Chonai saikin yurai kogen shigeki o uketa meneki saibo no saibonai taisha dotai kaimei
Bebas
10
0
0
Teks penuh
(2) Elucidation of the intracellular metabolic dynamics of immune cells stimulated with gut microbiota-derived antigen. 腸内細菌由来抗原刺激を受けた免疫細胞の細胞内代謝動態解明 慶應義塾大学総合政策学部4年 伊藤優太郎. 要旨 近 年 ,宿 主 と 共 生 細 菌 と の 相 互 作 用 が 互 い の 生 理 機 能 に 様 々 な 影 響 を 与 え る こ と が 報 告 さ れ て お り , 宿主と共生細菌とが織りなす複雑で洗練された複合生態系を包括的に理解することが,真の生命体の 理 解 に つ な が る と 考 え ら れ る . 卒 業 論 文 で は こ の よ う な 宿 主 と 共 生 細 菌 と の 相 互 作 用 を , マルチ オ ミ ク ス 解 析 を 用 い て 解 明 す べ く , 「植 物 疾 患 に お け る 土 壌 細 菌 叢 の 影 響 解 明 」 , 「 腸内細菌由来の刺激 に よ る 哺 乳 類 免 疫 細 胞 の 応 答 性 解 明 」 という 2 つ の テ ー マ に お け る 研 究 結 果 を 報 告 す る . 本 卒 業 論 文 ダ イジx ストでは後者に関して簡潔にまとめる. するといわれている.. ヒ ト の 腸 内 に は 約 100兆 個 , 数 百 種 類 の 腸 内 細 菌 が 存 在. こ う し た 腸 内 細 菌 は 宿 主 の 免 疫 細 胞 と 相 互 に 作 用 し 合 い ,疾患や恒 常 性 の 維 持. など様々な形で宿主に影響を与えていることが知られている. 我々はこれまでに,生体内の自然免疫系 に お い て 重 要 な 役 割 を 担 う マ ク ロ フ ァ ー ジ へ の 微 生 物 刺 激 に よ る 応 答 性 を ,遺 伝 子 発 現 レ ベ ル や ,代 謝物レベルで解析してきた.. メタボローム解析の結果, マクロファージ様細胞にグラム陰性菌の細胞. 膜構成成分であるリポ多糖( Lipopolysaccharide; LPS)刺 激 を 加 え る こ と で , DNA メ チ ル 化 の 基 質 と し て 利 用 さ れ る S -ア デ ノ シ ル メ チ オ ニ ン ( S-adenosylmethionine; た.. SAM )が 細 胞 内 に 蓄 積 す る と い う 知 見 を 得. こ の 結 果 は LPS 刺 激 に よ っ て マ ク ロ フ ァ ー ジ DNA の メ チ ル 化 が 生 じ , そ の 後 の 応 答 性 の 変 化 に 閨 与. す る 可 能 性 を 示 唆 し て い る . 先 行 研 究 に お い て , 免 疫 細 胞 に 対 す る LPS 刺 激 に よ る SAM 蓄 積 の 明 確 な メ 力 ニ ズ ム は 報 告 さ れ て お ら ず , ど の よ う に し て SAM が 細 胞 内 に 蓄 積 す る の か は 不 明 瞭 で あ る .し た が つ て 本 研 究 で は SAM 蓄 積 の メ カ ニ ズ ム 解 明 に 向 け , LPS 刺 激 直 後 の マ ク ロ フ ァ ー ジ 細 胞 内 代 謝 物 の 変 動 を メ夕ボローム解析により網羅的に探ることで,代謝物レベルでの細胞動態を明らかにすることを目指 した . そ の 進 渉 を 本 卒 業 論 文 ダ ^ ジ ェ ス ト で 報 告 す る .. Keyword: 腸 内 細 菌 , マ ク ロ フ ァ 一 ジ , SAM, LPS , メ タ ボ ロ ー ム 解 析. 99.
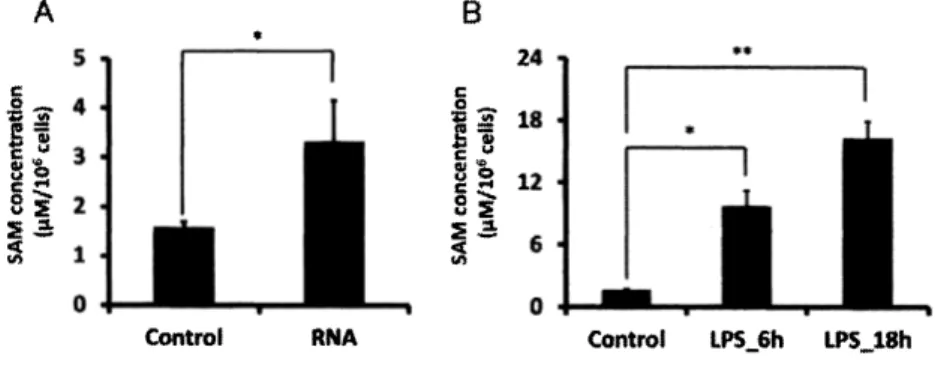
(3) 1•背景 1.1•腸内細菌による宿主免疫細胞のエピジェネティクス制御 ェピジヱネティクスとは, 「 DNA 配 列 の 変 化 を 伴 わ な い 細 胞 分 裂 後 も 継 承 さ れ る 遺 伝 子 機 能 を 研 究 す る 学 問 で あ る 」 とRiggs ら は 1996年 に 提 唱 し た ( R ig g s が a/. ,1 9 9 6 ) . 現 在 で は DNA , ヒ ス ト ン 修 飾 に よ るェピジェネティック制御が免疫記憶や細胞増殖など,宿主の免疫応答において重要な役割を担うこと が知られている.. さらに近年, そ う し た エ ピ ジ ェ ネ テ ィ ッ ク 制 御 が 共 生 細 菌 に よ っ て 引 き 起 こ さ れ ,宿. 主 の 生 体 応 答 に 影 響 を 与 え る こ と が 明 ら か と な っ て き た .腸 内 細 菌 に 対 す る 過 剰 な 免 疫 応 答 を 防 ぐ メ 力 ニ ズ ム と し て , 腸 管 上 皮 細 胞 の Toll様 受 容 体 ( Toll like receptor; TLR)4遺 伝 子 が 腸 内 細 菌 に よ る ェ ピ ジェネティック制御を受けることで抑制されることや( Takahashi.,2010), Clostridialesの 産 生 す る 酪 酸 が ナ イ ー ブ T細 胞 の ヒ ス ト ン ア セ チ ル 化 を 促 進 す る こ と で 制 御 性 T 細 胞 へ の 分 化 を 誘 導 し , 腸 内 の 恒 常 性の維持に閨わることが報告されている( Furusaw a が a/. ,2 0 1 3 ).さ ら に 腸 内 細 菌 と 宿 主 の 共 生 に も ェ ピ ジェネティックな制御が関与することが報告されている( O bata が a/., 2014). これまでの実験において,腸内細菌由来の核酸やグラム陰性菌の膜の構成要素であるリポ多糖. (Lipopolysaccharide; LPS) と い っ た 微 生 物 抗 原 を マ ク ロ フ ァ ー ジ 様 細 胞 で あ る RAW264.7 に 添 加 し た 際 の 細 胞 の 遺 伝 子 発 現 , お よ び 代 謝 動 態 を 解 析 し て き た . そ の 中 で , 微 生 物 抗 原 刺 激 に よ り S-アデノシルメ チオニン( S-adenosylmethionine; SAM)の 細 胞 内 蓄 積 が RAW264.7 に お い て 生 じ る と い う 知 見 が 得 ら れ て い る ( 図 1 ) . SAM は メ チ ル ト ラ ン ス フ ェ ラ ー ゼ が DNA や ヒ ス ト ン の メ チ ル 化 を 起 こ す 際 の 基 質 と し て 利 用されることが知られている. 従って, この結果は腸内細菌由来抗原刺激により免疫細胞にエピジェネ テ ィ ッ ク な 変 化 が 起 き , 免 疫 細 胞 の そ の 後 の 応 答 性 に 影 響 を 及 ぼ す こ と が 示 唆 さ れ て い る . 近 年 , マク ロファージの分化や機能にェピジェネティックな制御が閨与することが注目されているが( Ivashkiv .,. 2013) , マ ク ロ フ ァ ー ジ の 代 謝 研 究 の 領 域 に お い て , ェ ピ ジ ェ ネ テ ィ ッ ク 制 御 に か か わ る SAM を含むメ チ オ ニ ン 代 謝 の 動 態 は 着 目 さ れ て い な い ( Ghesquiere e? a/.,2 0 1 4 ) . し た が っ て 本 研 究 で は , 腸 内 細 菌 由 来 抗 原 の 1つ で あ る LPS 刺 激 に よ る 免 疫 細 胞 の ェ ピ ジ ヱ ネ テ イ ッ ク 制 御 を , 細 胞 代 謝 動 態 の 観 点 か ら 明 らかにすることを目指した. A s会. { S H S S O I / w a. f /s -. c o l s s c o u s<^. Control. RNA. Control. LPS^Sh. LPS_18h. 図 1 RAW 264.7 における,細胞内 SA M の蓄積 (A ) 3. Mg/Wの濃度で腸内細菌由来R N A を6 時間添加した際のR A W 264.7 細胞内のS A M の蓄積量を示す. M am wW hitney のU. 検定を用いて統計解析を行った.. ( B )5 0 0. n g /p l の濃度でL P S を6 時間,18 時間添加した際の細胞内S A M 蓄積量. Convert-. Iman法 を 用 い て 統 計 解 析 を 行 っ た .縦 軸 は lx l 〇° 細 胞 当 た り の 代 謝 物 濃 度 ( 心 ) を表す. 試験群は n=3で実施し, エラーバーは標準偏差を示す .. 100. *pc〇.〇5, **p<〇.〇〇5 を 示 す .各.
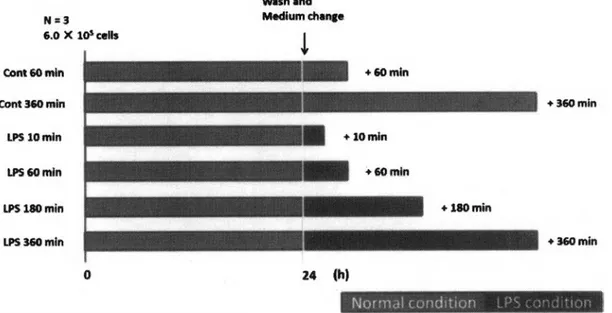
(4) 1.2•抗原刺激と内毒 素 耐性 ( Endotoxin tolerance) Endotoxin toleranceとは,細胞に対して LPS等の抗原刺激を2回行うと,2回目の刺激に対してTNF-a等 の炎症性サイト力インの遺伝子発現を抑制するのに対し,IL-10のような抗炎症性サイト力インは抑制 されない現象のことである (Biswas era /., 2009), この現象は1946年, Millerらがウサギに対し,腸チフ スのワクチンを複数回投与すると, ワクチンが引き起こす熱が弱まることを報告したものが最初の知 見にあたる( Miller が 2 0 0 6 ) . その後ヒトにおいても同様の現象が起こることが報告されており,現 在 で は が ん や 敗 血 症 ,嚢 胞 性 線 維 症 と い っ た 難 病 患 者 の 細 胞 に み ら れ る 現 象 と し て 知 ら れ て い る (Lopez-Collazo d. 2 0 1 3 ) . その分子メカニズムは TLRシグナル経路におけるキナーゼの不活性化や,. micro RNAによる転写後調節, エピジェネティックな制御が示唆されている (Biswas etal .,2009) . 本研 究では, この現象のメカニズムの一^9 であるエピジヱネティック制御が,抗原刺激による SAM生産に 向けた代謝のリプログラミングにより生じているという仮説の検証を目的としている.. 2.対象と手法 2.1. LPS 刺激直後の細胞内代謝物動態解析 マウスのマクロ フ. ァ ー. ジ様細胞株である R A W 2 6 4 . 7 細胞に L P S 刺激を行った際の細胞内代謝物動態を. 調べるため, L P S の有無や L P S 刺激時間を変えた 6 群を用意し,細 胞 内 の 代 謝 物 を 比 較 し た ( 図2 ) . R A W 2 6 4 .7 は 5 0 0 m lの ダ ル ベ ッ コ. 変法イ一グ ノレ培 地 (D u l b e c c o ’s. M o d i f ie d E a g l e M e d iu m 培地; DM EM. 培地,. S I G M A ) に5 0 m l のウシ胎児血清 ( F e t a l b o v i n e se r u m ; F B S ) と5 m l の抗生物質の混合液( ペ ニ シ リ ン 1 0 ,0 0 0 u n it/m l,ストレプトマイシン 1 0 ,0 0 0. アムホテリシンB. 2 5 p g / m l ) を加えたもので培養した. 2 度継代. を 行 っ た Raw264.7 を 6-well プ レ ー ト に 6><105細 胞 ず つ 播 種 し , 培 地 を 2 m l ず つ 加 え た 状 態 で 24 時 間 の 前 培 養 を 行 っ た . 24 時 間 後 , l x リ ン 酸 緩 衝 生 理 食 塩 水 (Phosphate buffered saline; PBS)で ウ ォ ッ シ ュ を 1度 行 っ た 後 , 最 終 濃 度 が 500 ng/ml に な る よ う に LPS. co// 055:B5, SIGMA)を 培 地 に 添 加 し た .. 対 照 群 に 対 し て は , lxPB S で ウ ォ ッ シ ュ 後 , 培 地 交 換 の み 行 っ た . LPS 刺 激 か ら 10,60 ,180,360 分 の タ イ ミンダで, 代 謝 物 測 定 用 の 前 処 理 を 行 っ た . Wash and. 図2. L P S 刺激直後の細胞内代謝物動態解析の概要. R A W. 264.7細胞を6.〇x l〇5細胞ずつ各ウヱルに添加し,24時間前培養した. 24時 間 後 lxPBSでウォッシユを1度行い, LPS. を最終濃度が500 ng.mlになるように添加した.また対照群として培地交換を行った群を設けた.LPS刺激から10, 60, 180,360分の タイミングで細胞を回した.赤色の バーは培地のみの状態を,黒色のバーはLPSを培地に添加している状態 を示す.. 101.
(5) 2.2. メ 夕 ボ ローム解 析 の 前 処 理 2.1•に 不 し た 条 件 で 細 胞 を 処 理 し た 後 , ア ス ピ レ ー タ ー を 用 い て 各 weii の 培 地 を 除 去 し , 5 ° /〇 (w/v )の マ ン ニ ト ー ル を 含 む Ultra Pure Distilled Water (Pure Water, In vitrogen)を 5 mr添 加 し ウ ォ ッ シ ュ を 行 っ た • ウ 才 ッ シ ュ 後 こ の 溶 液 を 抜 き , 再 び 1 m l の 5% (w /v) の マ ン ニ ト ー ル を 含 む Pure Waterを添加!し た . 添 加 後. Cell Lifter (SIGMA)を 用 い て 細 胞 を 剥 が し , こ の 溶 液 を 1.5 m l チ ュ ー ブ に 移 し た . 再 度 各 well に 3 5 0 一の 5% (w/v) の マ ン ニ ト ー ル を 含 む Pure Waterを 添 加 し 細 胞 の 回 収 を 行 っ た .. こ の 溶 液 を よ く 撹 拌 し , 50 pi. を 回 収 し て 血 球 計 算 盤 を 用 い て 細 胞 数 の カ ウ ン ト を 行 っ た • 残 り の 細 胞 は 13,000x g , 1 m in ,4 て の 条 件 で 遠心分離を行い,上 清 を 完全に除いた. その後,細胞ペレットが入っているチューブをチューブラック に こ す り つ け て ペ レ ッ ト を ほ ぐ し , 4 0 0 …の 内 部 標 準 入 り メ タ ノ ー ル ( 2 0 卩M し メ チ オ ニ ン ス ル ホ ン , 20. pM 2- モ ル ホ リ ノ エ ス ル ホ ン 酸 , 20 pM D-10- カ ン フ ァ ー ス ル ホ ン 酸 ) を 添 加 し て 使 用 時 ま で -8 0 T で 保 存 し た.. 400 p i の 内 部 標 準 入 り メ タ ノ ー ル を 添 加 し た 細 胞 溶 液 に 対 し 400 pi の クロ ロ ホ ル ム と 200 pi の Milli-Q を 加 え , よ く 撹 拌 し た 後 9,100x g , 15 min, 4 て の 条 件 で 遠 心 分 離 を 行 っ た . 400 pi の 水 層 を 限 外 濾 過 カ ラ ム. (Millipore)に 移 し , 9,100x g , 180 min, 4°C の 条 件 で 限 外 濾 過 を 行 っ た . フ ロ ー ス ル 一 を 濃 縮 遠 心 乾 燥 機 (LABCONCO)を 用 い て 乾 固 し , 25 |il の 内 部 標 準 入 り Milli-Q (200 juM 3 -ア ミ ノ ピ ロ リ ジ ン , 200 1,3,5- ベ ン ゼ ン ト リ カ ル ボ ニ ル ト リ ク ロ リ ド ) を 用 い て 溶 解 し た . こ の 溶 液 を キ ャ ピ ラ リ ー 電 気 泳 動 時 間 飛 行 型 質 量 分 析 (CE-TOFMS)に 用 い る こ と で 代 謝 物 の 測 定 を 行 っ た .. 2.3. SAM阻 害 剤 を 用 い た 細 胞 の 応 答 性 解 析 RAW264.7 細 胞 に お け る SAM の 役 割 を 調 べ る べ く , メ チ オ ニ ン か ら の SAM を 合 成 す る メ チ オ ニ ン ア デ ノ シ ル 基 転 移 酵 素 2 (Methionine adenosyltransferase; Mat2)の 阻 害 剤 で あ る シ ク ロ ロイ シ ン ( Cycloleucine) や , DNA メ チ ル 化 酵 素 で あ る DNA メ チ ル 基 転 移 酵 素 (DNA metyltransferase; DNMT )の 阻 害 剤 と し て 働 く. 5- ア ザ シ チ ジ ン (5-azacytidine) を 利 用 し た 実 験 を 行 っ た . 2 回 継 代 を 行 っ た RAW264.7 細 胞 を , 6-well プ レ ー ト に 6.〇x l 〇5細 胞 ず つ 播 種 し た . 24 時 間 培 養 の 後 , 阻 害 剤 と し て Cydoleucin (最 終 濃 度 10 mM), 5-. azacytidine ( 最 終 濃 度 100 pM ) を LPS (最 終 濃 度 500 ng/m l) と 共 添 加 し た . ま た , 対 照 群 と し て 同 様 の 条 件 に お い て LPS 非 添 加 群 , ジ メ チ ル ス ル ホ キ シ ド (Dimethyl sulfoxide; DMSO)添 加 群 を 用 意 し た . 12時 間 培 養 後 , LPS 刺 激 を 行 っ て い な い 群 で は 培 地 交 換 の み を 行 い , LPS 刺 激 を 行 っ た 群 で は 最 終 濃 度 が 500 ng/. m l に な る よ う 培 地 交 換 を 行 っ た . 6 時 間 培 養 後 , こ れ ら の 細 胞 に 対 し , 1% の 2 -メ ル カ プ ト エ タ ノ ー ル (2-mercaptoethanol)を 含 ん だ 600 |iil の Buffer RLT (RNeasy Plus Mini Kit) を 添 加 し て 細 胞 を 回 収 し , 使 用 時 まで -80°Cで 保 存 し た . mt41um ctmm#. 0 + 刚 bitsir. __. im/ips iHwiso. 0. 3&. 24. 102. 42 (h).
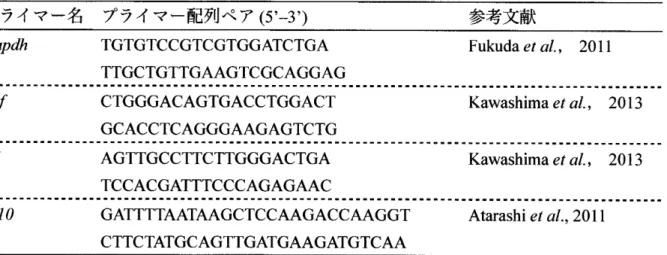
(6) 図3 阻害剤による細胞応答性解析実験の概要. RAW264.7細胞を 6.〇xl 〇5細胞ずつ各ウエルに添加し,24時間前培養した . 24時 間 後 lxPBSでウォッシユを 1度行い , LPS を最終濃度が 500 ng/ml, Cycloleucineを最終濃度が 10 mM, 5-azacytidineを 最 終 濃 度 が 100 |liMになるように添加した . DMSOは5-azacytidineの対照群として等量添加した . 処 理後 12時間経過時,再 度 lxPBSでウォッシユを 1度行い, LPSを最 終濃度が 500ng/mlになるよう添加した . また対照群として培地交換を行った群を設けた . 処理後 6時間時に各細胞を回収 した. 赤色のバーは培地のみの状態を,黒色のパーは LPSを培地に添加している状態を示す .. 2.4.培 養 細 胞 の 遺 伝 子 発 現 量 解 析 R N A 抽 出 に は R N easy Plus M ini Kit (Q IA G E N ) を 用 い た . 各 処 理 の 後 , 1% の 2m ercaptoethanol を 含 ん だ Buffer RLTに 溶 解 し た 状 態 で -80 〇 Cで 保 存 し て い た 溶 液 全 量 を gDNA Eliminationカ ラ ム (RNeasy Plus Mini Kit付 属 ) に 移 し , 8,000xg ,25 °C ,1 min の 条 件 で 遠 心分離した.. そ の 後 は キ ッ ト に 付 属 の プ ロ ト コ ル に 従 い RNA 抽 出 処 理 を 行 っ た . 抽 出 し た. RNA は Nanodrop (Thermo)で 濃 度 , 純 度 の 測 定 を 行 っ た . 抽 出 し た RNA を ReverTraAce(TOYOBO) を 用 い , 3(TC, lOmin , 42°C,20m in , 99°C ,5min の 条 件 で 逆 転 写 し , cD N A を 合 成 し た .. 合 成 さ れ た cD N A は 各 種 サ イ ト 力 イ ン ( TNF-a,. IL-6, IL-10)の 遺 伝 子 に 特 異 的 な プ ラ イ マ ー を 用 い , 95°C , 30 秒 の 初 期 変 性 の 後 , 95°C, 5 秒 , 60°C, 30 秒 の 条 件 で 40 サ イ ク ル の q-PCR を 行 っ た .. また, 比 較 定 量 法 を 行 う た め に , ハ. ウ ス キ ー ピ ン グ 遺 伝 子 で あ る 说 の 特 異 的 プ ラ イ マ ー を 用 い , 内部標準として利用する こ と で 遺 伝 発 現 量 の 正 規 化 を 行 っ た . 各 特 異 的 プ ラ イ マ ー の 塩 基 配 列 を 表 1に 示 す . 表 1本研究で使用したブライマー. プライマー名. プライマー配列ペア( 5’- 3 , ). 参考文献. Gapdh. TGTGTCCGTCGTGGATCTGA. Fukuda etal.,. 2011. TTGCTGTTGAAGTCGCAGGAG. Tnf. CTGGGACAGTGACCTGGACT. Kawashima et al.,. 2013. Kawashima et al.,. 2013. GCACCTCAGGGAAGAGTCTG. 116. AGTTGCCTTCTTGGGACTGA TCCACGATTTCCCAGAGAAC. IL10. GATTTTAATAAGCTCCAAGACCAAGGT. Atarashi et al., 2011. CTTCTATGCAGTTGATGAAGATGTCAA 2.5.デ ー タ 解 析 CE-TOFMS に よ っ て 得 ら れ た デ 一 夕 は マ ス タ ー ハ ン ズ を 用 い て ピ ー ク の 切 り 出 し , 濃 度 の決定を行った.. ヒ ー ト マ ッ プ は EXCEL に よ り z-score を 算 出 し , 各 代 謝 物 の z-score の 値 を. も と に MeV で 描 写 し た . 描 写 さ れ た ヒ ー ト マ ッ プ は ス ピ ア マ ン の 相 閨 係 数 を 用 い た 階 層 的 クラス夕リングによりクラスタリングされた. を用いて解析した.. 各 代 謝 物 群 の 関 与 す る 代 謝 経 路 は Metacore. また,本研究では各種サイト力インの遺伝子発現に関して統計的な検. 定 を 行 っ た . 2 群 間 比 較 と し て , EXCEL を 用 い た Studentの T検 定 を 行 っ た .. 103.
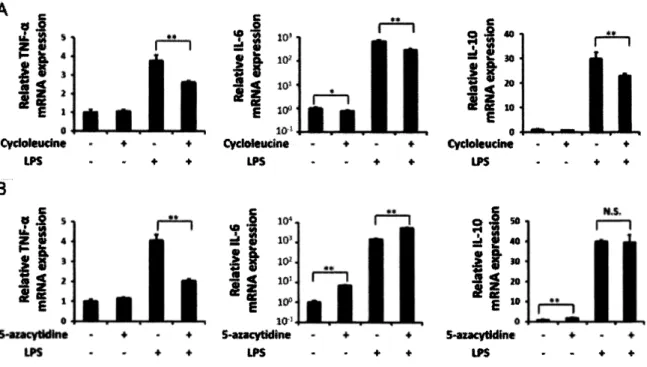
(7) 3•結 果 3.1. SAM蓄 積 の 代 謝 経 路 探 索 こ れ ま で の 研 究 に よ り , LPS 刺 激 後 6 時 間 , 18時 間 後 に SAM の 細 胞 内 蓄 積 が 生 じ る こ と 示 さ れ た ( 図. 1 ) . SAM の 細 胞 内 蓄 積 の さ ら な る 知 見 を 得 る べ く , LPS 刺 激 直 後 で あ る 10分 か ら 360 分 後 の 細 胞 内 の 代 謝 物 を 測 定 し , そ の 動 態 を 代 謝 物 ご と に z-score で 標 準 化 す る こ と で ヒ ー ト マ ッ プ を 作 成 し た ( 図 4A). 赤枠で囲った代謝物群の多くはグノレ夕ミン代謝やアルギニン代謝, ク エ ン 酸 回 路 ( TCA 回 路 ), Y-アミノ 酪 酸 ( GABA)合 成 経 路 , メ チ 才 ニ ン 代 謝 等 に 閨 わ る 代 謝 物 で あ る こ と が 示 さ れ た .ま た グ レ ー で 囲 っ た 代 謝 物 は LPS 刺 激 後 180分 の 時 点 で は 多 く 存 在 す る の に 対 し , 360 分 時 に 減 少 し て い る 代 謝 物 を 示 す .こ れ ら の 代 謝 物 は ベ ー タ ア ラ ニ ン 代 謝 や グ リ シ ン 代 謝 , メ チ オ ニ ン 代 謝 , ア ル ギ ニ ン 代 謝 , リジン代 謝 , タ ウ リ ン 代 謝 に 関 わ る 代 謝 物 で あ る こ と が 示 さ れ た . 特 に グ リ シ ン 代 謝 , メ チ オ ニ ン 代 謝 , タウ リ ン 代 謝 は SAM 周 辺 の 代 謝 経 路 で あ っ た ( 図 4B). Cont. [3 60mm 画. m. 網 m椒. B. 級 扣. _. min. 鲂 讲 抝. 臞 180min _ 360m in. pathway Beta>alan{ne m e t a b o l i s m Glycine, serine, cysteine, t h r e o n i n e m e t a b o l i s m Alanine, Cysteine a n d m e t h i n i n e m e t a b o i i s m Argin»n« m e t a b o l i s m lysine m e t a b o l i s m Taurine a n d hypot a u r i n e m e t a b o l i s m. pathway HIstldine-glutamate>glutamine m e t a b o i i s m Arginine mctai>otism Trtcarbonic acid cycle 6 A B A biotynthesband metabofism Abinine, Cysteine a n d m e t h i n i n e m e t a b o l i s m. 図4. lvalue p s 3.787e-5 P = 1.244e-4 P » 2.256 5.353e-4 P * 7.990e-4. l»=rl.008e*4. P -va lue. P»3.382e>8 P « 1,432 か 6 P = 1.624 P * 1.973e*6 2.603«-6. L P S 剌激直後の細胞内代謝物動態解析. (A)LPS 刺激直後の細胞内代謝物動態を, Z-SC〇reを用いたヒートマップで示した . 縦軸に各代謝物を,横軸に各試験群を 示 す •対照群とし て 培地交換後 10, 360分時の細胞内代謝物, LPS刺激後のタイムコースとして 10, 60,180, 360分の細胞内 代謝物を測定し,対照群, LPS刺激群の順番で時間経過ごとに i-viと番号を振り横軸に示した.青で示すプロットは細胞 内に少ない代謝物を, 黄色で示すプロットは細胞内に多い代謝物を示す.本研究では四角で囲った代謝物群に着目して おり, Metacoreを用いて算出されたこれらの代謝物の閨与する代謝経路とその p値を横に記した . 代謝経路はその p値の 値が小さなものから順に上から記した . グレーで囲った代謝物群は LPS刺 激 後 180分時に増加し, 360分時に減少してい た代謝物群を示す • 赤で囲った代謝物群は LPS刺 激 時 180分から 360分にかけて増加した代謝物群を示す . ( B) SAM周辺の 代謝物の濃度変化を KEGG pathwayに基づいてマッピングしたもの . グラフのない代謝物は,本研究で測定できなかった 代謝物を示す • 横軸の Cは対象群, 10, 60,180, 360はそれぞれ 10, 60,180, 360分の LPS刺 激 群 を 示 す .縦 軸 は lx l 〇6細胞当 たりの代謝物の濃度( HM)を表す. 矢 印は 1つの酵素による反応を,破線矢印は複数の酵素による反応を示す.各試験は n=3で実施されており,エラーバーは標準偏差を示す .. 3.2. SAM 阻 害 剤 利 用 に よ る 細 胞 の 遺 伝 子 発 現 動 態 Cycloleucine を 用 い た SAM 合 成 阻 害 , お よ び 5-azcytidine を 用 い た DNA メ チ ル 化 阻 害 に よ る 細 胞 の 遺 伝 子 発 現 動 態 を 調 べ た . L P S と C y c lo le u c in e 共 添 加 時 の T N F -a や IL -6 , IL -1 0 の 遺 伝 子 発 現 量 は ,. Cycloleucine非 添 加 時 と 比 べ 有 意 に 減 少 し た (図 5 A ) . そ れ に 対 し L PS と5-azacytidineの 共 添 加 を 行 う と , 5-azacytidine 非 添 加 時 と 比 べ TNF-a の 遺 伝 子 発 現 量 は 有 意 に 減 少 す る が IL-10 は 変 化 が な く , IL-6 に関し ては有意に上昇した( 図 5 B ) . また LPS 非 添 加 群 に お い て , 5-azacytidine の 添 加 は IL-10 や IL-6 の 遺 伝 子 発 現量を有意に上昇させた.. こ れ に 対 し , C ycloleucine の 添 加 は IL -6 の 遺 伝 子 発 現 量 を 有 意 に 減 少 さ せ. た.. 104.
(8) A 5. 箱 踟. 炉 i. I. 0 3一i. 1 が i. f. I. 0 *. 4 0. 妒舻 s. s. !. 10. «l n. 細. M. 一. 一. ^. 8 锄. 1. o 0f 0 1 - 1. i. » •1. 騸 锄. i. W. . ^r 0. 1. i. 選. 8. 0 0 沪 1. 2 0 1 0. 0- w I to. 図5 SAM阻害による LPS2回刺激時のサイト力イン遺伝子発現量解析 (A )L P S とC y c lo le u c in e の共添加による T N F -a,. IL -6 ,IL -1 0 の遺伝子発現量への影響を調べた .. ( B ) L P S と5-azacytid in e の共. 添加による T N F -a ,IL-6, IL -1 0 の遺伝子発現量への影響を調べた . L P S はL P S の2 回刺激を行った群を示す.-は各処理を 行っていないものを, + は各処理を行ったものを示す . IL -6 の縦軸は対数軸で表示して い る . 各試験は n = 3 で実施して お. り,エラ一バーは 標準偏差を示す.統計検定は 2 群検定の Stu dent のT テストを行って い る . *. p < 0 .0 5 ,* * p < 0 .0 0 5 を,N S は. 統計的に有意差がないことを示す .. 4. 讓論 4.1.. 細胞内SAM 蓄積の代謝経路解明. 本 研 究 に よ り 得 ら れ て い る 知 見 で あ る SAM の 細 胞 内 蓄 積 に つ い て , ど の よ う な 経 路 を 介 し て 生 じ て い る の か を 調 べ る べ く , LPS 刺 激 直 後 の 細 胞 内 代 謝 物 動 態 を 調 べ た ( 図 4A). LPS 刺 激 後 180, 360分 後 に 多 くの代謝物が増加していることが明らかとなった.. こ れ ら の 代 謝 物 は グ ル タ ミ ン 摂 取 の 増 加 に よ る TCA. 回 路 物 質 の 上 昇 や GABA シ ャ ン ト に よ る コ ハ ク 酸 の 増 加 , ア ル ギ ニ ン 代 謝 に よ る 一 酸 化 窒 素 産 生 な ど , 炎症性マクロファージの代謝物指標として知られる代謝経路と関連していることが示唆された. (Ghesquiere W «/•,2014, M ills が a/., 2 0 1 4 ) . 次 に グ レ ー の 枠 で 囲 わ れ た 180分 に 増 加 し , 360 分 で 減 少 し て いる代謝物群に着目した.. こ れ ら の 代 謝 物 群 の 閨 わ る 代 謝 経 路 の う ち , グ リ シ ン 代 謝 , メチオニン代. 謝 , タ ウ リ ン 代 謝 は SAM 周 辺 の 代 謝 経 路 で あ る ( 図 4B). SAM 周 辺 の 代 謝 物 が 180分 か ら 360分 の 間 で 減 少 し て い る の に 対 し , SAM 自 体 は 180分 か ら 360 分 の 間 で 増 加 し て い る .. こ の 結 果 か ら , 細 胞 は LPS 刺 激. 後 180 分 か ら 360 分 の 間 に SAM 生 産 に 向 け て 代 謝 の リ プ ロ グ ラ ミ ン グ を 行 っ て い る 可 能 性 が 示 唆 さ れ た . 赤 枠 で 囲 っ た 代 謝 物 群 は 180分 の 時 点 で 増 加 し て い る の に 対 し , グ レ ー の 枠 の 180分 か ら 360分 で 変 動 し て い る 代 謝 物 は 比 較 的 遅 い 応 答 性 を 示 し て い る と 考 え ら れ , LPS 刺 激 か ら の 直 接 的 な 応 答 で は な く, 2 次 応 答 の よ う な 形 で 応 答 し て い る と 考 え ら れ る . グ レ ー の 枠 内 で あ げ ら れ て い る ア ル ギ ニ ン 代 謝 は サ イ ト 力 イ ン で あ る IFN-P を 介 し て 亢 進 す る こ と が 知 ら れ て お り ( Jacobs 以 a/.,2001) , SAM 生 産 の 増 加 もLPS 刺 激 に よ っ て 生 じ る サ イ ト 力 イ ン 等 を 介 し て い る と 推 測 し て い る .. 4.2.. 細胞内SAM 蓄積の役割の解明. 次 に , 細 胞 内 の SAM 蓄 積 の 役 割 を 解 明 す る こ と を 目 指 し た .. こ れ ま で に 報 告 さ れ て い る SAM の働き. として, 外 因 性 の SAM 投 与 に よ る SAM の 細 胞 内 蓄 積 は , 免 疫 細 胞 の エ ピ ジ ヱ ネ テ ィ ッ ク 修 飾 を 介 し て. 105.
(9) TNF-a な ど の 炎 症 性 サ イ ト 力 イ ン 生 産 を 抑 制 し , IL-10 な ど の 抗 炎 症 性 サ イ ト 力 イ ン 生 産 を 促 進 す る こ と が 報 告 さ れ て い る (Kharbanda W a/., 2013, Pfalzer が <. 2 0 1 4 . ) . こ れ ら の 炎 症 抑 制 能 は 外 因 性 の SAM に起. 因 す る と さ れ て い る が , 本 研 究 で の 免 疫 細 胞 に お け る SAM の 蓄 積 の 結 果 か ら , 細 胞 内 で こ れ ら の 応 答 が 生 じ て い る と 考 え ら れ る . つ ま り 微 生 物 抗 原 に よ る 炎 症 刺 激 の 後 , 細 胞 の 代 謝 が SAM 生 産 へ と リ ブ ロ グ ラ ミ ン グ さ れ , TNF-a 等 の 炎 症 性 サ イ ト 力 イ ン が エ ピ ジ ヱ ネ テ ィ ッ ク 制 御 に よ り コ ン ト ロ ー ル さ れ ることで炎症終結に寄与するのではないかという仮説である. そ の 仮 説 を 検 証 す べ く , 本 研 究 で は Endotoxin. to le ra n c e と 呼 ば れ る 現 象 に 着 目 し た . Endotoxin. tolerance は 抗 原 に 対 す る 過 剰 応 答 に よ る 宿 主 へ の ダ メ ー ジ を 抑 制 す る , い わ ば 炎 症 終 結 機 能 の 1つ で あ る.. こ の 現 象 で は LPS の 2 回 刺 激 に よ っ て TNF-a 等 の 炎 症 性 サ イ ト 力 イ ン の 抑 制 が 生 じ る こ と が 知 ら れ て. い る が , 1度 目 の LPS 刺 激 後 に 起 こ る SAM 蓄 積 や エ ピ ジ ェ ネ テ ィ ッ ク 制 御 を 阻 害 す る こ と で , 2 度 目 の. LPS 刺 激 の 際 に 炎 症 性 サ イ ト 力 イ ン が 抑 制 さ れ な い の で は な い か と 考 え た (図 5 ) . し か し な が ら , 予 想 し て い た 結 果 と は 逆 に , 2 回 刺 激 時 の TNF-a の 遺 伝 子 発 現 量 は 対 照 群 と 比 べ , 抑 制 さ れ て い る と い う 結 果 が 得 ら れ た . Cycloleucine を 用 い た 際 に は 1し 6 , IL-10 も 同 様 に , 対 照 群 と 比 べ て 抑 制 さ れ て い る . こ れらの結果から,. Cycloleucine は 1度 目 の LPS刺 激 の 炎 症 抑 制 を 促 し て お り , 2 度 目 の 刺 激 後 に こ れ ら の. サイト力インの遺伝子発現が減少しているように観察されたのではないかと考えている. 一 方 Iし 6 の 遺 伝 子 発 現 量 は , Cycloleucine 処 理 で は 有 意 に 抑 制 さ れ る が , 5-azacytidine 処 理 で は 有 意 に 増 加 し た . 塍 臓 が ん 細 胞 に お い て , IL-6 は DNA の メ チ ル 化 修 飾 に よ り 遺 伝 子 発 現 が 抑 制 さ れ て お り , 5-. azacytidineに よ る DNA メ チ ル 化 の 阻 害 は IL-6 の 発 現 を 上 昇 さ せ る こ と が 報 告 さ れ て い る ( D a n d re a 办 a /., 2009) . マ ク ロ フ ァ ー ジ に お い て も IL-6 は 同 様 の 発 現 抑 制 機 構 を 備 え て い る 可 能 性 が 考 え ら れ る . こ れ ら の 結 果 を 踏 ま え る と , 今 回 行 っ た 実 験 系 で は SAM 蓄 積 に よ る 炎 症 終 結 が 生 じ る と い う 仮 説 を 証 明 す る こ と は で き な か っ た . 今 後 は 阻 害 剤 を 入 れ る タ イ ミ ン グ の 変 更 や CE-TOFMS を 用 い た 阻 害 剤 の 効 果 の 検討などを行い, より詳細な検証を重ねていく.. 謝辞 本研究を行うにあたり,実験から解析の方法まで,幾度となくミーティングを行ってくださいました 福田真嗣特任准教授には大変感謝いたします.. また, 細 胞 の 扱 い や 結 果 な ど に 関 し ま し て 相 談 し て く だ. さいました田畑祥特任助教にも大変感謝いたします.. さ ら に , ア ド バ イ ザ ー と し て TP の添削等行ってく. だ さ い ま し た 村 上 慎 之 介 氏 に も こ の 場 を お 借 り し て 御 礼 申 し 上 げ ま す. 最 後 に , このような研究の場を 与えてくださった冨田勝教授に感謝申し上げます.. 参考文献 A tarash i,K ., Tanoue, T., Sm m a, T., Imaoka, A ., Kuwahara, T., M om ose, Y.? e t aL (2 011). Induction o f colon ic regulatory T cells bv indigenous Clostridium sp ecies. Scien ce (N ew York, N.Y.)t 3 3 7 (6 0 1 5 ), 3 3 7-341. B isw as, S. K., & L op ez-C ollazo, E. (2009). E naotoxin tolerance: N e w m ech anism s, m o lecu les and clinical significance. Trends in Im m unology, 3 0 (1 0 ), 4 75-487. Dandrea, M .5 D onadelli, M .? C ostanzo, C., Scarpa, A ., & Palmieri, M. (2 0 0 9 ). M eC P 2/H 3m eK 9 are in volved in IL-6 gene silen cing in pancreatic adenocarcinom a cell lines. N u cleic A cid s R esearch, 37 (2 0 ), 668 1 -6 6 9 0 . Edward S. M iller and Paul B. B eeson . (1 9 4 6 ) M urine typhus fever. M edicine, 2 5 (1 ), 1-16. Fukuda, S .? Toh5 H., H ase, K ., O shim a, K .? Nakanishi, Y., Yoshim ura, K ., e t ah (2011). Bifidobacteria can protect from enteropathogenic infection through production o f acetate. Nature, \ 543-547. Furusawa, Y., Obata, Y.? Fukuda, S., Endo, T. A ., Nakato, G ., Takahashi, D ., e t aL (2 0 1 3 ). C om m ensal m icrobe-derived butyrate induces the differentiation o f colon ic regulatory T cells. Nature, 5 0 4 (7 4 8 0 ), 44 6 -4 5 0 . G hesquiere, B .? W ong, B . W.? K uchnio, A ., & Carmeliet, P. (2 014). M etabolism o f strom al and im m une cells in health and disease. Nature, 5 7 /(7 5 0 8 ), 167-176. Ivashkiv, L. B. (2013). E pigenetic regulation o f m acrophage polarization and fiinction. Trends in Im m unology, 34(5), 216-223. Jacobs, A . r., & Ignarro, L. J. (2001). Lipopolysaccharide-induced expression o f interferon-beta m ediates the tim ing o f inducible nitric-oxide synthase induction in RAW 2 64.7 m acrophages. The Jou rn al o f B io lo g ic a l Chem istry, 2 7 6(51), 47950-47957. Jacobs, A . T., & Ignarro, L. J. (2001). L ipopolysaccharide-induced expression o f interferon-beta m ediates the tim ing o f inducible nitric-oxide synthase induction in RAW 2 64.7 m acrophages. The J o u rn al o f B io lo g ic a l Chem istry, 275(51), 47950-47957. K awashim a, T.? Kosaka, A ., Yan, H .? Guo, Z .? U chiyam a, R., Fukui, R., e t aL (2 0 1 2 ) Double-Stranded R N A o f Intestinal Com m ensal but N o t Pathogenic Bacteria Triggers Production o f Protective Interferon-p. Immunity. 38,1 1 8 7 -97.. 106.
(10) Kharbanda, K. K .? Bardag-G orce, F.? Barve, S.? M olina, P. E.? & Osna, N . A . (2013). Impact o f altered m ethylation in cytokine signaling and proteasom e function in alcohol and viral-m ediated diseases. A lcoh olism , C lin ical a n d E xperim ental R esearch , 3 7 (1 ) , 1-7. L opez-C ollazo, E .? & del Fresno, C. (2013). Pathophysiology o f endotoxin tolerance: M echanism s and clinical consequences. C ritica l C are (London , England), 1 7 (6 ) ,242. M ills, E.5 & O 'N eill,L . A . (2014). Succinate: A m etabolic signal in inflam m ation. Trends in C ell Biology, 24(5% 313-320. Obata, Y., Fxirusawa, Y., Endo, T. A ., Sharif, J.5 Takahashi, D ., A tarash i,K ., et a l . (2014). The epigen etic regulator U h rfl facilitates the proliferation and maturation o f colon ic regulatory T cells. N ature Im m unology, 75(6), 571-579. Obata, Y.? Furusawa, Y., Endo, T. A .? Sharif, J., Takahashi, D .? Atarashi, K .? e t aL (2014). The epigen etic regulator U h rfl facilitates the proliferation and maturation o f colon ic regulatory T cells. Nature Im m unology, 75(6), 571-579. Pfalzer, A . C., C hoi, S. W.? Tammen, S. A ., Park, L. K., Bottiglieri, T.? P a rn ell,L . D ., e t a l (2 014). S-adenosylm ethionine m ediates inhibition o i inflam m atory response and changes in D N A m ethylation in human m acrophages. P h ysio lo g ica l Genom ics, 必(1 7 ) , 6 1 7 -6 2 3 . R iggs A D ., R u sso V E A ., M artienssen R N A . 1996. Epigenetic m echanism o f gene regulation. Plainview, N.Y. C o ld S prin g H arb o r L a b o ra to ry Press. Takahashi, K. (2 0 10). Interaction betw een the intestinal immxme system and com m ensal bacteria and its effect on the regulation o f allergic reactions. B ioscience, B iotechnology, a n d B iochem istry, 74(4), 691-695.. 107.
(11)
Gambar
Dokumen terkait
Perusahaan-perusahaan transnasional dan revolusi hijau untuk Afrika telah memperkenalkan pertanian kontrak [di mana petani berkomitmen untuk memproduksi produk dengan cara
Studi yang berjudul “English Prepositional Phrases Using Prepositions “By”,“With”, and “Without” and Their Translation Equivalences in Indonesian adalah mengenai
Mempertahankan image dari bahasa sumber (bsu) dalam menerjemahkan metafora adalah untuk menyampaikan makna yang sama kepada para pembaca di bahasa sasaran (bsa), sedangkan
Demikianlah, melalui beberapa KBK di atas diharapkan lulusan Program Studi Pendidikan Bahasa Arab memiliki keterampilan yang unggul serta relevan dengan kebutuhan
Dengan kata lain, selain membawa pesan, kegiatan penerjemahan juga merupakan kegiatan untuk mengubah bentuk bahasa dengan tujuan agar teks hasil terjemahan bisa
tersebut sudah menjadi tanah negara bebas.Oleh karena itu diperlukan pembuktian tertulis yang diketahui oleh Lurah atau Kepala Desa, dan Camat setempat, sehingga dapat
Dataset yang didapatkan berformat .csv (comma separated values). File film tersebut berisi data- data film yang memiliki atribut: movieid, title, genres, imdbid, plot,
