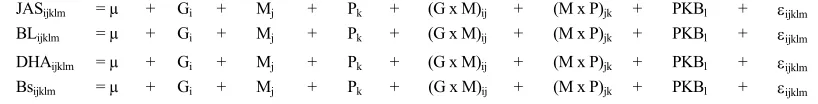ISMETH INOUNU1, BESS TIESNAMURTI1, SUBANDRIYO1, danHARIMURTI MARTOJO2
1
Balai Penelitian Ternak, P.O. Box 221, Bogor 16002, Indonesia
2
Program Pascasarjana, Institut Pertanian Bogor, Jalan Rasamala, Dramaga, Bogor 16680, Indonesia
(Diterima dewan redaksi 5 Oktober 1998)
ABSTRACT
INOUNU, ISMETH, BESS TIESNAMURTI, SUBANDRIYO, and HARIMURTI MARTOJO. 1999. Lamb production of prolific sheep. Jurnal Ilmu Ternak dan Veteriner 4(3): 148-160.
The research was aimed to study factors that affected litter size (JAS), lamb survival (DHA) and litter weight at birth (BL) and at weaning (BS) on prolific ewes. Production records during the year of 1981-1993 were used in this study (1,437 records). Ewe genotypes (G), management levels (M), ewe parity (P) were the fixed effect and ewe gestation gain (PKB) was a covariable that included in the model. Analysis of variance for unbalanced data with general linear model procedure were used. Mean JAS was 1.77. Sources of variation that affected JAS were G, M, P-M interaction, and PKB. One copy of FecF gene increased the number of lambs born by 0.8. Changing toward a better management levels was followed by an increase of JAS. An increased of one kilogram PKB was also followed by an increase of 0.04 litter size born. The mean of BL was 3.43 kg and the sources of variation that affected BL were G, M and P-M interaction. At all management levels, primipara ewes had lower litter size at birth and litter birth weight than the pluripara ewes. Mean DHA found was 73.93%, it was affected by G-M interaction, P and PKB. First parity ewes has 11% lower of DHA than second parity ewes. An increase of 1 kg PKB was followed by an increase of 2.3% of DHA. Sources of variation that affected BS were G-M interaction, P and PKB. Mean BS in this study was 13.12 kg.
Key words : Prolific sheep, lamb production
ABSTRAK
INOUNU, ISMETH, BESS TIESNAMURTI, SUBANDRIYO, dan HARIMURTI MARTOJO. 1999. Produksi anak pada domba prolifik. Jurnal Ilmu Ternak dan Veteriner 4(3): 148-160.
Penelitian dilakukan untuk mempelajari faktor-faktor yang mempengaruhi JAS, bobot lahir anak (BL), daya hidup (DHA) dan bobot sapih (BS) per induk pada domba prolifik. Data yang digunakan diambil dari catatan produksi induk (1.437 catatan) dari tahun 1981-1993. Peubah tetap yang diikutsertakan di dalam model adalah genotipe (G), manajemen (M), paritas (P), dan pertambahan bobot badan induk (PKB) sebagai covariable. Data dianalisis dengan sidik ragam untuk pengamatan yang tidak sama menggunakan prosedur general linear model. Rataan jumlah anak sekelahiran (JAS) yang didapatkan adalah 1,77 ekor per induk. Keragaman JAS dipengaruhi oleh genotipe, manajemen dan interaksi antara manajemen dengan paritas induk serta pertambahan bobot badan induk (PKB). Kehadiran satu duplikat gen FecJF dapat meningkatkan JAS sebanyak 0,8 ekor per induk. Perubahan manajemen ke tingkat yang lebih baik juga meningkatkan JAS. Peningkatan satu kg bobot badan induk (PKB) diikuti dengan meningkatnya JAS sebanyak 0,04 ekor. Rataan total bobot lahir anak per induk (BL) adalah 3,43 kg. Keragaman BL dipengaruhi oleh genotipe, manajemen dan interaksi manajemen dengan paritas induk. Pada semua tingkat manajemen, induk-induk primipara menghasilkan JAS dan BL lebih rendah dibandingkan induk-induk pluripara. Rataan daya hidup anak prasapih (DHA) didapatkan sebesar 73,93%, keragamannya dipengaruhi oleh interaksi genotipe dan manajemen, paritas induk dan pertambahan bobot badan induk. Pada paritas pertama, induk mempunyai DHA 11% lebih rendah dibandingkan pada paritas kedua. Kenaikan pertambahan bobot badan induk sebesar 1 kg meningkatkan DHA sebesar 2,3%. Keragaman total bobot badan anak saat sapih (BS) dipengaruhi oleh interaksi genotipe dan manajemen, paritas induk dan pertambahan bobot badan induk (PKB). Rataan BS yang didapat adalah 13,12 kg.
Kata kunci : Domba prolifik, produksi anak
PENDAHULUAN
Suatu populasi domba dapat dikelompokkan menjadi domba prolifik bila mempunyai rataan jumlah anak lahir yang banyak (≥ 1,75 ekor). Jumlah anak sekelahiran ditentukan oleh tingkat laju ovulasi dan
fertilisasi. Laju ovulasi pada domba Booroola Merino (PIPER et al., 1985) dan pada domba Jawa (BRADFORD
et al., 1991), dipengaruhi oleh gen tunggal,
hubungannya dengan tingkat manajemen yang diterapkan (INOUNU et al., 1997).
HOHENBOKEN dan CLARKE (1981) dan BRADFORD (1985) menyatakan bahwa peningkatan jumlah anak sekelahiran secara ekonomis menguntungkan dibandingkan induk yang menghasilkan satu ekor anak saja setiap kali beranak. BINDON et al. (1984) melaporkan bahwa kenaikan jumlah anak saat lahir 0,77 ekor per induk beranak, menghasilkan kelebihan keuntungan sebesar 24%. Hal ini disebabkan total bobot sapih induk yang beranak kembar lebih tinggi dibandingkan dengan induk yang beranak tunggal. Pada ternak-ternak dengan prolifikasi tinggi diperlukan manajemen yang memadai untuk dapat meningkatkan daya hidup anak. Sebaliknya, apabila tingkat manajemen yang sama diterapkan untuk domba non-prolifik, akan terjadi pemborosan yang menurunkan nilai laba.
INOUNU et al. (1993) melaporkan bahwa rataan bobot sapih dari induk yang melahirkan tunggal, kembar dua dan tiga adalah sebesar 10,5; 12,3 dan 13,1 kg per induk secara berturut-turut pada kondisi lingkungan yang normal. Namun, dengan perbaikan kondisi lingkungan (peningkatan kualitas dan kuantitas pakan), bobot sapih tersebut dapat ditingkatkan secara berturut-turut menjadi 15,2; 20,1 dan 24,4 kg.
Hasil-hasil di atas menunjukkan bahwa penanganan induk pra-sapih berdasarkan genotipe sangat penting karena erat hubungannya dengan nilai laba yang akan diterima. Dalam makalah ini dilaporkan hubungan antara tingkat manajemen yang diterapkan dengan produktivitas induk domba prolifik sampai pada periode sapih.
BAHAN DAN METODE
Lokasi penelitian
Penelitian dilakukan di Stasiun Pemuliaan Ternak, Cicadas, Gunung Puteri, 28 km dari Bogor, dengan rataan suhu udara 33oC dan rataan curah hujan 3.112
mm per tahun. Sejak bulan Agustus 1990, lokasi ternak dipindahkan ke Stasiun Penelitian Ternak Bogor, dengan rataan suhu udara 25oC dan rataan curah hujan 4.230 mm per tahun. Pada lokasi Bogor, fasilitas yang tersedia relatif sama dengan lokasi Cicadas.
Ternak
Sejak tahun 1983, perkawinan diarahkan untuk pembentukan domba galur prolifik, dengan menggunakan pejantan-pejantan dari Garut. BRADFORD
et al. (1991) mengklasifikasikan ternak-ternak ini ke
dalam kelompok genotipe berdasarkan kriteria-kriteria sebagai berikut :
FecJ+FecJ+ (prolifikasi rendah): induk-induk yang
tidak pernah menghasilkan corpus luteum (CL) atau jumlah anak sekelahiran (JAS) > 2. Pada jumlah pengamatan ≥ 3 kali, rataan CL atau JAS ≤ 1,7.
FecJFFecJ+ (prolifikasi medium): induk-induk yang minimal mempunyai satu kali pengamatan menghasilkan CL atau JAS=3, atau mempunyai frekuensi catatan CL atau JAS=2 yang tinggi. Pada jumlah pengamatan ≥ 3 kali, rataan CL atau JAS > 1,7.
FecJFFecJF (prolifikasi tinggi): induk-induk yang minimal mempunyai satu kali pengamatan menghasilkan CL atau JAS=4.
Pencatatan jumlah CL dilakukan dengan teknik laparoskopi secara periodik pada hari ke-3 - 10 setelah induk menampakkan berahi. Berdasarkan kriteria tersebut di atas ternak dikelompokkan ke dalam masing-masing kelompok genotipe, untuk kemudian dikawinkan dalam masing-masing kelompoknya.
Tatalaksana pemeliharaan
Di lokasi Stasiun Pemuliaan Ternak di Cicadas, 1981-1989, ternak diberi pakan konsentrat sebanyak 300 g/ekor/hari, dengan kualitas yang berbeda-beda (kisaran protein kasar 10-13%). Namun pada saat "sulit" ternak tidak mendapatkan konsentrat. Hijauan rumput Gajah diberikan sebanyak 2-3 kg/ekor/hari. Peningkatan jumlah pakan penguat menjadi 400-500 g/ekor/hari dilakukan pada saat mengawinkan domba betina, dan saat akan beranak sampai masa penyapihan (90 hari setelah kelahiran). Secara umum kondisi ketersediaan pakan di Stasiun Percobaan Cicadas dapat dibagi menjadi dua kelompok, yaitu kelompok kondisi ketersediaan pakan baik (tahun 1981, 1984 1987, dan 1989), sedangkan data tahun 1990 tidak diikutsertakan dalam analisis karena pada saat itu terjadi perpindahan ternak bunting ke lokasi Bogor. Kondisi ketersediaan pakan buruk (tahun 1982, 1985, 1986, dan 1988), selanjutnya kondisi ini secara berturut-turut disebut sebagai manajemen sedang (MNJ-2) dan manajemen buruk (MNJ-1).
dari jumlah sebelumnya, berdasarkan asumsi induk akan tumbuh paling tidak sebanyak 5 kg sampai dengan beranak.
Perkawinan ternak
Sebelum tahun 1983 perkawinan ternak dilakukan secara kontinyu, untuk mendapatkan selang beranak 8 bulan. Mulai tahun 1983, untuk memudahkan pengumpulan data, dilakukan penyerentakan berahi dengan menggunakan spons intra vaginal Chronogest atau Repromap, yang dimasukkan ke dalam vagina dan dibiarkan selama 14 hari. Pemeriksaan berahi dimulai pada hari pencabutan spons. Ternak betina yang menunjukkan berahi dapat diketahui dengan mempergunakan pejantan vasektomi. Setelah berahi terdeteksi, ternak dikawinkan dengan pejantan yang telah ditetapkan. Setelah dilakukan perkawinan secara individual selama dua minggu, pejantan dibiarkan di dalam masing-masing kandang selama dua minggu lagi untuk memberi kesempatan kawin bagi ternak-ternak yang kembali berahi ataupun yang belum kawin. Ternak betina dijaga agar tetap berada di dalam kelompok yang sama seperti pada saat perkawinan sampai beranak dan menyapih anaknya, agar
identifikasi nomor pejantan tidak tertukar. Pada saat perkawinan, dilakukan pencatatan nomor domba betina, nomor pejantan, tanggal kawin, dan bobot kawin (BK).
Pada saat kelahiran, induk dan anaknya dimasukkan ke dalam kandang sekat berukuran 1x1m2,
dan dibiarkan selama satu sampai tiga hari agar induk dan anak dapat saling mengenal.
Data
Peubah-peubah yang dicatat pada saat kelahiran adalah jumlah anak sekelahiran (JAS), total bobot anak saat lahir (BL), kemudian dicatat pula daya hidup anak (DHA) sampai dengan umur sapih (90 hari) dengan cara membandingkan jumlah anak yang hidup sampai saat disapih dengan jumlah anak pada saat kelahiran; total bobot anak saat sapih (BS) dan juga diamati pertambahan bobot badan induk selama kebuntingan (PKB) dengan cara mengurangi bobot badan induk saat beranak (BB) dengan bobot induk saat kawin (BK).
Data dianalisis dengan sidik ragam untuk pengamatan yang tidak sama, menggunakan prosedur
general linear model paket SAS (1987). Model
matematik peubah yang diamati serta faktor yang mempengaruhinya adalah sebagai berikut:
JASijklm = µ + Gi + Mj + Pk + (G x M)ij + (M x P)jk + PKBl + εijklm BLijklm = µ + Gi + Mj + Pk + (G x M)ij + (M x P)jk + PKBl + εijklm
DHAijklm = µ + Gi + Mj + Pk + (G x M)ij + (M x P)jk + PKBl + εijklm Bsijklm = µ + Gi + Mj + Pk + (G x M)ij + (M x P)jk + PKBl + εijklm
Keterangan: µ=rataan umum; G=Genotipe: FecJ+FecJ+=11; FecJFFecJ+=22; FecJFFecJF=33; M=Manajemen: rendah=1; sedang=2; tinggi=3; P=Paritas (1, 2, 3, 4, dan ≥ 5); PKB=Pertambahan bobot badan induk (kawin-beranak)
HASIL DAN PEMBAHASAN
Jumlah anak sekelahiran
Gambar 1 memperlihatkan fluktuasi jumlah anak sekelahiran (JAS) dari masing-masing genotipe dari tahun ke tahun. JAS dari ternak dengan genotipe FecJFFecJF tampak lebih bervariasi dibandingkan kedua
genotipe lainnya. Hal ini membuktikan bahwa jumlah anak lahir pada induk-induk prolifik erat kaitannya dengan lingkungan (pakan, manajemen, musim kering yang panjang). FAHMY (1989) melaporkan adanya pengaruh musim pada saat perkawinan terhadap JAS pada domba Romanov.
Rataan JAS pada penelitian ini adalah 1,77 ekor per induk (N=1.437; SD=0,64; CV=35,99%). Distribusi JAS dari penelitian ini adalah 44,5% tunggal, 37,6% kembar dua, 14,5% triplet, 3,1% kembar empat, dan
0,3% anak kembar ≥ lima. Sifat beranak banyak ini juga dilaporkan oleh MAIJALA (1996) yang menyatakan bahwa pada domba Finnsheep dijumpai induk-induk yang beranak 9 ekor per kelahiran, induk-induk dengan 8 ekor anak sering dijumpai dan induk-induk dengan 7 ekor anak selalu terjadi setiap tahun.
Jumlah anak kembar pada ternak dengan genotipe FecJ+FecJ+ adalah 17%, sedangkan anak tunggal adalah
23, dan 16% masing-masing untuk genotipe FecJFFecJ+ dan FecJFFecJF (Gambar 2). Hasil ini lebih memperkuat
menunjukkan adanya variasi JAS yang besar pada kedua bangsa domba ini.
Gambar 1. Rataan jumlah anak sekelahiran (JAS) pada masing-masing genotipe dari tahun 1981-1993
Gambar 2. Distribusi (%) jumlah anak sekelahiran berdasarkan genotipe
Sumber keragaman yang mempengaruhi nilai JAS (Tabel 1) adalah genotipe (P<0,01), manajemen (P<0,05), paritas induk (P<0,05) dan pertambahan bobot induk (P<0,01). Pada domba Booroola Merino, yang telah terlebih dahulu dinyatakan produktivitasnya dipengaruhi oleh gen prolifik dengan simbol gen FecBF,
rataan JAS pada masing-masing genotipenya lebih tinggi dibandingkan JAS dari domba pada penelitian ini, yaitu masing-masing 1,48; 2,38; dan 2,74 untuk genotipe FecB+FecB+, FecBFFecB+ dan FecBFFecBF secara berturut-turut (PIPER et al., 1985).
Seperti sudah diduga sejak semula terdapat perbedaan JAS antara ternak-ternak non-karier dengan ternak karier gen FecJF. Secara keseluruhan, satu duplikat gen FecJF, terlihat meningkatkan jumlah anak
sekelahiran sebanyak 0,80 ekor. PIPER et al. (1985) mendapatkan pada domba Booroola Merino satu duplikat pertama gen FecBF meningkatkan JAS sebanyak 0,90 ekor, sedangkan untuk satu duplikat yang kedua sebanyak 0,40 ekor per induk.
Secara keseluruhan perbaikan manajemen menyebabkan terjadinya kenaikan JAS, terutama dari MNJ-1 ke MNJ-2 terjadi peningkatan JAS sebesar 4,3%. Tidak demikian halnya apabila peningkatan dilakukan dari MNJ-2 ke MNJ-3. LAND (1977), GUNN dan DONEY (1979), GUNN (1983), PIPER et al. (1985) , dan BRADFORD dan INOUNU (1996) melaporkan bahwa JAS dipengaruhi oleh faktor genetik dan faktor lingkungan serta interaksinya. COOP (1966) melaporkan adanya pengaruh tingkat pakan terhadap JAS, serta hubungan yang erat antara JAS dengan bobot badan induk. HOHENBOKEN dan CLARKE (1981) melaporkan adanya peningkatan JAS pada domba keturunan Finnsheep yang dipelihara di daerah beririgasi, yang mencerminkan ketersediaan pakan yang kontinyu, dibandingkan dengan betina yang dipelihara di daerah pegunungan yang ketersediaan hijauannya sangat tergantung pada musim. Sebaliknya, pada domba keturunan Cheviot di daerah yang beririgasi dihasilkan JAS yang lebih rendah dibandingkan dengan JAS dari betina Cheviot yang dipelihara di daerah pegunungan. Selanjutnya, REESE et al. (1990) melaporkan adanya kenaikan JAS dengan peningkatan energi pada pakan tambahan yang diberikan, JAS meningkat dari 1,27 menjadi 1,49 pada domba Sumatera yang beranak pertama kali. Hasil yang berlawanan dilaporkan oleh MAIJALA (1996) bahwa tidak ada perbedaan JAS antara induk-induk Finnsheep yang dipelihara di negara-negara lain dengan yang dipelihara di Finlandia, hal ini menunjukkan bahwa pada domba Finnsheep JAS yang tinggi tidak dipengaruhi oleh kondisi lingkungan di mana domba tersebut dipelihara.
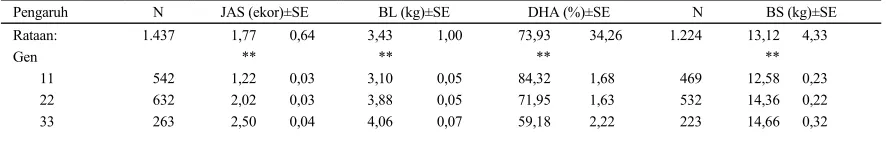
Tabel 1. Rataan kuadrat terkecil dan salah baku (SE) untuk jumlah anak sekelahiran (JAS), total bobot lahir (BL), daya hidup anak (DHA) dan total bobot sapih (BS)
Pengaruh N JAS (ekor)±SE BL (kg)±SE DHA (%)±SE N BS (kg)±SE
Rataan: 1.437 1,77 0,64 3,43 1,00 73,93 34,26 1.224 13,12 4,33
Gen ** ** ** **
MNJ: * ** ** **
1 446 1,83 0,04 3,48 0,06 62,62 2,17 337 10,08 0,32
2 705 1,91 0,02 3,52 0,04 74,70 1,50 621 13,12 0,20
3 286 1,99 0,04 4,04 0,07 78,12 2,30 226 18,40 0,30
Paritas: * * * **
1 419 1,79 0,04 3,06 0,06 66,84 1,89 334 12,28 0,26
2 409 1,91 0,03 3,63 0,05 75,46 1,86 369 13,87 0,25
3 269 1,93 0,05 3,74 0,07 72,03 2,46 229 14,12 0,34
4 191 1,99 0,05 3,99 0,08 72,08 2,72 165 15,43 0,38
5 149 1,94 0,06 3,97 0,09 72,67 3,08 127 13,63 0,41
Gen x MNJ:
n.s. n.s. P<0,06 **
11 1 174 1,19 0,05 2,97 0,08 79,78 2,79 138 9,92 0,40 11 2 272 1,19 0,04 2,90 0,06 86,56 2,18 242 11,50 0,29 11 3 96 1,28 0,07 3,42 0,11 86,61 3,64 89 16,32 0,48 22 1 213 1,91 0,07 3,60 0,08 64,36 2,70 162 10,12 0,39 22 2 312 1,98 0,04 3,72 0,06 75,63 2,02 273 13,52 0,27 22 3 107 2,16 0,07 4,31 0,11 75,87 3,62 97 19,43 0,48 33 1 59 2,41 0,09 3,86 0,13 43,73 4,57 37 10,20 0,72 33 2 121 2,55 0,06 3,92 0,09 61,91 3,15 106 14,33 0,42 33 3 83 2,54 0,07 3,92 0,11 71,89 3,89 80 19,44 0,50
MNJ x Paritas: * ** n.s. *
1 1 179 1,71 0,05 2,91 0,08 53,92 2,86 124 8,62 0,45 1 2 124 1,93 0,06 3,65 0,09 63,82 3,23 105 9,82 0,46 1 3 47 1,90 0,10 3,55 0,15 64,10 5,14 36 10,34 0,74 1 4 40 1,76 0,10 3,52 0,16 70,59 5,54 31 11,37 0,79 1 5 56 1,86 0,09 3,77 0,14 60,69 4,66 41 10,25 0,69 2 1 165 1,85 0,05 3,15 0,08 70,67 2,72 141 12,27 0,37 2 2 203 1,87 0,05 3,33 0,07 79,20 2,50 186 12,89 0,33 2 3 167 1,85 0,05 3,37 0,08 73,52 2,74 142 13,80 0,37 2 4 103 1,92 0,06 3,77 0,10 76,37 3,44 92 13,80 0,46 2 5 67 2,05 0,08 3,95 0,12 73,74 4,22 60 12,85 0,57 3 1 75 1,81 0,08 3,12 0,12 75,92 4,09 69 15,94 0,54 3 2 82 1,93 0,07 3,91 0,11 83,38 3,87 78 18,90 0,50 3 3 55 2,04 0,09 4,29 0,14 78,46 4,67 51 18,23 0,61 3 4 48 2,28 0,09 4,67 0,15 69,27 4,99 42 21,11 0,68 3 5 26 1,91 0,13 4,18 0,20 83,59 6,82 26 17,79 0,86
PKB: -0,04** -0,03n.s. 2,32** 0,185**
Keterangan : Gen=Genotipe: 11=FecJ+
FecJ+
; 22=FecJF
FecJ+
; 33=FecJF
FecJF
; MNJ=Manajemen: 1=rendah; 2=sedang; 3=tinggi;
PKB=Pertambahan bobot badan Kawin-Beranak; **=P<0,01; *=P<0,05; n.s.=P>0,05; N=jumlah pengamatan untuk JAS, BL dan DHA;
n=jumlah pengamatan untuk BS
Angka-angka pada Tabel 1 memperlihatkan kenaikan JAS dengan bertambah dewasanya induk. Ternak yang beranak untuk kedua kalinya mempunyai jumlah anak sekelahiran 6,3% lebih tinggi dibandingkan dengan ternak-ternak primipara. Pada MNJ-1, JAS tertinggi dicapai pada paritas kedua, pada MNJ-2 pada paritas keempat, dan pada MNJ-3 pada paritas ketiga dan baru menurun pada paritas kelima. FAHMY (1989) dan INIGUEZ et al. (1991) melaporkan adanya pengaruh paritas terhadap JAS. Sementara itu, REESE et al. (1990) melaporkan adanya hubungan
tinggi pada domba persilangan Finnsheep dan Booroola Merino. Pada domba persilangan tersebut kenaikan JAS pada paritas kedua adalah sebesar 22,8%. FAHMY (1996) dalam studi pustakanya melaporkan penelitian MARIN dan PEYRAND (1975) pada domba Romanov dari 6 kelompok ternak di Perancis yang meliputi 1.500 kelahiran, didapatkan bahwa JAS meningkat dari 2,87 ekor pada induk yang berumur 2 tahun hingga mencapai 3,32 ekor pada induk-induk yang berumur 4 tahun, tetapi hanya 2,07 ekor pada induk-induk yang dikawinkan pada umur 7-8 bulan.Meningkatnya satu kg PKB diiringi dengan menurunnya jumlah anak sekelahiran sebanyak 0,04 ekor (P<0,01). Hal ini menunjukkan pada induk-induk yang beranak banyak, seluruh zat makanan yang didapat, digunakan untuk merawat kebuntingan, sehingga peningkatan bobot badannya lebih rendah dibandingkan betina-betina yang beranak lebih sedikit jumlahnya. Kematian embrio yang tinggi dapat pula terjadi pada betina yang telah dideteksi menghasilkan LO tinggi pada saat perkawinan dan terlanjur diberi pakan yang lebih tinggi dibandingkan betina yang menghasilkan LO rendah, sehingga zat-zat makanan yang seharusnya digunakan untuk merawat sejumlah besar anak tidak termanfaatkan dan tertimbun mengakibatkan bobot badan induk menjadi meningkat. Hasil sebaliknya, pada domba Sumatera dilaporkan oleh REESE et al. (1990), didapatkan bahwa pada kelompok betina yang mempunyai PKB <70 g/hari dihasilkan kelahiran anak kembar sebanyak 20%, sedangkan pada kelompok betina yang mempunyai PKB antara 70-85 g/hari dihasilkan kelahiran anak kembar sebanyak 60-75%. Namun perlu diingat bahwa domba pada penelitian tersebut mempunyai rataan JAS yang lebih rendah dibandingkan pada penelitian ini.
Bobot lahir total per induk
Gambar 3 memperlihatkan fluktuasi rataan bobot lahir total (BL) per induk dari masing-masing genotipe dari tahun ke tahun. Secara umum, BL dari induk dengan genotipe FecJFFecJF lebih berfluktuasi dan lebih tinggi dibandingkan dengan genotipe lainnya, bahkan pada tahun-tahun tertentu grafik BL untuk ternak dengan genotipe FecJFFecJF berhimpit dengan grafik BL ternak dengan genotipe FecJFFecJ+.
Gambar 3. Rataan total bobot lahir per induk (BL) pada masing-masing genotipe dari tahun 1981-1993
Rataan BL pada penelitian ini adalah 3,43 kg per induk per kelahiran (N=1.437; SD=1,00; CV =29,21%). INIGUEZ et al. (1991) melaporkan pada domba lokal Sumatera rataan BL per induk per kelahiran sebesar 2,74 kg. Rendahnya BL pada domba Sumatera erat hubungannya dengan bobot induk yang rendah pula (22,2 kg), sedangkan pada penelitian ini bobot badan induk rata-rata pada saat beranak adalah 26,4 kg. HAFEZ (1969) melaporkan bahwa induk yang berukuran tubuh kecil akan membatasi ukuran fetus agar proses kelahiran menjadi mudah, sebaliknya induk-induk yang berukuran besar akan memberikan pengaruh yang menguntungkan pada anaknya, setidaknya pada ukuran tubuh anak pada saat dilahirkan. DONALD dan RUSSELL (1970) melaporkan bahwa BL berhubungan erat dengan bobot induk saat kawin, induk dengan bobot kawin sebesar 25 kg mempunyai anak dengan BL sebesar 9,6% dan 15,2% dari bobot kawin induk untuk anak tunggal dan kembar dua, sedangkan induk dengan bobot kawin 100kg mempunyai anak dengan BL sebesar 6,5% dan 10,6% dari bobot kawin induk untuk anak tunggal dan kembar dua secara berturut-turut. Selanjutnya, INOUNU et al. (1995) melaporkan bahwa dengan menyilangkan domba Garut betina dengan pejantan St. Croix didapatkan bobot lahir yang sama antara keturunan domba Garut murni dengan hasil persilangannya, namun hasil persilangan antara domba St. Croix dengan betina lokal Sumatera masih lebih kecil dibandingkan hasil persilangan antara St. Croix dengan Garut. Ukuran tubuh domba betina Sumatera lebih kecil dibandingkan dengan ukuran tubuh domba Garut. Hasil ini menunjukkan bahwa ukuran tubuh induk merupakan faktor pembatas dari BL. Hasil yang sejalan dilaporkan oleh HUNTER (1956), yang melakukan perkawinan silang resiprokal antara domba Border Leicester yang berukuran tubuh besar dengan domba Welsh yang berukuran tubuh kecil. Anak dari induk bertubuh besar, lebih besar dibandingkan dari induk yang bertubuh kecil. Pejantan kecil mempunyai sedikit pengaruh pada ukuran tubuh anak dari induk besar dibandingkan dengan anak dari pejantan besar dengan induk kecil.
sedangkan pertambahan bobot induk (PKB) tidak mempengaruhi BL (P>0,05). SMEATON et al. (1983) melaporkan adanya pengaruh tingkat pakan dengan bobot lahir, selanjutnya SITORUS dan INOUNU (1983) melaporkan adanya pengaruh umur induk terhadap bobot lahir. Angka-angka pada Tabel 1 menunjukkan adanya peningkatan BL, satu duplikat gen FecJF
meningkatkan BL sebesar 0,78 kg.
Selanjutnya, perbedaan yang terjadi terutama antara genotipe FecJ+FecJ+ dengan genotipe FecJFFecJ+ maupun dengan genotipe FecJFFecJF dapat dikatakan
disebabkan oleh meningkatnya jumlah anak yang dilahirkan. INOUNU et al. (1994) melaporkan adanya kenaikan bobot lahir per induk dengan meningkatnya jumlah anak lahir, masing-masing 2,6; 3,2; 4,5; dan 5,5 kg untuk induk-induk yang beranak tunggal, kembar dua, tiga, dan empat secara berturut-turut. PRIYANTO et al. (1992) melaporkan ternak-ternak di desa-desa di daerah Cirebon dengan jumlah anak sekelahiran 1, 2, dan 3 secara berturut-turut mempunyai BL sebesar 2,6; 3,3; dan 6,0 kg, bila dikandangkan dan bila digembalakan didapatkan BL sebesar 2,1 dan 3,0 untuk ternak dengan jumlah anak sekelahiran 1 dan 2 secara berturut-turut.
Perbaikan manajemen baru terlihat meningkatkan BL pada MNJ-3 sebanyak 0,56 kg dibandingkan pada MNJ-1 (Tabel 1). HAFEZ (1969) menyatakan bahwa pada keadaan induk sangat kekurangan gizi
(undernourishment) fetus akan terus tumbuh dan akan
mencapai bobot lahir yang mendekati normal. Namun pada kondisi tertentu dijumpai pula bobot fetus yang proporsional dengan jumlah kalori yang dikonsumsi induk. Itulah sebabnya pada penelitian ini antara MNJ-1 dan MNJ-2 tidak terdapat perbedaan BL pada masing-masing genotipe, akan tetapi pada MNJ-3 perbedaannya menjadi jelas terlihat. Hasil yang sejalan dilaporkan oleh PRIYANTO et al. (1992) bahwa BL pada ternak domba yang dikandangkan di daerah Cirebon lebih tinggi dibandingkan dengan ternak yang digembalakan, karena kondisi padang pengembalaan yang lebih miskin pakan dibandingkan dengan induk-induk yang dikandangkan di samping mendapat pakan hijauan, peternak biasa pula memberikan pakan tambahan.
Tingkat paritas induk menunjukkan kedewasaan seekor induk. Hal ini terlihat dari kian meningkatnya BL dengan bertambahnya pengalaman ternak dalam beranak (Tabel 1). Ternak yang beranak untuk kedua kalinya menghasilkan BL yang lebih berat dibandingkan ternak-ternak yang beranak pertama kali, dan terus meningkat dengan bertambah dewasanya induk. Hal ini terjadi pada semua kondisi manajemen yang diterapkan. Hasil yang sejalan dilaporkan pula oleh SINGH et al. (1985) yang mendapatkan BL dari induk primipara lebih tinggi dibandingkan dengan BL dari induk pluripara (2,78 vs 3,20 kg). HAFEZ (1969)
menyatakan bahwa paritas induk mempengaruhi derajad perkembangan uterus dan vaskularitas dari uterus, yang kemudian kondisi ini mempengaruhi BL. Dinyatakan pula, pada induk-induk yang telah tua dan dengan pengalaman beranak yang banyak, uterus akan dipenuhi oleh lemak dan hal ini mengakibatkan pengaruh yang negatif pula terhadap bobot lahir anak.
Daya hidup anak
Gambar 4 memperlihatkan rataan daya hidup anak (DHA) pra sapih dari masing-masing genotipe ternak dari tahun ke tahun. Secara umum, induk dengan genotipe FecJFFecJF mempunyai DHA yang paling rendah dibandingkan genotipe lainnya. Seperti telah dilaporkan oleh INOUNU et al. (1997) bahwa induk-induk dengan genotipe FecJFFecJF lebih dipengaruhi
oleh lingkungan dibandingkan dengan ternak-ternak dengan genotipe lainnya, sedangkan induk-induk dengan genotipe FecJ+FecJ+ lebih tahan terhadap fluktuasi kondisi lingkungan. Rendahnya DHA pada induk-induk prolifik disebabkan tingginya jumlah anak yang dilahirkan. INOUNU et al. (1986) melaporkan kematian anak pada domba Ekor Tipis meningkat dengan meningkatnya jumlah anak sekelahiran, masing-masing 16,7; 18,4; 35,5; 42,9; dan 60% untuk JAS 1, 2, 3, 4, dan 5 secara berturut-turut.
Rataan DHA prasapih pada penelitian ini adalah 73,93% (N=1.437; SD=34,26; CV=46,36%). Sumber keragaman yang mempengaruhi nilai DHA adalah interaksi genotipe dan manajemen (P<0,06), paritas induk (P<0,05) dan pertambahan bobot induk (PKB, P<0,01).
Gambar 4. Rataan daya hidup anak pra-sapih per induk (DHA) pada masing-masing genotipe dari tahun 1981-1993
meningkatnya DHA (Tabel 1). HAFEZ (1969) menyatakan bahwa anak dari induk-induk yang kekurangan pakan secara umum akan sedikit lebih ringan bobot lahirnya dibandingkan dengan anak dari induk-induk yang cukup pakan dan juga anak dari induk-induk yang kurang pakan akan lahir prematur dan lebih ringan dibandingkan dari anak yang lahir dengan lama kebuntingan yang normal. Hal ini pula yang menyebabkan kematian yang lebih tinggi pada anak-anak yang dilahirkan dari induk dengan manajemen yang buruk. HAFEZ (1969) menjelaskan lebih lanjut bahwa induk yang menerima pakan buruk selama akhir kebuntingan menyebabkan pengurangan kandungan glikogen pada otot fetus dan terutama hati fetus. Penimbunan glikogen pada fetus akan meningkat pada akhir kebuntingan dan glikogen ini akan bertindak sebagai sumber energi sesaat setelah kelahiran. Oleh karena itu nutrisi induk yang buruk dapat meningkatkan kematian anak saat kelahiran.
INOUNU (1991) melaporkan bahwa 70% dari total kematian prasapih terjadi pada umur antara 1-6 hari setelah kelahiran. Selanjutnya, BOUJENANE (1996) melaporkan bahwa kematian yang tinggi pada awal kelahiran disebabkan oleh BL yang rendah dari JAS yang tinggi. OWENS et al. (1985) melaporkan bahwa pada semua tingkat JAS, anak-anak domba yang lebih berat bobot lahirnya lebih cepat berdiri dan menyusu, sehingga meningkatkan DHA dan kenaikan satu kg BL meningkatkan DHA sebesar 28%.
Kehadiran gen FecJF berakibat negatif terhadap DHA. Pada kondisi MNJ-1, ternak homosigot (FecJFFecJF) mempunyai DHA yang paling rendah. Namun sebaliknya, pada ternak-ternak prolifikasi rendah (FecJ+FecJ+), dengan MNJ-1, angka DHA telah menyamai angka DHA pada ternak-ternak pembawa gen FecJF, baik pada kondisi MNJ-2 maupun MNJ-3. Jelas terlihat bahwa ternak-ternak dengan prolifikasi rendah (FecJ+FecJ+) dapat bertahan dengan kondisi MNJ-1. Perbaikan manajemen memang menghasilkan DHA yang lebih tinggi. Hal ini disebabkan pada induk-induk prolifikasi rendah (FecJ+FecJ+) hanya ada 17% dari induk yang melahirkan anak kembar dua. Pada kondisi demikian kematian anak lebih banyak disebabkan oleh induk yang tidak mau menyusui anaknya, baik karena kondisi kelenjar susunya yang kering atau adanya infeksi pada puting susu yang menyebabkan iritasi.
Pada ternak-ternak homosigot (FecJFFecJF) penerapan MNJ-1 menghasilkan DHA paling rendah dari semua kelompok. Hal ini dapat dimengerti karena pada kelompok ini ada 84% dari induk yang melahirkan anak kembar dua atau lebih. Kekurangan gizi pada induk-induk ini akan berakibat buruk, karena fetus akan berebut untuk mendapatkan gizi yang memang telah terbatas, akibatnya dihasilkan anak yang bobot lahirnya
beragam. Pada anak-anak dengan bobot badan rendah biasanya kondisi tubuhnya lemah, lambat untuk berdiri atau sama sekali tidak dapat berdiri dan menyusu pada induknya. Hal ini menyebabkan kematian anak sangat tinggi. OWENS et al. (1985) melaporkan satu menit perpanjangan waktu dari sejak kelahiran ke saat anak mulai belajar berdiri, berdiri atau belajar mencari puting susu induk mengakibatkan menurunnya DHA sebesar 1%. SHRESTHA et al. (1992) melaporkan penurunan DHA dengan meningkatnya jumlah anak yang dilahirkan. Pada anak tipe kelahiran satu didapatkan DHA=84%, dan menurun dengan kenaikan tipe kelahiran, sehingga pada tipe kelahiran 5 daya hidupnya hanya sebesar 55%. Selanjutnya BRADFORD (1972) dan HINCH et al. (1985) melaporkan adanya hubungan yang erat antara bobot lahir dan daya hidup anak. INOUNU et al. (1993) melaporkan bahwa 60-84% dari DHA pada berbagai tipe kelahiran dipengaruhi oleh bobot lahir.
Peningkatan manajemen ke MNJ-2 menghasilkan DHA yang lebih tinggi, namun peningkatan lebih lanjut ke MNJ-3, secara statistik tidak terlihat adanya peningkatan yang nyata (P>0,05), kecuali pada induk-induk dengan genotipe FecJFFecJF terjadi peningkatan DHA sebanyak 9,98% dibandingkan pada MNJ-2. Hal ini menunjukan bahwa pada domba dengan genotipe FecJFFecJF masih dapat ditingkatkan daya hidup anaknya dengan perbaikan manajemen. Pada domba betina Barbados Blackbelly, dilaporkan bahwa pada umumnya domba ini tidak memproduksi cukup banyak air susu untuk merawat lebih dari dua ekor anak walaupun dipelihara pada kondisi manajemen yang baik (RASTOGI, 1996).
Paritas induk mempengaruhi daya hidup anak, seperti juga pada sifat-sifat yang lainnya. Pada paritas pertama DHA lebih rendah 9% dibandingkan pada paritas kedua. Pada paritas induk yang ke-2 didapatkan DHA tertinggi, kemudian menurun pada paritas induk yang ke-3 dan bertahan sampai pada paritas ke-5. Hasil yang serupa juga dilaporkan oleh SHRESTHA et al. (1992), bahwa DHA pada betina-betina muda lebih rendah dibandingkan pada betina dewasa (67% vs
Pertambahan bobot induk (PKB) sebanyak satu kg mengakibatkan kenaikan daya hidup anak sebesar 2,3%. Perbedaan pertambahan bobot badan induk selama kebuntingan dapat terlihat pada Tabel 2. Angka-angka pada Tabel 2 menunjukkan bahwa pada MNJ-1, ternak dengan genotipe FecJFFecJF paling menderita akibat cekaman kurang pakan, yang ditunjukkan dengan pertumbuhan induk selama kebuntingan yang paling rendah. Hal ini disebabkan semua cadangan kalori induk digunakan untuk merawat kebuntingan dengan jumlah anak yang banyak. Namun, pada ternak dengan genotipe FecJ+FecJ+ dengan MNJ-1, pertambahan bobot badan induk selama kebuntingan telah sama dengan yang dicapai pada induk-induk pembawa gen FecJF pada tingkat MNJ-2. Tidak
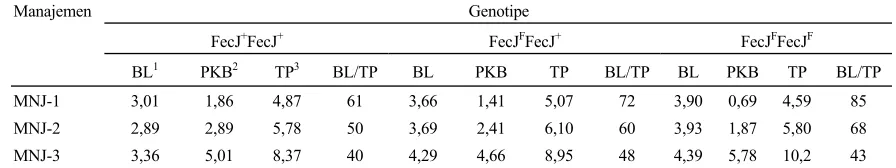
mengherankan apabila selanjutnya kondisi tubuh induk yang terjadi akibat kebuntingan akan berpengaruh terhadap daya hidup anak yang dilahirkan. INOUNU et al. (1993) melaporkan bahwa pertambahan bobot induk saat kebuntingan mempengaruhi DHA antara 11-27%, tergantung jumlah anak yang dikandungnya. Apabila nilai PKB pada Tabel 2 ditambah dengan nilai BL dari masing-masing kelompoknya, maka hasil dari penjumlahan nilai-nilai tersebut akan mencerminkan total produksi (TP) seekor induk pada saat beranak, yaitu berupa pertambahan bobot badannya sendiri dan bobot anak yang dihasilkannya. Selanjutnya apabila total bobot lahir per induk dibagi dengan nilai total produksi per induk maka akan didapatkan angka
persentase bobot lahir total dibandingkan dengan total produksi per induk (Tabel 3).
Angka-angka pada Tabel 3 menunjukkan bahwa pada tingkat MNJ-1 dan MNJ-2, kecuali pada ternak dengan genotipe FecJ+FecJ+, lebih dari 60% dari total
produksi induk digunakan hanya untuk produksi anak (BL). Terutama pada ternak dengan genotipe FecJFFecJF pada MNJ-1, hanya 15% saja dari total produksi yang digunakan untuk meningkatkan bobot tubuh induk. Padahal pertambahan bobot badan induk pada saat kebuntingan erat hubungannya dengan persiapan induk untuk memproduksi air susu. Pada induk-induk yang tercukupi kebutuhan gizinya, kondisi tubuh induk lebih baik dibandingkan induk yang kekurangan gizi, sehingga produksi susu induk akan lebih tinggi dibandingkan dengan induk-induk yang kurang pakan dan daya hidup anak pun akan lebih tinggi. Di samping itu pada induk-induk yang tercukupi gizinya penimbunan glikogen pada otot fetus berlangsung normal. Sesaat setelah kelahiran, anak domba perlu tenaga untuk berdiri dan menyusu pada induknya, sedangkan pada anak-anak dari induk yang kurang gizi, sumber energi yang diperlukan untuk berdiri tidak tercukupi. Lebih jauh lagi, HAFEZ (1969) menyatakan kekurangan gizi pada induk bunting mengakibatkan memburuknya otot lokomotor, hal ini akan menyebabkan keterlambatan anak untuk menyusu pada induknya dan mengakibatkan kematian dini pada anak yang baru lahir.
Tabel 2. Jumlah pengamatan (N), rataan kuadrat terkecil dari pertambahan bobot badan induk selama kebuntingan (PKB, kg) dan salah baku (SE) dari tiga genotipe prolifikasi pada manajemen yang berbeda
Genotipe
Manajemen FecJ+FecJ + FecJFFecJ+ FecJFFecJF
N PKB SE N PKB SE N PKB SE
MNJ-1 174 1,86a (0,28) 213 1,41a (0,26) 59 0,69a (0,53)
MNJ-2 272 2,89b (0,21) 312 2,41b (0,20) 121 1,87a (0,32)
MNJ-3 96 5,01c (0,35) 107 4,66c (0,33) 83 5,78b (0,36)
Keterangan : Huruf superskrip yang sama menunjukkan tidak berbeda nyata (P>0,05)
Tabel 3. Nisbah BL/TP (%) dari tiga genotipe prolifikasi dan manajemen yang berbeda
Manajemen Genotipe
FecJ+FecJ+ FecJFFecJ+ FecJFFecJF
BL1 PKB2 TP3 BL/TP BL PKB TP BL/TP BL PKB TP BL/TP
MNJ-1 3,01 1,86 4,87 61 3,66 1,41 5,07 72 3,90 0,69 4,59 85
MNJ-2 2,89 2,89 5,78 50 3,69 2,41 6,10 60 3,93 1,87 5,80 68 MNJ-3 3,36 5,01 8,37 40 4,29 4,66 8,95 48 4,39 5,78 10,2 43
Keterangan :
1
:BL=bobot lahir per induk (kg);
2
3
:TP=BL+PKB (kg)
Total bobot sapih per induk
Gambar 5 memperlihatkan total bobot sapih (BS) per induk dari ketiga genotipe ternak dari tahun ke tahun. Fluktuasi BS dari tahun ke tahun menunjukkan adanya gabungan pengaruh musim, ketersediaan pakan dan manajemen terhadap BS. Secara umum, BS dari induk dengan genotipe FecJFFecJF lebih tinggi dibandingkan dengan kedua genotipe lainnya. FAHMY (1989) melaporkan adanya pengaruh musim terhadap BS pada domba Romanov. Induk-induk yang dikawinkan pada musim dingin, panas dan gugur menghasilkan BS masing-masing sebesar 36,1; 35,5; dan 45,3 kg secara berturut-turut.
Gambar 5. Rataan total bobot sapih anak per induk (BS) pada masing-masing genotipe dari tahun 1981-1993
Produksi seekor induk dinilai dari jumlah anak dan jumlah kilogram dari anak-anak yang dihasilkan pada satuan waktu tertentu, yang dalam hal ini adalah bobot sapih total. Rataan BS yang diperoleh dalam penelitian ini adalah 13,12±4,33 kg/induk/kelahiran (N=1.224; CV=33,0%). Hasil ini lebih tinggi dari BS yang dilaporkan oleh INIGUEZ et al. (1991) pada domba lokal Sumatera sebesar 11,45 kg, tetapi lebih rendah dari BS domba Dorset dan Finnsheep dari negeri beriklim dingin, yaitu masing-masing sebesar 21,3 dan 17,6 kg (INIGUEZ et al., 1986), dan juga lebih rendah bila dibandingkan dengan BS domba Romanov sebesar 39,0 kg seperti yang dilaporkan oleh FAHMY (1989).
Nisbah antara produksi anak (BS) per induk dengan bobot tubuh induk (BS/BB induk) pada penelitian ini adalah 0,55. Nilai nisbah ini lebih tinggi dari yang didapat pada domba Sumatera maupun Dorset dan Finnsheep (INIGUEZ et al., 1991) yang masing-masing mempunyai nilai nisbah produksi sebesar 0,52; 0,41; dan 0,42. Hal ini menunjukkan bahwa domba
pada penelitian ini mempunyai efesiensi produksi yang tinggi dibandingkan domba-domba yang disebutkan tadi.
BS dipengaruhi oleh genotipe dan manajemen (P<0,01; Tabel 1). Secara keseluruhan, peningkatan manajemen ke arah yang lebih baik meningkatkan bobot sapih total per induk pada semua genotipe. Namun pada tingkat MNJ-1, tidak ada keistimewaan dari ternak-ternak pembawa gen FecJF, semua induk menghasilkan bobot sapih di bawah 11 kg. Hal ini merupakan pencerminan dari kombinasi kondisi tubuh induk yang buruk diiringi rendahnya produksi susu induk, rendahnya bobot lahir anak dan tingginya angka kematian prasapih. HAFEZ (1969) menyimpulkan bahwa pada induk-induk yang kekurangan pakan pada saat kebuntingan, tidak saja menyebabkan pertumbuhan yang rendah pada saat fetus masih dalam kandungan (pranatal) tetapi juga berakibat pada rendahnya pertumbuhan pascalahir. Hal tersebut terbukti setelah dilakukan peningkatan manajemen pakan. Pada MNJ-2 terlihat perbedaan antara ternak normal dengan ternak pembawa gen FecJF, bahkan pada MNJ-3 ternak pembawa gen FecJF dapat menghasilkan bobot sapih lebih dari 19 kg. Dengan demikian dapat disimpulkan bahwa peningkatan manajemen ke arah yang lebih baik tidak saja memperbaiki kondisi induk tetapi juga sekaligus meningkatkan produksi induk pada saat sapih.
Hal lain yang mempengaruhi BS adalah interaksi manajemen dan paritas induk (P<0,05; Tabel 1) dan PKB (P<0,01). Namun, INIGUEZ et al. (1991) melaporkan tidak ada pengaruh yang nyata dari paritas induk terhadap BS pada domba Sumatera.
Bertambah dewasanya induk diiringi pula dengan meningkatnya kemampuan induk untuk merawat anaknya, sehingga dihasilkan bobot sapih yang meningkat. Pada semua tingkat manajemen bobot sapih tertinggi dicapai pada paritas keempat dan kemudian menurun kembali pada paritas kelima. FAHMY (1989) melaporkan hasil penelitiannya pada induk-induk domba dengan paritas 1, 2, 3, 4, 5, dan ≥6 menghasilkan BS sebesar 29,8; 42,2; 41,2; 38,3; 43,1; dan 39,2 kg secara berturut-turut.
KESIMPULAN
Dari hasil penelitian ini dan pembahasan maka dapat ditarik kesimpulan sebagai berikut:
Rataan jumlah anak sekelahiran (JAS) yang didapatkan adalah sebesar 1,77 ekor per induk. Keragaman JAS dipengaruhi oleh genotipe, manajemen dan interaksi antara manajemen dengan paritas induk serta pertambahan bobot badan induk (PKB). Kehadiran satu duplikat gen FecJF dapat meningkatkan
JAS sebanyak 0,8 ekor per induk. Perubahan manajemen ke tingkat yang lebih baik juga meningkatkan JAS. Pada induk-induk yang baru pertama kali beranak dihasilkan JAS lebih rendah dibandingkan induk-induk yang telah beranak dua kali, hal ini terjadi pada semua tingkat manajemen. Peningkatan satu kg bobot badan induk (PKB) diikuti dengan meningkatnya JAS sebanyak 0,04 ekor.
Rataan total bobot badan anak saat lahir (BL) adalah 3,43 kg. Keragaman BL dipengaruhi oleh genotipe, manajemen dan interaksi manajemen dengan paritas induk. Pada semua tingkat manajemen induk-induk beranak pertama kali (primipara) menghasilkan BL lebih rendah dibandingkan induk-induk yang telah beranak beberapa kali (pluripara).
Rataan daya hidup anak prasapih (DHA) dari domba prolifik didapatkan sebesar 73,93%, keragamannya dipengaruhi oleh interaksi genotipe dan manajemen, paritas induk dan pertambahan bobot badan induk. Pada paritas pertama induk mempunyai DHA 11% lebih rendah dibandingkan pada paritas kedua. Kenaikan pertambahan bobot badan induk sebesar 1 kg meningkatkan DHA sebesar 2,3%.
Keragaman total bobot badan anak saat sapih (BS) dipengaruhi oleh interaksi genotipe dan manajemen, paritas induk dan pertambahan bobot badan induk (PKB). Rataan BS yang didapat adalah 13,12 kg.
DAFTAR PUSTAKA
BINDON, B.M., L.R. PIPER, and T.S. CH'ANG. 1984. Reproductive performance of crossbred ewes derived from Booroola and Control Merinos and Joined to rams of two terminal sire breeds. In : Reproduction in Sheep. D.R. LINDSAY and D.T. TORELL (Eds.). Australian Academy of Science, Canberra. pp.243-246.
BOUJENANE, I. 1996. The D'Man. In : Prolific Sheep. M. H. FAHMY (Eds.). CAB International. pp. 109-120.
BRADFORD, G.E. 1972. The role of maternal effect in animal breeding. VII Maternal effects in sheep. J. Anim. Sci.
35:1324-1334.
BRADFORD, G.E. 1985. Selection for litter size. In : Genetic of Reproduction in Sheep. R.B. LAND and D.W. ROBINSON (Eds.). Butterworths, London. pp. 3-18.
BRADFORD, G.E., J.F. QUIRKE, P. SITORUS, I. INOUNU, B. TIESNAMURTI, F.L. BELL, I.C. FLETCHER, and D.T. TORELL. 1986. Reproduction in Javanese Sheep: Evidence for gene with large effect on ovulation rate and litter size. J. Anim. Sci. 63:418-431.
BRADFORD, G.E., I. INOUNU, L.C. INIGUEZ, B.TIESNAMURTI, and D.L. THOMAS. 1991. The prolificacy gene of Javanese sheep. In : Major Genes for Reproduction in Sheep. J.M. ELSEN, L. BODIN, and J. THIMONIER (Eds.). Proc. 2nd Int. Workshop, Toulouse, France, July 16-18, 1990. pp. 67-74.
BRADFORD, G.E. and I. INOUNU. 1996. Prolific breeds of Indonesia. In : Prolific Sheep. M.H. FAHMY (Eds.). CAB International. pp. 137-145.
COOP, I.E. 1966. Effect of flushing on reproductive performance of ewes. J. Agric. Sci. (Camb.). 67:305-325.
DONALD, H.P. and W.S. RUSSEL. 1970. The relationship between live weight of ewe at mating and weight of newborn lamb. Anim. Prod. 12:273-280.
FAHMY, M.H. 1989. Reproductive performance, growth and wool production of Romanov sheep in Canada. Small Ruminant Res. 2:253-264.
FAHMY, M.H. 1996. The Romanov. In : Prolific Sheep. M.H. FAHMY (Eds.). CAB International. pp. 47-72.
FITZUGH, H.A. and G.E. BRADFORD. 1983. Productivity of hair sheep and opportunities for improvement. In : Hair
Sheep of Western Africa and the Americas. H.A.
FITZUGH and G.E. BRADFORD (Eds.). Westview Press, Boulder-Colorado. pp. 23-54.
GUNN, R.G. and J.M. DONEY. 1979. Fertility in Cheviot ewes. I. The effect of body condition at mating on ovulation rate and early embryo mortality in North and South Country Cheviot ewes. Anim. Prod. 29:11-16.
GUNN, R.G. 1983. The influence of nutrition on the reproductive performance of ewes. In : Sheep
Production. W. HARESIGN (Eds.). Butterworths,
London. pp. 99-110.
HAFEZ, E.S.E. 1969. Prenatal growth. In: Animal Growth and Nutrition. E.S.E. HAFEZ and I.A. DYER (Eds.). Lea and Febiger. Philadelphia. pp. 21-39.
HINCH, G.N., S.F. CROSBIE, R.W. KELLY, J.L. OWENS, and G.H. DAVIS. 1985. Influence of birth weight and litter size on lamb survival in high fecundity Booroola-Merino crossbred flocks. N. Z. J. Agric. Res. 28:31-38.
HOHENBOKEN, W.D. and S.E. CLARKE. 1981. Genetics environmental and interaction effects on lamb survival, cumulative lamb production and longevity of crossbred ewes. J. Anim. Sci. 53:956-976.
HUNTER, G.L. 1956. The maternal influence on size in sheep.
J. Agric. Sci. 48:36-60.
Dorset and Finnsheep ewes and their F1 crosses. J. Anim. Sci. 63:715-728.
INIGUEZ, L.C., M. SANCHEZ, and S.P. GINTING. 1991. Productivity of Sumatran sheep in a system integrated with rubber plantation. Small Ruminant Res. 5:303.
INOUNU, I. 1991. Production Performance of Prolific Javanese Sheep. MS Thesis. Program Pascasarjana Institut Pertanian Bogor. Bogor.
INOUNU, I., SUBANDRIYO, N. THOMAS, P. SITORUS, and M. BELL. 1986. Lambing characteristics of Javanese thin-tail ewes at Cicadas experiment station and under village conditions. Ilmu dan Peternakan II(2): 79-82.
INOUNU, I., L.C. INIGUEZ, G.E. BRADFORD, SUBANDRIYO, and B. TIESNAMURTI. 1993. Performance production of prolific Javanese ewes. Small Ruminant Res. 12:243-257.
INOUNU, I., L.C. INIGUEZ, and A. DJAJANEGARA. 1994. Production traits of prolific sheep under different feeding levels. Proc. 7th AAAP Anim. Sci. Conf. July, 1994. Kuta-Bali, Indonesia. I:11-12.
INOUNU, I., B. TIESNAMURTI, dan A. PRIYANTI. 1995. Pemanfaatan Gen FecJF dalam Pengembangan Usaha Ternak Domba untuk Tujuan Komersial. Laporan. Balai Penelitian Ternak dan Proyek Pembinaan Kelembagaan Penelitian Pertanian (ARMP). Bogor.
INOUNU, I., B. TIESNAMURTI, SUBANDRIYO, dan H. MARTOJO. 1997. Laju ovulasi dan daya hidup embrio pada domba prolifik. Media Veteriner 4(3): 25-38.
LAHLOU-KASSI, A. and M. MARIE. 1985. Sexual and ovarian function of the D'man ewe. In: Genetic of Reproduction
in Sheep. R.B. LAND and D.W. ROBINSON (Eds.).
Butterworths, London. pp. 245-260.
LAND, R.B. 1977. Reproduction in young sheep. Some genetic and environmental sources of variation. J. Reprod. Fert. 52:427-436.
MAIJALA, K. 1996. The Finnsheep. In: The Prolific Sheep.
M.H. FAHMY (Eds.). CAB International. pp. 10-46.
MASON, I.L. 1980. Prolific tropical sheep. FAO. Animal production and health paper No. 17. Food and Agriculture Organization, Rome. p. 124
OWENS, J.L., B.M. BINDON, T.N. EDEY, and L.R. PIPER. 1985. Behavior at parturition and lamb survival of Booroola Merino sheep. Livestock Prod. Sci. 13:359-372.
PIPER, L.R., B.M. BINDON, and G.H. DAVIS. 1985. The single gene inheritance of the prolificacy of the Booroola Merino. In: Genetic of Reproduction in Sheep. R.B. LAND and D.W. ROBINSON (Eds.). Butterworths, London. pp. 115-125.
PRIYANTO, D., B. SETIADI, I. INOUNU, dan SUBANDRIYO. 1992. Produktivitas domba pedesaan pada kondisi pemeliharaan tradisional di Cirebon. Ilmu dan Peternakan 5(1):15-19.
RASTOGI, R.K. 1996. The Barbados Blackbelly. In : Prolific
Sheep. M. H. FAHMY (Eds.). CAB International. pp.
121-136.
REESE, A.A., S.W. HANDAYANI. S.P. GINTING, S. SINULINGGA, G.R. REESE, and W.L. JOHNSON. 1990. Effect of energy supplementation on lamb production of Javanese thin-tail ewes. J. Anim. Sci. 68:1827-1840.
SAS. 1987. SAS/STAT Guide for Personal Computers. Version 6 Edition. SAS Institute Cary., NC, USA.
SETIADI, B., I. INOUNU, dan SUBANDRIYO. 1995. Produktivitas induk domba ekor tipis di Kabupaten Bogor (Studi Kasus Desa Ciburuy, Kecamatan Cijeruk). Media. Ed. Khusus. Fakultas Peternakan, Universitas Diponegoro, Semarang. pp. 337-345.
SHRESTHA, J.N.B., D.P. HEANEY, and R.J. PARKER. 1992. Productivity of three synthetic Arcott sheep breeds and their crosses in terms of 8-mo breeding cycle and artificially reared lambs. Small Ruminant Res. 9: 283-296.
SINGH, G., A.K. GUPTA, and D.C. GUPTA. 1985. Effect of foetal genotype and ewes parity on the birth weight of the lamb. Indian J. Anim. Sci. 55: 706-709.
SITORUS, P. and I. INOUNU. 1983. The effect of altitude and ewe's body weight on litter size, total birth weight, and weaning weight of the thin tailed sheep. Proc. 5th World Conf. Anim. Prod. Vol 2. Tokyo.
SMEATON, D.C., P.V. RATTRAY, P. MACKISACK, and S. HEATH. 1983. Nutrition and management of ewes before and after lambing. Proc.N. Z. Soc. Anim. Prod. 43: 37-40.