TINJAUAN PUSTAKA
Proses Fermentasi yang Melibatkan Konsorsium Mikroba
Makanan fermentasi adalah substrat makanan yang ditumbuhi oleh mikroba, dapat dimakan (edible) dan mengandung berbagai macam enzim yaitu: amilase, protease, lipase dan enzim penghidrolisis polisakarida. Selain itu pada makanan fermentasi dapat ditemukan pula protein dan lipid sebagai produk non toksik serta cita rasa, aroma serta tekstur yang menyenangkan dan menarik untuk dikonsumsi oleh manusia (Steinkraus 1997). Proses fermentasi makanan dapat dikategorikan menjadi proses fermentasi spontan yang tidak menggunakan kultur starter (NKS) dan proses fermentasi menggunakan kultur starter (KS). Kultur starter yang digunakan dapat berupa isolat tunggal atau kultur campuran. Proses fermentasi spontan (NKS) maupun fermentasi KS, masih memungkinkan masuknya berbagai mikroba lain yang akhirnya terlibat dalam proses fermentasi tersebut (Holzapfel 2002). Proses tersebut dapat pula melibatkan berbagai jenis bakteri terkultur maupun tidak terkultur (Ampe et al. 1999).
Tahapan proses fermentasi dapat didominasi oleh mikroba yang berbeda-beda. Hal ini disebabkan karena adanya kondisi pertumbuhan yang dibentuk oleh komunitas mikroba dalam proses fermentasi (Ampe et al. 1999; Randazo et al. 2002; Ogier et al. 2004). Variasi jenis dan lokasi mikroba pada proses fermentasi juga melibatkan perannya yang berbeda-beda. Adanya interaksi mikroba yang berkorelasi secara negatif maupun positif dengan menekan pertumbuhan organisme tertentu atau mendukung pertumbuhan organisme lain (Caplice & Fitzgerald 1999; Monier et al. 2008). Perubahan populasi mikroba dalam suatu proses fermentasi terjadi karena metabolit tertentu dapat dibentuk oleh mikroba tertentu untuk menunjang suksesinya dengan menciptakan lingkungan yang memungkinkan pertumbuhannya (Van der Meulen et al. 2007). Jenis mikroba berbeda dapat mensintesis metabolit berbeda walaupun pada bahan makanan yang sama. Jenis metabolit yang disintesis pada bahan makanan tertentu sangat dipengaruhi oleh jenis mikroba dalam proses fermentasinya (de Reu et al. 1994; Bauman & Bisping 1995; Denter et al. 1998; Hubert et al. 2008).
Komposisi mikroba yang terlibat dalam proses fermentasi makanan sangat penting karena berkontribusi terhadap cita rasa, tekstur dan nilai gizi dari makanan. Produksi komponen aroma juga dapat dihasilkan atau dimodifikasi dari eksopolisakarida dan protein yang ada pada bahan makanan. Komponen nutrisi seperti vitamin juga dapat dihasilkan karena peranan mikroba dalam proses fermentasi (Holzaffel 2002; Hugenholtz 2008). Secara tradisional komposisi mikroba dalam proses fermentasi yang menghasilkan produk tertentu dapat digunakan kembali untuk mempercepat kondisi fermentasi pada proses fermentasi selanjutnya. Hal ini dilakukan dengan menggunakan kultur backslopping atau dengan menggunakan residu fermentasi kering (Holzaffel 2002; Kourkoucas et al. 2006).
Pentingnya Fingerprinting komunitas Sebagai Sistem Barcoding Pada Proses Fermentasi
Produk fermentasi yang beragam dapat dihasilkan dari bahan baku yang sama. Mikroba merupakan parameter paling sulit untuk dikontrol selama proses fermentasi. Mikroba dalam proses fermentasi keju, dapat berasal dari kultur bakteri asam laktat yang ditambahkan maupun kultur bakteri asam laktat lain dalam starter. Bakteri lain, khamir dan fungi dapat juga timbul secara spontan dari lingkungan. Populasi mikroba yang tumbuh selama proses fermentasi sangat sulit dikontrol karena kompleksnya dinamika dan interaksi antar mikroba. Pengetahuan tentang struktur dan dinamika dari komunitas mikroba yang terlibat menjadi penting karena dapat memberi pemahaman lebih baik tentang variasi karakteristik produk fermentasinya. Hal ini disebabkan adanya pertumbuhan dan metabolisme mikroba yang ada. Kontrol komposisi mikroba akan memudahkan pemilihan karakteristik tertentu dan mencegah penurunan kualitas produk fermentasi (Jani & Barbier 2008).
Komposisi mikroba dalam suatu proses fermentasi sangat tergantung pada teknologi yang digunakan seperti pada keju (Ogier et al. 2004) serta daerah geografis tempat proses fermentasi tersebut dilakukan (Scheirlinck et al. 2007). Upaya memperoleh gambaran tentang hasil produksi fermentasi yang reprodusibel perlu dilakukan dengan diawali tahapan pembuatan database dan fingerprinting
mikroba dalam proses fermentasinya secara menyeluruh (Ogier et al. 2004). Studi tentang fingerprinting komunitas kultur starter akan membantu memberi pemahaman hubungan antara keragaman mikroba dan karakteristik spesifik yang dihasilkan. Analisis ini dapat menjadi dasar perancangan suatu kultur starter dan pengontrolan proses fermentasi (Lv et al. 2012)
Teknik Culture-Independent untuk Mempelajari Mikroba yang Terlibat dalam Suatu Proses Fermentasi Makanan
Upaya untuk mempelajari komunitas mikroba dalam suatu proses fermentasi makanan secara tradisional perlu dilakukan dan tidak hanya terbatas pada teknik culture-dependent saja tetapi juga penting untuk melihat keragaman komunitas dengan menggunakan teknik berbasis culture-independent (Ampe et al. 1999). Keterbatasan metode culture-dependent dalam kurun waktu dua dekade
menyebabkan adanya pengembangan teknik culture-independent yang
berdasarkan pada analisa asam nukleat. Teknik ini juga telah banyak digunakan dalam mempelajari komunitas mikroba pada ekosistem makanan untuk mengontrol produksi dan jaminan kualitas makanan yang baik (Juste et al. 2008).
Teknik culture-independent merupakan teknik yang mampu memberikan gambaran komunitas dalam suatu sistem makanan. Keterbatasan teknik culture-independent sangat dipengaruhi oleh kemampuan metode ekstraksi DNA yang digunakan. Ekstraksi DNA dari makanan dan ada tidaknya nuklease maupun inhibitor PCR yang dapat terekstraksi dari bahan makanan tersebut sangat mempengaruhi kualitas DNA hasil ekstraksi (Metwally et al. 2008). Metode yang digunakan haruslah metode terbaik untuk sampel spesifik, kriteria utamanya didasarkan pada kualitas dan kuantitas DNA hasil ekstraksi (Jara et al. 2008). Kuantitas dan kualitas DNA bersifat sangat kritis untuk interpretasi analisis komunitas mikroba (Thakuria et al. 2008). Ekstraksi DNA dari tempe belum banyak dipelajari padahal metode ekstraksi sangat bersifat spesifik untuk matriks atau bahan dasar makanan yang berbeda (Abrioruel et al. 2006; Jara et al. 2008).
Teknik culture-independent umumnya melibatkan teknik PCR sehingga penting untuk memperhatikan gen yang menjadi target dasar analisa serta metode ekstraksi DNA-nya. Pemilihan gen sebagai marker genetik harus dapat
membedakan variasi organisme secara luas. Sekuen gen yang dipilih harus memiliki sekuen variabel dan konservatif dimana domain yang variabel memungkinkan diskriminasi dalam selang yang luas dari suatu tingkatan taksonomi. Sekuen target ini harus diapit oleh daerah yang koservatif. Gen yang dapat digunakan sebagai penanda (marker) dapat dikelompokkan menjadi dua yaitu ubiquitously conserved genes dan functional genes (Juste et al. 2008).
Automated Intergenic Spacer Analysis (ARISA)
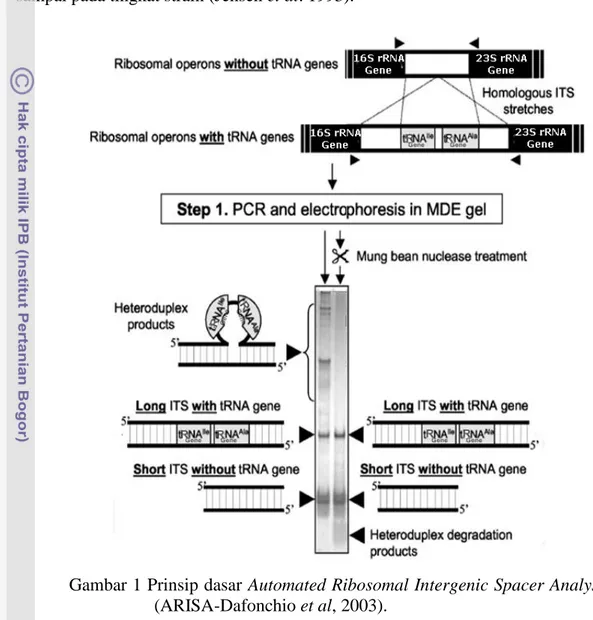
Metode rRNA Intergenic Spacer Analysis (RISA) merupakan salah satu metode culture-independent yang melibatkan amplifikasi PCR daerah intergenik antara gen 16S dan 23S rRNA. Daerah ini menunjukkan heterogenitas tinggi yang bergantung pada spesies baik dalam hal panjang amplikon maupun sekuen yang dihasilkan (Gambar 1). Kedua tipe variasi pada teknik ARISA ini telah banyak digunakan untuk membedakan strain-strain maupun spesies yang berkerabat secara dekat (Jensen et al. 1993; Maes et al. 1997) dan daerah ini dapat pula digolongkan sebagai ubiquitously conserved genes. Fisher dan Triplett (1999) menggunakan teknik ini dengan memodifikasi teknik RISA dan menggunakan primer oligonukleotida terlabel fluoresens pada proses PCR. Teknik ini
selanjutnya disebut Automated RISA (ARISA). Metode ARISA sangat
memudahkan analisis data yang dihasilkan.
Analisis ARISA digunakan sebagai tahapan lanjut analisis konsorsium mikroba tempe karena daerah sekuen intergenik 16S-23S rRNA memiliki variasi panjang dengan variasi ketidak miripan (dissimilarity) sekuen yang tinggi (35-100%) (Song et al. 2004). Organisme dengan 99% kesamaan pada analisis gen 16S rRNA dapat terdeteksi sebagai organisme berbeda dengan teknik ini. Teknik ini mampu mengkarakterisasi dan membedakan struktur genetik komunitas mikroba berdasarkan kehadiran masing-masing populasi mikroba dalam komunitas tersebut (Ranjard et al. 2001; Danovaro et al. 2006). Tempe diproduksi dengan standar prosedur bervariasi sehingga sangat membutuhkan teknik ARISA untuk menganalisis konsorsium mikroba yang ada dan diduga cukup kompleks. Teknik ini memungkinkan mempelajari struktur komunitas mikroba pada tempe karena memiliki sensitivitas tinggi dan dapat mengukur intensitas profil yang
terdeteksi secara tepat (Ranjard et al. 2001). Profil ARISA (Gambar 1) sangat bergantung pada jumlah kopi operon gen rRNA yang ada pada sel (Dafonchio et al. 2003) dan bersifat spesifik karena mampu memilah keragaman mikroba sampai pada tingkat strain (Jensen et al. 1993).
Gambar 1 Prinsip dasar Automated Ribosomal Intergenic Spacer Analysis (ARISA-Dafonchio et al, 2003).
Aplikasi teknik ARISA perlu ditunjang dengan penggunaan primer yang tepat dan andal terutama dalam menganalisa bahan makanan. Hal ini perlu diperhatikan agar tidak menutupi kemungkinan ditemukannya mikroba yang secara teknis tidak diinginkan atau belum pernah terdeteksi keberadaannya pada bahan makanan dan sulit teridentifikasi dengan teknik yang berbasis culture-dependent (Ogier et al. 2004). Cardinale et al. (2004), telah mencoba membandingkan satu set primer dan menemukan bahwa pasangan primer ITSF dan ITSReub yang didisainnya mampu menunjang analisa ARISA. Pemanfaatan
primer tersebut mampu mengeksplorasi keragaman bakteri dengan memberikan tingkat keragaman yang tinggi, tidak menyebabkan bias dan menghasilkan fingerprinting molekuler yang mudah dianalisis (Gambar 2). Pasangan primer ini telah diuji terhadap berbagai macam sumber sampel dan mampu mereduksi bias dalam analisa ARISA yang memungkinkan diperolehnya gambaran secara global tentang komunitas suatu sistem ekologi. Pada analisis keragaman fungi, Ranjard et al. (2001) telah merancang pasangan primer 2234C dan 3126T yang mampu mendeteksi variasi fungi dan dapat digunakan untuk melihat keragaman fungi dalam kelompok taksonomi utama fungi (Gambar 3). Analisis keragaman dilakukan dengan melihat fragmen DNA yang muncul dengan ukuran berbeda sebagai peak dalam ARISA type. Fragmen DNA ini didefinisikan sebagai OTU (Operational Taxonomic Unit) (Hewson & Fuhrman 2004; Ramete 2009).
Profil ARISA yang baik sangat bergantung pada kualitas DNA total hasil isolasi. Metode ekstraksi DNA sangat mempengaruhi kuantitas dan kualitas DNA untuk berhasilnya tahapan amplifikasi (Simon et al. 1996; Abriouel et al. 2006; Thakuria et al. 2008). Metode yang digunakan dalam ekstraksi DNA total perlu dioptimasi untuk mendapatkan metode terbaik untuk memberi gambaran menyeluruh tentang total komunitas mikroba dalam suatu sistem ekologi. Bahan makanan dapat mengandung bahan pengganggu PCR dan metode yang digunakan harus mampu mengekstrak semua DNA dari sampel. Hal ini diperlukan agar dapat memberikan profil yang sebenarnya tentang semua mikroba yang ada (Abriouel et al. 2006; Ranjard et al. 2001). Faktor yang perlu diperhatikan dalam proses tersebut adalah korelasi nyata antara jumlah ribotipe reprodusibel yang dihasilkan dan ekstrak DNA hasil ekstraksi. Metode yang digunakan dalam ekstraksi DNA
total juga perlu dioptimasi untuk mendapatkan metode terbaik (Ranjard et al. 2001).
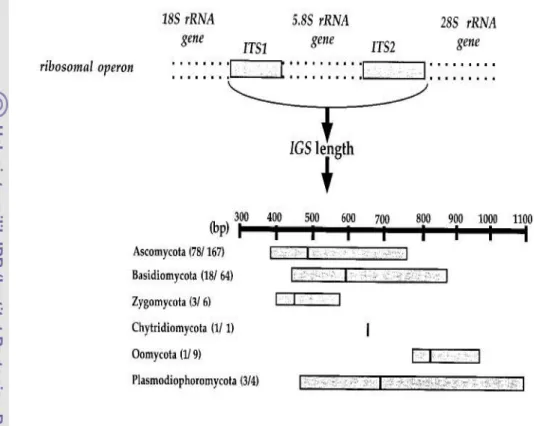
Pustaka Gen 16S rRNA
Gen 16S rRNA merupakan gen yang selalu ada pada semua prokariot dan menjalankan fungsi sama. Perubahan sekuen pada gen ini akan menandai jarak evolusi antara mikroba yang berkerabat. Intensitas perubahan dapat terjadi dengan kecepatan dan lokasi dalam gen 16S rRNA yang tidak seragam antara setiap mikroba. Panjang gen 16S rRNA adalah sekitar 1550 bp dan tersusun atas daerah variable dan konservatif. Gen ini cukup besar dan memiliki sifat polimorfisme interspesifik yang dapat digunakan sebagai pembeda nyata secara statistik. Primer universal dirancang sebagai fragmen komplemen pada awal dan ujung gen. Daerah gen yang variable diapit oleh primer dan dapat digunakan dalam analisis komparatif taksonomi (Clarridge 2004).
Gambar 3 Kedudukan dan ukuran amplikon yang dihasilkan dari primer 2234C dan 3126T (Ranjard et al.2001).
Weisburg et al. (1991) telah menemukan bahwa amplifikasi gen 16S-rRNA, kloning dan sekuensing merupakan metode untuk mengidentifikasi mikroorganime tanpa harus dikulturkan. Pendekatan pustaka gen 16S rRNA dapat dimanfaatkan dalam mengidentifikasi spesies spesifik yang berpotensi sebagai indikator dalam mempelajari struktur suatu komunitas (Hartman & Widmer, 2006). Marchesi et al. (1998) telah merancang sepasang primer yang dapat digunakan untuk mengamplifikasi gen 16S rRNA dari kelompok bakteri yang disebut primer 63F dan 1387R.
Tempe Sebagai Makanan Tradisional yang Kaya Nutrisi
Kedelai merupakan bahan makanan yang unik karena kaya akan nutrisi. Kedelai mengandung karbohidrat kompleks, protein nabati, lipid, serat, oligosakarida, senyawa fitokimia (khususnya isoflavon) dan mineral. Protein kedelai dikenal sebagai protein yang lengkap dan setara dengan albumin telur. Substitusi protein hewani dengan protein kedelai akan mereduksi ekskresi kalsium urin dan mereduksi osteoporosis. Kandungan serat yang larut dan tidak larut juga memberi keuntungan bagi kesehatan, sedangkan oligosakaridanya berperan sebagai komponen prebiotik di kolon. Isoflavon genistein dan daidzein dari kedelai memiliki sejumlah fungsi biologis dan mineralnya berada dalam bentuk yang tersedia sehingga menjadi penting karena mereduksi osteoporosis dan hipertensi (Anderson et al. 1999). Kedelai juga mengandung asam lemak gliserid seperti asam palmitat, stearat, oleat, linoleat dan linoleat (de Reu et al. 1994). Zamora & Veum (1979) menyatakan bahwa nutrisi kacang kedelai yang difermentasi dengan adanya fungi lebih signifikan meningkatkan average daily gain (ADG) dan rasio gain:feed (G:F) dari tikus. Hal ini menunjukkan bahwa proses fermentasi berperan penting dalam menyediakan nutrisi yang lebih siap untuk diserap.
Tempe merupakan makanan fermentasi tradisional Indonesia yang secara prinsip dibuat dari kedelai. Tempe di Indonesia terutama diproduksi oleh industri skala kecil dengan skala produksi 10 kg sampai 4 metrik ton per hari. Diduga terdapat lebih dari 100.000 produsen tempe yang tersebar di Indonesia (Astuti et al. 2000). Chan et al. (2007) melaporkan bahwa tempe merupakan sumber zinc,
besi dan kalsium yang terbaik bagi bayi karena kandungan mineralnya relatif tinggi dan kandungan phytat-nya rendah. Tempe yang berbahan dasar kedelai dan diproses melalui fermentasi tradisional diproses melalui tahapan proses soaking (perendaman dalam kondisi asam), dehulling (pengelupasan mantel biji), cooking (perebusan) dan fermentasi. Karena kedelai sebagai bahan baku tempe mengandung sejumlah senyawa antinutrisi dan senyawa yang bersifat toksik, Egounlety & Aworh (2003) melaporkan bahwa tahapan preparasi sebelum proses fermentasi perlu dilakukan sebagai tahapan persiapan substrat untuk selanjutnya mengalami pemecahan secara enzimatik oleh fungi. Pada tahapan preparasi ini juga, proses cooking secara signifikan mereduksi inhibitor trypsin pada kacang-kacangan, dehulling mengeliminasi tannin dan fermentasi dengan Rhizopus oligosporus mereduksi asam phytat sebanyak 30,7% pada kedelai. Proses Soaking-dehulling-cooking dan fermentasi dengan Rhizopus oligosporus mengeliminasi stachyose yang merupakan oligosakarida yang sangat flatulent.
Galur Rhizopus yang digunakan sebagai starter sangat beragam. Baumann & Bisping (1995) pernah melaporkan adanya 36 strain Rhizopus asal Indonesia yang biasa digunakan untuk membuat tempe. Rhizopus yang digunakan dalam pembuatan tempe mampu membentuk karotenoid dan menurunkan jumlah tocopherol dengan jumlah vitamin E yang tetap, menurunkan asam lemak gliserid (de Reu et al. 1994), mampu melakukan proteolisis, meningkatkan jumlah asam amino bebas dalam suatu proses fermentasi tempe (Baumann & Bisping 1995), serta memproduksi ergosterol (Denter et al. 1998). Proses fermentasi ini dapat memberikan hasil yang berbeda bila galur fungi dikombinasikan dengan bakteri dalam proses fermentasi (de Reu et al. 1994; Baumann & Bisping 1995; Denter et al. 1998). Kemampuan proteolisis R. stolonifer, R. oryzae dan R oligosporus dalam melakukan proteolisis sangat beragam, bahkan sangat bergantung pada galur dan bukan spesies (Baumann & Bisping 1995). Rhizopus juga telah
dilaporkan mampu mensintesis β-carotene namun kemampuan ini tidak dimiliki oleh semua spesies ini (Denter et al. 1998). Jenis asam lemak yang didegradasi dan dihasilkan juga sangat beragam (de Reu et al. 1994).
Walaupun proses fermentasi tempe umumnya menggunakan kultur starter tunggal yang terdiri atas fungi dari kelompok Rhizopus, beberapa penelitian telah
melaporkan adanya keterlibatan mikroba lain seperti bakteri dan kapang dalam proses ini. Liem et al. (1977) pernah membuktikan peran bakteri dalam membentuk vitamin B12 pada tempe (Tabel 1).
Tabel 1 Bakteri dan khamir yang dilaporkan berasosiasi dengan proses fermentasi tempe
Bakteri dan Khamir Produk yang dihasilkan Sumber
Lactobacillus casei Asam, Mulyowidarso et al. 1989
Streptococcus faecium Asam, invertase Mulyowidarso et al. 1989 Sterptococcus dysgalactiae Asam Mulyowidarso et al. 1989 Staphilococcus epidermidis Asam, invertase Mulyowidarso et al. 1989
Citrobacter diversus - Mulyowidarso et al. 1989
Enterobacter aglomerans - Mulyowidarso et al. 1989
Enterobacter cloacae - Mulyowidarso et al. 1989
Klebsiella pneumoniae Asam, invertase, α -galactosidase
Mulyowidarso et al. 1989
Klebsiella Ozaenae - Mulyowidarso et al. 1989
Bacillus brevis Asam Mulyowidarso et al. 1989
Citrobacter freundii Vitamin B12 Keuth & Bisping 1994 Klebsiella pneumoniae Vitamin B12 Keuth & Bisping 1994; Barus et al.
2008 Brevibacterium epidermidis Factor 2 Klus et al. 1993 Microbacterium aborescens Factor 2 dan Glycitein Klus et al. 1993 Micrococcus Isoflavon polyhydroxylasi Klus & Bars 1998 Arthrobacter Isoflavon polyhydroxylasi Klus & Bars 1998 Enterococcus faecium LMG 19827 Bakteriosin B1 Moreno et al. 2002 Enterococcus faecium LMG 19828 Bakteriosin B2 Moreno et al. 2002 Bacillus subtilis Protease serin dari kelompok
subtilisin
Kim et al. 2006; Barus et al. 2008 Enterobacteria bersifat proteolitik Barus et al. 2008
Acetobacter indonesiensis - Barus et al. 2008
Flavobacterium sp. - Barus et al. 2008
Klebsiella sp. - Barus et al. 2008
Brevundimonas sp. - Barus et al. 2008
Bacillus sp. - Barus et al. 2008
Pseudomonas putida - Barus et al. 2008
Acinetobacter - Barus et al. 2008
Pichia burtonii (khamir) Invertase Mulyowidarso et al. 1989. 1991 Candida diddensiae (khamir) - Mulyowidarso et al. 1989
Rhodotorula rubra (Khamir) - Mulyowidarso et al. 1989
Analisa yang berdasarkan teknik culture-independent menggunakan Amplified Ribosomal DNA Restriction Analysis (ARDRA) telah pula dilakukan dan ditemukan 60 filotipe bakteri pada tempe tidak pahit (Stefania 2009) dan 58 filotipe bakteri dari tempe dengan rasa pahit (Sari 2009) yang menggambarkan
tingginya keragaman bakteri pada tempe. Keterlibatan khamir dalam fermentasi tempe di Malaysia juga telah dilaporkan oleh Moreno et al. (2002).
Bakteri asam laktat juga telah dilaporkan berpengaruh terhadap komposisi beberapa produk makanan dengan bahan baku kedelai. Bakteri ini dilaporkan berpengaruh terhadap komposisi senyawa fitokimia dan sifat antioksidan dari kecambah kedelai sebagai hasil proses fermentasinya (Hubert et al. 2008). Pada produk pasta kedelai (doenjang), bakteri asam laktat dan bakteri-bakteri lain seperti Bacillus dilaporkan berperan dalam membentuk komponen-komponen pembentuk cita rasa, protein dan bakteriosin pada makanan ini (Kim et al. 2009). Kondisi lingkungan yang berbeda antara daerah penghasil tempe juga dapat berpengaruh terhadap hasil fermentasi. Baumann dan Bisping (1995) menyatakan bahwa suhu yang rendah akan menurunkan kecepatan fermentasi, namun justru dapat meningkatkan kandungan asam amino bebas pada tempe.
Mengacu pada penelitian yang dikemukakan di atas terlihat bahwa dalam proses fermentasi tempe di Indonesia melibatkan sangat banyak spesies Rhizopus sebagai kultur starter pada daerah produsen tempe yang berbeda-beda. Selain itu karena kondisi lingkungan produsen tempe yang berbeda-beda, dapat terjadi bahwa proses fermentasi melibatkan banyak sekali mikroba lain selain kultur starter. Varietas biji kedelai yang digunakan produsen tempe juga berbeda pada daerah penghasil tempe yang berbeda. Hal ini dapat berpengaruh pada hasil akhir fermentasi. Peran kapang Rhizopus, selain menyediakan beberapa komponen penting juga berperan untuk menyediakan substrat bagi bakteri dalam mensintesis komponen cita rasa dan komponen nutrisi lain.
Perbedaan rasa pada tempe dari berbagai daerah di Indonesia diduga dapat disebabkan karena adanya variasi komunitas mikroba, karena variasi bahan serta tempat proses fermentasi yang juga beragam (Astuti et al. 2000). Hal ini tentu saja menjadi faktor penunjang ciri khas rasa tempe yang dihasilkan pada daerah berbeda di seluruh Indonesia. Scheirlinck et al. (2007, 2008) menemukan bahwa dalam proses fermentasi sourdough yang dilakukan secara spontan melibatkan suatu konsorsium mikroba. Penelitian tersebut menemukan bahwa tidak terdapat kesamaan komunitas bakteri asam laktat pada proses fermentasi sourdough pada daerah yang berbeda. Skor plot dari Principal Component Analysis (PCA)
menunjukkan bahwa sampel sourdough yang berasal dari depositor yang sama akan dikelompokkan bersama. Korelasi antara tipe tepung, propinsi penghasil atau karakteristik teknologi (pH, waktu fermentasi dan suhu) tidak ditemukan demikian pula asosiasi signifikan antara spesies bakteri asam laktat tertentu dengan salah satu dari variabel yang diamati. Keragaman variasi komunitas bakteri asam laktat ini secara signifikan sangat dipengaruhi oleh lingkungan bakery tempat terjadinya fermentasi sourdough.
Walaupun berkontribusi secara positif, ternyata mikroba yang terlibat dalam proses fermentasi tempe dapat juga memberi dampak yang merugikan. Rhizonin adalah hepatotoksik siklopeptida dari kultur fungi Rhizopus microspores yang tumbuh pada kacang-kacangan di Mozambique. Penelitian menunjukkan bahwa toksin ini sebenarnya tidak diproduksi oleh fungi tetapi oleh bakteri dari genus Burkhoderia yang merupakan endosimbion dalam sitosol fungi (Partida-Martinez et al. 2007a, 2007b.). Rohm et al. (2010) menunjukkan bahwa bakteri toksigenik ini juga ditemukan pada kultur starter dalam produksi tempe atau sufu. Profil metabolik dari fungi selama kondisi standar untuk produksi tempe atau sufu, menunjukkan bahwa toxin dan rhizonin diproduksi dalam jumlah cukup kritis.
Biogenik amin berimplikasi terhadap kejadian keracunan yang berasosiasi dengan konsumsi makanan hasil fermentasi. Pada proses fermentasi makanan berbahan kedelai telah dilaporkan terjadi pada fermentasi sufu di Taiwan. Dari sembilan biogenik amin pada semua sampel sufu kurang dari 5 mg/100g, ternyata diperoleh satu sampel sufu cokelat yang memiliki kandungan histamin terbesar (15.8 mg/100 g) yang lebih besar dari 5.0 mg/100 g, limit standar yang diijinkan oleh US Food and Drug Administration (Kung et al. 2007). Pada bahan makanan fermentasi, bakteri asam laktat sangat berperan dalam memberikan aroma dan tekstur yang baik (Hugenholtz 2008). Walau demikian, bakteri asam laktat juga merupakan bakteri yang banyak memproduksi senyawa biogenik amin ini sehingga pengontrolan dan seleksi komposisi mikroba dalam proses fermentasi perlu dilakukan untuk menjaga keamanan pangan makanan hasil fermentasi (Marcobal et al. 2005).