ABSTRAK
Tanaman kedelai (Glycine max L.) dapat membe-rikan respons positif dan negatif terhadap perubahan lingkungan tumbuh di atas tanah maupun di dalam tanah. Respons tersebut dapat diketahui dari peru-bahan fenotipik dan fisiologis tanaman. Lingkungan di atas tanah yang berpengaruh terhadap pertum-buhan tanaman kedelai terutama adalah lama dan intensitas penyinaran, suhu udara, dan kandungan CO2 di atmosfer. Lama penyinaran yang optimal adalah 10–12 jam. Berkurangnya intensitas cahaya matahari menyebabkan tanaman tumbuh lebih tinggi, ruas antarbuku lebih panjang, jumlah daun dan jumlah polong lebih sedikit, dan ukuran biji semakin kecil. Respons kedelai terhadap perubah-an suhu tergperubah-antung pada fase pertumbuhperubah-an. Suhu yang sesuai pada fase perkecambahan adalah 15– 22 oC, fase pembungaan 20–25 oC, dan pada fase
pemasakan 15–22 oC. Peningkatan CO
2 atmosfer dari
349 μL menjadi 700 μL meningkatkan laju pertu-karan karbon (C), menurunkan laju transpirasi, dan meningkatkan efisiensi penggunaan air. Kondisi lingkungan di dalam tanah yang berperan terhadap pertumbuhan kedelai terutama adalah tekstur tanah, kadar air tanah dan unsur hara, unsur-unsur toksik, kemasaman tanah, suhu tanah, dan salinitas. Kedelai tumbuh baik pada tanah bertekstur ringan hingga berat, namun tanah yang padat (BI >1,38 kg/m3) tidak sesuai untuk kedelai. Kebutuhan air
tanaman kedelai pada fase generatif lebih tinggi dibandingkan pada fase vegetatif, sehingga pada fase generatif lebih peka terhadap kekeringan terutama pada fase pembungaan hingga pengisian polong. Kandungan air optimal adalah 70–85% dari kapasitas lapangan. Kandungan unsur hara tanah harus di atas batas kekahatan agar tanaman tumbuh optimal. Nilai kritis suatu unsur hara dalam tanah beragam tergantung jenis tanah dan metode analisis yang digunakan. Pengaruh suhu tanah terutama pada fase perkecambahan, dan suhu tanah optimal adalah 24,2–32,8°C. Kedelai agak sensitif terhadap kemasaman tanah, unsur-unsur toksik, dan salinitas. Nilai kritis pH, Al, Mn, dan salinitas berturut-turut adalah pH 5,5, Al-dd 1,33 me/100 g, Mn 3,3 ppm, dan 1,3 dS/m. Rhizobium berperan dalam memasok kebutuhan N tanaman kedelai, namun inokulasi
RESPONS TANAMAN KEDELAI TERHADAP
LINGKUNGAN TUMBUH
Abdullah Taufiq dan Titik Sundari 1)
tidak efektif pada tanah yang sering ditanami kedelai.
Kata kunci: Glycine max, interaksi, lingkungan
ABSTRACT
The response of soybean (Glycine max L.) to above and below ground environmental change can be posi-tive and negaposi-tive. The response can be identified from phenotipic as well as physiological change of the plant. The above ground environment affect soybean growth is mainly intensity and photoperi-ods, temperature and CO2 concentration. Optimum photoperiods is 10–12 hours. Decreasing sun shine intensity reduced plant hight, increase the intern-ode, decrease number of leaf, pod set and seed size. Response of soybean to temperature depends on the growth phase. The optimum temperature during seedling, flowering, and maturing stage is 15–22oC,
20–25oC, and 15–22oC respectively. Increasing CO 2
atmospher from 349 μL to 700 μL increase carbon exchange rate, reduce transpiration rate, and in-crease water use efficiency. The below ground con-dition affect soybean growth is mainly soil texture, soil moisture and nutrient content, soil acidity, toxic element, soil temperature, and salinity. Soybean is favourable on light to heavy texture soil. However, compacted soil (bulk density >1.38 kg/m3) is
unfavourable for soybean. Soybean of generative stage need more water than that of vegetative stage, so that soybean more sensitive to water stress dur-ing generative stage, especially durdur-ing flowerdur-ing and pod filling stages. Optimum soil moisture content for soybean is 70–85% of field capacity. Nutrient con-centration of soil must be above the critical deficiency level in order soybean grows normally. The critical deficiency level of a nutrient is various depends on soil type and the method of analysis. Soil tempera-ture affect mainly during seedling stage, and opti-mum is 24.2 to 32.8°C. Soybean is moderately sen-sitive to salinity, and the critical level is 1.3 dS/m. Rhyzobium has an important role for N supply for soybean, but inoculation mostly ineffective on soil where soybean is often cultivated.
Keywords: Glycine max L., interaction, environment
1 Balai Penelitian Kacang-kacangan dan Umbi-umbian
Jl. Raya Kendalpayak km 8 Kotak Pos 66 Malang 65101. email [email protected].
Naskah diterima tanggal 10 November 2011, disetujui untuk diterbitkan tanggal 13 Februari 2012.
Diterbitkan di Buletin Palawija No. 23: 13–26 (2012).
LINGKUNGAN TUMBUH
tanaman di atas tanah terdiri atas sinar matahari, suhu udara, kelembaban udara, kandungan gas di udara, dan hujan. Faktor lingkungan bagian tanaman di dalam tanah terdiri atas suhu tanah, kandungan air tanah, salinitas, pH, kandungan unsur hara, kan-dungan unsur toksik, tekstur dan struktur tanah, dan aerasi tanah. Komponen-komponen faktor lingkungan tersebut secara individu mau-pun interaksinya berpengaruh langsung maupun tidak langsung terhadap pertumbuhan tanaman.
Respons tanaman terhadap lingkungan ber-beda-beda tergantung jenis dan kultivar tanam-an. Tanaman dapat memberikan respons positif maupun negatif terhadap perubahan ling-kungan tumbuh. Respons yang beragam ter-sebut menimbulkan terjadinya interaksi antara lingkungan dengan genotipe, dan fenomena tersebut sering ditemui dalam pengujian multi-lokasi. Respons tersebut dapat diketahui dari perubahan fisik tanaman berupa perubahan pertumbuhan, dan perubahan fenotipik tana-man. Respons tanaman juga dapat diketahui dari perubahan proses fisiologis misalnya kecepatan fotosintesis, dan translokasi foto-sintat.
RESPONS TERHADAP CAHAYA, SUHU, DAN CO2
Kedelai adalah tanaman berhari pendek, yaitu tidak mampu berbunga bila penyinaran melebihi 16 jam, dan cepat berbunga bila kurang dari 12 jam. Lama penyinaran matahari di Indonesia umumnya sekitar 12 jam. Di In-donesia kedelai berbunga pada umur 25–40 hari dan panen pada umur 75–95 hari, sedangkan di wilayah subtropika dengan panjang hari 14– 16 jam kedelai berbunga umur 50–70 hari dan panen pada umur 150–160 hari. Lama penyi-naran optimal adalah 10–12 jam, penyipenyi-naran kurang dari 10 jam atau lebih dari 12 jam menyebabkan pembungaan lambat, penurunan jumlah bunga, polong, dan hasil, tetapi ukuran biji tidak terpengaruh dan menjadi lebih kecil bila penyinaran <6 jam (Arifin 2008). Penyi-naran terus menerus dengan sinar buatan menghasilkan total biomas dan total kapasitas source untuk fotosintesis yang lebih tinggi dibandingkan penyinaran selama 10 jam, akan tetapi laju fotosintesis/satuan luas daun dan produksi polong lebih rendah (Kasai 2008).
Kedelai sesuai pada lingkungan dengan intensitas cahaya dan suhu sedang, serta tanah cukup lembab. Pada daerah panas pertum-buhannya terhambat karena enzim RuBisCO (Ribulose-1,5-bisphosphate carboxylase oxyge-nase)mengikat banyak oksigen dengan mening-katnya suhu sehingga memacu fotorespirasi yang menyebabkan kehilangan karbon dan nitrogen sehingga menghambat pertumbuhan.
Kedelai memerlukan penyinaran penuh, tetapi dalam praktik budidaya di Indonesia, kedelai sering ditumpangsarikan dengan tana-man lain. Intensitas cahaya yang diterima kede-lai pada tumpangsari dengan jagung berkurang sekitar 33% (Asadi et al. 1997). Berkurangnya intensitas sinar matahari menyebabkan tana-man tumbuh lebih tinggi, ruas antar buku lebih panjang, jumlah daun lebih sedikit, jumlah polong makin sedikit, dan ukuran biji semakin kecil (Susanto dan Sundari 2010). Tanggapan tanaman kedelai terhadap cahaya berbeda antarvarietas. Varietas Pangrango lebih tang-gap terhadap peningkatan kuanta cahaya dari-pada varietas Wilis dan Brawijaya (Sitompul 2003).
Berdasarkan tipe fotosintesis, kedelai meru-pakan kelompok tanaman C3. Tingkat keje-nuhan fotosintesis kelompok tanaman C3 dicapai pada intensitas cahaya lebih rendah dibandingkan tanaman C4. Tingkat kejenuhan cahaya bagi individu daun kelompok tanaman C3 dicapai pada kisaran 50% cahaya penuh, sedangkan tanaman C4 hampir linier hingga 100% cahaya penuh (Odum 1983). Laju foto-sintesis lambat pada kondisi intensitas cahaya dan suhu tinggi. Fotosisntesis pada kondisi suhu dingin, lembab, namun cahaya normal lebih efisien dibandingkan tanaman C4. Pada kondisi panas dan kering, stomata menutup untuk mengurangi kehilangan air, tetapi menghambat pertukaran CO2 sehingga
menu-runkan laju fotosintesis. Pada tanaman C3, enzim yang menyatukan CO2 dengan substrat
untuk pembentukan karbohidrat (fotosintesis) pada saat bersamaan dapat mengikat O2 untuk proses fotorespirasi (pembongkaran karbohidrat untuk menghasilkan energi yang terjadi pada siang hari).
Peningkatan konsentrasi CO2 di atmosfer
Kedelai lebih adaptif pada kondisi CO2
atmosfer tinggi. Peningkatan konsentrasi CO2
dari 349 μL menjadi 700 μL meningkatkan laju pertukaran karbon (C), menurunkan laju transpirasi, dan meningkatkan efisiensi peng-gunaan air, serta sebagian besar C terpartisi ke dalam bentuk pati (Huber et al. 1984). Kedelai yang ditumbuhkan pada CO2 atmosfer
yang tinggi menunjukkan laju pertumbuhan yang lebih tinggi (Jones et al. 1984), luas daun yang lebih besar (Peet 1984), dan hasil yang lebih tinggi (Kimball 1983, Teramura et al. 1990). Peningkatan CO2 atmosfer pada suhu rendah menyebabkan pertumbuhan kedelai lebih baik (Sionit et al. 1987). Namun sebalik-nya, setiap peningkatan suhu 1oC menurun-kan hasil kedelai 17% (Lobell dan Asner 2003).
Kedelai banyak dibudidayakan di lingkung-an tropis dlingkung-an subtropis, dapat tumbuh hingga ketinggian tempat 1000 m dpl. Daya adaptasi yang luas tersebut karena kedelai relatif tahan pada kondisi suhu rendah dan tinggi. Suhu udara yang paling sesuai adalah 20–25 oC. Kecepatan pertumbuhan mengalami penu-runan pada suhu >35 °C maupun pada suhu <18 °C. Kondisi iklim dengan suhu dan radiasi UV-B tinggi dapat menyebabkan kerusakan organ reproduktif seperti morfologi bunga dan serbuk sari pada kedelai (Koti et al. 2005).
Sensitifitas kedelai terhadap perubahan suhu tergantung pada fase pertumbuhan. Suhu udara minimum untuk pertumbuhan vegetatif adalah 10 °C, untuk pembentukan polong dan biji adalah 15 °C. Suhu yang sesuai untuk kede-lai pada fase perkecambahan adalah 15–22 oC, fase pembungaan 20–25 oC, dan pada fase pemasakan 15–22 oC (Liu et al. 2008). Pening-katan suhu udara pada siang hari dari 30 oC menjadi 35 °C selama 10 jam dari fase pembu-ngaan hingga pemasakan menyebabkan penu-runan hasil 27%, akan tetapipeningkatan suhu pada malam hari dari 20 menjadi 30 oC tidak berpengaruh terhadap hasil dan komponen hasil (Gibson dan Mullen 1996).
Hasil biji kedelai meningkat seiring dengan peningkatan suhu antara 18/12 oC (siang/ malam) dan 26/20 oC, tetapi hasil mengalami penurunan pada suhu lebih tinggi dari 26/20
oC karena ukuran biji menjadi lebih kecil (Sionit
et al. 1987). Suhu tinggi (30/20oC) selama pem-bungaan dan pembentukan polong mengha-silkan jumlah polong yang lebih banyak, tetapi
suhu di atas 40 oC menghambat pembentukan polong (Lawn dan Hume 1985). Peningkatan suhu dari 19/20 oC menjadi 34/20 oC selama pengisian biji menurunkan hasil biji kedelai (Dornbos dan Mullen 1991).
RESPONS TERHADAP KONDISI FISIK TANAH
Karakter fisik tanah yang banyak berhu-bungan dengan pertumbuhan tanaman adalah tekstur, struktur, konsistensi, porositas, den-sitas, dan suhu tanah. Karakter fisik tanah ber-pengaruh terhadap perakaran dan proses fisiologi akar. Meskipun demikian hasil akhir dari pengaruh tersebut dapat terlihat pada bagian tanaman di atas tanah, misalnya ter-hambatnya pertumbuhan, dan menurunnya hasil biji. Karakter fisik tanah dapat berpe-ngaruh langsung maupun tidak langsung terhadap tanaman.
Kedelai dapat tumbuh pada tanah bertekstur ringan hingga berat. Akan tetapi Beutler et al. (2005) menunjukkan bahwa di Brazil pertum-buhan terhambat dan hasil kedelai turun mulai nilai ketahanan penetrasi tanah 0,85 Mpa dan berat isi tanah (BV) 1,48 kg/m3. Hal ini menun-jukkan bahwa tanah yang padat kurang baik untuk kedelai. Rismaneswati (2006) melapor-kan bahwa hasil kedelai sangat rendah (0,9– 1,0 t/ha) pada Alfisol yang mempunyai BV 1,29– 1,34 g/cm3. Botta et al. (2004) melaporkan bahwa peningkatan BV akibat pemadatan tanah dari 1,33 kg/m3 menjadi 1,38 kg/m3, 1,40 kg/m3, dan 1,51 kg/m3 menurunkan hasil kedelai dari 3,1 t/ha menjadi berturut-turut 2,8 t/ha, 2,4 t/ha, dan 1,9 t/ha atau terjadi penu-runan hasil berturut-turut 9,7%, 22,6%, dan 38,7%. Pemadatan tanah juga berpengaruh terhadap pembentukan bintil akar. Pada kondisi tanpa mulsa, jumlah bintil akar me-ningkat dengan makin padatnya tanah, namun terjadi sebaliknya jika diberi mulsa (Siczek dan Lipiec 2011).
RESPONS TERHADAP SUHU TANAH
RESPONS TERHADAP SALINITAS TANAH
Salinitas adalah tingkat kegaraman yang mengindikasikan jumlah garam terlarut dalam air. Ion yang dapat menyebabkan salinitas adalah ion-ion Na+, K+, Ca+2, Mg+2, dan Cl–. Satuan yang digunakan beragam seperti ppt (part per thousand), PSU (practical salinity unit = g/kg). Konduktivitas elektrik (EC) umum digunakan untuk menunjukkan tingkat sali-nitas dan dinyatakan dengan satuan dS/m (desi siemens/m; 1 dS/m = 1 mmhos/cm = 640 ppm atau mg/kg = 1000 μS/cm).
Pada daerah semi-arid, meningkatnya peng-gunaan air irigasi akibat peningkatan intensi-fikasi pertanian menyebabkan peningkatan salinitas tanah (Yeo 1998). Dari laporan FAO/ Unesco, di Afrika terdapat 69,5 juta ha lahan salin, di Asia, Amerika Latin, dan Australia masing-masing 19,5 juta ha, 59,4 juta ha, dan 84,7 juta ha (Brinkman 1980). Dengan semakin meningkatnya penggunaan air irigasi pompa, apalagi penggunaan pupuk yang cenderung meningkat maka di masa mendatang akan semakin banyak penumpukan garam di lahan pertanian.
Penilaian toleransi kedelai terhadap sali-nitas beragam tergantung parameter dan pembanding yang digunakan. Berdasarkan penurunan bobot kering tanaman dan kan-dungan N tanaman pada tingkat salinitas 50 dan 100 mM NaCl, kedelai tergolong toleran dibandingkan kacang tunggak, kacang hijau, dan faba-bean (Delgado et al. 1994). Berdasar-kan pengujian salinitas dari ECe 0,8 hingga 7,0 dS/m, kedelai termasuk mempunyai sensi-tivitas sedang dibandingkan dengan jagung, tomat, dan kentang (Katerji et al. 2000). Katerji et al. (2000) menggunakan istilah sensitivitas dan bukan toleransi. Biji kedelai tidak mampu berkecambah pada salinitas tanah (ECe) >7 dS/ m (Mindari 2009). Hasil kedelai tidak menga-lami penurunan pada salinitas 5 dS/m, tetapi pada salinitas 6,2, 7,5, dan 10 dS/m mengalami penurunan berturut-turut 25%, 50%, dan 100%. Berdasarkan penurunan hasil 10%, nilai kritis salinitas untuk kedelai adalah 1,3 dS/m (setara padi dan kacang tanah) (http://www. dpi. nsw.gov.au).
Pembentukan bintil akar dan fiksasi N turun pada salinitas 27 mM NaCl (Qifu dan Murray 1993). Peningkatan salinitas dari 0,8 dS/m menjadi 4,2 dS/m menurunkan total serapan
N sebesar 11% dan hasil sebesar 12%, pada peningkatan menjadi 7,0 dS/m menurunkan to-tal serapan N sebesar 44,7% dan hasil sebesar 46% (van Horn et al. 2001). Peningkatan salinitas dari 5 dS/m menjadi 10 dS/m menurunkan indeks luas daun, total klorofil kanopi daun, efisiensi penggunaan sinar mata-hari, akan tetapi indeks kandungan klorofil meningkat yang ditunjukkan juga oleh warna daun hijau gelap (Wang et al. 2001).
Varietas unggul kedelai mempunyai tingkat toleransi yang berbeda terhadap salinitas. Vari-etas Wilis, Malabar, dan Sindoro toleran ter-hadap salinitas hingga konsentrasi NaCl 70 mM (±6,4 dS/m), sedangkan varietas Lokon, dan Tidar mempunyai toleransi sedang (Yuniati 2004). Berdasarkan vigor benih pada konsen-trasi NaCl hingga 50 mM (±4,6 dS/m), varietas Orba tergolong toleran, Sinabung semi-toleran, dan Kawi tidak toleran (Muh. Farid 2006). Berdasarkan inisiasi tunas, varietas Wilis toleran hingga konsentrasi NaCl 8 g/L (±12,5 dS/m), varietas Kipas Putih, Jaya Wijaya, Tidar sudah terhambat pada konsentrasi NaCl 6 g/ L (±9,4 dS/m), dan varietas Lokon pada kon-sentrasi NaCl 2 g/L (±3,1 dS/m) (Lubis, http:/ /www.researchgate.net).
RESPONS TERHADAP KONDISI AIR TANAH
Kelebihan atau kekurangan air mempenga-ruhi pertumbuhan kedelai. Kelebihan air menyebabkan genangan dan menimbulkan cekaman aerasi, sedangkan kekurangan air menyebabkan cekaman kekeringan. Beberapa istilah yang banyak digunakan dalam kaitan dengan air tanah adalah kapasitas lapangan (field capacity), titik layu (wilting point), dan air tersedia (available water).
Tabel 1. Kandungan liat dan air tanah pada nilai pF 2,5 (kapasitas lapangan) dari beberapa jenis tanah.
Kandungan Kandungan air
Jenis tanah Asal liat (%) pada pF 2,5 (%) Pustaka
Ultisol Banten 34 37,8 Subowo et al. (2002)
53 39,2
Ultisol Subang 40 39,7 Pirngadi dan Pane (2004)
Entisol Kendalpayak, Malang 36 41,9 Sunardi et al. (2002)
Alfisol Muneng, Probolinggo 22 33,4 Sunardi et al. (2002)
Vertisol Ngale, Ngawi 81 83,0 Sunardi et al. (2002)
Entisol Pasuruan 42 37,2 Sunardi et al. (2002)
kapasitas lapangan. Makin tinggi kandungan oksida besi pada tanah-tanah yang berpe-lapukan lanjut juga meningkatkan air tersedia (Hidayat et al. 2002). Peningkatan kandungan C-organik sebesar 1% meningkatkan kandung-an air tersedia sebesar 2% (Olness dkandung-an Archer 2005). Kandungan liat dan air tanah pada nilai pF 2,5 dari berbagai jenis tanah disajikan dalam Tabel 1.
Air tersedia adalah kandungan air tanah antara kondisi kapasitas lapangan dan titik layu permanen (permanent wilting point). Kondisi titik layu permanen adalah kondisi air tanah dimana daun tanaman menunjukkan kondisi layu yang tidak dapat pulih kembali pada kondisi kelembaban udara jenuh, atau setara dengan kandungan air pada potensial – 15 bar atau nilai pF 4,2 (Landon 1984). Tidak semua air tersedia dapat diserap tanaman. Landon (1984) memperkirakan sekitar 66% dari total air tersedia yang dapat diserap tanaman.
Air dalam fase cair merupakan bagian pen-ting dalam siklus pertumbuhan tanaman. Sekitar 70–90% dari bobot tanaman merupakan zat cair, namun hanya sekitar 0,01% yang dibutuhkan dalam proses fotosintesis. Kedelai membutuhkan air pada fase pertumbuhan cepat sebesar 2,54–5,08 mm/hari, pada fase pembu-ngaan hingga polong isi penuh sebesar 5,08– 7,62 mm/hari, dan fase pemasakan polong hingga panen sebesar 1,27–5,08 mm/hari (http:/ /www.aragriculture. org). Secara umum, selama pertumbuhannya, (85–100 hari) kedelai mem-butuhkan air sebanyak 300–450 mm atau 2,5– 3,3 mm/hari.
Kebutuhan air pada fase generatif lebih tinggi dibandingkan pada fase vegetatif. Oleh karena itu, fase generatif lebih peka terhadap
kekeringan, terutama pada fase pembungaan hingga pengisian polong. Cekaman kekeringan selama fase generatif menurunkan hasil kedelai sebesar 34%–46%. Pada tanah bertekstur berat (seperti Vertisol) dan ringan (seperti Entisol), hasil kedelai yang tinggi dicapai pada kan-dungan air tanah masing-masing 70–85% dan 62–75% dari kapasitas lapangan (Suyamto et al. 1998). Tetapi hasil penelitian Syahbudin et al. (1998) menunjukkan bahwa serapan nitro-gen, fosfor, kalium dan hasil biji kedelai varietas Wilis pada Latosol tertinggi dicapai pada kan-dungan air tanah 125% dari air tersedia.
Kondisi air tanah saat tanam juga sangat penting diperhatikan karena menentukan perkecambahan biji. Rahmianna et al. (2000) melaporkan bahwa 81,3% benih kedelai berke-cambah pada kisaran potensial air –0,56 bar (pF 2,7) sampai –1,12 bar (pF 3,0). Dari 18,7% benih yang gagal berkecambah, sebanyak 10,9% karena busuk dan sisanya masih dalam kondisi utuh.
RESPONS TERHADAP UNSUR HARA
Nitrogen (N)
Nitrogen merupakan komponen utama penyusun protein, klorofil, enzim, hormon dan vitamin. Nitrogen diserap dalam bentuk ion NO3– dan NH4+ dan merupakan unsur yang
sangat mobil (mudah ditranslokasikan) dalam tanaman, dan oleh karena itu gejala kekahatan (kekurangan) N akan nampak pada daun tua. Gejala kekahatan N pada tanaman muda ditunjukkan oleh daun berwarna hijau pucat, dan pada kondisi kekahatan yang parah daun berwarna kuning pucat, batangnya lemah dan memanjang. Pada tanaman yang tua, daun-daun bagian bawah menunjukkan gejala kuning yang parah dan akhirnya gugur. Pertumbuhan tanaman kerdil, batang berwarna kemerahan, perkembangan polong terhambat, daun mengecil dan berdinding tebal sehingga daun menjadi kasar/keras dan berserat.
Kekahatan N umumnya terjadi pada tanah bertekstur pasir, tanah-tanah yang masam (pH rendah) dimana aktivitas mikroorganisme tanah terganggu. Tanaman kedelai mampu menfiksasi N setara dengan 46 kg N/ha. Sera-pan N pada kedelai dapat mencapai 280,9 kg N/ha di mana 141 kg N/ha berasal dari penam-batan N. Salvagiotti et al. (2008) menganalisis data dari 480 penelitian dari daerah subtropis dan tropis (termasuk Indonesia) dan menun-jukkan bahwa total serapan N pada kedelai sebesar 44–485 kg N/ha (rata-rata 219 kg N/ ha) dimana 52% dari hasil fiksasi. Secara umum, sekitar 50% dari N yang dibutuhkan tanaman berasal dari penambatan oleh
rizobium. Lahan yang pernah ditanami kedelai pada umumnya mempunyai populasi Rhizo-bium alami yang tinggi.
Tanah dengan kandungan N-total <0,1% N perlu dilakukan pemupukan N dosis 23–35 kg N/ha, terutama saat tanaman masih muda. Kandungan N sebesar 4,01–5,30% pada daun muda yang terbuka sempurna saat pemben-tukan polong dianggap cukup.
Fosfor (P)
Fosfor merupakan komponen utama penyusun nukleoprotein, asam nukleotida, fosfolipida, penyusun enzim yang berperan aktif dalam transportasi enerji. Fosfor juga penting dalam proses fosforilasi, fotosintesis, respirasi, sintesis, dan dekomposisi karbohidrat, protein, dan lemak. Unsur P sangat diperlukan untuk pembentukan biji. Fosfor diserap dalam bentuk ion H2PO4– dan bersifat mobil dalam tanaman.
Kekahatan P menurunkan nodulasi dan fiksasi N, meningkatkan karbohidrat, menurunkan kadar air tanaman. Kekahatan P biasanya mulai muncul pada minggu ke-4, tanaman terlihat kerdil, ukuran daun kecil. Daun tua berwarna hijau gelap kemudian dengan cepat berubah warna menjadi kuning dan gugur sebelum waktunya. Batang berubah warna menjadi ungu karena adanya akumulasi anto-sianin. Kekahatan P menghambat pemben-tukan bintil akar, perkembangan akar, polong dan biji.
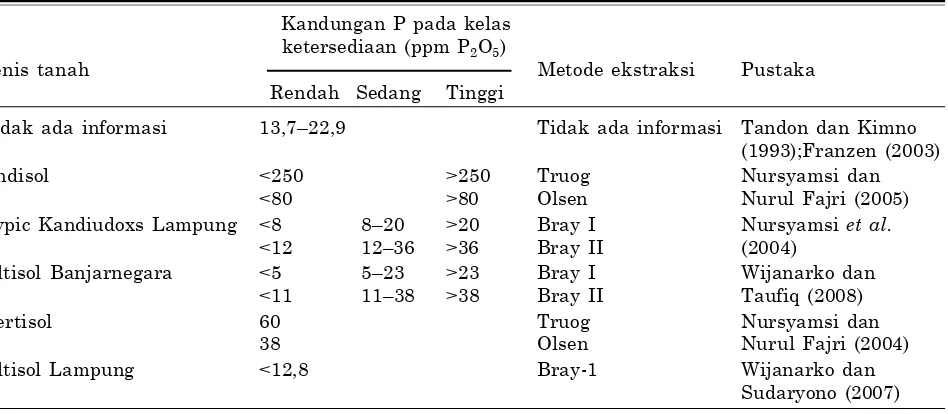
Kekahatan P umumnya terjadi pada Oxisol, Ultisol dan Inceptisol atau pada tanah masam yang mempunyai kandungan Fe, Al tinggi akibat adanya fiksasi. Kandungan P tersedia dalam klas ketersediaan yang sama beragam tergantung jenis tanah dan metode pengekstrak yang digunakan (Tabel 3).
KALIUM (K)
Kalium merupakan unsur penting dalam metabolisme protein, karbohidrat, lemak, dan transportasi karbohidrat dari daun ke akar. Kalium diserap dalam bentuk K+ dan bersifat mobil dalam tanaman. Gejala kekahatan K mulai nampak pada daun tua, yaitu timbulnya klorosis di antara tulang daun atau tepi daun. Pada kekahatan yang parah klorosis meluas hingga mendekati pangkal daun dan hanya meninggalkan warna hijau pada tulang daun, dan selanjutnya timbul gejala nekrosis. Tepi daun tua menguning, menggulung ke atas dan
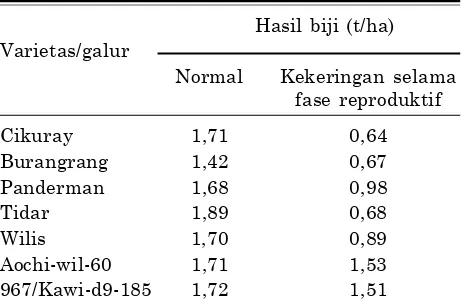
Tabel 2. Hasil beberapa varietas dan galur kedelai pada kondisi normal dan cekaman kekeringan.
Hasil biji (t/ha) Varietas/galur
Normal Kekeringan selama fase reproduktif
Cikuray 1,71 0,64
Burangrang 1,42 0,67
Panderman 1,68 0,98
Tidar 1,89 0,68
Wilis 1,70 0,89
Aochi-wil-60 1,71 1,53
967/Kawi-d9-185 1,72 1,51
pada akhirnya mengering. Batas kritis kan-dungan K-dd untuk kedelai adalah 0,2–0,3 me/ 100 g tanah (Franzen 2003).
Kekahatan K umum terjadi pada Oxisol, Ultisol dengan kejenuhan basa rendah atau pada tanah bertekstur pasir. Tanah yang mengandung K dapat ditukar (K-dd) 0,2–0,3 me/ 100 g perlu pemupukan K sebesar 22,5–45 kg K2O/ha. Kelas ketersediaan hara kalium pada Ultisol untuk kedelai berdasarkan pengekstrak HCl 25% adalah rendah (<340 ppm K2O), sedang (340–1150 ppm K2O) dan tinggi (>1150 ppm K2O). Kebutuhan pupuk untuk mencapai hasil maksimum adalah 210, 190, dan 150 kg KCl/ha, sedangkan untuk mencapai hasil opti-mum 85, 2, dan 0 kg KCl/ha masing-masing untuk K tanah rendah, sedang, dan tinggi (Nursyamsi 2006).
Kekahatan S, Ca, P menurunkan kandungan K tanaman, namun kekahatan N meningkatkan K dalam tanaman. Imbangan antara (Ca+Mg)/ K yang optimum untuk kedelai adalah 23–28 (Mascarenhas et al. 2004).
Kandungan K sebesar 1,71–2,50% dalam daun muda yang terbuka sempurna (young fully matured leaf, YFML) saat pembentukan polong dianggap cukup. Batas kritis kandungan K pada YFML pada fase R1 (berbunga) adalah 2,4% dan 2,6% masing-masing pada tanah yang diolah dan tanpa diolah (Yin dan Vyn 2005), pada fase R3–R4 (pembentukan polong) dan R5– R6 (pengisian polong) masing-masing adalah 0,88% dan 0,39% (Bell et al. 1987).
Kalsium (Ca)
Kalsium berperan penting dalam pengaturan air dalam tanaman. Kalsium diserap dalam bentuk ion Ca2+ dan mempunyai mobilitas rendah dalam tanaman, sehingga gejala ke-kahatan muncul pada daun muda atau titik tumbuh baik pada batang maupun akar. Meskipun demikian pada kekahatan yang parah, Ca dapat ditranslokasikan dari daun tua ke daun muda (Fegeria 2009). Kekahatan Ca ditandai dengan adanya bintik-bintik coklat atau hitam pada permukaan bawah daun, bila berlanjut terjadi nekrosis pada permukaan bawah maupun atas daun sehingga daun menjadi berwarna coklat, dan kadang daun nampak keriting mirip gejala serangan virus. Pada kondisi kekahatan yang akut ujung akar dan pucuk tanaman mati.
Kekahatan Ca umum terjadi pada tanah ber-tekstur pasir, Oxisol, Ultisol dengan pH masam, kejenuhan basa rendah dan Aluminium dapat ditukar (Al-dd) tinggi. Kandungan Ca-dd opti-mum untuk kedelai adalah 2,8 me/100 g (Fegeria 2009). Kandungan Ca 0,36–2,00% dalam daun muda yang terbuka sempurna saat pembentukan polong dianggap cukup.
Magnesium (Mg)
Magnesium adalah komponen penyusun klorofil daun dan karenanya sangat penting dalam proses fotosintesis. Dalam tanaman, Mg termasuk unsur yang agak mudah ditranslo-kasikan dari daun tua dan oleh karenanya
Tabel 3. Klas ketersediaan P tanah untuk kedelai pada berbagai jenis tanah dan metode ekstraksi.
Kandungan P pada kelas ketersediaan (ppm P2O5)
Jenis tanah Metode ekstraksi Pustaka
Rendah Sedang Tinggi
Tidak ada informasi 13,7–22,9 Tidak ada informasi Tandon dan Kimno (1993);Franzen (2003)
Andisol <250 >250 Truog Nursyamsi dan
<80 >80 Olsen Nurul Fajri (2005)
Typic Kandiudoxs Lampung <8 8–20 >20 Bray I Nursyamsi et al.
<12 12–36 >36 Bray II (2004)
Ultisol Banjarnegara <5 5–23 >23 Bray I Wijanarko dan
<11 11–38 >38 Bray II Taufiq (2008)
Vertisol 60 Truog Nursyamsi dan
38 Olsen Nurul Fajri (2004)
Ultisol Lampung <12,8 Bray-1 Wijanarko dan
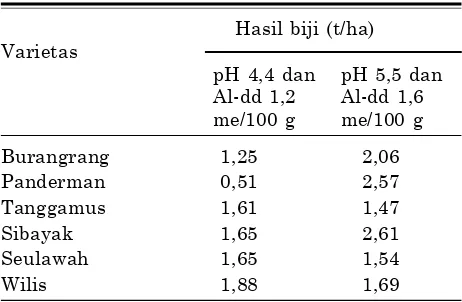
Tabel 5. Hasil beberapa varietas kedelai pada Ultisol Sumatera Selatan.
Hasil biji (t/ha) Varietas
pH 4,4 dan pH 5,5 dan Al-dd 1,2 Al-dd 1,6 me/100 g me/100 g
Burangrang 1,25 2,06
Panderman 0,51 2,57
Tanggamus 1,61 1,47
Sibayak 1,65 2,61
Seulawah 1,65 1,54
Wilis 1,88 1,69
Sumber: Purwantoro et al. (2007).
gejala awal kekahatan akan nampak pada daun-daun tua. Daun mengalami klorosis yang berawal dari tepi daun, kemudian menjalar ke bagian tengah di antara tulang daun. Keka-hatan yang meningkat menyebabkan terjadinya perubahan warna tepi daun menjadi merah kekuningan daun gugur, pertumbuhan ter-hambat dan hasil rendah.
Kekahatan Mg umum terjadi pada tanah bertekstur pasir, Oxisol, Ultisol dengan pH masam, kejenuhan basa rendah. Batas kritis kekahatan Mg-dd adalah 50 ppm Mg. Kan-dungan Mg-dd optimum untuk kedelai adalah 1,4 me/100 g (Fegeria 2009). Kisaran nilai cukup untuk kedelai pada daun muda adalah 0,26– 1,0%.
RESPONS TERHADAP LINGKUNGAN MASAM
Lahan kering masam umumnya berada pada daerah beriklim basah (curah hujan tahunan >2000 mm). Jenis tanah pada lahan kering ber-iklim basah didominasi oleh Ultisol dan Oxisol terutama tersebar di Kalimantan, Sumatera, Irian Jaya, sebagian Jawa dan Sulawesi.
Ciri utama lahan masam adalah pH tanah rendah (pH<6), dan umumnya kandungan Al tinggi. Kondisi pH tanah yang rendah dan Al tinggi tidak saja berpengaruh langsung hadap tanaman, tetapi juga berpengaruh ter-hadap ketersediaan hara bagi tanaman (Tabel 4). Ultisol dan Oxisol di Lampung mempunyai pH 4,3 dan kemasaman tertukar 57,2 cmol+/ kg (Setiawan 1997; Prasetyo dan Ritung 1998), mengandung Al monomerik 1,89 mM pada lapisan 0–15 cm dan 2,29 mM pada lapisan 15– 45 cm (Van der Heide et al. 1992), pH tanah 4,5–5 dengan kejenuhan Al 24,5–30,2% (Taufiq et al. 2004).
Adanya Al yang berlebihan dalam tanah masam menyebabkan pertumbuhan tanaman kedelai terganggu dan mengakibatkan rendah-nya hasil. Gejala awal keracunan nampak pada sistem parakaran, akar tumbuh tidak normal, percabangan akar tidak normal. Gejala pada daun adalah adanya bercak-bercak klorosis di antara tulang daun muda, tetapi tulang daun tetap hijau. Pada gejala yang parah tanaman kerdil dan daun berbentuk seprti mangkuk. Keracunan Al sering terjadi pada tanah masam dengan kejenuhan basa rendah. Aluminum menjadi tidak larut pada pH
≥
5,5 (Lindsay 1979).Kondisi pH tanah optimal adalah 5,5–7,5. Beberapa varietas kedelai seperti Wilis, Seu-lawah, dan Tanggamus toleran terhadap kondisi masam hingga pH 4,4 (Tabel 5).
Batas toleransi kedelai terhadap kejenuhan Al adalah 20%. Kandungan Al-dd dalam tanah sebesar 22 ppm atau sekitar 0,24 me Al/100 g termasuk tinggi. Beberapa varietas kedelai di Indonesia yang ada saat ini mempunyai batas kritis keracunan Al sekitar 1,33 me Al/100 g dalam larutan tanah. Varietas Wilis tergolong toleran terhadap kejenuhan Al tinggi. Berda-sarkan bobot kering akar, varietas Wilis tidak terpengaruh pada peningkatan kejenuhan Al dari 25% hingga 75%, sedangkan varietas Slamet, Sindoro, dan Dieng mengalami penu-runan 68% hingga 80% (Hanum et al. 2007).
Lahan masam juga dapat berupa lahan pasang surut dengan jenis tanah Histosol, Entisol, dan Inceptisol (Subagyo dan Widjaja-Adhi 1998). Masalah agro-fisik lahan rawa pasang surut terutama adalah lingkungan
Tabel 4. Unsur hara tanaman yang mungkin kahat dalam hubungannya dengan pH tanah.
Kisaran Unsur hara yang mungkin kahat pH tanah
4–5 Mo, Cu, Mg, B, Mn, S, N, P, K 5–6 Mo, Mg, S, N, P, K, Ca
6–7 Mg
8–9 Cu, B, Fe, Mn, N, Zn
9–10 Cu, Fe, Mn, Mg, Ca, Zn
perakaran yang jenuh air dan anaerobik (lum-pur, reduksi, kahat oksigen dan gas H2S),
adanya pirit atau bahan sulfidik, keracunan Al, Fe dan Mn, pH sangat masam dan kesuburan alami rendah (kahat P, N dan K serta miskin basa-basa) (Sudarsono 1999). Pada lahan-lahan yang piritnya sudah teroksidasi, masalah utama adalah keracunan Al, Mn, dan Fe karena pH tanah yang rendah, serta kahat unsur P.
Batas kritis Mn dengan pengekstrak DTPA untuk kedelai pada Ustochrepts adalah 3,3 mg/ kg tanah atau 3,3 ppm (Bansal dan Nayyar 1990). Batas kritis kekahatan Mn adalah 10
μg/g dan keracunan adalah 160 μg/g pada daun muda yang terbuka sempurna pada fase pertumbuhan R2 (Ohki 1981).
RESPONS TERHADAP INOKULASI RHIZOBIUM
Kedelai adalah tanaman Legume yang mem-bentuk bintil akar. Bintil akar termem-bentuk akibat adanya asosiasi antara akar tanaman dengan mikroba penambat N dari jenis Rhizobium. Bintil akar sesungguhnya berasal dari rambut akar tanaman yang terinfeksi oleh bakteri rhizobium dan kemudian mengalami perubahan bentuk. Jadi dalam satu bintil akar mengan-dung banyak bakteri rhizobium. Bakteri ini dapat hidup pada lingkungan oksigen (O2) terbatas (anaerobik) maupun pada kondisi kaya oksigen (aerobik), namun lebih menyukai kondisi yang aerobik. Waktu yang dibutuhkan dari terjadinya infeksi hingga terbentuknya bintil akar kecil sekitar 7 hari. Fiksasi N dimulai pada saat tanaman umur sekitar 25–30 hari.
Ciri bintil akar yang sehat dan tidak sehat:
1. Jika bintil akar dibelah, di dalamnya ber-warna merah atau pink menunjukkan tinggi-nya kandungan leghaemoglobin dan berarti aktif memfiksasi N. Semakin merah warna-nya berarti semakin aktif.
2. Bintil akar yang dalamnya berwarna putih atau abu-abu menunjukkan masih muda dan belum ada kegiatan menfiksasi N.
3. Bintil akar yang berwarna hijau menunjuk-kan tidak aktif menfiksasi N yang bisa disebabkan oleh rhizobium yang tidak efisien atau karena tanaman yang kurus.
Rhizobium dalam bintil akar dapat mengikat nitrogen (N2) dari udara dan kemudian
meng-ubahnya kedalam bentuk amonium (NH+4)
dengan bantuan enzim nitrogenase. Amonium
yang dihasilkan oleh rhizobium tersebut dapat dimanfaatkan oleh tanaman. Sebaliknya bakteri rhizobium mengambil karbohidrat, pro-tein, dan oksigen yang dihasilkan tanaman untuk hidup dan berkembang biak. Kebutuhan rhizobium akan oksigen dikontrol oleh protein yang mengandung unsur besi (Fe) yang disebut leghaemoglobin. Jumlah N yang difiksasi kede-lai yang tumbuh normal dapat mencapai rata-rata 219 kg N/ha, dan jumlah tersebut dapat memenuhi sekitar 50% dari kebutuhan tana-man kedelai.
Terbentuknya bintil akar dan efektivitas rhizobium dipengaruhi oleh beberapa faktor:
1. Kekeringan: terbentuknya bintil akar dan fiksasi N sangat terhambat oleh kondisi ke-keringan. Oleh karena itu kelembaban tanah harus dijaga optimal.
2. Kondisi tergenang: perkembangan rhizobium terhambat oleh adanya genangan sehingga fiksasi N juga terhambat.
3. Kondisi pH tanah yang rendah atau terlalu tinggi juga menghambat perkembangan rhizobium.
4. Pupuk N yang berlebihan akan menurunkan fiksasi N.
5. Aplikasi insektisida (yang diuji berbahan aktif carbofuran, thimet, dasanite, dan hep-tachlor) tidak berpengaruh terhadap kan-dungan leghaemoglobin, bintil akar.
6. Dipengaruhi oleh faktor tanah yang mempe-ngaruhi perakaran, misalnya kandungan Aluminium (Al) dan mangan (Mn).
7. Dipengaruhi oleh bakteri dan jamur anta-gonis terhadap rhizobium.
Secara alamiah, populasi Rhizobium dalam tanah yang pernah ditanami tanaman jenis le-gume cukup tinggi. Dari beberapa contoh tanah di lahan kering Alfisol di Jawa menunjukkan populasi Rhizobium yang cukup tinggi (Tabel 6). Populasi sel Rhizobium alami bervariasi tergantung jenis tanaman yang diusahakan, pola tanam, dan iklim. Lahan yang pernah dita-nami kedelai pada umumnya mempunyai popu-lasi Rhizobium alami lebih tinggi, namun ada pula yang populasinya tinggi meskipun belum pernah ditanami kedelai.
sangat lambat, terutama pada pH 4,5 atau lebih rendah.
Respons kedelai terhadap inokulasi rhizo-bium beragam. Pemberian Rhizorhizo-bium pada Ultisol Lampung dengan pH <4,5 meningkatkan bintil akar hingga 100% tetapi tidak mening-katkan hasil kedelai, dan pemberian pupuk 25 kg N/ha tidak meningkatkan hasil dan bahkan menekan pertumbuhan Rhizobium yang ditun-jukkan oleh menurunnya berat bintil akar (Sumadi 1985). Adisarwanto (1990) melaporkan bahwa inokulasi kedelai dengan Rhizogen dosis 2,5–5 g/kg benih meningkatkan hasil kedelai di lahan kering Alfisol Pasuruan, dan pening-katan dosis rhizogen menjadi >5 g/kg benih tidak meningkatkan hasil. Penggunaan Rhizo-plus dan Ginon di lahan kering Alfisol Probo-linggo dan Blitar pada beberapa varietas kedelai tidak meningkatkan hasil (Adie et al. 2000). Pemberian Rhizoplus dan pemupukan 25 kg N/ ha pada Ultisol Kalimantan Tengah tidak meningkatkan hasil kedelai (Asmarhansyah et al. 2001).
Sumadi (1987) melaporkan bahwa serapan N pada kedelai mencapai 280,9 kg N/ha dimana 141 kg N/ha atau 99% berasal dari fiksasi N. Salvagiotti et al. (2008) menganalisis data dari 480 penelitian dari daerah subtropis dan tropis (termasuk Indonesia) dan menunjukkan bahwa total serapan N pada kedelai sebesar 44–485 kg N/ha (rata-rata 219 kg N/ha) di mana 52% berasal dari hasil fiksasi oleh bakteri rhizobium.
Hasil-hasil penelitian ini menunjukkan bahwa penggunaan inokulan sangat tergantung dari pola tanam. Pada lahan yang sering
dita-nami kedelai, tidak diperlukan inokulasi untuk merangsang terbentuknya bintil akar.
KESIMPULAN
Lama penyinaran yang optimal adalah 10– 12 jam. Berkurangnya intensitas cahaya mata-hari menyebabkan tanaman tumbuh lebih tinggi, ruas antar buku lebih panjang, jumlah daun dan jumlah polong lebih sedikit, dan ukuran biji semakin kecil. Respons kedelai terhadap perubahan suhu tergantung pada fase pertumbuhan. Suhu yang sesuai pada fase perkecambahan adalah 15–22 oC, fase pembu-ngaan 20–25 oC, dan pada fase pemasakan 15– 22 oC. Peningkatan CO2 atmosfer dari 349 μL menjadi 700 μL meningkatkan laju pertukaran karbon (C), menurunkan laju transpirasi, dan meningkatkan efisiensi penggunaan air.
Faktor lingkungan di atas tanah dan di dalam tanah secara individu maupun inter-aksinya berpengaruh langsung maupun tidak langsung terhadap pertumbuhan tanaman. Respons tersebut dapat diketahui dari peru-bahan pertumbuhan, fenotipik tanaman, dan proses fisiologis. Lingkungan di atas tanah yang berpengaruh terhadap pertumbuhan tanaman kedelai terutama adalah lama dan intensitas penyinaran, suhu udara dan kandungan CO2
di atmosfer.
Kondisi lingkungan di dalam tanah yang berperan terhadap pertumbuhan kedelai ter-utama adalah tekstur tanah, kadar air tanah dan unsur hara, kemasaman tanah, unsur-unsur toksik, suhu tanah, dan salinitas. Kedelai tumbuh baik pada tanah bertekstur ringan
Tabel 6. Populasi Rhizobium endogen (alami) lahan kering Alfisol dari beberapa daerah di Jawa Timur.
Asal tanah Populasi Rhizobium Asal tanah Populasi Rhizobium
per gram tanah per gram tanah
Kalipare-Malang 2 x 103 Lamongan 8 x 101
Kademangan-Blitar 1,5 x 102 Gondosuli-Probolinggo 1,5 x 102
Besole-Tulungagung 6,5 x 101 Banyuglugur-Besuki 5,5 x 101
Watulimu-Trenggalek 3,75 x 102 Nguling-Pasuruan 2,3 x 103 Grujugan-Bondowoso 9 x 101 Panuran-Tuban 2,3 x 103
Kotakan-Situbondo 9 x 101 Ranuyoso-Lumajang 8 x 101
W.rejo-Banyuangi 4 x 103 Sumberjambe-Jember 8 x 101
Maron-Probolinggo 9 x 104 Kdungdung-Sampang 9 x 101
Galis -Bangkalan 2 x 103 Ketapang-Sampang 9 x 101
Burneh - Bangkalan 5,5 x 101
hingga berat, namun tanah yang padat (BI >1,38 kg/m3) tidak sesuai untuk kedelai. Kebu-tuhan air tanaman kedelai pada fase generatif lebih tinggi dibandingkan pada fase vegetatif, sehingga pada fase generatif lebih peka ter-hadap kekeringan terutama pada fase pembu-ngaan hingga pengisian polong. Kandungan air optimal adalah 70–85% dari kapasitas lapangan.
Kandungan unsur hara tanah harus di atas batas kekahatan agar tanaman tumbuh opti-mal. Nilai kritis suatu unsur hara dalam tanah beragam tergantung jenis tanah dan metode analisis yang digunakan. Pengaruh suhu tanah terutama pada fase perkecambahan, dan suhu tanah optimal adalah 24,2–32,8°C. Kedelai agak sensitif terhadap kemasaman tanah, unsur-unsur toksik, dan salinitas. Nilai kritis pH, Al, Mn, dan salinitas berturut-turut adalah pH 5,5, Al-dd 1,33 me/100 g, Mn 3,3 ppm, dan 1,3 dS/ m.
Rhizobium mempunyai peran penting dalam mendukung kebutuhan N tanaman kedelai, namun demikian inokulasi rizobium tidak diperlukan pada tanah yang sering ditanami kedelai.
DAFTAR PUSTAKA
Adisarwanto, T. 1990. Kajian inokulum Rhizogen dan PPC terhadap pertumbuhan dan hasil kedelai di lahan kering. Pen. Palawija 5(1):24–30.
Ariffin. 2008. Respons tanaman kedelai terhadap lama penyinaran. Agrivita 30(1): 61–66.
Asadi, B., D.M. Arsyad, H. Zahara, Darmijati. 1997. Pemuliaan kedelai untuk toleran naungan. Bul. Agrobio.1:15–20.
Asmarhansyah, M.A. Firmansyah, dan D.A. Suria-dikarta, 2001. Aplikasi pupuk Urea, SP-36, dan Rhizoplus pada kedelai di lahan kering masam Parenggean, Kalimantan Tengah. hlm. 89–93. Dalam D.M. Arsyad et al. (eds). Kinerja Teknologi untuk Meningkatkan Produktivitas Tanaman Ka-cang-kacangan dan Umbi-umbian. Puslitbangtan.
Bansal, R.L. and V.K. Nayyar. 1990. Critical manga-nese deficiency level for soybean grown in Ustochrepts. Fertilizer Res. 25:153–157.
Bell, R.W., D. Brady, D. Plaskett, and J. F. Lone-ragan. 1987. Diagnosis of potassium deficiency in soybean. J. of Plant Nut 10(9–16):1947–1953.
Beutler, A.N., J.F. Centurion1 and A.P. da Silva. 2005. Soil Resistance to Penetration and Least Limiting Water Range for Soybean Yield in a Haplustox from Brazil. Brazilian Archives of Biol and Tech 48(6): 863–871.
Botta, G.F., D. Jorajuria, R. Balbuena, and H. Rosatto. 2004. Mechanical and cropping behavior of direct drilled soil under different traffic inten-sities: effect on soybean (Glycine max L.) yields. Soil & Tillage Res. 78: 53–58.
Brinkman, R. 1980. Saline and sodic soils. In Land reclamation and water management, p. 62–68. In-ternational Institute for Land Reclamation and Improvement, Wageningen, The Netherlands.
Delgado, M.J., F. Ligero and C. Lluch. 1994. Effects of salt stress on growth and nitrogen fixation by pea, faba-bean, common bean and soybean plants. Soil Biol and Biochem 26(3):371–376.
Dornbos, D.L.Jr., and R.E.Mullen. 1991. Influence of stress during soybean seed fill on seed weight, germination, and seedling growth rate. J. of Plant Sci. 71: 373–383.
Fegeria, N.K. 2009. The Use of Nutrients in Crops Plants. CRC Press, Brazil. 430 P.
Franzen, D.W. Soybean Soil Fertility. http//www.ext. nodak.edu/extpubs/ plantsci/soilfert/sf1164w.htm. 24 Maret 2003.
Gibson, L.R. and R.E. Mullen. 1996. Influence of Day and Night Temperature on Soybean Seed Yield. Crop Sci 36: 98–104.
Hamim, Khairul Ashri, Miftahudin, dan Triadiati. 2008. Analisis status air, rolin dan aktivitas enzim antioksidan beberapa kedelai toleran dan peka kekeringan serta kedelai liar. Agrivita 30(3):201– 210.
Hanum, C, W.Q. Mugnisjah , S. Yahya, D. Sopandy, K. Idris, dan A. Sahar. 2007. Pertumbuhan Akar Kedelai pada Cekaman Aluminium, Kekeringan dan Cekaman Ganda Aluminium dan Keke-ringan. Agritrop 26(1):13–18.
Hidayat, A., S. Hardjowigeno, M. Soekardi dan S. Sabihan, 2002. Peranan oksida besi terhadap sifat tanah berpelapukan lanjut. J. Tanah dan Iklim 20:47–55.
Hillel, D. 1982. Introduction to Soil Physics. Academic Press, Inc., New York. 364 pages.
http://www.dpi.nsw.gov.au. Salinity tolerance in ir-rigated crops.
Huber,S.C., H.H. Rogers, dan F.L. Mowry. 1984. Ef-fects of Water Stress on Photosynthesis and Car-bon Partitioning in Soybean (Glycine max L. Merr.) Plants Grown in the Field at Different CO2 Levels. Plant Physiol 76:244–249.
ICAR, 1987. Fertilizer Use in Groundnut. Pub. and Information Dev. ICAR, New Delhi-India. Jenny, H. 1980. The Soil Resource: Origin and
Be-havior. Springer–Verlag, New York. 377 pages. Jones, P., L.H. Ellen, J.W.Jr. Jones, K.J. Boote, and
Kasai, M. 2008. Effect of growing soybean plants un-der continous light on leaf photosyntethic rate and other characteristics concerning biomass produc-tion. J of Agron 7(2):156–162.
Katerji, N., J.W. van Hoorn, A. Hamdy, and M. Mas-trorilli. 2000. Salt tolerance classification of crops according to soil salinity and to water stress day index. Agric Water Manag 43(1): 99–109. Kimball, B.A. 1983. Carbon dioxide and agricultural
yield; An assemblage and analysis of 430 prior observations. Agron J 75:779–788.
Koti, S., K.R. Reddy, V.G. Kakani, D. Zhao, V.R. Reddy. 2005. Interactive effects of carbon diox-ide, temperature and ultraviolet-B radiation on flower and pollen morphology, quantity and qual-ity of pollen in soybean (Glycine max L.) geno-types. J. Exp. Bot 56:725–736.
Landon, J.R. 1984. Booker Tropical Soil Manual: A handbook for soil survey and agricultural land evaluation in the tropics and subtropics. Longman Inc., New York. 450 p.
Lawn, R.J. and D.J.Hume. 1985. Response of tropi-cal and temperature soybean genotypes to tem-perature during early reproductive growth. Crop Sci. 25:137–142.
Lindsay, W.L. 1979. Chemical Equilibria in Soils. A Wiley-Interscience Publ. Toronto. 449 p.
Liu, X.J. Jian, W. Guanghua, and S.J. Herbert. 2008. Soybean yield physiology and development of high-yielding practices in Northeast China. Field Crops Res. 105:157–171.
Lobell, D.B. and G.P. Asner 2003. Climate and man-agement contributions to recent trens in US Agric. Yields. Sci 299, 1032.
Lubis, K. Respon Morfogenesis Embrio Beberapa Varietas Kedelai (Glycine max L. Merr) pada Berbagai Konsentrasi NaCl Secara in vitro. http:/ /www. researchgate.net.
Mapegau. 2006. Pengaruh cekaman air terhadap pertumbuhan dan hasil tanaman kedelai (Glycine max L. Merr). USU e-Journals Jurnal Kultura 41(1).
Mascarenhas, H.A.A., R.T. Tanaka, E.B. Wutke, N.R. Braga, and M.A.C. de Miranda. 2004. Potassium for soybeans. Better Crops 88(3):26–27.
Mindari, W., Maroeto, dan Syekhfani. 2009. Amelio-rasi Air salin menggunakan pupuk organik untuk meningkatkan produksi tanaman kedelai dan jagung dalam rotasi. Penelitian Hibah Bersaing DP2M Dikti TA. 2009.
Muchlis Adie, M. et al (eds). 2000. Laporan Tahunan Balitkabi Tahun 1999/2000.
Muh. Farid Bdr. 2009. Seleksi kedelai tahan kekeringan dan salinitas secara in vitro dengan NaCl. Agrivigor 6(1).
Nursyamsi, Ddan Nurul Fajri. 2005. Penelitian korelasi uji tanah hara phosphorus di tanah
Andisol untuk kedelai (Glycine max, L.). J Ilmu Tanah dan Lingkungan 5(2):27–37.
Nursyamsi, D. 2006. Kebutuhan hara kalium tanaman kedelai di tanah Ultisol.J Ilmu Tanah dan Lingkungan 6(2): 71–81.
Nursyamsi, D. dan Nurul Fajri. 2004. Metode ekstraksi dan batas kritis hara fosfor tanah Vertisol untuk kedelai (Glycine max, L.). Agric J Ilmu Pertanian No. 18.
Nursyamsi, D., M.T. Sutriadi, dan U. Kurnia. 2004. Metode ekstraksi dan kebutuhan pupuk P tanaman kedelai (Glycine max L.) pada tanah masam Typic Kandiudox di Papanrejo, Lampung. J Tanah dan Iklim No. 22.
Odum, E.P. 1983. Basic ecology. pp.368–443. Holt Sauders Inter. Eds. Japan.
Ohki, K. 1981. Manganese critical levels for soybean growth and physiological processes. J of Plant Nut 3(1–4):271–284.
Olness, A.E., and Archer, D.W. 2005. Effect of or-ganic carbon on available water in soil. Soil Sci 170(2):90–101. Morris, Minnesota </main/ site_main.htm? modecode=36–45-00-00>.
Peet, M. M. 1984. CO2 enrichment of soybean, effect of leaf/pod ration. Physiol. Plant. 60:38–42.
Pirngadi, K dan H. Pane, 2004. Pemberian bahan organik , kalium dan teknik persiapan lahan untuk padi gogorancah. Penel. Pert. Tan. Pangan. 23(3): 177–184.
Prasetyo, B.H. dan S. Ritung, 1998. Beberapa kendala pengembangan lahan kering di Indonesia. hlm. 267–275. Dalam Sudaryono dkk. (eds). Prosiding Seminar Nasional dan Pertemuan Tahunan Komisariat Daerah Himpunan Ilmu Tanah In-donesia Tahun 1998 (Buku 2).
Purwantoro, H. Kuswantoro, dan D.M. Arsyad. 2007. Keragaan beberapa galur kedelai di tanah Ultisol. Hlm 23–32. Dalam D. Harnowo et al. (peny.). Prosiding Seminar Peningkatan Produksi Kacang-kacangan dan Umbi-umbian Mendukung Kemandirian Pangan. Puslitbangtan, Bogor. 628 hlm.
Qifu, Ma and F. Murray. 1993. Effects of SO2 and salinity on nitrogenase activity, nitrogen concen-tration and growth of young soybean plants. Environ and Exp Bot 33(4):529–537.
Rahmianna, A.A., T. Adisarwanto, G. Kirchhof and H.B. So. 2000. Crop establishment of legumes in rainfed lowland rice-based cropping systems. Soil and Tillage Res 56(1–2): 67–82.
Rismaneswati. 2006. Pengaruh terracottem, kompos, dan mulsa jerami terhadap sifat fisik tanah, pertumbuhan dan hasil kedelai pada tanah Alfisol. J. Agrivigor 6(1):49–56.
Ni-trogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res 108 (1):1–13.
Setiawan, D. 1997. Keragaman susunan mineral liat beberapa tanah Sumatra Selatan (Buku II). Pros Kongres Nasional VI HITI. hlm. 33–40.
Siczek, A and J. Lipiec. 2011. Soybean nodulation and nitrogen fixation in response to soil surface straw mulching. Soil & Tillage Res 114:50–56.
Sionit, N., B.R.Strain, and E.P. Flint. 1987. Interac-tion of temperature and CO2 enrichment on soy-bean growth and dry matter partitioning. Can. J. Plant Sci. 67:59–67.
Sitompul, S.M. 2003. Potensi produksi dan pengem-bangan teknologi kedelai dan jagung dalam sistem agroforestri. Laporan Penelitian Hibah Bersaing. Univ Brawijaya.
Soedarjo, M., T. Adisarwanto, dan Sudaryono. 2001. Nodulasi dan pertumbuhan tanaman kedelai pada beberapa jenis tanah di lahan kering di Jawa Timur. Laporan Teknis Balitkabi. 9 hlm.
Strain, B.R. 1985. Physiological and ecological con-trols on carbon sequestering in ecosystems. Bio-geoch 1:219–232.
Subagyo, H dan IPG. Widjaja-Adhi. 1998. Peluang dan kendala pembangunan lahan rawa untuk pengembangan pertanian di Indonesia. Hlm. 13– 50. Dalam Kurnia, U. et al (eds). Pros Pertemuan pembahasan dan Komunikasi Hasil Penelitian Tanah dan Agroklimat. Pusat Penelitian Tanah dan Agroklimat. Badan Litbang Pertanian.
Subowo, I. Anas, G. Djajakirana, A. Abdurahman dan S. Hardjowigeno, 2002. Pemanfaatan cacing tanah untuk meningkatkan produktivitas Ultisol lahan kering. J. Tanah dan Iklim 20:35–46.
Sudarsono. 1999. Pemanfaatan dan pengembangan lahan rawa/pasang surut untuk pengembangan pangan. hlm. 81–94. Dalam Irsal Las, et al (eds). Prosiding Seminar Nasional Sumber Daya Lahan. Hasil Penelitian Tanah dan Agroklimat. Pusat Penelitian Tanah dan Agroklimat. Badan Litbang Pertanian.
Suhartina. 2007. Evaluasi galur harapan kedelai hitam toleran kekeringan dan berdaya hasil tinggi. hlm 153–161. Dalam D. Harnowo et al. (peny.). Prosiding Seminar Peningkatan Produksi Kacang-kacangan dan Umbi-umbian Mendukung Kemandirian Pangan. Puslitbangtan, Bogor. 628 hlm.
Sumadi, S. 1985. Tanggapan kedelai terhadap ino-kulasi Rhizobium dan pemupukan N. Pen. Perta-nian 5(3):137–140.
Sumadi, S. 1987. Estimation of fixed nitrogen by soy-bean root nodules of T 201 and Enrei cultivars. Pen. Pertanian 7(1):29–32.
Sunardi, A. Taufiq, Sutrisno, dan A. Winarto, 2002. Laporan Tahunan Balitkabi Tahun 2001. Balai
Penelitian Tanaman Kacang-kacangan dan Umbi-umbian, Malang. 104 hlm.
Susanto, G.W.A., and T. Sundari, 2010. Pengujian 15 genotipe kedelai pada kondisi intensitas cahaya 50% dan penilaian karakter tanaman berdasarkan fenotipnya J. Biologi Indonesia 6(3):459–471.
Suyamto, A.A. Rahmianna dan L. Sunaryo. 1998. Peningkatan efisiensi air pengairan. Hlm. 85– 95. Dalam. Sudaryono, M. Soedarjo, Y. Widodo, Suyamto, A.A. Rahmianna dan A. Taufiq. Pros Seminar Nasional dan Pertemuan Tahunan Komisariat Daerah Himpunan Ilmu Tanah donesia Tahun 1998. Himpunan Ilmu Tanah In-donesia. Komisariat Daerah Jawa Timur.
Syahbudin, H., Y. Apriana, N. Heryani, Darmijati S, dan I. Las, 1998. Serapan hara nitrogen, fosfor, kalium tanaman kedelai (Glycine max L.) di rumah kaca pada tiga taraf intensitas radiasi surya dan kadar air tanah Latosol. J. Tanah dan Iklim 16:20–28
Tandon, H.L.S., and I.J. Kimmo, 1993. Balance fer-tilizer use, Its practical importance and guidelines for agricultural in the Asia-Pasific Region. ESCAP/FAO/UNIDO, New York. 49 p.
Taufiq, A., H. Kuntyastuti dan A.G. Manshuri, 2004. Pemupukan dan ameliorasi lahan kering masam untuk peningkatan produktivitas kedelai. Hlm. 21–40. Dalam Prosiding Lokakarya Pengembangan Kedelai Melalui Pendekatan Pengelolaan Tanaman Terpadu di Lahan Masam. Balitkabi–BPTP Lampung. 72 hlm.
Teramura, A. H., J. H. Sulivan, and J. Lydon. 1990. Effect of UV-B radiation on soybean yield and seed quality. A six yiears field study. Physiol. Plant. 80:5–11.
Tyagi, S.K and R. P. Tripathi. 1983. Effect of tem-perature on soybean germination. Plant and Soil 74(2):273–280
Van der Heide, J., S. Setijono, M.S.Syekhfani, E.N. Flach, K. Hairiah, S.M. Sitompul, and M. van Moordwijk. 1992. Can low external input crop-ping systemon acid upland soilsin the humid trop-ics be sustainable? Agrivita 15:1–10.
van Hoorn, J.W., N. Katerji, A. Hamdy and M. Mastrorilli. 2001.Effect of salinity on yield and nitrogen uptake of four grain legumes and on bio-logical nitrogen contribution from the soil. Agric. Water Manag. 51(2): 87–98.
Wang, D., M.C. Shannon, and C.M. Grieve. 2001. Sa-linity reduces radiation absorption and use effi-ciency in soybean. Field Crops Res 69:267–277.
Wijanarko, A. dan Sudaryono, 2007. Uji kalibrasi P pada tanaman kedelai di tanah Ultisol Seputih Banyak Lampung Tengah. hlm. 233–242. Dalam D. Harnowo, et al. (Peny.). Pros. Sem. Pening-katan Produksi Kacang-kacangan dan Umbi-umbian Mendukung Kemandirian Pangan. Pus-litbangtan, Bogor. 628 hlm.
Yeo, A. 1998. Predicting the interaction between the effects of salinity and climate change on crop
plants. Scientia Horticulturae 78(1–4):159–174.
Yin, Xinhua and T.J. Vyn. 2005. Critical leaf potas-sium is higher in no-till soybeans. Better Crops 89(2):3–5.