 bc 37,8 b Rata-rata 0,03 0,46 1,86 37,7 BNT 5% tn 0,11 0,72 14,7
HSI = hari setelah infestasi. Angka sekolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji BNT 5%.
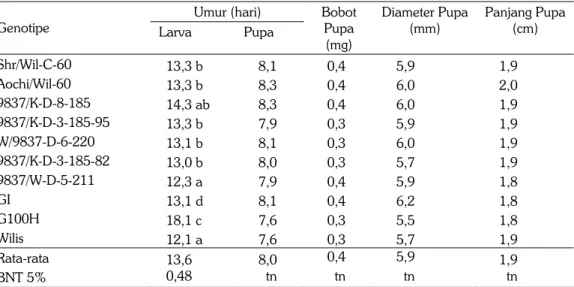
Pengaruh pakan asal 10 genotipe terhadap biologi ulat grayak hanya terjadi pada umur larva, sedangkan umur, bobot, diameter dan panjang pupa tidak dipengaruhi oleh asal pakan. Beberapa penelitian sebelumnya juga mengindikasikan bahwa ketidaksesuaian pakan akan mengganggu proses metabolisme larva sehingga memperpanjang umur larva. Larva yang mendapatkan pakan G100H memiliki umur rata-rata 18,1 hari, lebih lama dibandingkan dengan umur larva asal pakan genotipe GI yang hanya 13,1 hari (Tabel 3). Komatsu el al. (2010) membandingkan tingkat antibiosis dari varietas Himeshirazu dengan Fukuyutaka (peka) dan Sodendaizu (tahan), dimana larva yang mendapatkan pakan asal genotipe Himeshirazu memiliki bobot individu larva lebih ringan dan umur instar lebih panjang dibanding genotipe peka. Kajian pewarisan antibiosisnya pada populasi F2 dari persilangan Himeshirazu dengan Fukuyutaka menghasilkan heritabilitas arti luas untuk
antibiosis sebesar 71,3%; dengan pengendali faktor resesif. Kajian dengan SSR menun-jukkan bahwa faktor resesif dari antibiois berada pada kromosom kelompok pautan M. Tabel 3. Biologi ulat grayak asal pakan pada 10 genotipe kedelai. MK. 2005.
Umur (hari)
Genotipe Larva Pupa Bobot Pupa (mg) Diameter Pupa (mm) Panjang Pupa (cm) Shr/Wil-C-60 13,3 b 8,1 0,4 5,9 1,9 Aochi/Wil-60 13,3 b 8,3 0,4 6,0 2,0 9837/K-D-8-185 14,3 ab 8,3 0,4 6,0 1,9 9837/K-D-3-185-95 13,3 b 7,9 0,3 5,9 1,9 W/9837-D-6-220 13,1 b 8,1 0,3 6,0 1,9 9837/K-D-3-185-82 13,0 b 8,0 0,3 5,7 1,9 9837/W-D-5-211 12,3 a 7,9 0,4 5,9 1,8 GI 13,1 d 8,1 0,4 6,2 1,8 G100H 18,1 c 7,6 0,3 5,5 1,8 Wilis 12,1 a 7,6 0,3 5,7 1,9 Rata-rata 13,6 8,0 0,4 5,9 1,9 BNT 5% 0,48 tn tn tn tn
tn = tidak nyata. Angka sekolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji BNT 5%.
Kajian ketahanan dengan melihat preferensi peletakan telur menunjukkan bahwa Kyukei 279 tergolong moderat terhadap hama ulat grayak yang diukur dari tingkat kerusakan daun dan preferensi oviposisi yang lebih rendah dibandingkan dengan genotipe peka Fukuyutaka (Nobuyuki et al. 2002). Endo et al. (2007) membandingkan ketahanan dari tiga genotipe kedelai (Bay, Fukuyutaka, dan Himeshirazu), yang menunjukkan bahwa pada genotipe tahan Bay, konsumsi pakan dari larva lebih sedikit (78,2%), bobot larva lebih rendah (67,3%), umur larva lebih lama (114,2%), dan bobot pupa lebih ringan (79,7%) dibandingkan dengan tanaman yang tidak mengalami kerusakan (100%).
Untuk memahami penyebab ketahanan kedelai terhadap ulat grayak, diidentifikasi karakter morfologi daun. Sepuluh genotipe kedelai memiliki karakter morfologi daun (ketebalan, kerapatan trikoma, panjang trikoma dan luas daun) yang berbeda nyata. G100H memiliki daun yang lebih tebal (0,97 mm) dan yang paling tipis pada varietas Wilis (0,57 mm). Selain berdaun tebal, G100H juga memiliki trikoma yang sangat padat, baik pada permukaan daun bagian atas maupun pada bagian bawah, diikuti oleh trikoma yang lebih panjang. Varietas Wilis, yang relatif kurang tahan, memiliki kerapatan trikoma 16 buah/4 mm2 pada permukaan daun atas dan 23 buah/4 mm2 pada daun bagian
Tabel 4. Karakter morfologi daun dari 10 genotipe kedelai. Laboratorium, MK. 2005. Kerapatan trikoma/ 4 mm2 Panjang trikoma (mm) Genotipe Ketebalan daun (mm) Daun atas Daun bawah Daun atas Daun bawah Luas daun (cm2) Shr/W-C-60 0,8 cdef 15,3 a 11,0 ab 0,5 b 0,5 b 32,0 cd Aochi/Wil-60 0,7 abcd 15,7 a 10,0 a 1,0 c 0,9 c 59,4 e 9837/K-D-8-185 0,7 abc 15,3 a 16,0 c 0,6 b 0,6 b 25,4 bc 9837/K-D-3-185-95 0,7 abc 14,7 a 15,7 bc 0,6 b 0,6 b 51,7 e W/9837-D-6-220 0,6 ab 18,7 a 16,3 c 0,6 b 0,8 c 36,9 d 9837/K-D-3-185-82 0,9 def 14,7 a 15,7 bc 0,6 b 0,5 b 35,9 d 9837/W-D-5-211 0,9 ef 15,7 a 15,7 bc 1,4 d 1,1 d 36,0 d GI 0,8 bcde 15,0 a 10,0 a 0,2 a 0,1 a 20,7 a G100H 1,0 f 25,0 b 29,0 e 1,1 c 1,1 d 55,2 e Wilis 0,6 a 16,0 a 22,7 d 0,5 b 0,5 b 30,0 cd Rata-rata 0,8 16,6 16,2 0,7 0,7 38,3 BNT 5% 0,2 5,8 4,3 0,2 0,2 8,5
BNT = beda nyata terkecil. Angka sekolom yang diikuti oleh huruf yang sama tidak berbeda nyata menurut uji BNT 5%
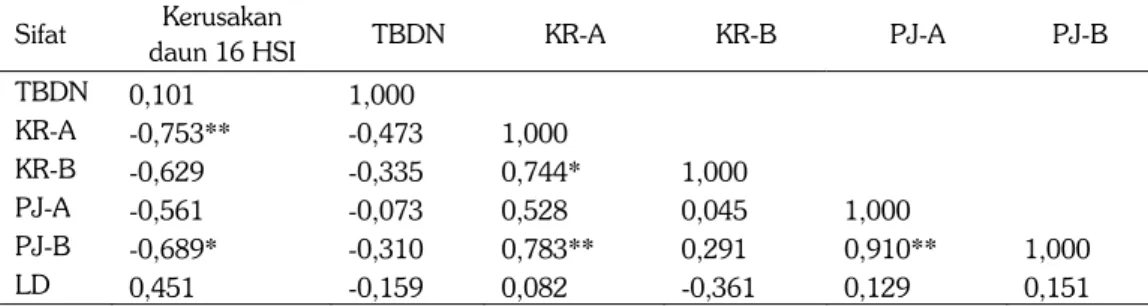
Tabel 5. Korelasi antara intensitas kerusakan daun pada 16 HSI dengan karakter morfologi daun. Sifat Kerusakan
daun 16 HSI TBDN KR-A KR-B PJ-A PJ-B
TBDN 0,101 1,000 KR-A -0,753** -0,473 1,000 KR-B -0,629 -0,335 0,744* 1,000 PJ-A -0,561 -0,073 0,528 0,045 1,000 PJ-B -0,689* -0,310 0,783** 0,291 0,910** 1,000 LD 0,451 -0,159 0,082 -0,361 0,129 0,151
* dan ** = nyata pada p = 0.05 dan p = 0.01. TBDN = tebal daun. KR-A = kerapatan trikoma permukaan daun atas. KR-B = kerapatan trikoma permukaan daun bawah. PJ-A = panjang trikoma daun bagian atas. PJ-B = panjang trikoma daun bagian bawah. LD = luas daun
Penelusuran faktor penentu kerusakan daun oleh hama ulat grayak pada 16 HSI, ternyata ditentukan oleh kerapatan trikoma yang ada pada daun bagian atas (r = -0.753**) dan diikuti oleh panjang trikoma daun pada bagian bawah (r = -0,689*). Orientasi trikoma, khususnya kerapatan dan panjang trikoma, berperan penting dalam mempengaruhi daya makan larva, semakin rapat dan semakin panjang trikoma meng-indikasikan genotipe kedelai semakin tidak disenangi sebagai sumber pakan larva ulat grayak. Di antara berbagai karakter morfologik daun, trikoma potensial sebagai perta-hanan fisik kedelai terhadap gangguan serangga pemakan daun, khususnya S.litura (Singh
et al. 1971). Fungsi trikoma sebagai alat pertahanan fisik daun adalah untuk mengganggu
mekanisme makan, oviposisi, pergerakan hingga hinggap, keracunan dan gangguan lain dari allelokinin dan kelenjar yang terdapat pada trikoma. Pada saat datang dan kontak dengan tanaman, serangga dapat terperangkap pada trikoma daun sehingga menghalangi
pergerakannya dan mencegah serangga mencapai permukaan daun untuk makan (Kush dan Panda 1995).
KESIMPULAN
1. Trikoma pada daun (panjang dan kerapatan) lebih berperan dibandingkan dengan karakter morfologi daun lainnya (ketebalan dan luas daun) dalam menentukan ketahanan kedelai terhadap hama ulat grayak.
2. Peran trikoma sebagai penyebab ketahanan kedelai terhadap ulat grayak diindikasikan dengan terganggunya daya makan larva sehingga bobot larva menjadi lebih rendah dan umur larva lebih panjang.
3. Trikoma pada daun kedelai prospektif digunakan sebagai karakter perbaikan ketahanan kedelai terhadap hama ulat grayak. Genotipe G100H yang memiliki trikoma rapat dan panjang dapat digunakan sebagai sumber gen ketahanan.
DAFTAR PUSTAKA
Al Ayedh, H. 1997. Antixenosis: the effect of plant resistance on insect behavior. Hayala@la-mar.colostate.edu. (Diakses 23 April 2012).
BPS (Biro Pusat Statistik). 1999. Luas dan intensitas serangan OPT dan bencana alam padi, palawija dan sayuran di Jawa, Tahun 1997. BPS, Jakarta.
Clark, W.J., F.A. Harris, F.G. Maxwell and E.E. Hartwig. 1972. Resistance of certain soybean cultivars to bean leaf beetle, striped blister beetle and bollworm. J. Econ. Entomol. 65: 1669−1672.
Endo, N, I. Hirakawa, T. Wada, and S.Tojo. 2007. Induced resistance to the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae) in three soybean cultivars. Applied Entomology and Zoology 42: 199−204.
Kilen, T.C., J.H. Hatchetth and E.E. Hartwig. 1977. Evaluation of early generation soybeans for resistance to soybean looper. Crop Sci. 17: 397−398.
Komatsu, K, M. Takahashi, Y. Nakazawa. 2010. Genetic study on resistance to the common cutworm and other leaf-eatinf insect in soybean. JARQ 44 : 117−125.
Kush, G.S, N. Panda. 1995. Host Plant Resistance to Insect. International Rice Research Institute. Manila. 431 hlm.
Lambert, L. and T.C. Kilen. 1984. Influence of three soybean plant genotypes and their F1 intercrosses on the development of five insect species. J. Econ. Entomol. 77 : 622−625. Nobuyuki, E., W. Takashi, M. Nobou and T. Masakazu. 2002. Possible resistance and tolerance
of a soybean breeding line, Kyukei 279 to the common cut worm, Spodoptera litura and soybean stink bugs. Proc of the Ass for Plant Protection of Kyushu 48: 68–71.
Singh, B.B., L.A. Andres and R.L. Bernard. 1971. Morphology of pubescence in soybeans and its relationship to plant vigor. Crop Sci. 11: 13−16.
Teestes, G.L. 1996. Plant resistance to inscts : a fundamental components of IPM. Radeliffes IPM World Textbook. The Univ. of Minnesota.
van Duyn, J.W., S.G. Turnipseed and J. D. Maxwell. 1971. Resistance in soybeans to the Mexican bean beetle. I. Source of resistance. Crop Sci. 11: 572−573.
Wier, A.T. and D.J. Boethel. 1996. Symbiotic nitrogen fixation and yield of soybean following defoliation by soybean looper during pod and seed development. J. Econ. Entomol. 89: 525−535.