EVALUASI Aspergillus PENGHASIL ENZIM HIDROLITIK DAN ANTAGONISMENYA
TERHADAP Fusarium oxysporum f.sp. cubense (Foc)
Suciatmih
Bidang Mikrobiologi, Puslit Biologi – LIPI, CSC Jl. Jakarta – Bogor Km 46, Cibinong, Bogor suciatmih2008@yahoo.ca
ABSTRAK
Evaluasi Aspergillus penghasil enzim hidrolitik dilakukan untuk mencari agen antijamur terhadap
Fusarium oxysporum f.sp. cubense (Foc), penyebab penyakit panama (layu fusarium) pada Musa spp.
Identifikasi jamur Aspergillus dilakukan secara morfologi, sedangkan evaluasi enzim kitinase, lipase, dan
protease dilakukan dengan menggunakan media spesifik. Uji antagonisme antara Aspergillus dan patogen
Foc menggunakan metode kultur ganda. Lima puluh tiga (45,7 %) dan 63 (54,3 %) dari 116 isolat Aspergillus yang dikoleksi dari berbagai subtrat masing-masing berhasil diidentifikasi sampai spesies dan
genus. Aspergillus yang sudah diidentifikasi sampai spesies adalah satu isolat sebagai A. clavatus, 3 isolat
sebagai A. chevalieri, 1 isolat A. flavipes, 1 isolat sebagai A. flavus, 10 isolat sebagai A. fumigatus, 4
isolat sebagai A. japonicus, 2 isolat sebagai A. kanagawaensis, 3 isolat sebagai A. neoglaber, 23 isolat
sebagai A. niger, dan 5 isolat sebagai A. terreus. Semua isolat jamur tidak menunjukkan aktivitas kitinase,
tetapi 45,7 %, 20,7 %, dan 11,2 % masing-masing mempunyai aktivitas lipase dan protease; lipase serta
protease. Hanya 22,4 % isolat jamur tidak mempunyai aktivitas lipase atau protease. Tiga contoh isolat Aspergillus (A. niger 917, A. terreus 583, dan Aspergillus sp. 920) yang mempunyai aktivitas lipase dan protease tinggi dapat menghambat pertumbuhan patogen Foc sekitar 21,67–36,81 %.
Kata Kunci: antagonisme, Aspergillus, Fusarium oxysporum f.sp. cubense (Foc), enzim hidrolitik
PENDAHULUAN
Salah satu ancaman utama tanaman pisang di seluruh dunia adalah penyakit layu fusarium (Vuylsteke et al., 1996). Penyakit tersebut disebabkan oleh Fusarium oxysporum f.sp. cubense (Foc) melalui infeksi sistemik vaskuler (Snyder & Hanson, 1940). Foc, seperti
kebanyakan isolat patogen F. oxysporum, mengolonisasi
jaringan vaskuler, menyebabkan gangguan translokasi air ke pucuk tanaman (Beckman & Roberts, 1995). Di Indonesia, penyakit ini dilaporkan telah menyebar hampir seluruh daerah pertanaman pisang. Sekitar 1,300 ha pisang barangan rusak berat akibat serangan Foc di
Sumatera Utara (Jumjunidang et al., 2005).
Pengendalian Foc menggunakan fungisida sintetis ke dalam tanah tidak efektif karena propagul jamur yang berdistribusi ke dalam tanah seringkali di luar jangkauan fungisida (Campbell, 1989). Selain itu, aplikasi fungisida dapat membunuh organisme bukan target, menyebabkan timbulnya strain patogen yang resisten terhadap fungisida, dan dapat membahayakan kesehatan serta
lingkungan (Djatnika et al., 2003). Oleh karena itu perlu
dicari alternatif pengendalian Foc yang aman dan ramah lingkungan. Salah satu pendekatan alternatif yang menjanjikan adalah dengan menggunakan jamur saprofit
seperti Aspergillus.
Aspergillus spp. adalah jamur yang hidup bebas dan ditemukan di berbagai substrat, seperti akar, batang, biji, buah, bunga, daun, umbi, bahan bakar, dan tanah rizosfer. Jamur tersebut dapat menjadi oportunis, simbion tanaman tidak virulen, dan parasit pada jamur lain. Aspergillus spp. digunakan untuk mengendalikan jamur
patogen tanaman melalui mikoparasitisme, lisis
miselium, dan antibiosis melalui sintesis metabolit volatil dan atau non-volatil (Bhattacharyya & Jha, 2011). Selulase, kitinase, pektinase, protease, dan lipase adalah enzim yang bertanggung jawab untuk melisis dinding sel
jamur (Smitha et al., 2014).
Spesies Aspergillus telah dilaporkan dapat
mengendalikan banyak jamur patogen, seperti Fusarium
acuminafum, F. equiseti, F. oxysporum f.sp. ciceri, F. monilliforme, F. sambucinum, F. solani, Macrophomia phaseolina, dan Rhizoctonia solani pada tanaman chickpea (Dolar, 2011), Fusarium oxysporum f. sp. lycopersici (Alwathnani & Perveen, 2012), Fusarium oxysporum var cumini (Sharma, 2012), dan F. solani f.sp. melongenae (Najar et al., 2011).
Penelitian ini bertujuan mengevaluasi aktivitas enzim hidrolitik, seperti kitinase, lipase, dan protease
secara in vitro dari Aspergillus koleksi LIPI-MC yang
Aspergillus penghasil aktivitas enzim hidrolitik tinggi kemudian diuji antagonismenya melawan patogen Foc. METODE
Identifikasi Aspergillus
Seratus enam belas isolat Aspergillus koleksi
LIPI-MC diidentifikasi secara morfologi menggunakan
panduan genus Aspergillus (Raper & Fennell, 1965).
Spesies Aspergillus dibiakan pada dua media berbeda,
yaitu Czapek Solution Agar (CZA) and Malt Extract
Agar (MEA). Setelah tujuh hari diinkubasi, koloni jamur
diamati karakter makroskopisnya, seperti diameter
koloni, eksudat, sebalik koloni; dan karakter
mikroskopisnya meliputi konidiofora, vesikel, metula, fialid, dan konidia.
Deteksi enzim hidrolitik
Deteksi aktivitas enzim hidrolitik Aspergillus,
seperti kitinase dilakukan dengan metode Lingappa &
Lockwood (1962), lipase dilakukan dengan metode
Peterson & Johnson (1949), serta protease dilakukan
dengan metode Uria et al. (unpublished).
Evaluasi antagonisme
Evaluasi antagonisme Aspergillus (penghasil
enzim hidrolitik tinggi) terhadap jamur patogen Foc
dilakukan dengan metode kultur ganda. Diameter jamur patogen diukur pada hari ke-5 setelah inokulasi, dan hambatan pertumbuhannya dihitung berdasarkan rumus Skidmore & Dickinson (1976):
PI = C – T x 100 C
PI = hambatan pertumbuhan miselium (%)
C = diameter miselium patogen pada cawan petri kontrol (cm)
T = diameter miselium patogen pada cawan petri perlakuan (cm)
HASIL
Empat puluh dua isolat Aspergillus diisolasi dari
tanah rizosfer (Tabel 1 dan 4). Sembilan belas (45,2 %)
isolat Aspergillus berhasil diidendifikasi sampai spesies,
sedangkan 23 (54,8 %) isolat sampai genus. Aspergillus
yang sudah diidentifikasi sampai spesies adalah satu
isolat sebagai A. flavipes, 4 isolat sebagai A. fumigatus, 4
isolat sebagai A. japonicus, 2 isolat sebagai A. neoglaber,
6 isolat sebagai A. niger, dan 2 isolat sebagai A. terreus.
Tabel 1.Aspergillus yang diisolasi dari tanah rizosfer
No Aspergillus Tanah rizosfer
Bambu Jagung Mangrove Pisang Tebu Teh Tomat Ubi
kayu 1. A. flavipes - - 1 - - - - - 2. A. fumigatus - - 3 - - - 1 - 3. A. japonicus 1 - 3 - - - - - 6. A. neoglaber - - 2 - - - - - 7. A. niger 1 - - 1 - - 2 2 8. A. terreus - - 2 - - - - - 9. Aspergillus 1 1 4 1 4 3 3 6 Total isolat 3 1 15 2 4 3 6 8
Keterangan: - = tidak ditemukan
Empat puluh tujuh isolat Aspergillus diisolasi dari berbagai bahan (Tabel 2 dan 5). Sembilan belas (40,4 %)
isolat Aspergillus berhasil diidendifikasi sampai spesies, sedangkan 28 (59,6 %) isolat sampai genus. Aspergillus yang
sudah diidentifikasi sampai spesies adalah satu isolat masing-masing sebagai A. chevalieri, A. clavatus, A. flavus, A.
kanagawaensis, A. neoglaber, dan A. terreus; 4 isolat sebagai A. fumigatus, dan 9 isolat sebagai A. niger. Tabel 2.Aspergillus yang diisolasi dari berbagai bahan
No Aspergillus
Bahan
Avtur Kain Kompos
Kon-taminan Minyak tanah Tanah hutan Tanah KRI Tanah Pantai 1. A. chevalieri - - 1 - - - - - 2. A. clavatus - - - 1 - - - - 2. A. flavus - - - - 1 - - 3. A. fumigatus 1 - - 3 - - - - 5. A. kanagawaensis - - - 1 - - 6. A. neoglaber - - - 1 - -
No Aspergillus
Bahan
Avtur Kain Kompos
Kon-taminan Minyak tanah Tanah hutan Tanah KRI Tanah Pantai 7. A. niger - - - 4 3 1 1 - 8. A. terreus - - - - 1 - - - 9. Aspergillus - 3 2 1 4 11 6 1 Total isolat 1 3 3 9 8 15 7 1
Keterangan: KRI = Kebun Raya Indonesia; - = tidak ditemukan
Dua puluh tujuh isolat Aspergillus diisolasi dari organ tanaman (Tabel 3 dan 6). Lima belas (55,6 %) isolat
Aspergillus berhasil diidendifikasi sampai spesies, sedangkan 12 (44,4 %) isolat sampai genus. Aspergillus yang sudah
diidentifikasi sampai spesies adalah dua isolat masing-masing sebagai A. chevalieri, A. fumigatus, dan A. terreus; 1
isolat A. kanagawaensis, serta 8 isolat sebagai A. niger.
Tabel 3.Aspergillus yang diisolasi dari organ tanaman
No Aspergillus Organ tanaman
Akar pisang Batang kopi Biji kedelai Buah tomat Bunga tomat Daun Umbi bawang merah 1. A. chevalieri 2 - - - - 2. A. fumigatus 2 - - - - 3. A. kanagawaensis - - 1 - - - - 4. A. niger 2 - 1 1 1 3 - 5. A. terreus - - 1 - - 1 - 6. Aspergillus 2 3 4 - - 2 1 Total isolat 8 3 7 1 1 6 1
Keterangan: - = tidak ditemukan
Semua (42) isolat Aspergillus yang diisolasi dari tanah rizosfer diketahui tidak terdeteksi menghasilkan enzim
kitinase, 28 (66,7 %) isolat Aspergillus menghasilkan enzim lipase, 26 (61,9 %) menghasilkan enzim protease, dan 10
(23,8 %) isolat Aspergillus tidak menghasilkan enzim lipase maupun protease (Tabel 4). Indeks lipase dihasilkan oleh
24 (57,1 %) isolat Aspergillus yang besarnya sekitar 1,11–2,33, sedangkan 4 (9,5 %) isolat Aspergillus hanya terdeteksi
positif menghasilkan lipase. Indeks protease dihasilkan oleh 17 (40,5 %) isolat Aspergillus yang besarnya sekitar 1,06–
1,67, sedangkan 9 (21,4 %) isolat hanya terdeteksi positif menghasilkan protease. Indeks lipase (2,33) dan protease
(1,67) tertinggi dihasilkan masing-masing oleh Aspergillus (514) dan Aspergillus (583). Ke dua Aspergillus tersebut
diisolasi dari tanah mangrove.
Tabel 4. Aktivitas enzim Aspergillus yang diisolasi dari tanah rizosfer No Aspergillus No koleksi Habitat Aktivitas kitinase Indeks lipase Indeks protease 1. A. fumigatus 501 Mangrove - 2,0 1,57 2. A. neoglaber 502 Mangrove - 1,2 1,11 3. Aspergillus 506 Mangrove - 2,0 1,63 4. A. fumigatus 509 Mangrove - 1,2 1,57 5. A. fumigatus 510 Mangrove - 1,14 1,5 6. A. japonicus 511 Mangrove - - - 7. Aspergillus 514 Mangrove - 2,33 1,13 8. Aspergillus 526 Mangrove - 1,14 1,5 9. A. neoglaber 580 Mangrove - - 1,63 10. A. terreus 583 Mangrove - 2,0 1,67 11. A. flavipes 584 Mangrove - - 1,25 12. A. japonicus 611 Mangrove - + - 13. A. japonicus 612 Mangrove - - - 14. Aspergillus 619 Mangrove - 1,4 1,13 15. A. terreus 623 Mangrove - + - 16. A. niger 692 Pisang - - -
No Aspergillus No koleksi Habitat Aktivitas kitinase Indeks lipase Indeks protease
17. Aspergillus 763 Ubi kayu - 2,14 1,14
18. Aspergillus 769 Pisang - 1,33 1,25
19. Aspergillus 811 Ubi kayu - 1,33 1,14
20. A. niger 815 Ubi kayu - - -
21. Aspergillus 824 Ubi kayu - + 1,45
22. Aspergillus 826 Ubi kayu - 1,91 -
23. Aspergillus 830 Ubi kayu - 1,2 +
24. A. niger 908 Ubi kayu - - -
25. Aspergillus 968 Jagung - 1,33 - 26. Aspergillus 996 Tebu - - 1,06 27. Aspergillus 1505 Tebu - 1,18 1,13 28. Aspergillus 1508 Teh - 1,11 + 29. Aspergillus 1525 Tebu - - - 30. Aspergillus 1526 Teh - + +
31. Aspergillus 1529 Ubi kayu - - -
32. Aspergillus 1533 Teh - 1,86 + 33. Aspergillus 1535 Tebu - 1,17 + 34. Aspergillus 1565 Bambu - 2,1 + 35. A .japonicus 1577 Bambu - - + 36. A. niger 1580 Bambu - - - 37. A. niger 2240 Tomat - - - 38. A. niger 2241 Tomat - - - 39. Aspergillus 2270 Tomat - 1,29 - 40. A. fumigatus 2273 Tomat - 1,38 + 41. Aspergillus 2275 Tomat - 1,67 + 42. Aspergillus 2276 Tomat - 1,33 -
Keterangan: - = tidak terdeteksi; + = terdeteksi
Semua (47) isolat Aspergillus yang diisolasi dari
berbagai bahan diketahui tidak terdeteksi menghasilkan
enzim kitinase, 33 (70,2 %) isolat Aspergillus
menghasilkan enzim lipase, 28 (59,6 %) isolat Aspergillus menghasilkan enzim protease, dan 8 (17,0 %)
isolat Aspergillus tidak terdeteksi menghasilkan enzim
lipase maupun protease (Tabel 5 & Gambar 1). Indeks
lipase dihasilkan oleh 29 (61,7 %) isolat Aspergillus yang
besarnya sekitar 1,13–3,11, sedangkan 4 (8,5 %) isolat Aspergillus hanya terdeteksi positif menghasilkan aktivitas lipase. Indeks protease dihasilkan oleh 17 (36,2
%) isolat Aspergillus yang besarnya sekitar 1,05–2,0,
sedangkan 11 (23,4 %) hanya terdeteksi positif menghasilkan aktivitas protease. Indeks lipase (3,11)
tertinggi dihasilkan oleh Aspergillus (1523), sedangkan
indeks protease (2,0) tertinggi dihasilkan masing-masing
oleh Aspergillus (798) dan Aspergillus (917). Aspergillus
(1523) yang menghasilkan indeks lipase tertinggi
diisolasi dari minyak tanah, sedangkan Aspergillus (798)
dan Aspergillus (917) yang menghasilkan indeks protease tertinggi masing-masing diisolasi dari tanah KRI.
Tabel 5. Aktivitas enzim Aspergillus yang diisolasi dari berbagai bahan No Aspergillus No koleksi Habitat Aktivitas kitinase Indeks lipase Indeks protease
1. Aspergillus 722 Tanah KRI - 1,5 1,29
2. Aspergillus 790 Tanah KRI - 1,2 -
3. Aspergillus 798 Tanah KRI - - 2,0
4. Aspergillus 822 Tanah KRI - 1,19 1,13
5. Aspergillus 867 Tanah KRI - 1,13 1,25
6. A. fumigatus 907 Avtur - 1,75 1,13
7. A. niger 909 Minyak tanah - 1,2 -
8. A. niger 911 Minyak tanah - - -
9. A. niger 912 Minyak tanah - - -
10. A. niger 913 Tanah Hutan - - -
11. A. niger 917 Tanah KRI - 2,0 2,0
No Aspergillus No koleksi Habitat Aktivitas kitinase Indeks lipase Indeks protease
13. Aspergillus 919 Tanah KRI - 2,67 1,05
14. Aspergillus 920 Tanah KRI - 2,0 1,82
15. A. terreus 921 Minyak tanah - - 1,24
16. Aspergillus 976 Tanah Hutan - 1,67 1,74
17. A. flavus 979 Tanah Hutan - 1,2 -
18. Aspergillus 992 Tanah Hutan - 2,44 1,34
19. Aspergillus 993 Tanah Hutan - - 1,11
20. Aspergillus 1507 Minyak tanah - 1,33 -
21. Aspergillus 1523 Minyak tanah - 3,11 -
22. Aspergillus 1528 Minyak tanah - 2,86 +
23. Aspergillus 1573 Kain - + +
24. Aspergillus 1591 Tanah Hutan - 1,6 -
25. Aspergillus 1601 Tanah Hutan - 1,23 +
26. Aspergillus 1605 Tanah Hutan - - +
27. Aspergillus 1606 Tanah Hutan - 1,71 -
28. Aspergillus 1620 Tanah Hutan - 1,18 -
29. Aspergillus 1621 Tanah Hutan - - -
30. Aspergillus 1633 Kain - 1,4 +
31. Aspergillus 1642 Tanah Hutan - 1,36 1,11
32. Aspergillus 1661 Kain - 1,14 +
33. A. clavatus 1881 Kontaminan - + 1,2
34. A.chevalieri 1886 Kompos - - -
35. Aspergillus 1888 Kompos - 1,45 +
36. Aspergillus 1895 Kompos - 1,33 +
37. Aspergillus 1913 Tanah Hutan - 2,17 +
38. A. neoglaber 1917 Tanah Hutan - 3,0 -
39. A. kanagawaensis 1923 Tanah Hutan - 1,4 +
40. A. niger 2149 Kontaminan - - - 41. Aspergillus 2154 Kontaminan - + - 42. A. fumigatus 2157 Kontaminan - 1,2 + 43. A. niger 2161 Kontaminan - - - 44. A. niger 2168 Kontaminan - + - 45. A. fumigatus 2172 Kontaminan - - 1,24 46. A. niger 2224 Kontaminan - - - 47. A. fumigatus 2258 Kontaminan - - 1,14
Keterangan: - = tidak terdeteksi; + = terdeteksi
Semua (27) isolat Aspergillus yang diisolasi dari
berbagai organ tanaman diketahui tidak terdeteksi menghasilkan enzim kitinase, 16 (59,3 %) isolat Aspergillus menghasilkan enzim lipase, 12 (44,4 %)
isolat Aspergillus menghasilkan enzim protease, dan 8
(29,6 %) isolat Aspergillus tidak menghasilkan enzim
lipase maupun protease (Tabel 6). Indeks lipase
dihasilkan oleh 14 (51,9 %) isolat Aspergillus yang
besarnya sekitar 1,14–2,0, sedangkan 2 (7,4 %) isolat Aspergillus hanya terdeteksi positif menghasilkan aktivitas lipase. Indeks protease dihasilkan oleh 8 (29,6
%) isolat Aspergillus yang besarnya sekitar 1,09–1,29,
sedangkan 4 (14,8 %) isolat Aspergillus hanya terdeteksi
positif menghasilkan aktivitas protease. Indeks lipase
(2,0) tertinggi dihasilkan masing-masing oleh A.
kanagawaensis (2278) dan A. terreus (2288), sedangkan indeks protease (1,29) tertinggi dihasilkan oleh Aspergillus (2132). Aspergillus kanagawaensis (2278)
dan A. terreus (2288) masing-masing diisolasi dari biji
kedelai, sedangkan Aspergillus (2132) diisolasi dari akar
Tabel 6. Aktivitas enzim Aspergillus yang diisolasi dari organ tanaman No Aspergillus No koleksi Habitat Aktivitas kitinase Indeks lipase Indeks protease 1. Aspergillus 1542 Daun - 1,44 - 2. A. terreus 1755 Daun - 1,4 - 3. Aspergillus 1878 Buah - 1,27 1,13 4. Aspergillus 1879 Daun - 1,25 1,12
5. Aspergillus 1961 Batang kopi - 1,14 +
6. Aspergillus 1967 Batang kopi - 1,44 -
7. Aspergillus 1982 Umbi bawang merah - - -
8. A. niger 1994 Bunga tomat - - -
9. A. niger 2026 Daun tomat - - -
10. A. niger 2080 Buah jeruk - - 1,11
11. A. fumigatus 2125 Akar pisang - 1,6 -
12. A. fumigatus 2126 Akar pisang - 1,29 1,11
13. Aspergillus 2132 Akar pisang - + 1,29
14. A. niger 2139 Akar pisang - - -
15. A. niger 2141 Akar pisang - - -
16. A. chevalieri 2142 Akar pisang - - 1,13
17. Aspergillus 2143 Akar pisang - - 1,22
18. A. chevalieri 2144 Akar pisang - + 1,09
19. A. niger 2255 Daun pisang - - -
20. A. niger 2256 Daun pisang - - -
21. Aspergillus 2277 Biji kedelai - 1,35 -
22. A. kanagawaensis 2278 Biji kedelai - 2,0 +
23. A. niger 2279 Biji kedelai - 1,33 -
24. Aspergillus 2282 Biji kedelai - - -
25. Aspergillus 2283 Biji kedelai - 1,25 +
26. A. terreus 2288 Biji kedelai - 2,0 -
27. Aspergillus 2291 Biji kedelai - 1,33 +
Keterangan: - = tidak terdeteksi; + = terdeteksi
Secara total (116), 53 (45,7 %) isolat Aspergillus
terdeteksimenghasilkan lipase dan protease, 24 (20,7 %)
isolat Aspergillus menghasilkan lipase, 13 (11,2 %) isolat
Aspergillus menghasilkan protease, dan 26 (22,4 %)
isolat Aspergillus tidak menghasilkan enzim lipase
maupun protease (Tabel 4, 5 & 6).
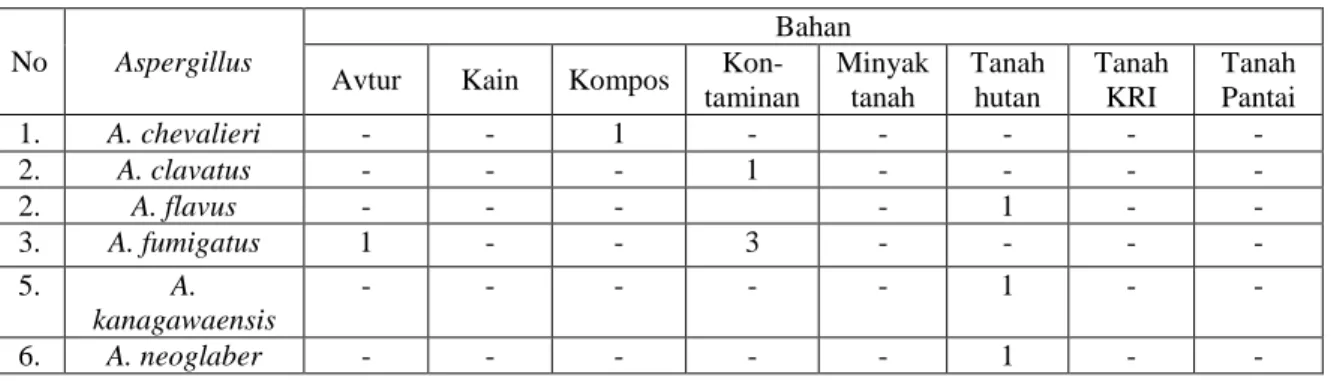
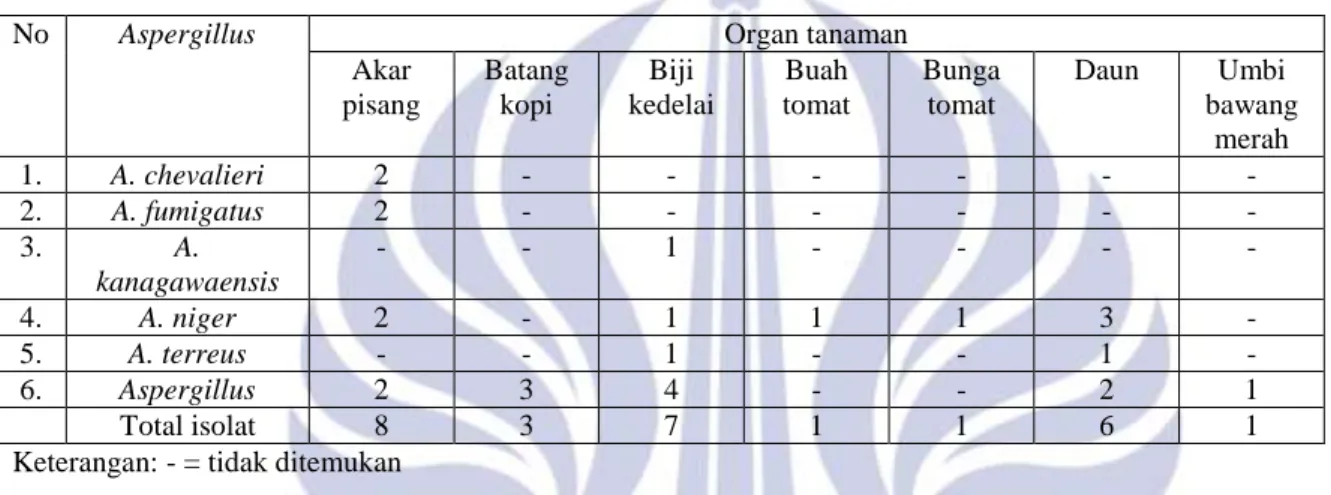
Gambar 1. Aktivitas protease Aspergillus (867) kiri dan
aktivitas lipase Aspergillus (790) kanan
Tiga contoh isolat Aspergillus yang menghasilkan
enzim lipase dan protease tinggi dapat menghambat pertumbuhan patogen Foc (Tabel 7 & Gambar 2). Persen hambatan pertumbuhan Foc tertinggi pada pengamatan
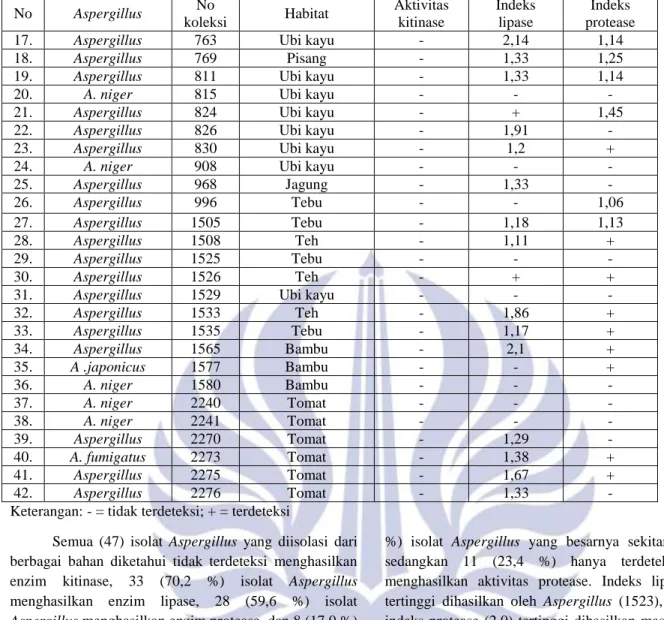
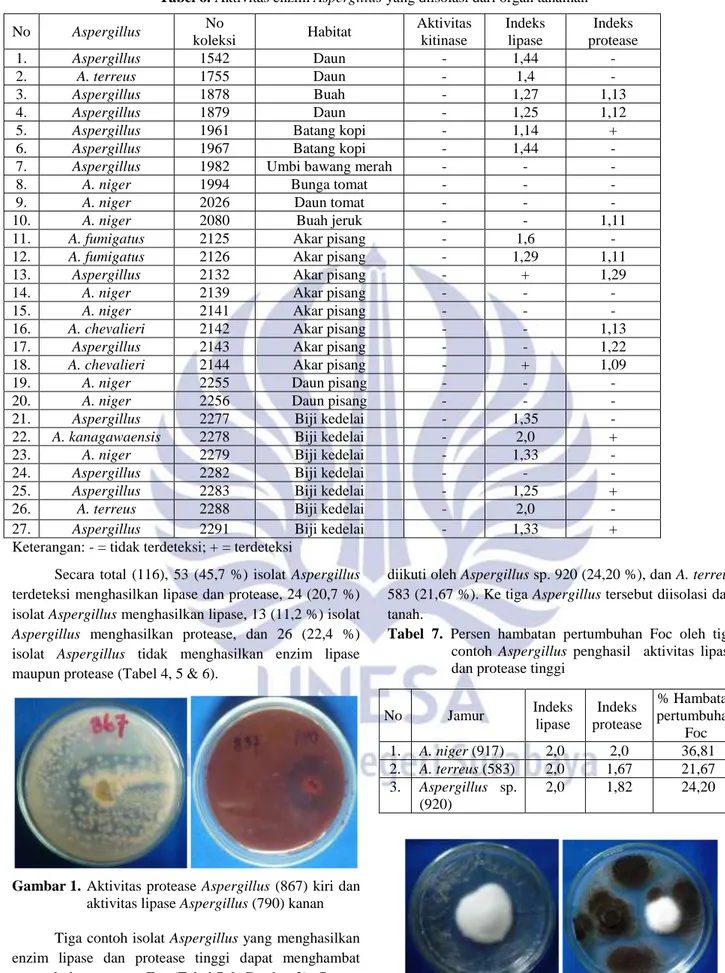
hari ke-5 diperlihatkan oleh A. niger 917 (36,81 %)
diikuti oleh Aspergillus sp. 920 (24,20 %), dan A. terreus
583 (21,67 %). Ke tiga Aspergillus tersebut diisolasi dari
tanah.
Tabel 7. Persen hambatan pertumbuhan Foc oleh tiga
contoh Aspergillus penghasil aktivitas lipase
dan protease tinggi
No Jamur Indeks lipase Indeks protease % Hambatan pertumbuhan Foc 1. A. niger (917) 2,0 2,0 36,81 2. A. terreus (583) 2,0 1,67 21,67 3. Aspergillus sp. (920) 2,0 1,82 24,20 Gambar 2. Foc (kiri) dan A. niger (917) vs Foc (kanan)
PEMBAHASAN
Banyaknya (116) isolat Aspergillus yang terisolasi
dari berbagai subtrat pada penelitian ini merupakan sumber bioaktif alami berpotensi yang dapat dieksploitasi untuk keperluan industri, obat, dan pertanian (Strobel &
Daisy, 2003). Masih banyaknya isolat Aspergillus (54,3
%) yang belum teridentifikasi sampai spesies secara morfologi merupakan kendala tersendiri sehingga ke depannya perlu diusahakan identifikasi dengan metode molekuler.
Sebanyak 45,7 % Aspergillus menghasilkan
aktivitas lipase maupun protease. Hasil ini tentunya sangat menggembirakan karena ada kemungkinan ke dua enzim tersebut dapat melisis polimer kecil (protein dan lipid) yang menyusun dinding dan membran sel jamur yang mungkin terlibat dalam respon antagonisme dari Aspergillus (de la Cruz & Llobell, 1999). Hal ini
terbukti bahwa tiga contoh isolat Aspergillus yang
menghasilkan aktivitas lipase dan protease tinggi (A.
niger 917, A. terreus 583 dan Aspergillus sp. 920) ternyata dapat menghambat pertumbuhan jamur patogen
Foc secara in-vitro sekitar 21,67-36,81 %. Antagonisme
Aspergillus melawan patogen Foc telah dilaporkan secara
intensif oleh Garoé et al. (2013) dan Suciatmih &
Rahmansyah (2013).
Beberapa penelitian berikut yang mendukung
penelitian Aspergillus yang menghasilkan enzim
melaporkan bahwa A. flavus, A. fumigatus, dan A. niger
menghasilkan aktivitas lipase dan protease (Aboul-Nasr et al., 2013). Protease dihasilkan oleh A. clavatus
(Sampaio e Silva et al., 2011) dan A. terreus (Budak et
al., 2014), sedangkan lipase dihasilkan oleh A. japonicus
(Jayaprakash & Ebenezer, 2012) dan A. terreus
(Mahmoud et al., 2015).
Ketidakmampuan Aspergillus menghasilkan
aktivitas kitinase, lipase, dan protease; serta Aspergillus
yang hanya terdeteksi positif menghasilakn enzim lipase atau protease tetapi indeks aktivitasnya tidak dapat diukur dalam penelitian ini mungkin disebabkan oleh faktor biologi seperti sifat genetik dari organisme yang dapat mempengaruhi sifat metabolisme dan biokimia strain mikroba sehingga akhirnya mempengaruhi sistem produksi enzimnya. Hasil yang berbeda telah dilaporkan oleh beberapa penelitian berikut bahwa aktivitas kitinase
dihasilkan oleh A. flavus (Gunalan et al., 2012), A.
fumigatus (Xia et al., 2001), A. niger (Brzezinska &
Jankiewicz, 2012), dan A. terreus (Farag & Al-Nusarie,
2014).
Metode kultur ganda telah digunakan secara
ekstensif untuk mempelajari antagonisme (Bhale et al.,
2013). El-Katatny (2001) menginformasikan bahwa pembentukkan zona hambatan pada titik kontak antara Aspergillus dan patogen Foc dapat disebabkan oleh
produksi metabolit volatil dan bukan volatil atau enzim hidrolitik, seperti lipase dan protease yang dihasilkan Aspergillus.
SIMPULAN
Aspergillus dapat diisolasi dari tanah rizosfer, organ tanaman, dan bahan lain (avtur, kain, kompos, minyak tanah, dan kontaminan yang berasal dari udara).
Tiga isolat Aspergillus (A. niger 917, A. terreus 583, dan
Aspergillus sp. 920) yang menghasilkan aktivitas lipase dan protease tinggi dapat menghambat patogen Foc. Ke tiga isolat tersebut akan diuji lebih lanjut pada skala
rumah kaca. Aspergillus yang belum berhasil
diidentifikasi secara morfologi akan diidentifikasi secara molekuler pada penelitian mendatang.
DAFTAR PUSTAKA
Aboul-Nasr MB, Zohri ANA, Amer EM, 2013. Enzymatic and toxigenic ability of opportunistic fungi contaminating intensive care units and operation rooms at Assiut University Hospitals,
Egypt. Springerplus, 2: 347. Doi:
10.1186/2193-1801- 2-347.
Alwathnani HA, Perveen K, 2012. Biological control of fusarium wilt of tomao by antagonist fungi and
Cyanobacteria. African Journal of Biotechnology,
11: 1100-1105.
Beckman C, Roberts E, 1995. On the nature and genetic basis for resistance and tolerance to fungal wilt
diseases of plants. Advances in Botanical Research,
21: 35-77.
Bhale UN, Wagh PM, Rajkonda JN, 2013. Antagonistic
confrontation of Trichoderma spp. against fruit rot
pathogens on sapodilla (Manilkara zapota L.).
Journal of Yeast and Fungal Research, 4(1): 5-11. Bhattacharyya PN, Jha DK, 2011. Optimization of
cultural conditions affecting growth and improved bioactive metabolite production by a subsurface Aspergillus strain TSF 146. International Journal of Applied Biology and Pharmaceutical Technology,2: 133-143.
Brzezinska MS, Jankiewicz U, 2012. Production of
antifungal chitinase by Aspergillus niger LOCK 62
and its potential role in the biological control. Current Microbiology, 65(6): 666-672.
Budak SO, Zhou M, Brouwer C, Wiebenga Ad, Benoit I, Falco MD, Tsang A, de Vries RP, 2014. A genomic
survey of proteases in Aspergilli. BMC Genomics,
15:523 DOI:10.1186/1471-2164-15-523.
Campbell R, 1989. Biological control of microbial plant pathogens. Cambridge: Cambridge University Press.
de la Cruz J, Llobell A, 1999. Purification and properties of a basic endo-β-1,6-glucanase (BGN16.1) from
the antagonistic fungus Trichoderma harzianum.
European Journal of Biochemistry,265: 145-151. Djatnika I, Hermanto C, Eliza, 2003. Pengendalian hayati
layu fusarium pada tanaman pisang dengan Pseudomonas flourescens dan Gliocladium. Jurnal Hortikultura, 13: 203-211.
Dolar FS, 2001. Antagonistic effect of Aspergillus
melleus Yukawa on soilborne pathogens of
chickpea. Tarim Bilimleri Dergisi, 8(2): 167-170.
El-Katatny MH, Gudelj M, Robra KH, Elnaghy MA, Gubitz GM, 2001. Characterization of a chitinase
and an endo-b-1,3-glucanase from Trichoderma
harzianum Rifai T24 involved in control of the
phytopathogen Sclerotium rolfsii. Applied
Microbiology and Biotechnology, 56: 137-143.
Farag MA, Al-Nusarie ST, 2014. Production,
optimization, characterization and antifungal
activity of chitinase produced by Aspergillus terrus.
African Journal of Biotechnology, 13(14): 1567-1578.
Garoé N-T, Cabrera R, Cosoveanu A, Martín TT, Giménez C, 2013. Survey of banana endophytic fungi isolated in artificial culture media from an
applied viewpoint. Journal of Horticulture, Forestry
and Biotechnology, 17(2): 22-25.
Gunalan G, Sadhana D, Ramya RP, 2012. Production,
optimization of chitinase using Aspergillus flavus
and its biocontrol of phytopathogenic fungi. Journal
of Pharmacy Research, 5(6): 3151.
Jayaprakash A, Ebenezer P, 2012. Optimization of Aspergillus japonicus lipase production by response
surface methodology. Journal of Academia and
Industrial Research, 1(1): 23-30.
Jumjunidang, Narsis N, Riska, Handayani H, 2005.
Teknik pengujian in vitro ketahanan pisang terhadap
layu fusarium menggunakan filtrat toksin dari kultur Fusarium oxysporum f.sp. cubens. Jurnal Hortikultura, 15(2): 135-139.
Lingappa Y, Lockwood JL, 1962. Chitin medium for
selective isolation and culture of Actinomycetes. Phytopathology, 52: 317-323.
Mahmoud GAL, Koutb MM, Morsy FM, Bagy MM, 2015. Characterization of lipase enzyme produced
by hydrocarbons utilizing fungus Aspergillus
terreus. European Journal of Biological Research, 5(3): 70-77.
Najar AG, Anwar A, Masoodi L, Khar MS, 2011. Evaluation of native biocontrol agents against Fusarium solani f.sp. melongenae causing wilt
disease of brinjal in Kashmir. Journal of Phytology,
3(6): 31-34.
Peterson MH, Johnson MJ, 1949. Delayed hydrolysis of
butterfat by certain Lactobacilli and Micrococci
isolated from cheese. Journal of Bacteriology, 58:
701-708.
Raper KB, Fennell DI, 1965. The Genus Aspergillus.
Baltimore: Williams and Wilkins.
Sampaio e Silva TA, Knob A, Tremacoldi CR,
Brochetto-Braga MR, Carmona EC, 2011.
Purification and some properties of an extracellular
acid protease from Aspergillus clavatus. World
Journal of Microbiology and Biotechnology, 27: 2491-2497.
Sharma S, 2012. Antagonistic interaction between Fusarium oxysporum var cumini and Aspergillus niger. Journal of Research in Biology, 1: 032-034. Skidmore AM, Dickinson CH, 1976. Colony interactions
and hyphal interference between Septoria nodurum
and phylloplane fungi. Transactions of the British
Mycological Society, 66: 57-64.
Smitha C, Finosh GT, Rajesh R, Abraham PK, 2014. Induction of hydrolytic enzymes of phytopathogenic
fungi in response to Trichoderma viride influence
biocontrol activity.International Journal of Current
Microbiology and Applied Sciences, 3(9): 1207-1217.
Snyder W, Hanson H, 1940. The species concept in Fusarium. American Journal of Botany, 27: 64–67. Strobel G, Daisy B, 2003. Bioprospecting for microbial
endophytes and their natural products. Microbiology
and Molecular Biology Reviews, 67(4): 491-502. Suciatmih, Rahmansyah M, 2013. Endophytic fungi
isolated from mangrove plant and have antagonism
role against Fusarium wilt. ARPN Journal of
Agricultural and Biological Science, 8(3): 251-257. Vuylsteke D, Swennen R, de Langhe E, 1996. Field performance of somaclonal variants of plantain. Journal of the American Society of Horticultural Science, 121: 42-46.
Xia G, Jin C, Zhou J, Yang S, Zhang S, Ji C, 2001. A Novel Chitinase Having a unique mode of action
from Aspergillus fumigatus YJ-407. European