POLA PEWARISAN BEBERAPA KARAKTER
KUALITATIF DAN KUANTITATIF PADA CABAI
(Capsicum annuum L.)
Oleh
Muhammad Dzikri Alif A34403052
PROGRAM STUDI
PEMULIAAN TANAMAN DAN TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
2008
POLA PEWARISAN BEBERAPA KARAKTER
KUALITATIF DAN KUANTITATIF PADA CABAI
(Capsicum annuum L.)
Skripsi sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian Institut Pertanian Bogor
Oleh
Muhammad Dzikri Alif A34403052
PROGRAM STUDI
PEMULIAAN TANAMAN DAN TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
2008
BEDANYA AKU.. ORANG TUAKU DAN GURUKU : BELIAU MEMILIKI
PENGALAMAN YANG TIDAK AKU MILIKI
Orang
bijak
bilang
:
“Kita tidak bisa bercermin pada air yang mengalir
dan hanya bisa pada air yang diam”dan aku ingin mencobanya...diam
untuk kedua orang tuaku,guruku, dan sahabatku
M. Dzikri Alif, Februari 2008
IPB BOGOR
Mohon doa.. semoga hidupku bermakna
RINGKASAN
MUHAMMAD DZIKRI ALIF. Pola Pewarisan Beberapa Karakter Kualitatif dan Kuantitatif pada Cabai (Capsicum annuum L.). Dibimbing oleh MUHAMAD SYUKUR dan SRIANI SUJIPRIHATI.
Penelitian dilaksanakan pada bulan Februari hingga Juli 2007 di Kebun Percobaan IPB Leuwikopo Darmaga. Penelitian ini bertujuan untuk mempelajari pewarisan sifat kualitatif dan kuantitatif serta heritabilitas pada beberapa karakter cabai.
Bahan tanaman yang digunakan adalah P1 (IPB C15), P2 (IPB C10), F1(15x10), F1R (10x15), BCP1 ((15x10)x15), BCP2 ((15x10)x10) dan F2. Penelitian ini tidak menggunakan rancangan percobaan, tetapi setiap populasi (kecuali F2) diulang dua kali.
Hasil penelitian menunjukkan bahwa pada populasi F2, pola segregasi sifat permukaan buah, lekukan di pangkal buah, dan orientasi buah sesuai dengan nisbah Mendel dengan perbandingan 3:1. Hal ini dapat berarti bahwa sifat permukaan buah, lekukan pada pangkal buah, dan orientasi buah dikendalikan oleh satu pasang gen dominan penuh. Sifat posisi bunga menunjukan hasil yang nyata dengan perbandingan 9:3:4. Posisi bunga dikendalikan oleh dua pasang gen dengan interaksi epistasis resesif. Sifat warna daun menunjukkan nisbah perbandingan 9:7. Hal ini berarti warna daun dikendalikan oleh dua pasang gen. Perbandingan tersebut menunjukkan interaksi epistasis resesif ganda.
Tinggi dikotomus menunjukkan sebaran normal yang mengindikasikan bahwa karakter tersebut dikendalikan oleh gen minor. Nilai heritabilitas arti sempit adalah 0.47 dan heritabilitas arti luas adalah 0.45 untuk karakter tinggi dikotomus. Pada karakter panjang buah, diameter, tinggi dikotomus, bobot total, dan bobot layak pasar dan orientasi buah terdapat pewarisan ekstrakromosomal.
Judul : POLA PEWARISAN BEBERAPA KARAKTER KUALITATIF DAN KUANTITATIF PADA CABAI (Capsicum annuum L.) Nama : Muhammad Dzikri Alif
NRP : A34403052
Menyetujui, Dosen Pembimbing
Pembimbing I Pembimbing II
Dr. Muhamad Syukur, SP, MSi Prof.Dr. Ir. Sriani Sujiprihati, MS NIP. 132 258 034 NIP. 131 284 838
Mengetahui, Dekan Fakultas Pertanian
Prof. Dr. Ir. Didy Sopandie, MAgr NIP. 131 124 019
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 29 Juli 1985 di Jakarta. Penulis merupakan anak kedua dari tiga bersaudara dari keluarga Bapak Abdullah dan Ibu Etty Latifah.
Penulis lulus dari Sekolah Dasar Negeri Pancoran 01 Pagi pada tahun 1997, tahun 2000 penulis menyelesaikan pendidikan di SLTP Negeri 115. Penulis melanjutkan studi di SMU Negeri 3 Jakarta dan lulus pada tahun 2003.
Penulis diterima di IPB pada tahun 2003 melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB). Selama menjadi mahasiswa penulis aktif mengikuti berbagai kegiatan di IPB seperti, peserta Seminar Nasional Pertanian yang diselenggarakan BEM Fakultas tahun 2004, peserta pelatihan pembuatan web
design yang diselenggarakan BEM FMIPA tahun 2005, peserta GRADASI
Himagron dalam cabang Bulutangkis 2006, peserta Festival Musik yang diselenggarakan di lingkungan kampus tahun 2007 Pengawas ujian TPB tahun 2006 dan 2007.
Penulis juga aktif dalam berbagai kegiatan kepanitiaan bedah kampus di SMUN 3 Jakarta mewakili IPB tahun 2006, panitia fieldtrip PMTTB 40 tahun 2005-2006, panitia Seminar dan pameran Pertanian dan panitia AMT (Achievment
Motivating Training) KKP IPB di Kabupaten Tegal. Penulis juga aktif dalam
kegiatan organisasi di IPB seperti di Himpunan Mahasiswa Agronomi (Himagron) tercatat sebagai anggota Himagron pada tahun 2004, dalam kepanitiaan MPD Lintas Desa sebagai senior pada tahun 2006 dan 2007, panitia Open House Departemen Agronomi dan Hortikultura tahun 2006 dan 2007, Koordinator Olimpiade Mahasiswa IPB dalam seleksi Bulutangkis di Faperta tahun 2007, Badan Eksekutif Mahasiswa Faperta pada tahun 2007 dalam kepanitiaan MPF 43, BEM KM IPB dalam kepanitiaan IEE (International Education Expo) di Botani Square pada tahun 2007, panitia Seminar Nasional Peningkatan Perolehan HKI dari Hasil Penelitian Yang Dibiayai Oleh Hibah Kompetitif Departemen Agronomi dan Hortikultura tahun 2007.
KATA PENGANTAR
Puji syukur penulis panjatkan kehadirat Alah SWT atas segala rahmat dan karunia-Nya. Penulis dapat menyelesaikan penelitian yang berjudul Pendugaan Parameter Genetik Beberapa Karakter Kualitatif dan Kuantitatif pada Cabai
(Capsicum annuum L.) sebagai tugas akhir mahasiswa Strata 1 untuk memperoleh
gelar Sarjana Pertanian pada Departemen Agronomi dan Hortikultura Fakultas Pertanian IPB.
Penulis menyampaikan terima kasih kepada berbagai pihak yang telah memberikan perhatian, semangat, motivasi, bimbingan, pengarahan, dan doa selama kegiatan penelitian dan penulisan skripsi antara lain :
1. Kepada kedua orang tua Bapak saya Abdullah dan almarhumah Ibu saya tercinta Etty Latifah yang telah memberikan perhatian dan doa selama penulis kuliah di IPB mulai dari masa TPB sampai dengan akhir hayatnya, teteh Tia, dan adik Aat.
2. Dr. Ir. Trikoesoemaningtyas selaku pembimbing akademik atas bimbingan dan pengarahannya selama penulis kuliah.
3. Dr. Muhamad Syukur, SP., MSi dan Dr. Ir. Sriani Sujiprihati, MS selaku pembimbing skripsi yang telah memberikan arahan dan bimbingan dalam penelitian dan penyusunan skripsi ini.
4. Dr Yudiwanti selaku dosen penguji skripsi yang telah memberikan kritik, saran, dan masukan dalam penyusunan skripsi ini.
5. Ibu Rahmi, Bapak Willy Bayuardi, Bapak Sarju, Mba Mawi, Mba Cici, Mas Undang, Pak Anen, Pak Maman, Pak Mamad yang telah membantu selama proses penelitian sampai dengan penyusunan skripsi.
6. Ibu dan Bapak guru yang telah mendidik dan mengajar sewaktu di SDN Pancoran 01 Pagi, SLTPN 115, dan SMUN 3 Jakarta.
7. Para dosen selama penulis masih di TPB sampai dengan penulis kuliah di program studi Pemuliaan Tanaman dan Teknologi Benih atas bimbingan dan arahannya selama penulis kuliah
8. Teman-teman satu tim penelitian cabai (Tri, Tedi, Habib, Regi, Sita, Iqra, Isma, Mite, dan Mba Indah).
9. Sahabat-sahabat penulis selama kuliah di IPB Ipul, Toni, Angga, Tedi, Mas Yudi, Bambang, Anto. Terima kasih atas kerjasama, kebersamaan, bantuan, dan semangatnya selama ini. Nanda dan Wili yang telah bersedia hadir dalam seminar penulis. Teman-teman satu band CRAZH (Cristian, Roni, Adit, dan Habib) video klip di Tamara kenangan tersendiri buat penulis. Teman-teman Ana, Farah, Tika, Mira, Elmyra, Vita, Febi, Mila, Lina, Yuseffa, Ratih dan keluarga besar PMTTB 40, PMTTB 41, IE 42, AGB 42, terima kasih atas kebersamaannya selama penulis kuliah.
10.Teman –Teman KKP 2006 Kabupaten Tegal (Iyus, Rina, Ana, Hadi, Joko) atas kebersamaan selama 2 bulan bersama penulis.
11.Semua pihak yang telah membantu selama kuliah dan penelitian yang tidak bisa disebutkan satu persatu.
Bogor, Februari 2008 Penulis
DAFTAR ISI
Halaman PENDAHULUAN Latar Belakang………... 1 Tujuan………... 2 Hipotesis………... 2 TINJAUAN PUSTAKA Botani dan Morfologi Cabai... 3Pemuliaan Tanaman Cabai... 4
Studi Pewarisan Beberapa Karakter Kualitatif dan Kuantitatif pada Cabai ... ... 5
BAHAN DAN METODE Waktu dan Tempat Penelitian... 9
Bahan dan Alat... 9
Metode Penelitian... 9
Pelaksanaan Penelitian... 9
Pengamatan Penelitian... 10
Analisis Data... 11
HASIL DAN PEMBAHASAN Kondisi Umum Pertanaman... ... 13
Pola Pewarisan Sifat Kualitatif... 13
Karakter Kuantitatif... 22
Derajat Dominansi... 24
Pewarisan Ekstrakromosomal... ... 25
KESIMPULAN DAN SARAN Kesimpulan... 27
Saran... 27
DAFTAR PUSTAKA... 28
LAMPIRAN... 30
DAFTAR TABEL
Teks
Nomor Halaman
1. Nisbah Fenotipik Sifat Resistensi yang dikendalikkan oleh gen Mayor pada
populasi F2... 12
2. Beberapa Karakter Kualitatif Cabai pada Populasi P1, P2, F1, dan F1R... 13
3. Nisbah Fenotipik Karakter Kualitatif Cabai pada Populasi BCP1, BCP2, dan F2... 14
4. Hasil Uji Khi Kuadrat Karakter Kualitatif Cabai Populasi F2... 14
5. Hasil Uji Normalitas Karakter Kuantitatif Cabai pada Populasi F2... 22
6. Ragam Tetua, Backcross, F1, F2, dan Heritabilitas Karakter Kuantitatif... 23
7. Nilai Tengah Karakter Kuantitatif Cabai dan Aksi Gen... 24
8. Nilai t Hitung pada Karakter Kuantitatif Cabai Populasi F1 dan F1R... 25
Lampiran Nomor Halaman 1. Nisbah Segregasi Karakter Permukaaan Buah... 30
2. Nisbah Segregasi Karakter Leher Buah... 30
3. Nisbah Segregasi Karakter Orientasi Buah………... 30
4. Nisbah Segregasi Karakter Bentuk Calix... 30
5. Nisbah Segregasi Karakter Warna Daun... 31
6. Nisbah Segregasi Karakter Tipe Pertumbuhan... 31
7. Nisbah Segregasi Karakter Tipe Percabangan... 31
8. Nisbah Segregasi Karakter Posisi Bunga... 31
DAFTAR GAMBAR
Teks
Nomor
Halaman
1. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Permukaan Buah...15
2. Bagan Persilangan antara F1 dan P1 serta F1 dengan P2 untuk Karakter Permukaan Buah..16
3. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Lekukan di Pangkal
Buah...17
4. Bagan Persilangan antara F1 dan P1 serta F1 dengan P2 untuk Karakter Lekukan di Pangkal
Buah...17
5. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Orientasi Buah...18
6. Bagan Persilangan antara F1 & P1 serta F1 dengan P2 untuk Karakter Orientasi Buah...19
7. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Penyempitan Tangkai
Buah...20
8 Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Warna Daun...21
9. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Posisi Bunga...22
Lampiran
Nomor
Halaman
1. Gambar Aksi Gen Karakter Panjang Buah...31
2. Gambar Aksi Gen Karakter Diameter Buah...31
3. Gambar Aksi Gen Karakter Tinggi Dikotomus...31
4. Gambar Aksi Gen Karakter Bobot Total...31
5. Gambar Aksi Gen Karakter Bobot Layak Pasar...31
6. Kondisi Umum Pertanaman Cabai di Lapang...32
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) adalah anggota famili Solanaceae, merupakan tanaman yang dibudidayakan secara luas di seluruh dunia. Kandungan gizi pada cabai membuat cabai dapat dikonsumsi baik sebagai sayuran, bumbu bahkan sebagai sumber baku obat – obatan. Zat gizi yang terkandung dalam cabai antara lain protein, lemak, karbohidrat, kalsium, fosfor, besi, vitamin A, B, C, dan senyawa alkaloid yaitu capsaicin (rasa pedas), flavonoid, dan minyak esensial (Agfi, 2006).
Produkiivitas cabai di Indonesia masih tergolong rendah walaupun termasuk negara yang memiliki luasan produksi terbesar di Asia. Produktivitas cabai khususnya di Asia dapat mencapai 18 ton/ha. Deptan (2007) mencatat produktivitas cabai di Indonesia tahun 2000-2006 berturut-turut sebesar 4.17, 4.07, 4.22, 6.05, 5.66, 5.65, dan 5.79 ton/ha.
Salah satu cara untuk meningkatkan produktivitas cabai adalah melalui pemuliaan tanaman. Kegiatan pemuliaan tanaman adalah koleksi, seleksi, peningkatan keragaman dan evaluasi. Dalam melakukan kegiatan pemuliaan tanaman, diperlukan adanya keragaman genetik yang terkait dengan heritabilitas. Menurut Nasir (2001) heritabilitas adalah proporsi besaran ragam genetik terhadap besaran total ragam genetik ditambah dengan ragam lingkungan. Dengan kata lain, heritabilitas merupakan proporsi besaran ragam genetik terhadap besaran ragam fenotipe untuk suatu karakter tertentu. Nilai heritabilitas sangat menentukan kemajuan genetik yang akan dicapai. Pada karakter yang mempunyai heritabilitas yang tinggi, seleksi dapat dilakukan pada generasi awal.
Karakter – karakter tanaman dapat berupa karakter kualitatif atau kuantitatif. Menurut Nasir (2001) karakter kualitatif merupakan wujud fenotipe yang saling berbeda tajam antara satu dengan yang lain secara kualitatif dan masing-masing dapat dikelompokkan dalam bentuk kategori. Karakter ini dikendalikan oleh sedikit gen. Sementara itu karakter kuantitatif dikendalikan oleh
2
Pola pewarisan masing-masing karakter diperlukan dalam menentukan strategi pemuliaan tanaman.
Tujuan
1. Mempelajari pewarisan sifat kualitatif dan kuantitatif pada beberapa karakter agronomi pada cabai.
2. Mempelajari tingkat heritabilitas beberapa karakter kuantitatif penting pada genotipe cabai.
Hipotesis
1. Terdapat beberapa karakter yang dikendalikan oleh gen mayor.
TINJAUAN PUSTAKA
Botani, Klasifikasi dan Syarat Tumbuh Tanaman Cabai
Cabai adalah tanaman tahunan tropika yang biasanya ditanam sebagai tanaman setahun. Jenis tanaman herba tersebut sebagian besar menjadi berkayu pada pangkal batangnya, dan beberapa jenis menjadi semak. Tanaman cabai merah tergolong dalam kingdom Plantae, divisi Magnoliophyta, kelas Magnoliopsida, ordo Solanes, famili Solanaceae, genus Capsicum dan spesies Capsicum annuum L. (Kusandriani, 1996). Terdapat sekitar 20-30 spesies yang termasuk ke dalam famili solanaceae. terdapat sekitar 20-30 spesies yang termasuk ke dalam genus Capsicum, termasuk diantaranya adalah lima spesies yang telah dibudidayakan, yaitu : C.baccatum, C. pubescens, C. annuum, C. chinense, dan C. frutescens (Greenleaf, 1986).
Menurut Kusandriani (1996) tanaman cabai berasal dari Meksiko. Tanaman tersebut termasuk komoditi penting dan banyak dibudidayakan di Meksiko dan negara-negara lain di dunia. Sebelum abad ke-15 spesies ini lebih banyak dikenal di Amerika Tengah dan Selatan, kemudian diintroduksi ke daratan Eropa tahun 1943. Setelah Colombus membawa dan menyebarkan cabai ke Eropa, cabai. menyebar cepat dari Eropa ke Asia dan Afrika.
Tanaman cabai termasuk tanaman dikotil berbentuk semak, batangnya berkayu, tipe percabangannya tegak atau menyebar (Kusandriani dan Permadi, 1996). Tanaman cabai memiliki sistem perakaran yang dangkal, diawali dengan akar tunggang (akar primer) kemudian tumbuh akar rambut ke samping (akar lateral/akar sekunder). Panjang akar primer berkisar 35-50 cm dan akar lateral sekitar 35-45 cm (Rubatzky dan Yamaguchi, 1997). Akar lateral cepat berkembang di dalam tanah dan menyebar pada kedalaman 10 -15 cm (Messiaen, 1992).
Bunga tanaman cabai umumnya bersifat tunggal dan tumbuh pada ujung ruas, serta merupakan bunga sempurna (hermaprodit). Bunga jantan dan betina terdapat pada satu bunga. Mahkota bunga berwarna putih atau ungu tergantung varietasnya, helaian mahkota bunga berjumlah lima atau enam helai. Pada dasar bunga terdapat daun buah berjumlah lima helai kadang-kadang bergerigi. Setiap bunga mempunyai satu putik (stigma), kepala putik berbentuk bulat. Terdapat
4
lima sampai delapan helai benang sari dengan kepala sari yang berbentuk lonjong, berwarna biru keunguan (Kusandriani, 1996).
Capsicum mempunyai buah yang bervariasi baik dari bentuk maupun
warna. Permukaan kulit dan warna buah cabai mempunyai variasi dari halus sampai bergelombang, mengkilat sampai kusam, hijau, kuning, coklat atau kadang-kadang ungu pada waktu muda dan menjadi merah waktu matang. Ukuran buah cabai beragam dari pendek sampai panjang. Panjang buah cabai berkisar antara 0.8-25 cm (Greenleaf, 1986).
Tanaman cabai dapat tumbuh pada berbagai jenis tanah, asalkan mempunyai drainase dan aerasi yang baik. Tanah yang paling ideal untuk tanaman cabai adalah yang mengandung bahan organik sekurang-kurangnya 1.5% dan mempunyai pH antara 6.0-6.5. Keadaan pH tanah sangat penting karena erat kaitannya dengan ketersediaan unsur hara. Apabila ditanam pada tanah yang mempunyai pH lebih dari 7, tanaman cabai akan menunjukan gejala klorosis, yakni tanaman kerdil dan daun menguning yang disebabkan kekurangan unsur hara kalsium (Ca) dan magnesium (Mg) atau keracunan alumunium (Al) dan mangan (Mn) (Sumarni, 1996).
Suhu udara optimal untuk pertumbuhan cabai adalah 18o – 27oC. Suhu udara yang paling cocok untuk pertumbuhan cabai rata-rata adalah 16oC pada
malam hari dan minimum 23oC pada siang hari. Bila suhu udara malam hari di bawah 16oC dan siang hari di atas 32oC, proses pembungaan dan pembuahan tanaman cabai akan gagal. Cabai merah tidak menghendaki curah hujan yang tinggi atau iklim yang basah, karena pada keadaan tersebut tanaman akan mudah terserang penyakit, terutama yang disebabkan cendawan. curah hujan yang sesuai untuk pertumbuhan tanaman cabai adalah 600-1.200 mm/tahun (Sumarni, 1996).
Pemuliaan Tanaman Cabai
Pemuliaan tanaman adalah suatu metode yang secara sistematik merakit keragaman genetik menjadi suatu bentuk yang bermanfaat bagi kehidupan manusia. Dalam pemuliaan tanaman diperlukan adanya keragaman genetik, sistem-sistem logis dalam pemindahan dan fiksasi gen, konsepsi dan tujuan atau sasaran yang jelas dan mekanisme penyebarluasan hasilnya kepada masyarakat (Makmur, 1992).
5
Tanaman cabai merupakan tanaman menyerbuk sendiri, tetapi penyerbukan silang terjadi karena serangga dan angin dengan persentase persilangan berkisar 7.6-36.8%.. Menurut Permadi dan Kusandriani (1996), diantara genotipe-genotipe cabai terdapat perbedaan dalam hal letak kepala putik terhadap kotak sari yang disebut heterostyle. Persilangan sering terjadi pada bunga yang memiliki tangkai putik (stilus) yang panjang dan kepala putik (stigma) lebih tinggi dari kotak sari. Penyerbukan sendiri terjadi pada bunga yang memiliki tangkai putik yang pendek, sehingga letak kepala putik lebih rendah daripada kepala sari. Bunga cabai pertama terbentuk pada umur 23-31 hari setelah tanam (HST) dan buah pertama mulai terbentuk pada umur 29-40 HST. Buah akan matang dalam waktu 45 hari setelah pembuahan (Greenleaf,1986). Struktur buah terdiri atas kulit, daging buah dan plasenta tempat melekatnya biji. daging buah umumnya renyah, tetapi kadang-kadang lunak, tergantung varietasnya. Biji cabai berwarna kuning jerami dan melekat di sepanjang plasenta, berjumlah sekitar 140 biji/buah (Kusandriani, 1996).
Menurut Kusandriani dan Permadi (1996) terdapat beberapa tujuan pemuliaan cabai, yaitu: (1) perbaikan daya hasil dan kualitas hasil; (2) perbaikan daya resistensi terhadap hama dan penyakit tertentu; (3) perbaikan sifat-sifat hortikultura; (4) perbaikan terhadap kemampuan mengatasi cekaman lingkungan.
Studi Pewarisan Beberapa Karakter Kualitatif dan Kuantitatif pada Cabai ( Capsicum annuum L.)
Menurut Nasir (2001) karakter kualitatif merupakan wujud fenotipe yang saling berbeda tajam antara satu dengan yang lain secara kualitatif dan masing-masing dapat dikelompokkan dalam bentuk kategori. Pada karakter kuantitatif umumnya dikendalikan oleh banyak gen dan merupakan hasil akhir dari suatu proses pertumbuhan dan perkembangan yang berkaitan langsung dengan karakter fisiologi dan morfologis, karakter morfologis lebih mudah diamati, misalnya produksi tanaman sering dijadikan obyek pemuliaan tanaman.
Ciri yang dapat digunakan untuk membedakan karakter kualitatif dan karakter kuantitatif menurut (Allard, 1960 dan Burns,1976) adalah sebagai berikut:
6
1). Pada karakter kualitatif terdapat ragam terputus pada kurva sebaran frekuensi dengan munculnya kembali ragam tetua di dalam generasi bersegregasi (F2, BC, F3), dan munculnya kembali salah satu ragam tetua bila terdapat
pengaruh dominansi penuh dalam generasi F1.
2). Pada karakter kuantitatif terdapat ragam kontinu pada kurva sebaran frekuensi di dalam generasi bersegrerasi (F2, BC, F3) dengan ragam F2 (VF2) yang
lebih besar dari ragam F1 (VF1).
Pada penelitian pewarisan suatu karakter, sering diperlukan analisis segregasi dari populasi yang bersegregasi (populasi F2). Dengan demikian analisis
statistik dan analisis genetik yang digunakan untuk melacak gen-gen pengendali karakter tersebut dapat dilakukan sesuai dengan persyaratan/asumsi : (1) tidak ada efek lingkungan, (2) tidak ada efek dominansi antar alel, (3) tidak ada efek epistasis, (4) gen memberikan efek yang sama dan bersifat aditif untuk semua lokus, (5) tidak ada pautan gen, dan (6) tetua dalam keadaan homozigositas lengkap, dan tanaman F1 dalam keadaan heterozigositas lengkap (Burns, 1976;
Poehlman, 1979).
Pada gen-gen yang mengikuti prinsip Mendel (disebut gen mayor) peranan ragam lingkungan relatif kecil dibandingkan peranan ragam gen-gen minor karena jumlah gen mayor umumnya tidak banyak dan peranan faktor lingkungan relatif kecil, maka ragam fenotipe yang ditampilkan dalam populasi bersegregasi sebagian besar merupakan ragam genetik, bersifat diskontinu dan merupakan akibat adanya efek dominan. Analisis genetik terhadap karakter yang dikendalikan oleh gen mayor, umumnya dilakukan dengan bantuan uji Chikuadrat (χ2)
(Strickberger, 1976).
Untuk menentukan apakah ragam pada karakter tersebut disebabkan oleh faktor genetik atau faktor lingkungan dilakukan pendugaan nilai heritabilitas. Heritabilitas sering juga dipakai sebagai tolok ukur kemajuan genetik yang dapat diharapkan dalam suatu program seleksi (Allard, 1960). Heritabilitas adalah proporsi keragaman teramati yang disebabkan oleh sifat menurun (Poespodarsono, 1988). Heritabilitas dapat juga diartikan rasio ragam genotipe terhadap ragam fenotipe (Sjamsudin, 1990). Nilai heritabilitas merupakan pernyataan kuantitatif peranan faktor keturunan dibanding faktor lingkungan dalam memberikan
7
pengaruh pada penampilan akhir atau sifat fenotipe yang bersangkutan (Poespodarsono, 1988).
Pendugaan heritabilitas arti sempit dilakukan dengan pendugaan nilai ragam lingkungan dengan mengikutsertakan satu set tanaman induk kedua tetua (P1 dan P2), F1 (P1 x P2), silang balik B1 (F1 x P1), silang balik B2 (F1 x P2) dan F2
(F1 x F1) (Warner, 1952). Studi pewarisan beberapa karakter kuantitatif pada cabai
menggunakan metode statistika yaitu setiap gen dan kombinasi dari beberapa gen diteliti secara menyeluruh (Khambanonda, 1948). Menurut Nasir (2001) heritabilitas dalam arti luas dapat dianggap sebagai suatu batas dugaan tertinggi dari heritabilitas dalam arti sempit. Oleh karena itu, selama heritabilitas dalam arti sempit dapat dihitung, nilai heritabilitas dalam arti luas tidak banyak digunakan, bila nilai heritabilitas arti sempit tinggi, maka metode seleksi yang paling tepat digunakan adalah seleksi massa sebaliknya apabila rendah sebaiknya digunakan seleksi silsilah, uji kekerabatan (sib-test), dan uji keturunan (progeny test).
Potensi rasio (hp) adalah peran atau aksi gen pengendali sifat informasi genetik yang terkait langsung dengan potensi sifat yang diwariskan oleh suatu tanaman. Menurut Petr dan Frey (1966) klasifikasi nilai potensi rasio 0<hp<1. termasuk ke dalam aksi gen dominan tidak sempurna. Menurut Crowder (1986) derajat dominansi tidak lengkap adalah kontribusi alel aktif A lebih besar daripada satu unit tertentu sehingga pengaruh dua alel aktif tidak sama dengan dua kali pengaruh dari alel aktif tunggal, yaitu AA tidak aditif; kombinasi Aa mendekati AA.
Menurut Petr dan Frey (1966) klasifikasi nilai potensi rasio hp > 1 atau hp < -1 termasuk ke dalam aksi gen over dominan. Menurut Crowder (1986) individu heterozigot memberi kontribusi pada fenotipe lebih besar daripada homozigot yang mempunyai alel aktif. Aa > AA, secara fisiologis alel-alel itu menghasilkan substansi berbeda yang komplementer. Istilah lewat dominansi umumnya digunakan dalam hubungannya dengan sifat-sifat fitness biologis seperti ukuran, produktivitas, dan daya hidup (viabilitas), persilangan antara individu dengan fitness kurang baik untuk sifat tertentu kadang-kadang menghasilkan keturunan yang lebih unggul (superior) dari kedua orang tuanya karena banyaknya gen yang terlibat sulit untuk menentukan hubungan dominansi dari gen-gen tertentu.
8
Menurut Crowder (1986) pewarisan di luar inti terjadi karena pewarisan partikel sitoplasma yang memiliki kelangsungan hidup. Pewarisan yang dikendalikan oleh gen di luar inti, disebut pewarisan ekstrakromosomal. Beberapa hal yang dapat dijadikan bukti bahwa suatu sifat diwariskan secara ekstrakromosomal adalah sebagai beikut:
1. Zuriat hasil persilangan berbeda dengan zuriat hasil persilangan resiprokalnya.
2. Sifat ditransmisikan melalui maternal.
3. Gen-gen tidak dapat dipetakan pada kromosom atau kelompok keterpautan tertentu.
4. Tidak terjadi segregasi untuk sifat tersebut. 5. Nisbah segregasi tidak mengikuti rasio Mendel.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan pada bulan Februari ampai dengan Juli 2007 di kebun percobaan IPB Leuwikopo, Darmaga, Bogor dan Laboratorium Genetika dan Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura. Tempat percobaan terletak 250 m dpl dengan jenis tanah Latosol.
Bahan dan Alat
Bahan tanaman yang digunakan adalah P1 (IPB C15), P2 (IPB C10), F1(15x10), F1R (10x15), BCP1 ((15x10)x15), BCP2 ((15x10)x10) dan F2. Pupuk yang digunakan adalah pupuk NPK Mutiara 16-16-16, pupuk kandang, pupuk Gandasil D dan Gandasil B. Pestisida yang digunakan adalah insektisida Curacron, Furadan 3G, akarisida Kelthane, fungisida Antracol, dan Dithane M-45. Bahan dan alat lainnya adalah Mulsa plastik hitam perak, ajir, meteran, timbangan analitik AND GF 3000, jangka sorong, tray, cangkul, kored, dan sprayer.
Metode Penelitian
Tanaman yang digunakan adalah P1: 25 tanaman, P2: 25 tanaman, F1 : 24 tanaman, F1R : 26 tanaman, BCP1 : 25 tanaman, BCP2 : 23 tanaman, dan F2 : 255 tanaman. Percobaan ini dilakukan tanpa rancangan percobaan tetapi setiap populasi (kecuali F2) diulang dua kali. Luas lahan yang digunakan adalah 200 m2, dengan 16 bedeng. Masing – masing bedeng berukuran 1 m x 5 m, dengan jarak tanam 50 cm x 50 cm.
Pelaksanaan Penelitian
Penyemaian dilakukan pada tray yang berisi media tanam (merk : media tanam plus). Media tanam disterilisasi terlebih dahulu pada suhu 150 OCselama tiga jam. Setelah benih disemai, dilakukan pemeliharaan antara lain penyiraman pada pagi atau sore hari. Pupuk diberikan dalam bentuk cair berupa campuran NPK Mutiara (10 g/l) dan fungisida Antracol (2 g/l) yang diberikan satu minggu sekali. Setelah berumur satu bulan, bibit dipindahkan ke lahan. Penanaman dilakukan pada delapan bedeng yang berukuran 1 m x 5 m. Satu minggu sebelum penanaman dibuat lubang tanam dengan jarak 50 cm x 50 cm dengan kedalaman lubang 30 cm, kemudian diberi pupuk kandang 1 kg dan kapur 0.5 kg per lubang
10 tanam. Bedengan ditutup dengan mulsa plastik hitam perak dan dibuat lubang sesuai jarak tanam.
Pengamatan
Pengamatan dilakukan pada karakter kualitatif dan kuantitatif. Peubah yang diamati pada karakter kualitatif adalah :
1. Permukaan buah (licin, agak kasar, kasar ) 2. Lekukan di pangkal buah
3. Orientasi Buah (samping, bawah, atas) 4. Penyempitan tangkai buah
0 1
5. Warna Daun ( hijau, hijau tua)
Peubah pada karakter kuantitatif yang diamati adalah :
1. Tinggi dikotomus (cm), diukur dari pangkal batang sampai cabang dikotomus setelah panen kedua.
2. Diameter buah (cm), diambil 5 buah setiap tanaman, diukur pada bagian pangkal, tengah, dan ujung dengan menggunakan jangka sorong. Penentuan bagian tengah, yaitu setengah dari panjang buah dengan ukuran relatif sama. Buah yang digunakan adalah buah panen pertama.
3. Panjang buah (cm), diambil 5 buah setiap tanaman dan diukur dari pangkal buah sampai dengan ujung buah pada saat panen pertama. 4. Bobot buah layak pasar (g), ditimbang dari bobot buah yang tidak
terserang hama dan penyakit serta mempunyai penampilan normal. Keterangan : 0 Tidak ada 1 Ada Keterangan : 0 Tidak ada 1 Ada
11 6. Posisi Bunga
7. Bobot buah total (g), ditimbang dari bobot buah total yang mempunyai penampilan normal.
Analisis Data
1). Untuk menguji kesesuaian perbandingan Mendel (karakter yang dikendalikan oleh gen mayor), digunakan rumus sebagai berikut :
χ2 =
‡”
[
(
oi-ei2/e)
]
Keterangan :
oi = nilai pengamatan populasi ke-i
ei = nilai harapan populasi ke –i
2). Heritabilitas digunakan untuk menentukan apakah ragam pada karakter tersebut disebabkan oleh faktor genetik atau oleh faktor lingkungan. Menurut Allard (1960) heritabilitas sering juga dipakai sebagai tolok ukur kemajuan genetik yang dapat diharapkan dalam suatu program seleksi. Rumus heritabilitas arti luas dan sempit adalah :
h2 NS = 2VF2- (VBCP1+VBCP2) VF2 Keterangan : h2
NS = heritabilitas arti sempit
h2BS = heritabilitas arti luas
VBCP1 = ragam populasi BCP1 VBCP2 = ragam populasi BCP2 VP1 = ragam populasi P1 VP2 = ragam populasi P2 VF2 = ragam populasi F2 h2 BS = V F2- (VF1+VP1+VP2 )/3 VF2 Keterangan : 3 Tidak tegak 5 Semi Tegak 7 Tegak
12 Tabel 1. Nisbah Fenotipik Sifat Resistensi yang dikendalikan oleh Gen Mayor
pada Populasi F2 (Burn,1976; Griffiths et al.,1996) Tipe Resistensi Resisten
(R) Resisten sedang (MR) Rentan sedang (MS) Rentan (S) 1. Resistensi dikendalikan pasang gen a. Dominan penuh 3 - - 1 b. Resesif 1 - - 3 2. Resistensi dikendalikan pasang gen
a. Dominan penuh pada
kedua lokus A dan B 9 3 3 1
b. Resesif epistasis Aa epistasis terhadap B dan b
9 3 - 4 c. Dominan epistasis A
epistasis terhadap B dan b
12 - 3 1 d. Dominan dan resesif
epistasis A epistasis terhadap B dan b; bb epistasis terhadap A dan a
13 - - 3
e. Duplikat resesif epistasis Aa epistasis ke B dan b; bb epistasis ke A dan aa 9 - - 7 f. Duplikat dominan epistasis A epistasis ke B dan b; B epistasis ke A dan a 15 - - 1 g. Interaksi duplikat 9 6 - 1 h. Interaksi komplementer 9 - - 7 i. Interaksi kompleks 10 3 - 3 3. Resistensi dikendalikan 3 pasang gen Interaksi epistasis : A 37 - - 27 B 45 - - 19 C 55 - - 9 3). Derajat dominansi menurut Petr dan Frey (1966) dihitung menggunakan
rumus : hp =F1-MP HP-MP Keterangan : hp = derajat dominansi F1 = rata-rata nilai F1
HP = rata-rata nilai tetua tertinggi MP = nilai tengah kedua tetua
HASIL DAN PEMBAHASAN
Kondisi Umum Pertanaman
Pada saat pindah tanam yaitu setelah umur bibit lebih kurang satu bulan, kondisi lahan cukup air dan lembab. Kondisi tersebut menyebabkan bibit banyak yang terserang penyakit rebah kecambah (Phytium debaryanum). Penyakit tersebut menimbulkan gejala busuk pada batang bawah atau pada pangkal akar.
Hama yang menyerang tanaman antara lain jangkrik, belalang, trips, kutu daun, dan tungau. Sementara itu patogen yang menyerang selain rebah kecambah antara lain bakteri, antraknosa dan virus gemini yang mengakibatkan daun-daun menjadi kuning.
Pola Pewarisan Sifat Kualitatif
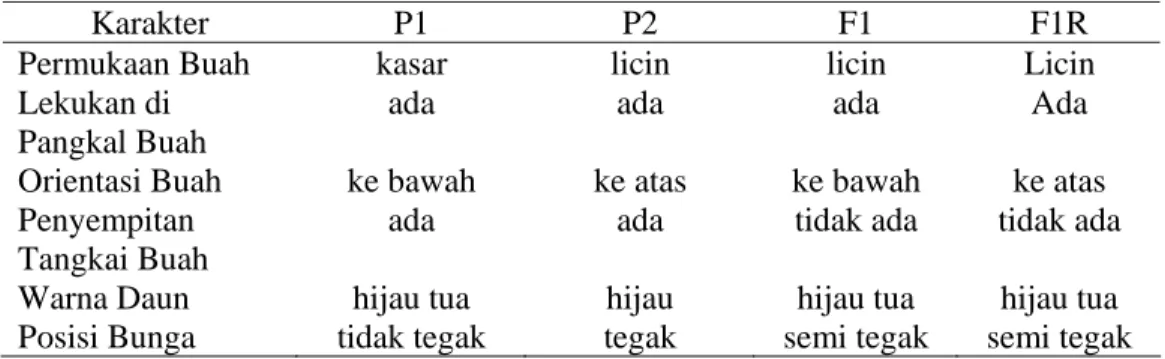
Studi pola pewarisan dilakukan dengan cara mengkelaskan sifat yang diamati pada populasi F2. Nisbah dari kelas-kelas yang didapat dari hasil pengamatan tersebut diuji apakah berbeda nyata atau tidak berbeda nyata dengan nisbah fenotipik gen mayor berdasarkan Tabel 1. Tabel 2 menunjukkan data karakter kualitatif pada populasi P1, P2, F1, dan F1R tanaman cabai yang diuji.
Tabel 2. Beberapa Karakter Kualitatif Cabai pada P1, P2, F1, dan F1R
Karakter P1 P2 F1 F1R
Permukaan Buah kasar licin licin Licin
Lekukan di Pangkal Buah
ada ada ada Ada
Orientasi Buah ke bawah ke atas ke bawah ke atas Penyempitan
Tangkai Buah
ada ada tidak ada tidak ada
Warna Daun hijau tua hijau hijau tua hijau tua Posisi Bunga tidak tegak tegak semi tegak semi tegak
14
Tabel 3. Nisbah Fenotipik Karakter Kualitatif Cabai pada Populasi BCP1, BCP2, dan F2
Karakter BCP1 BCP2 F2
Permukaan Buah
16 kasar : 9 licin 15 kasar : 8 licin 167 kasar :71 licin
Lekukan di Pangkal Buah
20 ada : 5 tidak ada 15 ada : 8 tidak ada 181 ada : 57 tidak ada
Orientasi Buah
18 ke bawah : 7 ke atas 13 ke bawah : 10 ke atas
169 ke bawah : 69 ke atas
Penyempitan Tangkai Buah
14 ada : 11 tidak ada 7 ada : 16 tidak ada 133 ada : 105 tidak ada
Warna Daun 17 hijau : 8 hijau tua 15 hijau : 8 hijau tua
130 hijau : 108 hijau tua
Posisi Bunga 16 tidak tegak : 7 semi tegak : 2 tegak 11 tidak tegak : 4 semi tegak : 8 tegak 140 tidak tegak : 42 semi tegak : 56 tegak
Nisbah fenotipik frekuensi pola pewarisan yang dikendalikan oleh gen mayor dalam populasi BCP1, BCP2, dan populasi F2 dapat dilihat pada Tabel 3. Hasil uji khi kuadrat untuk karakter permukaan buah, lekukan pada pangkal buah, orientasi buah, penyempitan tangkai buah, warna daun, dan posisi bunga pada populasi F2, disajikan pada Tabel 4.
Tabel 4. Hasil Uji Khi Kuadrat Karakter Kualitatif Cabai Populasi F2
Karakter Fenotipe F2
Nisbah
segregasi X hitung X tabel 5%
Permukaan Buah 167 kasar : 71
licin 3 : 1 2.96 tn 3.84 Lekukan di Pangkal Buah 181 ada : 57 tidak ada 3 : 1 0.14tn 3.84
Orientasi Buah 169 bawah : 69
atas 3 : 1 2.02tn 3.84
Penyempitan Tangkai Buah
133 ada : 105
tidak ada 9 : 7 0.01tn 3.84 Warna Daun 130 hijau : 108
hijau tua 9 : 7 0.26tn 3.84 Posisi Bunga 140 tidak tegak : 42 semi tegak : 56 tegak 9 : 3 : 4 0.64tn 5.99
15
Permukaan Buah
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter permukaan buah adalah 3 kasar : 1 licin (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter permukaan buah dikendalikan oleh satu gen dua alel. Karakter permukaan buah kasar diduga mempunyai genotipe KK dan Kk, sedangkan karakter permukaan buah licin mempunyai genotipe kk. Alel K diduga dominan terhadap alel k.
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur homozigot P1 (kasar) disilangkan dengan galur homozigot P2 (licin) akan menghasilkan F1 (kasar). Hasil ini tidak sesuai dengan data pengamatan di lapangan yaitu populasi F1 mempunyai karakter permukaan buah licin (Tabel 2). Ketidaksesuaian ini diduga karena tanaman P1 belum homozigot. Tanaman P1 yang demikian (bergenotipe Kk), jika disilangkan dengan tanaman P2 (homozigot kk) menghasilkan populasi F1 bergenotipe Kk dan kk. Tanaman F1 yang bergenotipe Kk jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 3 K- : 1 kk (Gambar 1). Tetua P1 x P2 Kk kk (kasar) (licin) F1 Kk & kk kasar licin Kk (kasar) F2 1 KK : 2 Kk : 1 kk 3 kasar : 1 licin
Gambar 1. Bagan Persilangan antara P1 (Kasar) & P2 (Licin) serta Selfing F1 untuk Karakter Permukaan Buah
Untuk mendukung dugaan genotipe pada tanaman P1, P2, dan F1 dilakukan persilangan antara F1 dan P1 (BCP1) serta F1 dan P2 (BCP2). Hasil yang diperoleh pada populasi BCP1 dan BCP2 berturut-turut menunjukkan perbandingan 16 kasar : 9 licin dan 15 kasar : 8 licin. Berdasarkan perbandingan tersebut diduga genotipe F1, P1, dan P2 berturut-turut adalah Kk, Kk, dan kk. Persilangan anatara F1 (Kk, fenotipe kasar)
16
dengan P1 (Kk, fenotipe kasar) menghasilkan populasi BCP1 dengan fenotipe kasar dan licin, demikian juga dengan persilangan F1 dan P2 menghasilkan populasi BCP2 dengan fenotipe kasar dan licin (Gambar 2).
F1 x P1 F1 x P2
Kk Kk Kk kk
(kasar) (kasar) (kasar) (licin)
BCP1 KK, Kk, dan kk BCP2 Kk & kk kasar licin kasar licin
Gambar 2. Bagan Persilangan antara F1 dengan P1 dan F1 dengan P2 untuk Karakter Permukaan Buah
Lekukan di Pangkal Buah
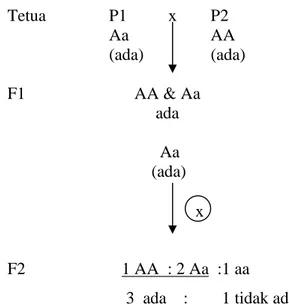
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter lekukan di pangkal buah adalah 3 ada : 1 tidak ada (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter lekukan di pangkal buah dikendalikan oleh satu gen dua alel. Karakter ada lekukan di pangkal buah diduga mempunyai genotipe AA dan Aa, sedangkan karakter tidak ada lekukan buah mempunyai genotipe aa. Alel A diduga dominan terhadap alel a.
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur P1 (ada) disilangkan dengan galur P2 (tidak ada) akan menghasilkan F1 (ada). Hasil ini tidak sesuai dengan data pengamatan di lapang yaitu populasi P2 mempunyai karakter ada lekukan di pangkal buah (Tabel 2). Salah satu dari galur P1 dan P2 diduga mempunyai genotipe Aa atau AA. Jika galur P1 mempunyai genotipe AA maka galur P2 mempunyai genotipe Aa, demikian juga sebaliknya. Tanaman F1 yang bergenotipe Aa jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 3 A- : 1 aa (Gambar 3).
17 Tetua P1 x P2 Aa AA (ada) (ada) F1 AA & Aa ada Aa (ada) F2 1 AA : 2 Aa :1 aa 3 ada : 1 tidak ada
Gambar 3. Bagan Persilangan antara P1 (Ada) & P2 (Ada) serta Selfing F1 untuk Karakter Lekukan di Pangkal Buah
Untuk mendukung dugaan genotipe pada tanaman P1, P2, dan F1 dilakukan persilangan antara F1 dan P1 (BCP1) serta F1 dan P2 (BCP2). Hasil yang diperoleh pada populasi BCP1 dan BCP2 berturut-turut menunjukkan perbandingan 20 ada : 5 tidak ada dan 15 ada : 8 tidak ada. Berdasarkan perbandingan tersebut diduga genotipe F1, P1, dan P2 berturut-turut adalah Aa, Aa, sedangkan P2 diduga mempunyai genotipe Aa. Tanaman ini berbeda dengan tanaman P2 untuk membentuk populasi F1. Persilangan antara F1 (Aa, fenotipe ada) dengan P1 (Aa, fenotipe ada) menghasilkan populasi BCP1 dengan fenotipe ada dan tidak ada, demikian juga dengan persilangan F1 dan P2 menghasilkan populasi BCP2 dengan fenotipe ada dan tidak ada (Gambar 4).
F1 x P1 F1 x P2
Aa Aa Aa Aa
(ada) (ada) (ada) ( ada)
BCP1 AA , Aa & aa BCP2 AA, Aa, & aa ada tidak ada ada tidak ada
Gambar 4. Bagan Persilangan antara F1 dan P1 serta F1 dan P2 untuk Karakter Lekukan di Pangkal Buah
18
Orientasi Buah
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter orientasi buah adalah 3 ke bawah : 1 ke atas (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter orientasi buah dikendalikan oleh satu gen dua alel. Karakter orientasi buah ke bawah diduga mempunyai genotipe BB dan Bb, sedangkan karakter orientasi buah ke atas mempunyai genotipe bb. Alel B diduga dominan terhadap alel b.
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur P1 (ke bawah) disilangkan dengan galur P2 (ke atas) akan menghasilkan F1 (ke bawah). Hasil ini sesuai dengan data pengamatan di lapang yaitu populasi F1 mempunyai karakter permukaan buah ke bawah (Tabel 2). Tanaman F1 yang bergenotipe Bb jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 3 B- : 1 bb (Gambar 5).
Tetua P1 x P2
BB bb
(ke bawah) (ke atas)
F1 Bb ke bawah F2 1 BB : 2 Bb :1 bb 3 ke bawah : 1 ke atas
Gambar 5. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Orientasi Buah
Untuk mendukung dugaan genotipe pada tanaman P1, P2, dan F1 dilakukan persilangan antara F1 dan P1 (BCP1) serta F1 dan P2 (BCP2). Persilangan antara F1 heterozigot dengan P1 homozigot dominan menghasilkan BCP1 yang mempunyai fenotipe orientasi buah ke bawah. Sementara itu persilangan antara F1 heterozigot dengan P2 homozigot resesif akan menghasilkan BCP2 yang mempunyai fenotipe orientasi buah ke bawah dan ke atas dengan perbandingan 1:1. Hasil ini tidak sesuai dengan data pengamatan di lapang pada populasi BCP1 dan BCP2 berturut-turut menunjukkan perbandingan 18 bawah : 7 atas dan 13 bawah : 10 atas. Berdasarkan perbandingan tersebut diduga genotipe F1, P1, dan P2 berturut-turut adalah Bb, Bb, dan bb. Persilangan antara F1 (Bb, fenotipe bawah) dengan P1 (Bb, fenotipe bawah)
19
menghasilkan populasi BCP1 dengan fenotipe bawah dan atas, demikian juga dengan persilangan F1 dan P2 menghasilkan populasi BCP2 dengan fenotipe bawah dan atas (Gambar 6). Tanaman P1 yang digunakan untuk membentuk populasi F1 berbeda dengan tanaman P1 untuk membentuk populasi BCP1. Dengan demkian galur P1 belum homozigot.
F1 x P1 F1 x P2
Bb Bb Bb bb
(ke bawah) (ke bawah) (ke bawah) (ke atas)
BCP1 BB, Bb & bb BCP2 Bb & bb ke bawah ke atas ke bawah ke atas
Gambar 6. Bagan Persilangan antara F1 dan P1 serta F1 dan P2 untuk Karakter Orientasi Buah
Penyempitan Tangkai Buah
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter penyempitan buah adalah 133 ada : 105 tidak ada atau 9:7 (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter penyempitan tangkai buah dikendalikan oleh dua pasang gen atau epistasis. Menurut Crowder (1986) perbandingan 9:7 pada populasi F2 menunjukkan interaksi epistasis resesif ganda. Hal ini berarti fenotipe yang sama dihasilkan oleh kedua genotipe homozigot resesif. Dua gen resesif bersifat epistasis terhadap alel dominan.
Karakter ada penyempitan tangkai buah diduga mempunyai genotipe A-T-, sedangkan karakter tidak ada penyempitan tangkai buah mempunyai genotipe A-tt, aaT-, dan aatt. Setiap ada alel A yang bersama – sama dengan T maka fenotipe yang muncul adalah ada penyempitan tangkai buah. Sementara itu, jika alel genotipe homozigot aa atau tt maka fenotipe yang muncul adalah tidak ada penyempitan tangkai buah (Gambar 7).
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur homozigot P1 (ada) disilangkan dengan galur homozigot P2 (tidak ada) akan menghasilkan F1 (ada). Hasil ini tidak sesuai dengan data pengamatan di lapang yaitu populasi P2 mempunyai karakter ada penyempitan tangkai buah sedangkan populasi F1 mempunyai karakter tidak ada penyempitan tangkai buah (Tabel 2). Ketidaksesuaian ini diduga karena tanaman P1 dan P2 belum homozigot. Tanaman P1 yang demikian (bergenotipe AaTt), jika disilangkan dengan tanaman P2 (bergenotipe AaTT)
20
menghasilkan populasi F1 bergenotipe AATT, AaTt, AATt, AaTT, aaTT, dan aaTt. Tanaman F1 yang bergenotipe AaTt jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 9 A-T- : 3 A-tt : 3 aaT- : 1 aatt (Gambar 7).
Tetua P1 x P2
AaTt AaTT
(ada) (ada)
F1 AATT, AaTt , AATt , AaTT, aaTT, aaTt ada tidak ada
AaTt
(ada)
F2 9 A-T : 3 A-tt : 3 aaT- : 1 aatt 9 ada : 7 tidak ada
Gambar 7. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Penyempitan Tangkai Buah
Warna Daun
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter warna daun adalah 130 hijau : 108 hijau tua atau 9:7 (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter warna daun dikendalikan oleh dua pasang gen atau epistasis. Menurut Crowder (1986) perbandingan 9:7 pada populasi F2 menunjukkan interaksi epistasis resesif ganda. Hal ini berarti fenotipe yang sama dihasilkan oleh kedua genotipe homozigot resesif. Dua gen resesif bersifat epistasis terhadap alel dominan.
Karakter warna daun hijau diduga mempunyai genotipe H-T-, sedangkan karakter warna daun hijau tua mempunyai genotipe H-tt, hhT-, dan hhtt. Setiap ada alel A yang bersama – sama dengan T maka fenotipe yang muncul adalah warna daun hijau. Sementara itu, jika genotipe homozigot hh atau tt maka fenotipe yang muncul adalah warna daun hijau tua (Gambar 8).
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur homozigot P1 (hijau tua) disilangkan dengan galur homozigot P2 (hijau) akan menghasilkan F1 (hijau). Hasil ini tidak sesuai dengan data pengamatan di lapang yaitu
21
populasi F1 mempunyai karakter warna daun hijau tua (Tabel 2). Ketidaksesuaian ini diduga karena tanaman P1 dan P2 belum homozigot. Tanaman P1 bergenotipe Hhtt, jika disilangkan dengan tanaman P2 bergenotipe HhTT menghasilkan populasi F1 bergenotipe HHTt, HhTt, dan hhTt. Tanaman F1 yang bergenotipe HhTt jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 9 T- : 3 H-tt : 3 hhT- : 1 hhH-tt (Gambar 8).
Tetua P1 x P2
Hhtt HhTT
(hijau tua) (hijau)
F1 HHTt, HhTt, hhTt hijau hijau tua HhTt (hijau) F2 9 H-T : 3 H-tt : 3 hhT- : 1 hhtt
9 hijau : 7 hijau tua
Gambar 8. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Warna Daun
Posisi Bunga
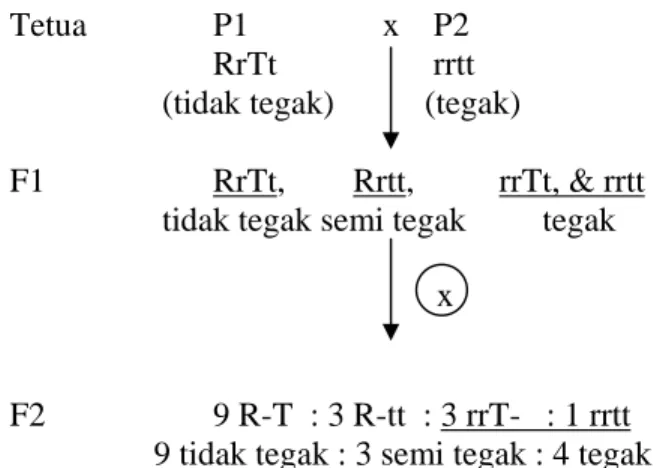
Berdasarkan data fenotipik pada populasi F2 diperoleh nisbah karakter posisi bunga adalah 140 tidak tegak : 42 semi tegak : 56 tegak atau 9: 3 : 4 (Tabel 4). Nisbah tersebut mengindikasikan bahwa karakter posisi bunga dikendalikan oleh dua pasang gen atau epistasis. Menurut Crowder (1986) perbandingan tersebut menandakan adanya interaksi epistasi resesif.
Karakter posisi bunga tidak tegak diduga mempunyai genotipe R-T-, karakter semi tegak mempunyai genotipe R-tt, dan rrT-, sedangkan genotipe karakter tegak adalah hhtt. Setiap ada alel R yang bersama – sama dengan T maka fenotipe yang muncul adalah posisi bunga tidak tegak. Sementara itu, jika ada genotipe R-tt atau rrT- maka fenotipe yang muncul adalah semi tegak, sedangkan apabila genotipe rrtt fenotipe yang muncul adalah tegak (Gambar 9).
Populasi F2 merupakan generasi kedua hasil persilangan dua galur homozigot. Galur homozigot P1 (tidak tegak) disilangkan dengan galur homozigot P2 (tegak) akan
22
menghasilkan F1 (tidak tegak). Hasil ini tidak sesuai dengan data pengamatan di lapang yaitu populasi F1 mempunyai karakter posisi bunga semi tegak (Tabel 2). Ketidaksesuaian ini diduga karena tanaman P1 dan P2 belum homozigot. Tanaman P1 bergenotipe RrTt jika disilangkan dengan tanaman P2 bergenotipe rrtt menghasilkan populasi F1 bergenotipe RrTt, Rrtt, rrTt dan rrtt. Tanaman F1 yang bergenotipe RrTt jika dilakukan penyerbukan sendiri akan menghasilkan populasi F2 dengan perbandingan 9 R-T : 3 R-tt : 3 rrT- : 1 rrtt (Gambar 9).
Tetua P1 x P2 RrTt rrtt (tidak tegak) (tegak)
F1 RrTt, Rrtt, rrTt, & rrtt tidak tegak semi tegak tegak
F2 9 R-T : 3 R-tt : 3 rrT- : 1 rrtt 9 tidak tegak : 3 semi tegak : 4 tegak
Gambar 9. Bagan Persilangan antara P1 & P2 serta Selfing F1 untuk Karakter Posisi Bunga
Karakter Kuantitatif
Karakter kuantitatif dikendalikan oleh banyak gen minor. Menurut Nasir (2001) karakter kuantitatif umumnya dikendalikan oleh banyak gen dan merupakan hasil akhir dari suatu proses pertumbuhan dan perkembangan yang berkaitan langsung dengan karakter fisiologi dan morfologi. Pada Tabel 5 dapat dilihat hasil uji normalitas karakter kuantitatif pada populasi F2.
Tabel 5. Hasil Uji Normalitas Karakter Kuantitatif Cabai pada Populasi F2 Karakter Normalitas Sebaran Bobot Layak Pasar < 0.01 tidak normal
Bobot Total Buah < 0.01 tidak normal
Diameter Buah < 0.01 tidak normal
Tinggi dikotomus > 0.1 normal
Panjang Buah < 0.01 tidak normal
Pada karakter bobot layak pasar, bobot total, diameter dan panjang buah menunjukkan sebaran yang tidak normal (Tabel 5). Menurut Murti et al.(2004)
23
parameter yang tidak mengikuti distribusi normal memiliki sebaran yang diskontinu, terdapat pengaruh gen mayor, sifat kelas yang dapat dibedakan dengan jelas karena dipengaruhi oleh beberapa gen.
Pada karakter tinggi dikotomus menunjukkan sebaran normal (Tabel 5). Menurut Crowder (1986) ragam kontinu fenotipe membentuk spektrum, nilai populasi cukup besar sering membentuk kurva normal menunjukkan karakter kuantitatif. Tabel 6 menunjukkan ragam tetua, ragam backcross, ragam F1, ragam F2, dan nilai heritabilitas karakter kuantitatif yang diteliti.
Tabel 6. Ragam Tetua, Ragam Backcross, Ragam F1, Ragam F2 dan Nilai Heritabilitas Karakter Kuantitatif
Menurut Nasir (2001) heritabilitas adalah proporsi ragam genetik terhadap besaran total ragam genetik ditambah dengan ragam lingkungan, dengan kata lain heritabilitas merupakan proporsi besaran ragam genetik terhadap besaran ragam fenotipe untuk suatu karakter tertentu. Ada dua nilai heritabilitas yang dikenal dalam pemuliaan tanaman yaitu heritabilitas dalam arti luas dan heritabilitas dalam arti sempit.
Pada karakter tinggi dikotomus dan diameter buah nilai heritabilitas arti luas lebih besar daripada nilai heritabilitas arti sempit. Menurut Nasir (2001) nilai heritabilitas dalam arti luas memperhatikan ragam genetik total dalam kaitannya dengan keragaman fenotipe. Dalam hal ini genotipe dianggap sebagai unit dalam kaitannya dengan lingkungan. Sementara itu heritabilitas dalam arti sempit yang menjadi fokus perhatian adalah keragaman yang diakibatkan oleh peran gen aditif merupakan bagian dari keragaman genetik total. Berdasarkan penjelasan ini dapat dipahami bahwa nilai Komponen Ragam Bobot Layak Pasar Bobot Total Buah Diameter Buah Tinggi Dikotomus Panjang Buah VP1 1058.34 5514.07 0.07 3.86 1.35 VP2 604.14 963.60 0.01 34.90 1.01 VSB1 648.93 2324.64 0.04 23.40 0.91 VSB2 589.21 1759.10 0.05 27.02 1.06 VF1 1356.55 2745.42 0.02 13.97 1.00 VF2 1349.68 2450.22 0.04 32.49 1.67 h2ns 1.08 0.33 -0.25 0.45 0.82 h2bs 0.25 -0.25 0.17 0.47 0.33
24
heritabilitas dalam arti sempit tidak akan pernah lebih besar dibandingkan dengan nilai heritabilitas dalam arti luas untuk suatu karakter tertentu.
Pada Tabel 6 dapat dilihat karakter bobot layak pasar, bobot total buah, dan panjang buah mempunyai nilai heritabilitas arti sempit lebih besar daripada nilai heritabilitas arti luas. Hasil ini tidak sesuai dengan pendapat Nasir (2001). Terjadinya perbedaan ini diakibatkan kurangnya populasi backcross. Pada karakter diameter dan bobot total nilai heritabilitas minus. Menurut Allard (1960) nilai heritabilitas minus dapat dianggap nol.
Derajat Dominansi
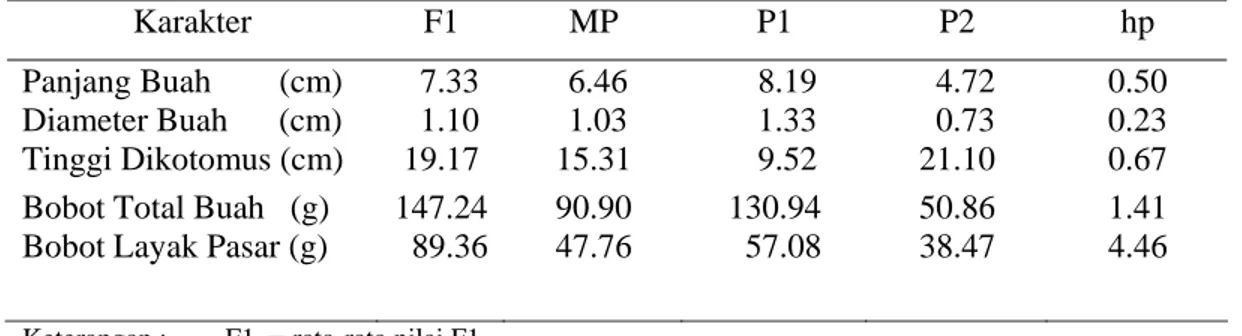
Derajat dominansi (hp) adalah peran atau aksi gen pengendali sifat genetik yang terkait langsung dengan potensi sifat yang diwariskan oleh suatu tanaman. Pada Tabel 7 disajikan nilai tengah dan aksi gen pada karakter kuantitatif yang diuji.
Tabel 7. Nilai Tengah Karakter Kuantitatif Cabai dan Aksi Gen
Karakter F1 MP P1 P2 hp
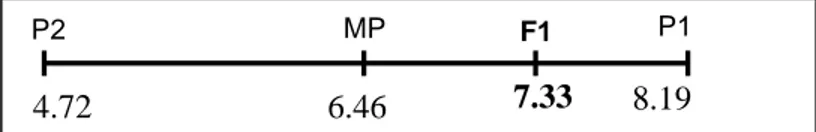
Panjang Buah (cm) 7.33 6.46 8.19 4.72 0.50 Diameter Buah (cm) 1.10 1.03 1.33 0.73 0.23 Tinggi Dikotomus (cm) 19.17 15.31 9.52 21.10 0.67 Bobot Total Buah (g) 147.24 90.90 130.94 50.86 1.41 Bobot Layak Pasar (g) 89.36 47.76 57.08 38.47 4.46
Keterangan : F1 = rata-rata nilai F1 P1 = rata-rata nilai P1 P2 = rata-rata nilai P2 MP = nilai tengah kedua tetua hp = derajat dominansi
Nilai derajat dominansi panjang buah, diameter, dan tinggi dikotomus menunjukkan angka 0.5, 0.23, 0.67 (Tabel 7). Hal ini berarti bahwa karakter panjang buah, diameter, dan tinggi dikotomus termasuk ke dalam aksi gen dominan tidak sempurna. Menurut Petr dan Frey (1966) klasifikasi nilai derajat dominansi adalah 0<hp<1. Sehingga panjang buah, diameter, dan tinggi dikotomus termasuk ke dalam aksi gen tidak sempurna. Menurut Crowder (1986) derajat dominansi tidak lengkap adalah kontribusi alel aktif A lebih besar daripada satu unit tertentu sehingga pengaruh dua alel aktif tidak sama dengan dua kali pengaruh dari alel aktif tunggal, yaitu AA tidak aditif; kombinasi Aa mendekati AA.
25
Nilai derajat dominansi bobot total dan bobot layak pasar buah menunjukkan angka 1.41 dan 4.46. Hal ini berarti karakter berat total dan berat layak pasar buah termasuk ke dalam aksi gen over dominan. Menurut Petr dan Frey (1966) klasifikasi nilai derajat dominansi hp > 1 atau hp < -1 termasuk ke dalam aksi gen over dominan. Menurut Crowder (1986) individu heterozigot memberi kontribusi pada fenotipe lebih besar daripada homozigot yang mempunyai alel aktif. Aa > AA, secara fisiologis alel-alel itu menghasilkan substansi berbeda yang komplementer istilah lewat dominansi umumnya digunakan dalam hubungannya dengan sifat-sifat fitness biologis seperti ukuran, produktivitas, dan daya hidup (viabilitas), persilangan antara individu dengan
fitness kurang baik untuk sifat tertentu kadang-kadang menghasilkan keturunan yang
lebih unggul (superior) dari kedua orang tuanya karena banyaknya gen yang terlibat sulit untuk menentukan hubungan dominansi dari gen-gen tertentu.
Pewarisan Ekstrakromosomal
Data kuantitatif pewarisan ekstrakromosomal dapat dilihat pada F1 dan F1R melalui uji t. Nilai t hitung dua genotipe F1 dan F1R dapat dilihat pada Tabel 8.
Tabel 8. Nilai t Hitung pada Karakter Kuantitatif Cabai Populasi F1 dan F1R
Sifat Genotipe T hit P Hit
F1 F1R
Panjang Buah (cm) 7.33 ± 1.00 6.56 ± 0.41 3.12** 0.005 Diameter Buah (cm) 1.10 ± 0.13 0.78 ± 0.09 8.35** 0.000 Tinggi Dikotomus (cm) 19.17 ± 3.74 14.00 ± 5.22 4.24** 0.000 Bobot Total Buah (g) 130.70 ± 55.40 58.20 ± 36.90 4.29** 0.001 Bobot Layak Pasar (g) 87.80 ± 39.10 30.50 ± 34.60 4.22** 0.000 Keterangan : ** = berbeda sangat nyata
Berdasarkan analisis data dengan menggunakan uji t dapat diketahui bahwa F1 dan F1R berbeda sangat nyata untuk karakter panjang buah, diameter buah, bobot total, bobot layak panen, dan tinggi dikotomus (Tabel 8). Hal ini menunjukkan bahwa terdapat perbedaan nilai antara populasi tanaman F1 dan F1 resiprokalnya sehingga karakter-karakter tersebut disebabkan oleh pewarisan ekstrakromosomal. Menurut Crowder (1986) pewarisan di luar inti terjadi karena pewarisan partikel sitoplasma yang memiliki kelangsungan hidup. Pewarisan yang dikendalikan oleh gen di luar inti disebut pewarisan ekstrakromosomal.
Pada data kualitatif pengaruh pewarisan ekstrakromosomal tidak dianalisis dengan uji t, tetapi hanya secara visual. Pengamatan visual menunjukkan terdapat
26
perbedaan orientasi buah pada populasi F1 & F1R. Tanaman F1 menunjukkan arah ke bawah dan F1R ke atas Hal ini menunjukkan bahwa karakter tersebut dipengaruhi oleh pewarisan ektrakromosomal (Tabel 2).
KESIMPULAN DAN SARAN
KesimpulanPada sifat permukaan buah, lekukan di pangkal buah, orientasi buah, penyempitan tangkai buah, warna daun, dan posisi bunga dikendalikan oleh gen mayor. Pada sifat permukaan buah, lekukan di pangkal buah, dan orientasi buah dikendalikan oleh satu pasang gen dominan penuh. Pada sifat posisi bunga dikendalikan oleh 2 pasang gen dengan interaksi epistasis resesif. Penyempitan tangkai buah dan warna daun dikendalikan oleh 2 pasang gen dengan interaksi epistasis resesif ganda.
Tinggi dikotomus menunjukkan sebaran normal yang mengindikasikan bahwa karakter tersebut dikendalikan oleh gen minor. Nilai heritabilitas arti sempit dan luas untuk karakter tinggi dikotomus adalah 0.47 dan 0.45.
Terdapat pewarisan ektrakromosomal pada karakter orientasi buah. Semua karakter kuantitatif yang diamati dipengaruhi oleh pewarisan ekstrakromosomal.
Saran
Pada karakter bobot layak pasar, bobot total, diameter dan panjang buah sebaiknya digunakan pengkelasan karena data menyebar tidak normal. Pada karakter bobot layak pasar, bobot total, diameter, dan panjang buah menunjukkan nilai heritabilitas arti sempit lebih besar daripada nilai heritabilitas arti luas sehingga sebaiknya populasi backcross ditambah.
DAFTAR PUSTAKA
Agfi, J. 2006. Aneka cabe atasi penggumpalan darah. http://www.mail- archieve.com/[email protected]/msg2929.html. (16 Maret 2006).
Allard, R. W. 1960. Principles of Plant Breeding. John Wiley & Sons. New York. 485 p.
Burns, G. W. 1976. The Science of Genetics: An Introduction to Heredity. 3rdedition. Macmillan Publ. Co. New York. 564 p.
Chandraratna, M. F. 1964. Genetics and Breeding of Rice : Genetics of Metric and Physiological Characters. Longmans, Green and Co Ltp. London. p.49-80.
Crowder, L.V. 1986. Genetika Tumbuhan. Edisi (Revisi ke-1). Gadjah Mada University Press. Yogyakarta. 499 hal.
Deptan. 2007. Pusat data dan informasi pertanian. www.deptan.go.id. (26 Oktober 2007).
Greenleaf, W. H. 1986. Pepper Breeding. p. 67-134. In Basset (Ed.). Breeding Vegetables Crops. AVI Publishing Co. Conecticut.
Griffiths, A.J.F. 1996. An Introduction to Genetic Analysis. 6nd. W.H. Freeman and Company, New York. 916 p.
Kusandriani, Y. 1996. Pembentukan Hibrida Cabai. Balai Penelitian Tanaman Sayuran. Lembang, Bandung. 19 hal.
Kusandriani, Y. dan A. H. Permadi. 1996. Pemuliaan tanaman cabai. p. 28-35.
Dalam: A.S. Duriat, A. Widjaja, W. Hadisoeganda, T.A. Soetiarso dan L.
Prabaningrum (Eds). Teknologi Produksi Cabai Merah. Balai Penelitian Tanaman Sayuran. Lembang.
Makmur, A. 1992. Pengantar Pemuliaan Tanaman. Rineka Cipta. Jakarta. 79 hal. Murti, R. H., T. Kurniawati, dan Nasrullah. 2004. Pola pewarisan karakter buah
tomat. Zuriat 15:40-149.
Messiaen, C. M. 1992. The Tropics; Vegetable Garden. ICTA Macmillan. New York. p.234-245.
Nasir, M. 2001. Keragaman Genetik Tanaman, hal 64. Dalam: Makmur, A (Ed). Pengantar Pemuliaan Tanaman. Direktorat Jenderal Pendidikan Tinggi Departemen Pendidikan Nasional. Jakarta.
Petr, F.C. and K. J. Frey. 1966. Genotypic correlation, dominance, and heritability of quantitative characters in oat. Crop Sci. 6:259-262.
29
Poehlman, J. M. 1979. Breeding Field Crops. 2nd. The AVI Publishing Company, Inc. Westport, Connecticut. 486 p.
Poespodarsono, S. 1983. Dasar-dasar Ilmu Pemuliaan Tanaman. PAU IPB. Bogor. 169 hal.
Rubatzky, V. E., dan M. Yamaguchi. 1997. World Vegetables. Principles, Production and Nutritive Values. Second Edition. Chapman and Hall. Newyork. 843 p.
Strickberger, M. W. 1976. Genetics. 2nd. Macmillan Publ. co. New York. 914 p.
Sumarni, N. 1996. Budidaya tanaman cabai merah, p.36-47. Dalam: A. S. Duriat, A. Widjaya, W.H. Thomas dan L. Prabaningrum (Eds.). Teknologi Produksi Cabai Merah. Balai Penelitian Tanaman Sayuran. Lembang.
30
Tabel Lampiran 1. Nisbah Segregasi Karakter Permukaan Buah
Nisbah segregasi
X-hitung
X-tabel 5%
3:1
2.96
tn3.84
9:3:4 33.90
**5.99
12:3:1 276.72
**5.99
9 : 7
18.73
**3.84
13 : 3
19.19
**3.84
Tabel Lampiran 2. Nisbah Segregasi Karakter Leher Buah
Nisbah segregasi
X-hitung
X-tabel 5%
3:1
0.14
tn3.84
1:3 330.81
**3.84
37:27
32.46
**3.84
45:19
3.75
tn3.84
9:7 37.92
**3.84
Tabel Lampiran 3. Nisbah Segregasi Karakter Orientasi Buah
Nisbah segregasi
X-hitung
X-tabel 5%
3:1
2.02
tn3.84
9:6:1 16.23
**5.99
10:3:3 48.11
**5.99
1:2:1
65.89
**5.99
13:3
25.45
**3.84
Tabel Lampiran 4. Nisbah Segregasi Karakter Bentuk Calix
Nisbah segregasi
X-hitung
X-tabel 5%
9:7
0.01
tn3.84
37:27 0.36
tn3.84
45:19 23.74
**3.84
55:9
177.90
**3.84
31
Tabel Lampiran 5. Nisbah Segregasi Karakter Warna Daun
Nisbah segregasi
X-hitung
X-tabel 5%
9:7
0.26
tn3.84
37:27 0.99
tn3.84
15:1 57.78
**3.84
3:1
52.71
**3.84
13:3
110.77
**3.84
Tabel Lampiran 6. Nisbah Segregasi Karakter Tipe Pertumbuhan
Nisbah segregasi
X-hitung
X-tabel 5%
1:2:1
3.71
tn5.99
10:3:3 34.86
**5.99
9:3:4 15.65
**5.99
12:3:1 200.39
**5.99
9:6:1 166.08
**5.99
Tabel Lampiran 7. Nisbah Segregasi Karakter Tipe Percabangan
Nisbah segregasi
X-hitung
X-tabel 5%
9:3:4
8.26
*5.99
12:3:1
54.95
**5.99
9:6:1
21.79
**5.99
10:3:3
27.99
**5.99
Tabel Lampiran 8. Nisbah Segregasi Karakter Posisi Bunga
Nisbah segregasi
X-hitung
X-tabel 5%
9:3:4
0.64
tn5.99
12:3:1 60.67
**5.99
9:6:1
62.13
**5.99
32
Gambar Lampiran 1. Aksi Gen Karakter Panjang Buah
Gambar Lampiran 2. Aksi Gen Karakter Diameter Buah
Gambar Lampiran. 3. Aksi Gen Karakter Tinggi Dikotomus
Gambar Lampiran 4. Aksi Gen Karakter Bobot Total
Gambar Lampiran 5. Aksi Gen Karakter Bobot Layak Pasar
P1 MP F1 9.52 15.31 13.97 21.10 P2 P2 MP F1 0.73 1.03 1.10 1.33 P1 P2 MP F1 4.72 6.46 7.33 8.19 P1 P2 MP P1 F1 130.94 147.24 90.90 50.86 P2 MP P1 F1 57.08 89.36 38.47 47.76
33 ss (b) (a)