Biawak Komodo plastis: Respon predator besar terhadap pulau kecil
Tim S. Jessop1,2, Thomas Madsen3, Claudio Ciofi4, M. Jeri Imansyah1, Deni Purwandana1, Achmad Ariefiandy1, John A. Phillips11 Conservation and Research for Endangered Species, Zoological Society of San Diego, Escondido Ca 92027, USA.
2 Department of Wildlife and Conservation Science, Zoos Victoria, Parkville Victoria 3052, Australia 3 School of Biological Sciences, University of Wollongong NSW 2522, Australia.
4 Department of Animal Biology and Genetics, University of Florence, Via Romana 17, 50125 Florence, Italy.
Alamat korespondensi: Tim S. Jessop
Department of Wildlife Conservation and Science Zoos Victoria
Parkville Vic 3052
Email: [email protected]
RINGKASAN
Pulau kecil mempunyai keterbatasan secara energetik sehingga menyulitkan predator besar, sebagai konsekuensi rantai makanan yang miskin mangsa. Didalam penelitian ini kami menguji kapasitas morfologi dan perbedaan tingkah laku biawak Komodo (Varanus komodoensis), kadal pemangsa terbesar di dunia, dalam kaitan responnya terhadap tingkatan kehadiran mangsa ungulata pada dua pulau besar dan dua pulau kecil. Kami mendemonstrasikan pola-pola berulang perbedaan morfologi biawak Komodo di dua pulau kecil, yang miskin mangsa, dan memiliki bentuk tubuh ektomorfik (ectomorphic) (tubuh lebih panjang, ekor lebih pipih, dan lebih ringan) dari pada populasi di dua pulau besar. Kewaspadaan, sebagai dampak tekanan akibat pemangsaan, digunakan untuk mengukur predasi intraspesifik, dan juga ditemukan mengikuti perbedaan pulau besar – pulau kecil. Biawak Komodo di pulau kecil secara signifikan lebih waspada dari pada komodo di pulau besar. Pola-pola perbedaan sifat bawaan banyak bertentangan dengan estimasi aliran gen di antara seluruh populasi, sehingga mekanisme yang berkaitan dengan bukan-makanan pada perbedaan fenotip kepulauan tidak diperhitungkan. Hasil ini mendukung argumen bahwa di pulau kecil, melalui kepadatan mangsa yang rendah, merupakan alasan paling sederhana untuk kemunculan bentuk-bentuk perbedaan populasi pada biawak Komodo. Pada akhirnya plastisitasdan atau perbedaan morfologi dan tingkah laku biawak Komodo diduga dapat meningkatkan kemampuan daya tahan hidup mereka di pulau kecil, dengan menyesuaikan ketersediaan mangsa, dan pada gilirannya secara energetik, yang secara khusus terlalu terhadap pemangsa besar lainnya.
Kata kunci: Mangsa besar; pulau; variasi mangsa pulau; seleksi alam; perbedaan fenotip.
Diterjemahkan oleh A Ariefiandy, D Purwandana, dan M J Imansyah (2007) dari artikel Jessop et al. 2007. Plastic dragons: A big predator’s responses to small islands.
Kutipan: Jessop, TS., Madsen T., Ciofi, C., Imansyah, M.J., Purwandana, D., Ariefiandy, A., Phillips, J.A. 2007. Biawak Komodo plastis: respon predator besar terhadap pulau kecil. Terjemahan. Ariefiandy, A., Purwandana, D., Imansyah, M.J. CRES-ZSSD/BTNK/TNC. Labuan Bajo, Flores, Indonesia.
PENDAHULUAN
Pada pulau, seleksi alam bekerja melalui variasi pulau terutama pada ketersediaan dan jenis mangsa, menyebabkan terjadinya seleksi alam sebagai contoh klasik perbedaan fenotip antar pulau dan pada akhirnya pembentukan struktur biota pulau (Case, 1978; Grant, 1998). Sebagai contoh, Predator besar berdarah panas, ketidakhadirannya di pulau kecil sangat jelas karena kebutuhan metabolismenya tidak didukung jaring makanan yang miskin mangsa dan sangat sederhana di pulau kecil (Burness et al. 2001). Di habitatnya kadal dan ular berdarah dingin, kebutuhan energinya hanya 20 kali lebih kecil dari pada predator puncak (Mc Nab 2002; Nagy et al. 1999; Nagy 2005).Meskipun demikian reptil dapat dipengaruhi oleh seleksi alam di pulau, disebabkan keterbatasan ataupun fluktuasi sumber makanan (Laurie & Brown 1990; Wikelski 1997). Sebaliknya, reptil kepulauan seperti vertebrata pulau lainnya dapat menunjukkan berbagai respon fenotip, termasuk perubahan bentuk tubuh yang mencolok untuk menyesuaikan kebutuhan energinya tehadap ketersediaan mangsa, dengan kata lain untuk meningkatkan daya tahan hidup (Case and Schwaner 1993; Boback
et al. 2003; Jessop et al. 2006; Wikelski 2005). Lebih lanjut populasi yang mendiami kepulauan
dapat dipengaruhi oleh perbedaan relatif relung makanan kepulauan yang dapat menghasilkan plastisitas fenotip yang nyata dan atau adaptasi lokal berkaitan pencernaan makanan (contohnya organ pencernaan, moncong, rahang, dan tulang kepala) (Grant,1999; Aubret and Shine, 2004). Biawak Komodo (Varanus komodoensis) merupakan kadal raksasa (berat mencapai 87 kg) dan memiliki kemampuan memangsa mamalia paling besar di habitatnya (dalam hal ini kerbau). Biawak Komodo terdapat di lima pulau di bagian timur Indonesia. Namun, seperti pulau endemik lainnya, Komodo nampaknya dipengaruhi seleksi alam yang berkaitan dengan makan, sebagai mana perbedaan populasi dalam hal ukuran tubuh maksimum (dalam hal ini empat variasi dalam bentuk tubuh) yang telah dipelajari di keempat populasi pulau yang mendiami Taman Nasional Komodo (Jessop et al., 2006). Ukuran tubuh Komodo merefleksikan kepadatan mangsa ungulata besar (dalam hal ini Rusa, Cervus timorensis) pada setiap pulau dan juga menunjukkan perbedaaan jenis mangsa yang terdapat diantara dua pulau besar kaya mangsa (270-380 km2) dengan dua pulau kecil miskin mangsa (< 11 km2) (Jessop et al., 2006). Dari pengamatan secara anekdot, kami mengetahui bahwa pola-pola variasi bentuk tubuh Biawak Komodo nampaknya mengikuti kepadatan independen rusa diantara keempat populasi pulau, sehingga tidak ada hubunganya dengan kekerabatan genetis diantara populasi. Sebagai contoh, komodo besar (dalam hal ini komodo dengan berat diatas 80 kg) ditemukan di pulau besar, Rinca, yang jaraknya hanya 800 meter terpisah oleh lautan dengan pulau kecil Nusa Kode, dimana hanya terdapat komodo-komodo kecil
(dengan berat tubuh lebih kecil dari 20 kg), (Gambar 1). Observasi awal ini menjadi dasar bagi penelitian yang sedang dilakukan, dan, secara khusus, penelitian ini difokuskan untuk dapat memahami keadaan yang saling mempengaruhi di antara mekanisme tambahan yang menyebabkan perbedaan populasi Komodo, juga untuk menentukan potensi efek aliran gen terhadap variasi morfologi dan perilaku pada kadal predator besar.
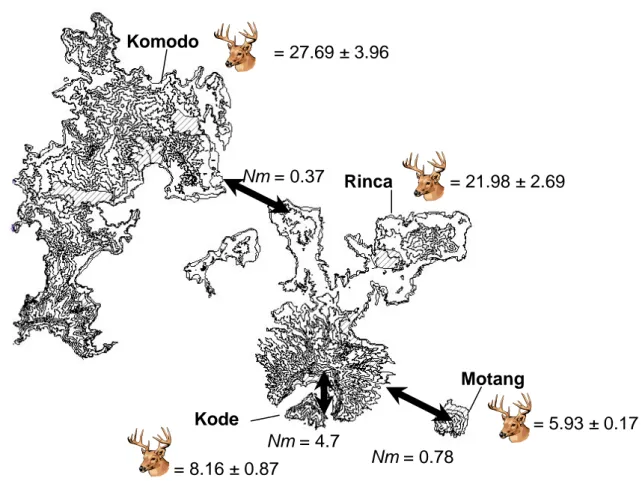
Nm = 0.37 Nm = 0.78 Nm = 4.7 = 27.69 ± 3.96 = 21.98 ± 2.69 = 5.93 ± 0.17 = 8.16 ± 0.87 Komodo Rinca Motang Kode
Gambar 1. Peta empat lokasi kajian di Tamana Nasional Komodo, menunjukkan kepadatan relatif mangsa Rusa (nilai tengah jumlah kotoran rusa per transek yang drata-ratakan dari 3 tahun penghitungan survey). Tanda panah menunjukkan tingginya nilai estimasi aliran gen (Nm) di antara dua populasi-pulau biawak Komodo. Daerah yang diarsir merupakan lokasi dan luas area kajian di setiap pulaunya.
Secara khusus kami berusaha untuk menemukan bukti tambahan (selain ukuran tubuh maksimum) mengenai perbedaan fenotip di antara populasi Komodo terkait alasan penentu keberadaan mereka di pulau kecil yang miskin mangsa tersebut. Bedasarkan dugaan kami, ketersediaan mangsa besar merupakan faktor seleksi alam yang menyebabkan perbedaan pada biawak Komodo. Kami menyeleksi beberapa ciri morfologi yang dapat mengindikasikan perbedaan sebagai refleksi variasi pada jenis dan ketersediaan mangsa tersebut. Perbedaan alat pencernaan termasuk bentuk kepala (panjang dan lebar), yang pada prinsipnya mempengaruhi struktur rahang reptil, dapat mewakili
dasar perbedaan fenotip sebagai mekaniseme untuk memudahkan kemampuan mendapatkan makan yang spesifik di pulau (Aubret and Shine, 2004), khususnya di pulau kecil dimana kepadatan rusa sangat rendah dan ketergantungan akan mangsa lebih kecil adalah sangat penting. Seleksi alam terkait makanan pada pulau sering kali nampaknya menunjukkan beban ketahanan hidup melalui ukuran tubuh yang besar (dan menyebabkan kekerdilan pada binatang ukuran besar) dan atau kebutuhan energetik yang tinggi (contohnya predator endoterm) (McNabb, 2004). Diantara spesies Varanus dengan ukuran panjang maksimum yang hampir sama, variasi berat tubuh menunjukkan perbedaan dalam pertumbuhan (sebagai contoh ringan hingga berat) secara umum sering teramati dan menunjukan kapasitas variasi tubuh dalam genus ini (Pianka, 1995). Pada V.
komodoensis, sebuah tingkat kesamaan variasi tubuh dapat mengarahkan kepada ektomorfisme
dengan pengurangan ukuran tubuh maksimal sehingga menurunkan berat relatif Biawak Komodo dan yang lebih penting adalahi kebutuhan energetik secara keseluruhan.
Pada jaring makanan yang sederhana, seperti danau di daerah empat musim, daya bertahan hidup ikan pemangsa tingkat atas (contohnya perch) terkait dengan kemampuannya untuk menggunakan interaksi pemangsa - mangsa yang fleksibel, termasuk peningkatan kanibalisme ketika sumber daya makanan alternatif menipis (Persson et al. 2003; Persson et al. 2004). Dalam sebuah cara yang sama, akankan jaring-jaring makanan sederhana di pulau kecil mengharuskan Biawak Komodo mengimbangi penurunan kepadatan mangsa dengan meningkatkan kanibalisme untuk memenuhi kebutuhan suplemen energetiknya? Dalam penemuan terdahulu, Auffenberg (1981) mencatat kanibalisme oleh Komodo dewasa di salah satu pulau besar, meskipun dalam frekuensi yang kecil. Hal ini menunjukkan bahwa perilaku tersebut merupakan salah satu bentuk interaksi pemangsa - mangsa.Apabila kanibalisme meningkat pada populasi pulau kecil, sebagai konsekuensi rendahnya kepadatan populasi rusa, hal tersebut dapat menghasilkan perbedaan antar pulau dalam tekanan predasi (selama disana tidak terdapat predator besar lainnya). Meningkatnya kewaspadaan sering digunakan sebagai pengukuran secara tidak langsung untuk tekanan predasi pada hewan vertebrata, dan disini kami menggunakannya untuk menilai perbedaan populasi dalam menduga perilaku yang merefleksikan ketersediaan mangsa diantara populasi-populasi kepulauan biawak Komodo. Penerapan pendekatan ini berdasarkan penelitian terdahulu yang menyebutkan bahwa vertebrata penggali di kepulauan (termasul kadal) meningkatkan kewaspadaan secara cepat ketika mendapatkan peningkatan tekanan predasi dari pemangsanya.(Stone et al. 1995; Gil-de-Costa et al. 2003).
Akhirnya, berdasarkan pola-pola berulang dalam ukuran tubuh maksimum antara pulau besar dan kecil (Jessop et al., 2006) kami menilai secara keseluruhan apakah perbedaan morfologi dan perilaku pada biawak Komodo paling berkaitan dengan kepadatan rusa di kepulauan, atau lebih mencerminkan kepada hubungan genetis. Di kepulauan, selain dari seleksi ekologis, populasi mungkin dapat berubah disebabkan karena proses acak termasuk pergeseran gen, inbreeding atau sejarah filogeni(Smith et al., 1997; Clegg et al., 2003). Jika pola-pola berulang yang sama di dalam perbedaan fenotip teramati diantara populasi biawak Komodo di pulau besar dan pulau kecil, meskipun perbedaan yang potensial dalam parameter genetis (dalam hal ini aliran gen), hal ini dapat medukung aturan seleksi alam dan memperkirakan bahwa gradien ekologi pada kepadatan mangsa besar sebagai faktor penting bagi perbedaan fenotip spesies ini.
METODE
(a) Prosedur penelitian lapangan
Biawak Komodo ditangkap dari sepuluh lokasi di empat pulau- Komodo (393.4 km2), Rinca (278.0 km2), Gili Motang (10.3 km2) dan Nusa Kode (9.6 km2) dalam kawasan Taman Nasional Komodo, Indonesia, pada tahun 2002-2006 (Gambar 1). Pada tahun 2002-2006 kami telah menangkap 498 komodo dari berbagai ukuran (tetasan tidak diikutkan) dengan menggunakan dua metode, yaitu perangkap yang diberi umpan dan tangkap tangan menggunakan tali laso (Jessop et al., 2006) Tujuh pengukuran morfologi: panjang mocong-kloaka “snout to vent length” (SVL), panjang ekor (komodo dengan ekor rusak tidak diikutkan), lingkar pangkal ekor (mendekati kloaka), lebar kepala maksimum, panjang kepala, lebar rahang atas (diukur pada lubang hidung) dan berat tubuh dicatat dengan menggunakan meteran plastik fleksibel, kaliper Vernier dan timbangan digital.
Kepadatan mangsa di hitung pada setiap pulau dengan mengunakan metode transek plot (terdapat dalam Jessop et al., 2006). Lokasi transek ditempatkan di lokasi yang sama dengan lokasi studi Mark Recapture biawak Komodo (dengan atau tanpa perangkap). Dalam penelitian ini kami menyajikan data perkiraan rata-rata kepadatan kotoran (rata-rata jumlah kelompok kotoran per transek) selama tiga tahun sebagai ukuran untuk mengukur kepadatan relatif rusa, yang merupakan sumber makanan utama bagi komodo dewasa (Auffenberg 1981), dan satu satunya mangsa ungulata yang hadir di keempat pulau (Jessop et al., 2006) (Gambar 1).
(b) Analisis morfologi dan tingkah laku
Analisis multivariasi covariansi (MANCOVA) digunakan untuk menguji apakah terdapat perbedaan signifikan untuk bentuk morfologi di keempat populasi-pulau V. komodoensis, yaitu ukuran tubuh independen (dalam hal ini panjang SVL). Kami menggunakan data yang ditransformasikan secara log untuk 6 variabel, yaitu panjang kepala, lebar kepala, lebar rahang atas, panjang ekor, lingkar pangkal ekor, dan berat tubuh untuk menilai perbedaan antar pulau. Untuk memenuhi asumsi MANCOVA, kami juga menguji untuk memastikan bahwa data kami tidak melanggar normalitas, linearitas, keseragaman regresi, multikolinearitas, dan keseragaman matriks variansi kovaransi. Untuk menilai perbedaan antar populasi pulau dalam tingkat kewaspadaan tingkah laku anti pemangsa, untuk mewakili predasi intraspesifik, kami mengukur variasi pulau dalam kelemahan tiap individu Komodo untuk dapat tertangkap dengan menggunakan tongkat bertali laso sepanjang 1,8 meter. Metode ini dapat digunakan untuk menangkap biawak Komodo semua ukuran dengan syarat komodo dapat didekati dan dilaso sebelum mereka melarikan diri. Sehingga perbedaan frekuensi relatif dalam metode tangkap tangan (dibandingkan dengan metode menggunakan perangkap) diantara populasi dapat digunakan untuk menduga perbedaaan dalam perilaku anti-pemangsa. Untuk menghindarkan terjadinya ulangan semu, maka hanya data dari tangkapan pertama saja yang digunakan dalam penelitian ini. Data perilaku ini telah dianalisis menggunakan Pearson chi-square
tests untuk independensi.
(c) Estimasi genetis dari aliran gen
Dalam kajian ini, kami menggunakan data genetik microsatelit dari penelitian terdahulu (Ciofi et al., 1999) dan dianalisis kembali untuk memperkirakan jarak dan polaritas aliran gen menggunakan pendekatan kemungkinan maksimal (the maximum likelihood approach) yang diimplementasikan dengan menggunakan perangkat lunak MIGRATE-1.6.9. MIGRATE meningkatkan pendekatan untuk memsukkan tingkat migrasi di antara populasi, dengan asumsi tinkat mutasi per-lokus adalah m, dan nilai Nm mewakili tingkat migrasi per generasi antar pulau yang diteliti (gambar 1) dan dilaporkan untuk skala arah yang lebih besar antara masing-masing pasangan dari pulau-pulau yang berdekatan.
Data genetik tersebut digunakan untuk menghitung aliran gen, dari Komodo di lokasi penelitian yang sama pada tahun 1994. Sehingga ketika estimasi tingkat populasi sepertinya tidak mewakili seluruh
binatang yang digunakan pada penelitian ini, kami menduga bahwa perbedaan waktu diantara kedua penelitian masih tetap dalam kurun waktu satu generasi(≈ 10-12 tahun untuk betina; Jessop
data yang tidak dipublikasikan), dan bahwa tidak ada bukti yang menyatakan perbedaan mendasar
dalam genetika populasi, data ini masih tetap dapat menghasilkan data estimasi yang cukup valid untuk parameter genetika populasi diantara populasi-populasi yang masih ada.
(d) Estimasi hubungan antar data
Perbedaan antar pulau, dengan penekananpada morfologi Komodo (menggunakan 7 ciri morfologi termasuk SVL) dan tingkah laku, dihitung dari data dengan menggunakan jarak nilai tengah
“Euclidean” multi demensi (multidimensional Euclidean distance means) (menggunakan SPSS 13)
menghasilkan matrik individu dari 6 nilai berpasangan. Matrik-matrik tersebut dibandingkan dengan perbedaan berpasangan pada kepadatan mangsa (yang juga diperoleh menggunakan jarak Euclidean) mengunakan analisis regresi dan ANOVA untuk menguji korelasi antara bentuk morfologi dan perilaku terhadap kepadatan mangsa rusa kepulauan. Untuk menyimpulkan adanya perbedaan antar populasi pada perubahan ciri biawak Komodo berkorelasi dengan hubungan genetis, dilakukan pembandingan antara data matrik morfologi dan perilaku dengan aliran gen berpasangan (Nm) mengunakan uji Mantel (dihitung denan program Poptools).
HASIL
(a) Variasi ukuran morfologi yang independen
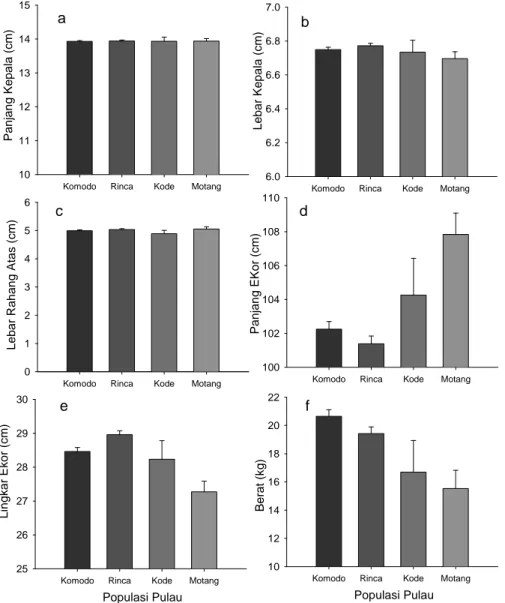
Hasil uji statistik MANCOVA (Wilk’s Lambda) sangat signifikan (F15, 1350.32 = 4.667; P< 0.0001) menunjukan adanya perbedaan ukuran pada biawak Komodo yang nyata untuk ukuran morfologi independen. Analisis univariasi pada keenam ciri-ciri morfologi independen menunjukkan sebagai berikut, lingkar ekor (ANCOVA: F 3,493 = 10.78, P < 0.001), tail length (ANCOVA: F 3,493 = 8.54, P = 0.001), lebar kepala(ANCOVA: F 3,493 = 2.69, P = 0.046),dan berat (ANCOVA: F 3,493 = 10.095, P = 0.001) adalah, masing-masing, berbeda secara nyata antar populasi, sedangkan panjang kepala (ANCOVA: F 3,493 = 0.635, P = 0.59)dan lebar rahang atas (ANCOVA: F 3,255 = 0.825, P = 0.48) tidak berbeda secara nyata.Perbedaan yang nyata pada bentuk morfologi menunjukkan pada dasarnya populasi pulau kecil Gili Motang dan Nusa Kode (Gambar2.) mempunyai bentuk ekor yang relatif lebih panjang dan kecil (lingkar ekor) bersamaan dengan mengecilnya tubuh dibandingkan dengan
populasi di kedua pulau besar. Lebih lanjut Biawak Komodo Gili Motang menunjukkan ukuran lebar kepala yang sedikit lebih kecil tetapi relatif cukup nyata terhadap populasi lainnya.
(b) Perilaku menghindari pemangsa
Kemampuan menangkap Komodo menggunakan tongkat laso berbeda secara nyata pada tiap populasi (χ2 3, 544 = 36.85; p < 0.001). Di pulau kecil Nusa Kode (1%) dan Gili Motang (0%), hampir tidak mungkin untuk mendekati dan menangkap langsung Komodo karena tingginya tingkat kewaspadaan dibandingkan dengan Pulau besar Rinca (34%) and Komodo (26%) (Gambar 3).
Komodo Rinca Kode Motang
Panj ang Kepala (cm ) 10 11 12 13 14 15 X Data
Komodo Rinca Kode Motang
Lebar Kepala (cm ) 6.0 6.2 6.4 6.6 6.8 7.0 X Data
Komodo Rinca Kode Motang
T a il le n g th (c m) 100 102 104 106 108 110 Populasi Pulau Komodo Rinca Kode Motang
Lingkar Ekor (cm ) 25 26 27 28 29 30 Populasi Pulau Komodo Rinca Kode Motang
B e rat (k g) 10 12 14 16 18 20 22
Komodo Rinca Kode Motang
Lebar Rahang Atas (cm
) 0 1 2 3 4 5 6 Panj ang E K or (cm ) a b c d e f
Gambar 2. Estimasi nilai tengah marginal dari enam ciri morfologi independen yang meggambarkan perbedaan relatif pada empat populasi pulau yang diteliti. Nilai tengah marginal menyajikan perbedaan relatif ciri morfologi Komodo yang distandarisasikan menjadi ukuran SVL yang seragam, yaitu 81,64 cm
Populasi Pulau
Komodo Rinca Kode Motang
qy p Frek uens i Relatif Ta ngkap Tangan (%) 0 10 20 30 40
Gambar 3. Perbedaan relatif dalam frekuensi tangkap tangan (tangkap langsung tanpa
menggunakan perangkap), sebuah penghitungan relatif untuk menunjukkan kewaspadaan di keempat populasi-pulau biawak Komodo. Frekuensi yang lebih rendah menandakan penurunan kapasitas biawak Komodo untuk mendekat dan ditangkap langsung sebelum komodo tersebut melarikan diri. Pada dasarnya di dua pulau kecil hampir tidak mungkin menangkap langsung biawak Komodo dibandingkan dengan di dua pulau besar.
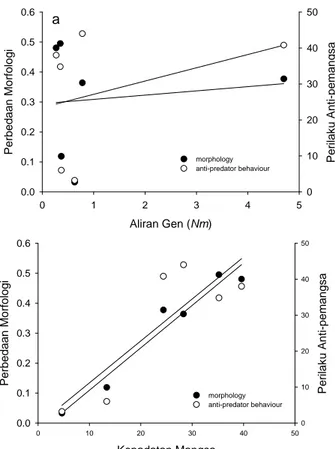
Aliran Gen (Nm) 0 1 2 3 4 5 Per beda an Mo rfolo gi 0.0 0.1 0.2 0.3 0.4 0.5 0.6 P e rilak u A n ti -pem an gsa 0 10 20 30 40 50 morphology anti-predator behaviour a Kepadatan Mangsa 0 10 20 30 40 50 Per bed aan Mo rfo logi 0.0 0.1 0.2 0.3 0.4 0.5 0.6 P e rilak u Ant i-pe m an gsa 0 10 20 30 40 50 morphology anti-predator behaviour
Gambar 4. Perbedaan morfologi (menggunakan 7 karakter morfologi) dan tingkah laku, sebagai standarisasi jumlahmigran (Nm) (a) dan kepadatan mangsa relatif (b) di antara pula-pulau. Kami menghitung karakter perbedaan ciri-ciri sebagai nilai berpasangan absolut dari perbedaan jarak euclidean, dalam dan antar tipe habitat. Kami menyajikan garis regresi sebagai ilustrasi saja, sebagaimana kami menggunakan korelasi matriks berpasangan (uji Mantel) untuk menghitung perbandingan non independen.
(c) Korelasi antara morfologi, mangsa dan aliran gen
Perbandingan berpasangan antara morfologi (R = ; ANOVA F1,4 = 80.46, P = 0.001) dan perbedaan anti-pemangsa (R = ; ANOVA F1,4 = 80.46, P = 0.001) pada populasi biawak Komodo dan kepadatan rusa kepulauan menujukkan korelasi yang nyata (Gambar 4a).
Sebaliknya tidak terdapat korelasi yang nyata antara aliran gen terhadap bentuk morfologi (uji Mantel untuk perbandingan matriks: R = 0.123, P = 0.50) atau perbedaan perilaku diantara keempat populasi pulau (Gambar 4b).
PEMBAHASAN
a) Perbedaan morfologi pada biawak Komodo
Perbedaan jenis mangsa antar pulau, ketersediaan mangsa antar pulau, dan kompetisi intraspesifik sudah diketahui secara jelas sebagai faktor seleksi penentu plastisitas fenotip, perubahan populasi (dan pada akhirnya spesiasi) dan pembentukan struktur komunitas dalam habitat kepulauan (Simpson, 1953; Grant, 1999, Schluter, 2000; Streelman & Danley, 2003). Hasil penelitian kami sangat menunjukan bahwa ketersediaan mangsa besar merupakan faktor yang sangat kuat dalam seleksi pada perubahan secara fenotip biawak Komodo, meskipun dalam kondisi aliran gen yang sangat tinggi (contohnya antara pulau Rinca dan Pulau Nusa Kode). -Pertambahan ukuran panjang ekor, penurunan lingkar ekor dan penurunan kondisi tubuh merupakan bukti bahwa dua populasi pulau menempati pulau kecil yang kurang mangsa. Perbedaan-perbedaan dalam proposi tubuh, dibandingkan dengan populasi biawak Komodo di pulau besar yang kaya akan mangsa, mengarahkan Komodo kepada perencanaan bentuk tubuh yang ektomorfik bersamaan dengan seluruh pengurangan dalam ukuran tubuh, kemudian lebih lanjut berperan dalam menurunkan kebutuhan energetik individu secara keseluruhan.
Ukuran kepala secara keseluruhan relatif sama diantara populasi dan diperkirakan tidak seperti vertebrata kepulauan lainnya yang terbiasa untuk makanan yang berbeda, bentuk kepalanya relatif stabil. Dalam bagian kesamaan proporsi kepala antara pulau dapat merefleksikan bahwa pada saat kepadatan mangsa besar sangat berkurang apabila di bandingkan dengan pulau kecil, rusa masih dapat ditemukan dan masih tetap menyediakan makanan bagi komodo besar, sehingga membatasi seleksi dalam modifikasi bentuk kepala. Bagaimanapun juga, lebar kepala Komodo untuk pulau yang
paling terisolasi (Gili Motang), relatif berkurang secara nyata dibandingkan dengan ketiga populasi lainnya. Kami menduga bahwa dengan pengurangan ukuran lebar kepala menjadi kecil dan tidak terjadi pada populasi pulau kecil lainnya dapat menjadi konsisten dengan proses melalui pergeseran gen.
(b) Perbedaaan tingkah laku pada biawak Komodo
Perbedaan tingkah laku yang ditunjukkan dengan adanya peningkatan kewaspadaan pada populasi biawak Komodo di pulau kecil yang miskin mangsa adalah konsisten dengan penemuan pada spesies lain sebagai respon terhadap peningkatan tekanan predasi (Cooper 2003; Heithaus et al.
2002; Stone et al. 1994). Kurangnya predator lain sangat menunjukkan predasi intraspesifik oleh Komodo yang lebih besar menjadi faktor utama perbedaan mencolok antara populasi Komodo di pulau besar kaya mangsa dan pulau kecil miskin mangsa. Meningkatnya kewaspadaan pada biawak Komodo di pulau kecil diduga menjadi perilaku anti pemangsa yang sangat penting dan diperlukan untuk mengurangi tingkat predasi intraspesifik. Secara sederhana, ketika kepadatan mangsa menurunkan proses kanibalisme, maka diharapkan akan meningkatnya interaksi pemangsa dan mangsa yang umum di antara populasi-pulau biawak Komodo. Ketika kanibalisme nampaknya sangat jarang pada binatang karnivor besar, sebaliknya pada pemangsa vertebrata kecil, terutama pada beberapa jenis ikan kanibalisme justru sangat umum dan bahkan dapat mempengaruhi secara dramatis dinamika dan keberadaan populasi (Persson et al. 2003; Persson et al. 2004). Pada beberapa mekanisme secara khusus dapat menimbulkan kanibalisme untuk meningkatkan daya tahan hidup individu dan untuk menjaga keberadaan populasi (Persson et al., 2004). Meningkatnya predasi intraspesifik menghasilkan pendapatan energetik yang mungkin mengakibatkan populasi bertahan dibawah kondisi dimana mangsa alternatif sangat rendah dan menyebabkan populasi non-kanibal dapat menjadi punah, sebuah fenomena yang biasa disebut life-boat mechanisme (Henson 1997; Persson et al. 2004). Lebih lanjut, kanibalisme dapat kembali menstabilkan dinamika populasi dan memperkecil fluktuasi populasi (Persson et al. 2004), kedua faktor tersebut dapat menjadi hal yang sangat penting dalam mempertahankan populasi pulau.
Sementara proses evolusi terarah dan adaptasi lokal konsisten dengan seleksi alam yang berkaitan dengan makanan sering kali dianggap sebagai penyebab perbedaan fenotip pada taxa di pulau, proses alternatif termasuk pergeseran gen, inbreeding atau sejarah filogenetiknya dapat menjadi penyebab terjadinya perubahan bentuk populasi (Smith et al., 1997; Clegg et al., 2003).Pada kasus lainnya kapasitas perbedaan fenotip, dapat terjadi melalui seleksi alam maupun proses acak (contoh
pergeseran), secara khusus dipengaruhi oleh isolasi geografi (dan demikian juga genetis) suatu populasi (Mayr 1963). Teori memperkirakan aliran gen dapat mendesak perbedan adaptif ada populasi yang mendiami habitat yang berbeda secara ekologis dan pada umumnya teori tersebut didukung oleh studi-studi empiris. (contohnya: Haldane 1948; Slatkin 1973; Endler 1977; Hendry et al. 2004).Akan tetapi beberapa penelitian menyangkal pentingnya aliran gen dalam penyeragaman perbedaan dan mencatat seleksi alam sudah cukup kuat, perbedaan sering kali terjadi ketika aliran gen pada tingkat yang cukup tinggi (misalnya Smith et al., 1997; Schneider et al., 1999; Brown et al,. 2001; Calsbeek & Smith 2003). Pada penelitian kami, tidak ditemukan adanya korelasi antara ciri-ciri morfologi dan perilaku yang diduga berkaitan dengan adanya aliran gen.
(c) Kesimpulan Penutup
Pada keempat populasi pulau biawak Komodo, teramati dua pola berulang dari perbedaan morfologi dan perilaku yang merefleksikan kepadatan mangsa besar kepulauan serta kekerabatan genetis independen.Di pulau yang kaya akan mangsa ungulata (Komodo dan Rinca) Komodo tidak hanya lebih besar ukurannya (Jessop et al., 2006), akan tetapi lebih kuat dan berani dari pada populasi Komodo di pulau kecil yang miskin mangsa (Nusa Kode and Gili Motang). Dengan demikian, seleksi alam pada pulau-pulau kecil ini, aliran gen independen,bekerja melalui variasi ketersedian mangsa, dan sepertinya menjadi faktor penting yang mengarahkan proses evolusi dan ekologi untuk memecahkan masalah berkaitan dengan ukuran tubuh Komodo yang besar. Perbedaan ukuran tubuh maksimum yang terjadi empat kali lipat pada Biawak Komodo sepertinya menunjukkan salah satu mekanisme penting untuk menyesuaikan ukuran individu terhadap mangsa pulau lokal (Jessop
et al. 2006). Namun, perbedaan lebih lanjut dalam morfologi dan perilaku, apakah itu fenotip maupun
mikroevolusi atau kombinasi kedua respon tersebut (sebagai contoh adaptasi plastsitas fenotip) adalah sejalan dengan penurunan ukuran tubuh maksimum. Hal tersebut dapat memungkinkan Komodo tidak memerlukan sumber daya nutrisi yang tidak tersedia, dan kedua, akan mengurangi kebutuhan energi secara keseluruhan di pulau kecil. Penelitian lebih lanjut diperlukan untuk mengetahui tambahan demografi, mekanisme fisiologi dan perilaku yang juga dapat menghasilkan bukti bagaimana kadal/biawak memecahkan dilema pemangsa besar pada pulau kecil.
UCAPAN TERIMA KASIH
Terima kasih kepada para Staf Taman Nasional Komodo yang telah membantu pelaksanaan penelitian di lapangan. Penelitian ini dilakukan sebagai program kerja sama antara Zoological Society of San Diego, The Nature Conservancy (Indonesia program) dan Direktorat Jenderal Perlindungan Hutan dan Konservasi Alam (PHKA). Dukungan dana disediakan oleh Millenium post-doctoral fellowship dari Zoological Society of San Diego (untuk T. S. Jessop), dan juga (untuk C. Ciofi) disediakan oleh American Zoo and Aquarium Association, The European Association of Zoos and Aquaria. The Offield Family Foundation juga menyediakan dukungan dana tambahan untuk proyek penelitian ini.
DAFTAR PUSTAKA
Auffenberg, W. 1981. The Behavioral Ecology of the Komodo Monitor. Gainesville: FloridaUniversity Press.
Boback, S. M. 2003. Body size evolution in snakes: evidence from island populations. Copeia, 81– 94.
Case, T. J. & Schwaner, T. D. 1993. Island/mainland body size differences in Australian varanid lizards. Oecologia94, 102-109.
Ciofi, C., & Bruford, M.W. 1999. Genetic structure and gene flow among Komodo dragon populations inferred by microsatellite loci analysis. Mol. Ecol.8, S17-S30.
Ciofi, C. & de Boer, M. E. 2004. Distribution and conservation of the Komodo monitor (Varanus
komodoensis). Herpetol.J.14, 99-107.
Claessen, D., de Roos A.M., & Persson, L. 2000. Dwarfs and giants: Cannibalism and competition in size-structured populations. Amer. Natur.155, 219-237.
Cooper, W. E. Jr. 2003. Rffect of risk on aspects of escape behavior by a lizard, Holbrookia
propinqua, in relation to optimal escape theory. Ethology109, 617-626.
Gil-da-Costa, R., Palleroni, A., Hauser, M., Touchton, J. & Kelley, J. P. 2003. Rapid acquisition of an alarm response by a neotropical primate to a newly introduced avian predator. Proc. R. Soc.
Lond. B270, 605-610.
Heithaus, M. R., Frid, A. & Dill, L. M. 2002. Shark-inflicted injury frequencies, escape ability and habitat use of green and loggerhead turtles. Mar. Biol.140, 229–236.
Jessop, T. S., Madsen, T., Sumner, J., Rudiharto, H., Phillips, J. A. & Ciofi C. 2006. Maximum body size among insular Komodo dragon populations covaries with large prey density. Oikos. Laurie, W. A., Brown, D. 1990. Population Biology of Marine Iguanas (Amblyrhynchus cristatus). 11.
Changes in the annual survival rates and the effects of size, sex, age and fecundity in a population crash. J. Anim. Ecol.59, 529-544.
McNab, B. K. 2002. Minimizing energy expenditure facilitates vertebrates persistence on oceanic islands. Ecol. Lett.5, 693-704.
Nagy, K.A., Girard, I. A. & Brown, T. K. 1999. Energetics of free-ranging mammals, reptiles, and birds. Annu. Rev. Nutr. 19, 247-77.
Nagy, K. A.. 2005. Field metabolic rate and body size. J. Exp. Biol. 208, 1621-1625.
Murdoch, W. W. 1969. Switching in generalist predators: experiments on predator specificity and stability of prey populations. Ecol. Monogr.39, 335-354..
Persson, L., de Roos, A.M., Claessen, D., Byström, P., Lövgren, J., Sjögren, S., Svanbäck, R., Wahlström, E. & Westman, E. 2003. Gigantic cannibals driving a whole-lake trophic cascade. Proc. Natl. Acad. Sci.100, 4035-4039.
Persson, L., Claessen, D. de Roos, A.M., Byström, P., Sjögren, S., Svanbäck, R., Wahlström E. &. Westman, E. 2004. Cannibalism in a size-structured population – energy extraction and control. Ecol. Mon.74, 135-157.
Purvis, A., Gittleman, J. L., Cowlishaw, G., & Mace, G. M. 2000. Predicting extinction risk in declining species. Proc. R. Soc. Lond. B267, 1947-1952.
Stone, P. A., Snell, H. L. & Snell, H. M. 1994. Behavioral Diversity as Biological Diversity: Introduced Cats and Lava Lizard Wariness. Cons.Biol.8,569-573.
Thomson, W. L., White, G. C. & Gowan, G. V. 1998. Monitoring vertebrate populations. Academic Press.
Wikelski, M., Carrillo, V. & Trillmich, F. 1997. Energy limits to body size in a grazing reptiles, the Galapagos marine iguana. Ecology78, 2204-2217.
Wikelski, M. 2005. Evolution of body size in Galapagos marine iguans. Proc. R. Soc. B272, 1985-1993.