Brain Research 880 (2000) 198–201
www.elsevier.com / locate / bres
Short communication
Tetanus-induced asynchronous GABA release in cultured hippocampal
neurons
a a ,
*
bKimmo Jensen , John D.C. Lambert
, Morten Skovgaard Jensen
a
Department of Physiology, University of Aarhus, DK-8000 Aarhus C, Denmark
b
Department of Neurobiology at the Institute of Anatomy, University of Aarhus, DK-8000 Aarhus C, Denmark
Accepted 25 July 2000
Abstract
Asynchronous GABA release was studied in cultured hippocampal neurons using paired whole-cell recordings. Tetanization of the presynaptic GABAergic neuron was accompanied by a train of IPSCs which showed tetanic depression. Asynchronous IPSCs (asIPSCs) also developed during the train and continued for 1.8560.3 s after the stimulation. The threshold frequency for evoking asIPSCs was 10
21
Hz, while maximal asynchronous activity was achieved at 40 Hz. Perfusion with EGTA-AM blocked asIPSCs. The elevation of [Ca ]i that accompanies presynaptic action potential firing triggers asynchronous release of GABA vesicles, thereby counteracting tetanic depression of synchronous IPSCs. 2000 Elsevier Science B.V. All rights reserved.
Theme: Excitable membranes and synaptic transmission
Topic: Mechanisms of neurotransmitter release
Keywords: IPSC; Asynchronous release; GABA; Cultured neurons; Hippocampus
The vesicle release trigger at central synapses binds 2.5, glucose 10, HEPES 10, CNQX (10mM) and DL-AP5
21
Ca with a relatively low affinity and rapid synchronous (50mM). pH was adjusted to 7.35 with NaOH (20–228C).
21
release of transmitter requires a local 100-fold elevation of Osmolality was 305 mosm kg . Patch-electrodes (3–6
21
[Ca ] , [3,13]. Nerve terminals are also capable ofi MV) were fabricated from borosilicate glass (o.d. 1.2 mm) releasing transmitters asynchronously, for example by on a P-97 puller (Sutter). The presynaptic electrode increasing the rate of spontaneous quantal release follow- contained (in mM): KOH 140, EGTA 11, CaCl 1, MgCl2 2
ing action potentials (APs) [12]. Although the asynchron- 2, NaCl 15, HEPES 10, leupeptin 0.1, MgATP 2; pH ous release of glutamate and acetylcholine has been well adjusted to 7.3 with methanesulfonic acid (290 mosm
21
described [1,4], less is known about asynchronous release kg ). The postsynaptic electrode contained (in mM): from GABAergic terminals [15]. We have recently ob- CsCl 120, TEACl 10, EGTA 11, CaCl2 1, MgSO4 1, served that tetanic stimulation of GABAergic neurons leupeptin 0.1, MgATP 4, QX-314 5, pH adjusted to 7.3
21
evokes a barrage of asynchronous postsynaptic GABAA with CsOH (290 mosm kg ). Dual whole-cell voltage-receptor mediated events that continued for several sec- clamp recordings were made using Axopatch 200 and onds after the train [9]. In the present report, we have 200A amplifiers (Axon Instruments). Stimulation pulse characterized this asynchronous activity further, and in- protocols were delivered by a pulse-generator (Master 8, vestigated its dependence on the presynaptic firing pattern. AMPI). The GABAergic neuron was stimulated by step-A detailed description of the methods has been given ping from270 mV to 0 mV for 2–3 ms, which evoked a previously [9]. During experiments, the neurons were GABA -mediated IPSC in the postsynaptic neuron. SingleA
perfused at 1 ml?min with an extracellular medium con- control stimuli were given at 0.2 Hz, while stimulus trains taining (in mM): NaCl 140, KCl 3.5, CaCl 2.5, MgCl2 2 were delivered at frequencies ranging from 5 to 80 Hz. Currents were filtered at 10 kHz, digitized using an AD-converter (Instrutech VR-100B), stored on a VTR and
*Corresponding author. Tel.:145-8942-27-84; fax:145-8612-90-65.
E-mail address: [email protected] (J.D.C. Lambert). analysed on a Pentium PC equipped with pClamp 6.0
K. Jensen et al. / Brain Research 880 (2000) 198 –201 199
(Axon Instruments). All data are presented as trace with respect to the baseline for 1 s following the train (Fig. 1B). In order that comparisons could be made means6S.E.M. with n indicating the number of pairs of
between neurons, this area was normalized to the mean neurons tested. Changes were considered to be significant
area of 10 single pre-tetanic IPSCs. asIPSC expression was at P-values, 0.05. Drugs and chemicals were purchased
examined in 20 pairs of neurons using 40 Hz stimulation. from Sigma, except CNQX and DL-APV (Tocris) and
The normalized asIPSCs area was found to depend on the QX-314 (Alomone).
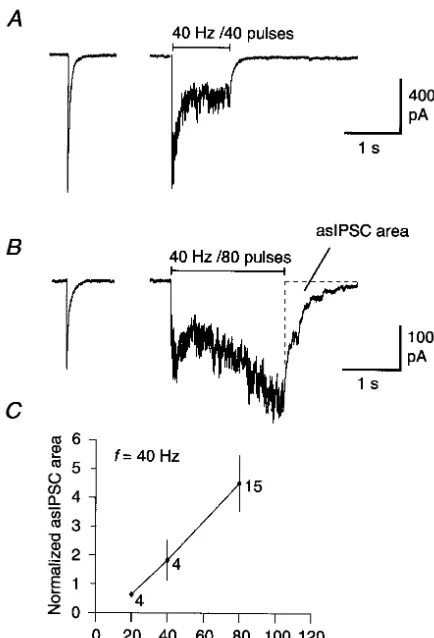
number of stimuli in the train. For fewer than 20 stimuli Tetanic stimulation of the GABAergic neuron at 40 Hz
there was no asIPSC activity. The current returned smooth-for 1 s (40 pulses) or 2 s (80 pulses) evoked a train of
ly to the baseline circumscribing a normalized area of synchronous IPSCs that showed tetanic depression (Fig.
0.660.06 (Fig. 1C, n54). Following 40 stimuli, the area 1A and B). Recovery to baseline following the train was
was 1.860.7 (n54) and after 80 stimuli the area was often accompanied by marked asIPSC activity (Fig. 1B).
4.561.0 (n515). Extrapolation showed that the threshold asIPSCs were quantified by measuring the area under the
for evoking asIPSCs at 40 Hz was around 20 stimuli (Fig. 1C).
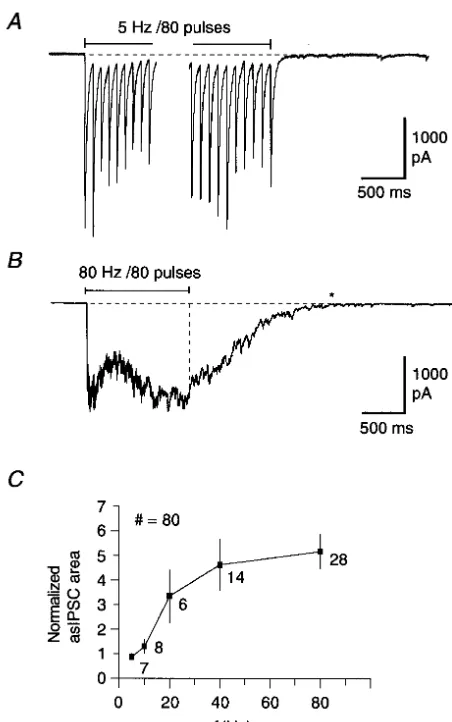
We then examined the dependency of asIPSC activity on the frequency of stimulation. Trains of 80 pulses were delivered at frequencies ranging from 5 to 80 Hz. asIPSC activity was essentially absent after a train of 5 Hz for 16 s (Fig. 2A), but was markedly expressed after 80 Hz for 1 s (Fig. 2B). Results from 63 trains (n553) are pooled in Fig. 2C and show that the threshold frequency for evoking asIPSCs is around 10 Hz. For trains of 80 Hz for 1 s, the duration of the macroscopic asIPSC activity from the end of the train to the return to baseline (indicated by the star in Fig. 2B) was also measured. asIPSC activity was present for 1.8560.3 s after the train (n519). We have previously shown that asIPSC activity is blocked by perfusion with EGTA-AM [9]. This allowed us to isolate asIPSC activity by subtracting the responses before and after perfusion with EGTA-AM (50 mM; Fig. 3). With stimulation at 80 Hz for 1 s, the subtraction showed that asIPSCs started to appear after a latency of about 20 pulses and continued for several seconds into the post-tetanic period. Similar results were found in four other pairs of neurons.
We report that repetitive presynaptic AP firing evokes asynchronous release of GABA at the level of unitary GABAergic connections [5]. This asynchronous activity contributes significantly to the GABA -mediated post-A
synaptic current, both during and for several seconds after the train. Since EGTA-AM blocks asynchronous release [4,15,18], it is likely to be caused by an increase in
21
Fig. 1. Post-tetanic asIPSCs depend on the number of pulses in the train. ambient [Ca ] . In cortical neuron boutons, ambient
i
(A) Trace from a postsynaptic neuron (Vhold5270 mV) showing IPSCs 21
[Ca ] is increased by 150 nM following a single APi
evoked by stimulation of the presynaptic GABAergic neuron. Five
[11]. Relating this value to our results suggests that
seconds after evoking a single IPSC (left), train stimulation at 40 Hz for 1
21
s was applied to the presynaptic neuron. This evoked a train of IPSCs [Ca ] increased to 3i mM after 20 APs, and to 12 mM
showing tetanic depression of about 60%. In this case, the current trace after 80 APs. On the assumption that asIPSCs are caused
21
returned smoothly to baseline immediately after stimulation. (B) Experi- by an elevation in ambient [Ca ] , the threshold would
i
ment from another pair of neurons. Following a train of 40 Hz of 2 s,
then be in the low micromolar range. We believe that
marked asynchronous IPSC (asIPSC) activity was observed. The area
tetanus-induced asIPSCs reflect bursts of quantal
ex-enclosed by the stippled line (asIPSC area) was measured and normalized
to the mean area of ten single pre-tetanic IPSCs, giving a ratio of 2.5 in ocytotic events at the active zones, as suggested by
this neuron. Note that, while the IPSCs showed tetanic depression during Behrends and ten Bruggencate [2]. Assuming that mIPSCs the initial part of the train, the overall current increased progressively have a t of 20 ms at 228C (M.S. Jensen et al.,
decay
thereafter and ultimately showed tetanic facilitation. (C) Summary of data
unpublished observation; [2]), calculations show that
quan-for 40 Hz trains consisting of 20, 40 and 80 pulses (23 trains; n520). The
tal events occurring at an inter-event interval smaller than
normalized asIPSC area increased steeply with the number of pulses in
200 K. Jensen et al. / Brain Research 880 (2000) 198 –201
Fig. 3. asIPSC activity isolated using EGTA-AM. The upper record shows a train of IPSCs evoked by presynaptic stimulation at 80 Hz for 1 s. Substantial asIPSC activity occurred during the train and continued into the post-tetanic period. Perfusion of EGTA-AM (50 mM) for 10 min abolished both the progressive increase in total current during the train and most of the asIPSC activity after the train. Subtraction of the responses shows the asIPSC activity in isolation. asIPSCs started to appear after about 0.25 s (20 pulses) and increased until the end of the train. asIPSC activity persisted for 2.8 s following stimulation.
This is in accordance with the observation that
replenish-21
ment of vesicles is accelerated by increased [Ca ]i
Fig. 2. Occurrence of asIPSCs depends on the frequency of stimulation.
[6,17,19]. If an increased replenishment of vesicles occurs,
(A) Trace showing the initial and final part of a train of IPSCs evoked by
it may seem surprising that synchronous release remained
presynaptic stimulation at 5 Hz for 16 s (80 pulses). Little asIPSC activity
is present following the stimulation. (B) Experiment from another pair of depressed, while asynchronous release increased. How-neurons. Substantial asIPSC activity is present following stimulation of ever, this disparity could reflect the fact that synchronous the presynaptic neuron at 80 Hz for 1 s. The normalized asIPSC area
release requires docking and priming of vesicles, and the
(calculated as in Fig. 1B) was 10.6 and the macroscopic current returned
binding of molecules that inhibit bursts of exocytotic
to baseline after 1.6 s (marked by *). (C) Summary of results showing the
events, such as Rab3A [8]. These steps could be rate
normalized post-tetanic IPSC area plotted against the frequency ( f ) for
trains containing 80 pulses. A normalized asIPSC area of about 0.7 limiting for synchronous release.
corresponds to a smooth return to baseline without evidence of asIPSC In conclusion, asynchronous release of GABA causes a activity. The threshold for evoking asIPSCs is around 10 Hz, while there
slowly rising macroscopic current that counteracts tetanic
is little further increase in asIPSCs at frequencies above 40 Hz (63 trains; 21
depression of synchronous IPSCs. When [Ca ] is raised
n553). i
to the micromolar level, GABA-vesicles could be mobil-ized from a reserve pool and released asynchronously. At
2
Cl current in the postsynaptic neuron. Therefore, the least 20 APs delivered above 10 Hz are required to activate major determinants for asIPSC expression would be the this process. This infers that asIPSCs would be prevalent
21
ability of ambient [Ca ] to rise to micromolar levels andi after sustained high-frequency firing of interneurons and in
21
spatio-temporal summation of mIPSCs, but might also be states with a decreased intracellular Ca buffering and influenced by re-uptake of GABA [14]. clearance.
In contrast to the enhancement of asIPSCs, synchronous IPSCs displayed tetanic depression [10,20]. Vesicles docked at the plasma membrane are thought to form a
readily releasable pool [7,16], inferring that tetanic depres- Acknowledgements
sion is caused by physical depletion of this pool. Our
K. Jensen et al. / Brain Research 880 (2000) 198 –201 201
[10] K. Jensen, J.D.C. Lambert, M.S. Jensen, Activity-dependent
depres-financial support. We thank K. Kandborg for preparation of
sion of GABAergic IPSCs in cultured hippocampal neurons, J.
the cultures and S. Kristensen for technical help.
Neurophysiol. 82 (1999) 42–49.
21
[11] P.J. MacKenzie, M. Umemiya, T.H. Murphy, Ca imaging of CNS axons in culture indicates reliable coupling between single action
References potentials and distal functional release sites, Neuron 16 (1996)
783–795.
[12] R. Miledi, R. Thies, Tetanic and post-tetanic rise in frequency of [1] P.P. Atluri, W.G. Regehr, Delayed release of neurotransmitter from
miniature end-plate potentials in low-calcium solutions, J. Physiol. cerebellar granule cells, J. Neurosci. 18 (1998) 8214–8227.
(Lond.) 212 (1971) 245–257. [2] J.C. Behrends, G. ten Bruggencate, Changes in quantal size dis- 21
[13] E. Neher, Vesicle pools and Ca microdomains: new tools for tributions upon experimental variations in the probability of release
understanding their roles in neurotransmitter release, Neuron 20 at striatal inhibitory synapses, J. Neurophysiol. 79 (1998) 2999–
(1998) 389–399. 3011.
[14] A. Roepstorff, J.D.C. Lambert, Factors contributing to the decay of [3] R. Bertram, A. Sherman, E.F. Stanley, Single-domain / bound
cal-the stimulus-evoked IPSC in rat hippocampal CA1 neurons, J. cium hypothesis of transmitter release and facilitation, J.
Neuro-Neurophysiol. 72 (1994) 2911–2926.
physiol. 75 (1996) 1919–1931. 21
[15] E. Rumpel, J.C. Behrends, Sr -dependent asynchronous evoked [4] D.D. Cummings, K.S. Wilcox, M.A. Dichter, Calcium-dependent
transmission at rat striatal inhibitory synapses in vitro, J. Physiol. paired-pulse facilitation of miniature EPSC frequency accompanies
(Lond.) 514 (1999) 447–458. depression of EPSCs at hippocampal synapses in culture, J.
Neuro-[16] T. Schikorski, C.F. Stevens, Quantitative ultrastructural analysis of sci. 16 (1996) 5312–5323.
hippocampal excitatory synapses, J. Neurosci. 17 (1997) 5858–
´ ¨
[5] D. Debanne, N.C. Guerineau, B.H. Gahwiler, S.M. Thompson,
5867. Physiology and pharmacology of unitary synaptic connections
[17] C.F. Stevens, J.F. Wesseling, Activity-dependent modulation of the between pairs of cells in areas CA3 and CA1 of rat hippocampal
rate at which synaptic vesicles become available to undergo slice cultures, J. Neurophysiol. 73 (1995) 1282–1294.
exocytosis, Neuron 21 (1998) 415–424. [6] J.S. Dittman, W.G. Regehr, Calcium dependence and recovery
´
[18] W. Van der Kloot, J. Molgo, Facilitation and delayed release at about kinetics of presynaptic depression at the climbing fiber to Purkinje
08C at the frog neuromuscular junction: effects of calcium chelators, cell synapse, J. Neurosci. 18 (1998) 6147–6162.
calcium transport inhibitors, and okadaic acid, J. Neurophysiol. 69 [7] L.E. Dobrunz, C.F. Stevens, Heterogeneity of release probability,
(1993) 717–729. facilitation, and depletion of central synapses, Neuron 18 (1997)
[19] L.-Y. Wang, L.K. Kaczmarek, High-frequency firing helps replenish 995–1008.
the readily releasable pool of synaptic vesicles, Nature 394 (1998) ¨
[8] M. Geppert, Y. Goda, C.F. Stevens, T.C. Sudhof, The small
GTP-384–388. binding protein Rab3A regulates a late step in synaptic vesicle
[20] K.S. Wilcox, M.A. Dichter, Paired pulse depression in cultured fusion, Nature 387 (1997) 810–814.
hippocampal neurons is due to a presynaptic mechanism indepen-[9] K. Jensen, M.S. Jensen, J.D.C. Lambert, Post-tetanic potentiation of
dent of GABAB autoreceptor activation, J. Neurosci. 14 (1994) GABAergic IPSCs in cultured hippocampal neurones, J. Physiol.