PENINGKATAN EFISIENSI TRANSFORMASI
GEN XYLOGLUCANASE'
PADA BERBAGAI EKSPLAN
Acacia mangium Willd
NELLY ANNA
SEKOLAH PASCASARJANA
INSTITUT PERTANLAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Peningkatan Efisiensi Transformasi Gen Xyloglucanase pada berbagai Eksplan Acacia mangium Willd adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip
dari
karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks clan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Bogor, Agustus 2007
RINGKASAN
NELLY ANSA. Peningkatan Efisiensi Transformasi Gel1 Xyloglucanase pada berbagai Eksplan Acacia rnangium Willd. Dibimbing oleh ULFAH JUNIARTI SZREGAR dan EWJY SUDARMONOWATI
ABSTRACT
NELLY ANNA. Increasing Efficiency of Xyloglucanase Gene Transformation
in Acacia mangium Willd Explants. Under the direction of ULFAH JUNIARTI
SIREGAR and ENNY SUDARMONO WATI
An effort to obtain transgenic Acacia mangium Willd, a fast growing species with high wood quality was done through transformation techniques. Transformation was performed by introducting xyloglucanase gene using
Agrobacterium tumefaciens, strain LBA4404 harboring pAaXEG300 which
contains XEG gene, nptII gene, and 35 S promoter. Different types of explants i.e. bud, stem, callus, multiple adventious shoots, and leaf pieces of Acacia mangium
Willd were used as target tissues. The selection for transgenic was performed through MS medium supplemented with 1 mg/l thidiadzuron, 0,25 mg/l indole-3- acetac acid, 225 mg/l carbenicillin, and concentrations of canamysin 400 mgll. Although the transformation experiment have not yet produced transgenic plants, adventious shoots could be obtained at 8,3% from transformed stem, and 5,5% from bud as explants. Confirmation of transgenic plants expressing xyloglucanase gene using Western blot is still on going.
O
Hak cipta milik IPB, tahun
2007
Hak
cipta dilindungi Undang-undang
I. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penulisan karya ilmiah, penyusunan Iaporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan clan memperbanyak sebagian atau
PENINGKATAN EFISIENSI TRANSFORMASI GEN
XYLOGLUCANASE PADA BERBAGAI EKSPLAN
Acacia mangium
Willd
NELLY ANNA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis
Nama
NRP
: Peningkatan Efisiensi Transformasi Gen Xyloglucanase pada berbagai Eksplan Acacia mangium Willd
: Nelly Mna
: E05 105025 1
Disetujui, Komisi Pembimbing
Dr. Ir. Ulfah Juniarti Siregar. M.Aw
Ketua
Dr. Ir. Ennv s&annonowati. APU Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Pengetahuan Kehutanan
.
C uDr. Ir. Rinekso Soekmadi, M.
PRAKATA
Puji clan syukur penulis panjatkan kepada Allah SWT atas segala karunia- Nya sehingga karya ilrniah ini berhasil diselesaikan. Tema yang dipilih dalarn penelitian yang dilaksanakan sejak bulan Agustus 2006 ini adalah transformasi gen, dengan judul Peningkatan Efisiensi Transformasi Gen Xyloglucanase pada berbagai Eksplan Acacia mangium Willd.
Terirna kasih penulis ucapkan kepada Ibu Dr. Ir. Ulfah Juniarti Siregar, M.Agr dan Ibu Dr. Ir. Enny Sudarmonowati, APU selaku pembimbing. Di sarnping itu, penghargaan penulis sampaikan kepada Research Institute for Sustainable Huymanosphere, Kyoto University, Jepang atas kerja samanya dengan Lembaga Ilmu Pengetahuan Indonesia sehingga penelitian ini dapat diselesaikan. Terima kasih juga penulis ucapkan kepada Ibu Sri Hartati dan semua s M Laboratorium Biologi Molekuler Bioteknologi-LIPI, yatlg telah membantu selama penelitian ini berjalan serta teman-teman yang telah membantu
dan
memberi motivasi. Ungkapan terimakasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya.Semoga karya ilrniah ini bermanfaat.
Bogor, Agustus 2007
RIWAYAT HIDUP
Penulis dilahirkan di Binjai, Surnatera Utara pada tanggal 10 Juni 198 1 dari ayah Agustarni Piliang dan ibu Hj. Nurani Koto. Penulis merupakan putri ke sepuluh dari sepuluh orang bersaudara. Pendidikan sarjana ditempuh mulai September 2000 di Program Studi Budidaya Hutan, Departernen Kehutanan, Fakultas Pertanian Universitas Sumatera Utara, dan lulus pada tanggal 12 Maret 2005, Selama menempuh pendidikan sarjana, penulis berkesempatan mendapatkan beasiswa dari PERTAMINA pada tahun 2002. Selain itu, penulis juga pernah menjadi asisten laboratorium Silvikultur pada tahun 2003 dan
Teknologi Benih pada tahun 2004.
DAFTAR IS1
Halaman
...
DAFTAR TABEL
xi
.
.
...
DAFTAR GAMBAR xi1
...
PENDAHULUAN 1
...
Latar Belakang 1
Perurnusan Masalah
...
3Tujuan Penelitian
...
3TINJAUAN PUSTAKA
...
4Botani Acasia mangium Willd
...
4Pemuliaan Tanaman secara Konvensional dan Peranan Bioteknologi
...
5...
Pertumbuhan dan Perkembangan Kayu 6 Gen Xyloglucanase...
7Transformasi Genetik Tanaman
...
8...
Seleksi Tanaman Transgenik 10...
Regenerasi In vitro 10 Kultur Jaringan...
1 1...
Eksplan 1 1...
Media. . .
12...
Kondisi Fisik Kultur 13...
Kondisi Lingkungan Kultur 13...
Induksi Embrio Somatik 14...
Analisis Tanaman Transgenik 15.
.
...
Uji GUS 15...
Analisis Polymerase Chain Reaction (PCR) 16...
Southern Blot 16 Northern Blot...
17Western Blot
. .
...
17...
Uji Hayati 18 BAHAN DAN METODE ... 19Tempat dan Waktu
...
19...
Bahan dan Alat 19...
Metode Penelitian 19 Persiapan eksplan...
19...
Optimasi media seleksi 2 1 Kultur Agrobacterium tumefaciens...
21Transforrnasi dan regenerasi
...
21Uji ekspresi Western blot
...
22X Induksi embriosomatik Acacia mangium Willd
...
25Seleksi terhadap resistensi Kanamisin pada
eksplan yang belum ditransformasi
...
28...
Penentuan nilai ODbo0 Apbacferium tumefaciens 30...
Transformasi berbagai eksplan Acacia mangium Willd 31 Regenerasi tanaman Acacia mangium Willdyang telah ditransformasi
...
34 Seleksi eksplan Acacia mangium Willd yang telahditrmfonnasi pada beberapa konsentrasi Kanamisin
...
37 Uji Ekspresi Western Blot...
42KESIMPULAN DAN SARAN
...
43...
Kesimpulan 43
Saran
...
43DAFTAR TABEL
Halaman
1 Persentase biji Acacia mangium Willd
yang berkecambah pada media MS tanpa zat pengatur tumbuh
...
.242
Persentase embrio dari biji Acacia mangium Willd yang berkalus Dan tumbuh tunas majemuk pada media MS+
0,25 mg/l IAA+
1 mg/l TDZ...
263 Uji konsentrasi Kanamisin pada berbagai eksplan
Acacia mangium Willd yang tidak ditransfomasi
...
294 Persentase eksplan Acacia mangium Willd yang hidup setelah direndam pada Agrobacterium tumefaciens
selama 5 menit pada beberapa nilai OD600
...
3 15 Persentase hidup eksplan Acacia mangium Willd yang telah
ditransformasi pada minggu ke 8 pada media seleksi (0,25 mgll IAA
+
1 mg/l TDZ+
225 Carbenicillin+
400 mg/l Kanamisin)...
326 Persentase regenerasi eksplan pucuk dan batang
yang telah ditransformasi setelah 8 minggu pada media seleksi
dan 5 minggu pada media regenerasi
...
3 57 Persentase hidup kontrol clan eksplan yang telah ditransformasi pada media seleksi yang mengandung 100 mg/l
DAFTAR GAMBAR
...
1 Diagram integrasi bioteknologi 6
...
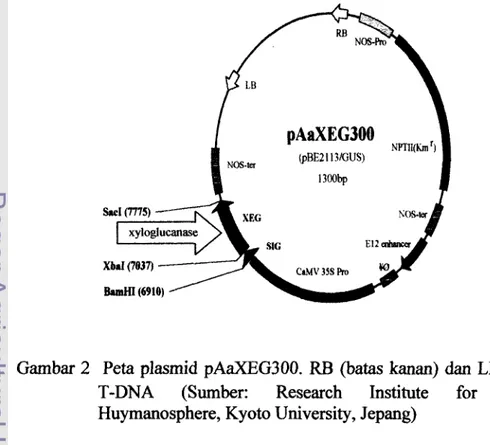
2 Peta plasmid pAaXEG300 RB (batas kanan) dan LB (batas kiri) 1 9
3 Eksplan Acacia mangium Willd berumur 3 bulan pada media MS (material untuk transformasi)
...
254 Kalus yang berasal dari embrio Acacia mangium Willd pada media
...
(MS+
0,25 mg/l IAA+
1 mg/l TDZ) 275 Eksplan Acacia mangium Willd yang telah ditransformasi pada media seleksi (0,25 mgA IAA
+
1 mg/l TDZ+
225 mg/l Carbenicillin
+
400 mg/l Kanamisin) pada minggu ke 8...
346 Eksplan yang telah ditransformasi pada media regenerasi
...
367 Eksplan Acacia mangium Willd setelah 8 minggu pada media seleksi (0,25 mg/l IAA
+
1 mg/l TDZ+
225 mg/l Carbenicillin
+
400 mg/l Kanamisin)... 39
8 Regenerasi eksplan Acacia mangium Willd yang telah
PENDAHULUAN
Latar Belakang
Kerusakan hutan dam dewasa ini semakin meningkat, sedangkan kebutuhan kayu semakin meningkat sejalan dengan meningkatnya pertumbuhan penduduk. Dalam rangka memenuhi kebutuhan kayu, berbagai usaha telah dilakukan diantaranya adalah dengan membangun hutan tanaman.
Laju penambahan hutan tanaman baru setiap tahun diperkirakan 4,5 juta ha. Dari pertumbuhan hutan tanaman tersebut, Asia terutama China, Indonesia, Malaysia, dan Vietnam memberikan kontribusi 70% terhadap pertumbuhan itu. Melalui reboisasi lahan alang-alang dan hutan sekunder, lebih
dari
9,9 juta ha hutan tanaman telah dapat dibangun, terrnasuk f juta ha hutan tanaman jati dan 1,4 juta ha hutan tanaman Acacia mangium Willd di Sumatra dan Kalimantan. Dengan dernikian, pada masa mendatang diharapkan Indonesia akan menjadi negara terkemuka di dunia dalam produksi pulpdari
hutan tanaman Acacia mangium Willd (Kedu dun Diy, 2005). Bagi Indonesia yang kehilangan banyak kayu berkualitas akibat illegal logging, kayu jenis Acacia mangium Willd dapat menjadi alternatif devisa negara dan bisa bersaing di pasaran dunia.Beberapa tahun terakhir, program bioteknologi telah terbukti memberikan sejumlah manfaat u n W mengatasi berbagai keterbatasan &lam metoda pemuliaan secara konvensional. Salah satunya adalah upaya transformasi genetik tanaman dengan pemanfaatm sejurnlah gen yang bermanfaat dari berbagai species untuk diekspresikan pada tanaman target.
Penerapan teknik transformasi genetik terbukti sangat membantu ddam perakitan spesies tahan (herbisida, virus, dan penyakit) atau spesies unggul, terutama jika ti& memunglunkan dilakukan dengan teknik konvensiond. Selain itu juga bermanfaat untuk meningkatkan kualitas dan kuantitas tanaman (Gunawan, 1992).
Tanaman cepat tumbuh, contohnya adalah Acacia mangium Willd yang dikembangkan dengan penerapan teknik transfonnasi genetik dapat digunakan sebagai solusi untuk meningkatkan produksi hutan. Kemajuan semacam itu dapat menguntungkan dunia industri kayu, karena dalarn waktu yang relatif singkat dapat menebang kayu untuk keperluan usahanya. Tanaman kayu yang biasanya memerlukan wakh 10 hingga 12 t&un untuk bisa ditebang, dengan transformasi genetik pohon tersebut sudah layak tebang dalam waktu 3 hingga 5 tahun dari waktu tanam.
Selain waktu tumbuh yang singkat, batang kayu juga bisa dibuat lurus sehingga dapat menghemat areal hutan tanaman yang pada akhinya hasil produksi kayu dapat ditingkatkan. Selain itu, kualitas kayu dapat ditingkatkan karena proses tramformasi genetik dalam pohon tersebut dapat meningkatkan berat jenis kayu sehingga kayu yang dihasilkan semakin bagus. Peningkatan berat jenis pohon juga berarti peningkatan kandungan gula atau karbohidrat (C&I1206)
dalam pohon. Kandungan zat ini
akan
membuat semakin banyak gas karbondioksida (C02) yang diserap oleh pohon. Sehingga adanya tanaman h a i l transformasi genetik ini juga dapat digunakan mtuk mengurangi pencemaran udara yang diakibatkan oleh gas C 0 2 .Berdasarkan atas penelitian Hartati et.al. (2005) maka perlu dilakukan percobaan selanjutnya untuk meningkatkan efisiensi transformasi pada berbagai eksplan
Acacia mangium Willd.
Sistem transformasi genetik yang paling umum digunakan adalah dengan menggunakan Agrobacterium tumefaciens. Sistem ini telah banyak digunakan karena efisien, sederhana
dan
stabil dalam mengintroduksikan suatu gen.Perurnusan Masalah
Transforrnasi gen xyloglucanase menggunakan Agrobacterium
tumefaciens dapat merupakan salah satu solusi untuk memperoleh bahan tanaman
Acacia mangium Willd yang unggul, tetapi ha1 yang harus diperhatikan adalah bagian eksplan apa yang dapat memberikan hasil transformasi yang terbaik. Dengan demikian pertanyaan yang muncul pada penelitian ini adalah: apakah dengan membandingkan berbagai eksplan Acacia mangium Willd dapat menghasilkan metoda transformasi gen xyloglucanase yang lebih efisien?
Tujuan Penelitian
Tujuan penelitian ini adalah mengembangkan metoda yang efektif untuk mengintroduksikan gen xyloglucanase ke dalam tanaman dan regenerasi tanman
TINJAUAN PUSTAKA
Botani Acacia mangium WilldAcacia mangium Willd jenis legurn yang termasuk dalam famili
Leguminosae, sub-famili Mimosoideae. Acacia mangium Willd secara umum dikenal sebagai brown salwood, black wattle, hickory wattle (di Australia).
Manggae hutan, tongke hutan, nak, laj, jerri (di Indonesia). Di Papua New Guinea dikenal sebagai arr, di Malaysia dikenal sebagai nama mangium, kayu sofada. Di Thailand dikenal sebagai kra thin tepa. Mangium mempunyai beberapa nama lain, diantaranya adalah : Mangium montanum Rump dan Acasia
glaucescena, serta Rancosperma mangium (Willd) (Awang dan Taylor, 1993).
Buahnya berupa polong kering yang merekah dan melingkar ketika masak, agak keras dengan panjang 7.8 cm dan lebar 3-5 rnrn. Biji berwarna hitam mengkilat, lonjong, funicle berwarna oranye. Daun besar, panjangnya mencapai 25 cm, lebar 3-10 cm, berwarna hijau gelap dengan empat urat longitudinal (tiga pada Acasia auriculiformis), daun majemuk. Secara umum Acacia mangium
Willd mencapai tinggi 25-35 m dengan bebas cabang melebihi setengah dari total tinggi. Diameternya dapat mencapai lebih dari 60 cm. Pada lahan yang miskin, pohon biasanya lebih kecil, dengan rata-rata tinggi antara 7 dan 10 m. Pohon yang masih muda berwarna hijau, kulit kasar dan beralur, berwma abu-abu atau coklat (Awang dan Taylor, 1993).
Pada tempat tumbuh yang baik, pohon berumur 9 tahun tingginya mencapai 23 m, dengan mta-rata riap diameter 2-3 c d t h dan produksi kayunya 41.5 m3/ha. Acacia mangium Willd tidak memerlukan persyaratan tumbuh yang tinggi. Jenis
ini dapat tumbuh pada tanah miskin hara, padang alang-alang, bekas tebangan, tanah tererosi, tanah berbatu dan juga tanah aluvial. Acacia mangium Willd dapat beradaptasi dengan tanah asam (pH 4.5-6.5) di dataran tropis yang lembab (Awang dan Taylor, 1993).
Acacia mangium Willd termasuk jenis yang tumbuh cepat, tidak
rata-rata 20 m3/ha/tahun. Acacia mangium Willd termasuk dalam kelas kuat III- IV, berat 0,56-0,60 dengan nilai kalori rata-rata antara 4800-4900 k.cal/kg (Dephut, 1994). Kegunaan kayu Acacia mangium Willd sebagai bahan konstruksi ringan sampai berat, rangka pintu dan jendela, perabot rumah tangga (a.1. lemari), lantai, papan dinding, tiang, tiang pancang, gerobak dan rodanya, pemeras minyak, gagang alat, alat pertanian, kotak dan batang korek api, papan partikel, papan serat, vinir dan kayu lapis, pulp dan kertas. Selain itu baik juga untuk kayu bakar dan arang (Mandang dun Pandit, 1997).
Pemuliaan Tanaman secara Konvensional dan Peranan Bioteknologi Pemuliaan tanaman merupakan kegiatan untuk mengubah susunan genetik tanaman secara tetap, sehingga memiliki sifat atau penampilan sesuai dengan tujuan yang diinginkan pelakunya. Pemuliaan tanaman umumnya mencakup tindakan penangkaran, persilangan, dan seleksi. Produk pemuliaan tanaman adalah kultivar dengan ciri-ciri yang khusus dan bermanfaat bagi penanamnya. Dalam kerangka usaha pertanian (agribisnis), pemuliaan tanaman merupakan bagian awalkulu dari mata rantai usaha tani dan memastikan tersedianya benih atau bahan tanam yang baik dan bermutu tinggi (Anonim, 2007).
Uji genetik
genotype Materi
genetik
Hibridisasi I Pembiakan
vegetatif
Rekayasa genetika Gambar 1 Diagram integrasi bioteknologi
Pertumbuhan dan Perkembangan Kayu
Kayu adalah bahan organik dengan susunan unsur 50% C, 6% H, 44% 0 (berdasar bobot), dan sedikit saja unsur lain. Kayu dapat juga disebut polimer alami, mengingat 97-99% bobotnya berupa polimer (sekitar 90% pada kayu tropis). Dari jumlah itu, sebesar 65-75% adalah golongan polisakarida. Dari persfektif kimia, jaringan kayu (termasuk bahan sel dan zat antarsel) merupakan bahan komposit yang dibangun dari berbagai polimer organik, yakni molekul yang terbuat dari ribuan subunit atau monomer. Struktur dasar atau materi kerangka dari semua dinding sel kayu ialah selulosa, yaitu molekul gula linear
Pertumbuhan kayu dalam dimensi memanjang dan melebar adalah berkat aktivitas sel khusus yang disebut meristem. Meristem apikal terletak di bagian ujung batang atau cabang, juga di ujung akar, dan berperan dalam pertumbuhan memanjang (pertumbuhan primer) dari kayu. Setelah melewati tahun pertama, mulailah kegiatan meristem lateral, atau lazim disebut kambium vaskuler. Semua sel di dalam zone kambium adalah hidup. Pada waktu pembentukan xylem, mulailah serangkaian transformasi yang mengubahnya menjadi unsur kayu dewasa. Sebagian dari turunan xylem dapat mengalami perusakan diri dan modifikasi dinding sel, sehingga terbentuklah lumen. Selama fase pembelahan dan pembesaran sel, dinding sel merupakan kantong yang tipis, lentur, dan dapat melar, yang disebut dinding primer. Menjelang akhir proses pembesaran, mulailah pembentukan dinding sekunder ke arah lumen dari dinding primer. Serat kayu, pembuluh, dan unsur xylem dan phloem tertentu yang tidak berfungsi sebagai penyalur d d a t a u pendukung, biasanya membentuk dinding sekunder (Haygreen et al., 2002).
Gen xyloglucanase
Xyloglucanase merupakan sejenis enzim yang mengkatalisis reaksi hidrolisis xyloglucan (Irwin et al., 2003). Xyloglucan adalah penyusun utama hemiselulosa polisakarida pada dinding sel tanaman dikotil termasuk di dalamnya sel kayu, yang dapat membentuk ikatan hidrogen dengan mikrofibril selulosa dan secara potensial membentuk ikatan silang dengan mikrofibril (Campbell dun Braam, 1998; Valls et al., 2006). Xyloglucan terdapat pada 20 % berat kering dinding sel primer (York dan Eberhard, 2003).
untuk merperkuat hubungan antara dinding sel primer dan dinding sel sekunder pada jaringan yang akan membentuk kayu (Kallas et al., 2005).
Transformasi Genetik Tanaman
Teknik transformasi genetik merupakan salah satu metode penting dalam biologi tanaman. Teknik transformasi genetik dapat dipergunakan untuk mempelajari regulasi gen, identifikasi f h g s i gen, pengujian metabolisme, mempelajari fisiologi serta perkembangan tanaman (Knight, 1992; Walkerpeach dan Velten, 1994). Keberhasilan dalam melakukan rekayasa genetika memerlukan beberapa faktor yaitu : tersedianya gen yang diinginkan, tersedianya cara untuk mentransfer dan mengintegrasikan gen tersebut ke dalam sel tanaman dan cara untuk meregenerasikan tanaman transgenik, dan kemampuan tanaman untuk mengekspresikan gen yang telah diintroduksikan. Metode transformasi genetik untuk mengintroduksikan gen terpilih ke dalam sel tanaman dapat dilakukan dengan beberapa cara antara lain menggunakan Agrobacterium (Sudarsono, 1994).
Terdapat dua spesies Agrobacterium yang bersifat pathogen, yaitu Agrobacterium tumefaciens sebagai penyebab penyakit tumor (crown gall) dan Agrobacterium rhizogenes sebagai penyebab penyakit akar rambut (hairy root) pada berbagai tanaman dikotil (Armitage et al., 1 987). Agrobacterium tumefaciens merupakan bakteri aerob obligat gram negatif yang hidup alami di tanah, secara genetik dapat mentransformasi sel inang dan secara agronomi merupakan penyakit yang penting menyerang tanaman dikotil. Interaksi antara Agrobacterium dan sel tanaman adalah contoh alami yang diketahui dapat mentransfer DNA (Deoxiribosa Nucleic Acid) antar kingdom. Pada proses ini, DNA dipindahkan dari Agrobacterium ke dalam inti sel tanaman. Ekspresi dari DNA yang ditransfer (T-DNA) mengakibatkan pertumbuhan tumor pada tanaman inang. Gen yang dibawa T-DNA membawa gen-gen yang terlibat dalam sintesis hormon pertumbuhan tanaman dan produksi opin (Sheng dan Citovsky, 1996).
dalam plasmid Ti yang berukuran 200 kb. Sedangkan daerah vir yang berukuran 35 kb terdiri dari tujuh lokus utarna (vir A, vir B, vir C, vir D, vir E, vir G, dan vir H). Gen-gen vir mensintesis protein virulens yang berperan untuk menginduksi terjadinya transfer T-DNA dan integrasi T-DNA ke tanaman. Gen vir berekspresi jika terdapat inducer yang antara lain berupa senyawa monosiklik fenolik seperti acetosyringone dan monosakarida seperti glukosa dan galaktosa. Disamping itu, kondisi pH juga mempengaruhi ekspresi gen vir. Nilai pH yang sesuai berkisar 5,O-5,s. Senyawa fenolik dan monosakarida terbentuk pada saat tanaman dikotil mengalami luka dan proses ini jarang terjadi pada monokotil (Sheng dun Citovsky, 1996).
Interakasi antara Agrobacterium dengan sel tanaman didahului dengan penginderaan (sensing) Agrobacterium terhadap sel rentan yang luka. Mekanisme penginderaan ini terjadi secara kimiawi dimana sel tanaman yang luka menghasilkan suatu metabolit yang berperan sebagai isyarat bagi Agrobacterium. Metabolit tersebut dapat berupa senyawa gula, asam amino, atau senyawa fenol. Dengan adanya isyarat tersebut maka Agrobacterium akan bergerak aktif menuju sel tanaman target. Gerakan yang bersifat kemotaksis dipandu oleh senyawa yang disekresikan oleh sel tanaman rentan yang luka (Schaad, 1988). Interaksi dilanjutkan dengan terjadinya kontak antara Agrobacterium dengan sel tanaman target. Untuk memperkuat kontak ini Agrobacterium mengeluarkan suatu metabolit, yaitu
P- 1,2-glukan. Beberapa gen dalam kromosom Agrobacterium
diketahui merupakan penyandi enzim yang berperan dalam sintesis berbagai senyawa glukan, yaitu chvA, chvB dan exoC. Gen lain pada kromosom yang peranannya seperti ketiga gen tersebut adalah cel, yang berperan dalam sintesis senyawa selulosa fibril (Douglas et al., 1985).Dengan menggunakan satu plasmid Ti dari Agrobacterium, maka beberapa transgen dapat digabung dan ditempatkan di antara T-DNA dan selanjutnya diintegrasikan ke dalam genom tanaman. Hal penting dalam proses transformasi melalui Agrobacterium tumefaciens ini adalah transfer T-DNA ke inti tanaman target, integrasi T-DNA tersebut ke dalam genom tanaman target yang diinduksi oleh ekspresi gen-gen vir serta ekspresi gen-gen yang tertransformasi (Cheng et al., 1998). Selain itu integrasi T-DNA yang membawa transgen ke dalam genom resipien, akan mengalami sedikit pengaturan kembali secara intra dan intermolekul, untuk memulihkan sistem iranskripsi dan translasi genom tanaman resipien. Transformasi melalui Agrobacterium lebih menjamin kestabilan genom tanaman resipien (Sheng dan Citovsky, 1996).
Seleksi Tanaman Transgenik
Tanaman transgenik yang terseleksi dapat diamati dengan adanya pembentukan tumor pada sel-sel tanaman yang mengalami transformasi atau dapat juga diamati melalui adanya pertumbuhan dalam kultur yang bebas hormon. Selain itu dapat juga digunakan penanda seleksi yang disisipkan pada T-DNA pada sel-sel yang mengalami transformasi (Nakas dan Hagedors, 1990).
Salah satu contoh penanda seleksi adalah gen ketahanan terhadap antibiotik yaitu gen resisten terhadap kanamisin yang telah berhasil digunakan sebagai penanda seleksi yang dapat terekspresi pada fenotipe untuk transformasi pada beberapa spesies tanaman. Resistensi terhadap kanamisin telah berhasil digunakan sebagai penanda seleksi pada beberapa tanaman. Resistensi terhadap kanamisin ini disebabkan adanya gen nptII yang diperoleh dari transposon Tn5. Gen ini menyandi enzim neomisin fosfotransferase dan cara pewarisannya pada tanaman transgenik mengikuti pewarisan hukum Mendel untuk gen-gen dominan (Nakas dan Hagedors, 1990).
Regenerasi In vitro
memperoleh regenerasi yang efisien sebagian besar dipusatkan pada pemilihan bagian tanaman yang paling responsif serta penentuan jenis dan konsentrasi zat pengatur tumbuh yang efektif. Perlakuan lain yang kadang-kadang perlu diuji adalah cahaya, panjang penyinaran serta reaksi dalam sub kultur (Bhaskaran dan Smith, 1990).
Kultur Jaringan
Kultur jaringan merupakan suatu metode untuk mengisolasi bagian dari tanaman seperti protoplasma, sel, sekelompok sel, jaringan, dan organ serta menumbuhkannya dalam kondisi aseptik, sehingga bagian-bagian tersebut dapat memperbanyak diri dan beregenerasi menjadi tanaman utuh kembali (Gunawan, 1992). Selain untuk perbanyakan tanaman, teknik ini juga dapat digunakan untuk memperbaiki tanaman, menghasilkan tanaman bebas virus, produksi metabolit sekunder dan preservasi tanaman (Hartmann et al., 1990).
Teori yang mendasari teknik ini adalah konsep totipotensi yaitu sel yang hidup memiliki kemampuan untuk berproduksi, membentuk organ dan berkembang menjadi individu sempurna jika ditempatkan pada media dan lingkungan yang sesuai untuk pertumbuhan (Pierik, 1 987). Keberhasilan menggunakan metode kultur jaringan dalam mempengaruhi pertumbuhan dan morfogenesis, sangat tergantung pada jenis dan fisiologi eksplan yang dikulturkan (seperti organ yang digunakan, umur fisiologi, umur saat diambil, dari tanaman asal, ulcuran
dan
kualitas tanaman asal), media yang digunakan, dan kondisi fisikkultur. Faktor-faktor tersebut dapat berinteraksi satu dengan yang lainnya. Eksplan
Pada dasarnya setiap bagian tanaman dapat digunakan sebagai eksplan. Pernilihan material eksplan yang tepat akan mempengaruhi kesuksesan kultur jaringan, baik dari segi organ, ukuran, umur, dan cara mengkulturkannya
(George dan Sherrington, 1 984).
daya hidupnya bila dikulturkan, sementara jika terlalu besar akan mempersulit untuk mendapatkan eksplan yang steril dan dalam proses manipulasinya (George dan Sherrington, 1984).
Kepadatan eksplan yang ditanam dalam tiap botol juga mempengaruhi diferensiasi sel. Semakin banyak jumlah eksplan tiap botol, maka semakin banyak jumlah sel yang tidak berdiferensiasi. Volume media kultur diduga ada interaksinya dengan kepadatan eksplan dalam mempengaruhi pertumbuhan dan perkembangan kultur. Interaksi ini diduga berhubungan dengan menurunnya senyawa inhibitor dalam media (George dun Sherrington, 1984).
Media
Media kultur jaringan pada prinsipnya harus bisa menyediakan unsur-unsur hara yang diperlukan untuk pertumbuhan seperti tanamm dilapang. Keberhasilan dalam penggunaan metode kultur jaringan sangat tergantung pada media yang digunakan. Pemilihan komposisi media dan jenis media tergantung pada jenis tanaman yang dikulturkan, faktor aerasi, dan bentuk pertumbuhan dari deferensiasi yang diinginkan (Pierik, 1 987).
Media kultur jaringan tanaman tidak hanya menyediakan unsur-unsur hara makro
(N,
P, K, Ca, Mg, dan S) dan mikro (Fe,Mn,
B,Cu,
dan Mo), tetapi juga karbohidrat yang pada umumnya berupa gula untuk menggantikan karbon yang biasanya didapat dari atrnosfer melalui fotosintesis. Hasil yang lebih baik juga akan diperoleh, bila ke dalam media tersebut ditambahkan vitarnin-vitamin, asam amino,dan
zat pengatur tumbuh. Pada keadaan tertentu media kultur jaringanjuga dilengkapi dengan arang aktif (Gunawan, 1992).
Dalam kultur jaringan terdapat dua zat pengatur tumbuh tanaman yang penting yaitu auksin dan sitokinin. Auksin berperan dalam merangsang pembentukan kalus, pemanjangan sel, pembesaran dan pembentukan akar. Beberapa eksplan secara alamiah memproduksi cukup auksin. Pengaruh sitokinin adalah merangsang pembelahan sel dan multiplikasi tunas (George dan Sherrington, 1984). Keseirnbangan auksin dan sitokinin pada media tumbuh juga
akan
menentukan arah perkembangan eksplan. Tunas akan terbentuk bilaperbandingan konsentrasi auksin lebih tinggi dari sitokinin (Gunawan, 1992). Kondisi fsik kultur
Kondisi fisik atau kepadatan media berpengaruh terhadap potensial air dan tekanan osmotik, serta penyerapan hara tanaman. Kepadatan media ditentukan oleh konsentrasi agar, pH media dan penambahan arang aktif. Konsentrasi agar semakin tinggi dapat mengurangi difusi persenyawaan dari dan ke eksplan, sehingga pengambilan hara dan zat pengatur tumbuh berkurang, sedangkan zat penghambat dari eksplan tetap berkumpul di sekitar eksplan. Selain agar, ada juga zat pemadat yang lain, yaitu gelrite yang dapat membentuk gel yang lebih
bening, pada konsentrasi 0,l-0,2% sudah dapat memadatkan media.
Derajat keasaman (pH) merupakan ha1 penting yang hams diperhatikan dalam penyiapan media kultur jaringan tanaman. Karena pH dapat mempengaruhi perturnbuhan dan perkembangan eksplan yaitu dapat mempengaruhi tersedianya nutrisi dan hormon pada jaringan tanaman serta mempengaruhi f h g s i membran sel dan pH sitoplasma. Pengaturan pH selain memperhatikan kepentingan fisiologi sel, juga hams memperhatikan : (1) kelarutan garam-garam penyusun media, (2) pangaturan pengambilan zat-zat pengahu tumbuh dan gararn-garam lainnya, dan (3) efisiensi pembekuan agar media (George & Sherrington, 1984). Keasaman media pada umumnya berkisar antara 5,5-5,8 sebelum disterilisasi (Gunawan, 1 992).
Kondisi lingkungan kultur
intensitas cahaya (George dun Sherrington, 1984). Kualitas cahaya yang paling baik untuk pertumbuhan kultur adalah putih. Banyak penelitian menunjukkan bahwa penggunaan panjang penyinaran selama (14-16) jam memberikan hasil yang baik. Intensitas cahaya dari lampu flourescent adalah antara (1000-4000) lux dan ditempatkan dengan jumlah lampu dan kekuatan tertentu pada jarak (40-50) cm
dari
tabung kultur, untuk luas area tertentu. Suhu di dalam ruang kultur oleh banyak peneliti dilaporkan pada kisaran (25-28) OC memberikan pengaruh yang baik untuk pertumbuhan tanaman in vitro. Suhu optimum untuk pertumbuhankultur
jaringan tergantung dari jenis tanaman dan tempat turnbuh alamidari
tanaman tersebut (Gunawan, 1992).Induksi Embrio somatik
Embriogenesis somatik yaitu suatu proses perkembangan nonseksual yang menghasilkan suatu sel embrio bipolar yang berasal dari jaringan somatik. Tahap- tahap perkembangannya serupa dengan embriogenesis normal dan menghasilkan embrio tanpa hubungan vaskular dengan jaringan asalnya (Haccius, 1978). Embriogenesis sornatik memiliki dua pola perkembangan yaitu embriogenesis langsung (direct embriogenesis), yaitu embrio langsung terbentuk pada eksplan tanpa melalui proses pengkalusan, dan embriogenesis tak langsung (indirect embriogenesis), yaitu sebelum terbentuk embrio, eksplan membentuk kalus terlebih dahulu. Embriogenesis langsung secara in vitro umurnnya terjadi pada sel-sel eksplan yang masih muda Quvenil) sedangkan embriogenesis
tak
langsung terjadi pada sel-sel yang telah mengalami diferensiasi, pembelahan sel, transformasi menjadi sel embriogenik.Embrio somatik dapat dicirikan dari strukturnya yang bipolar, yaitu mempunyai dua calon meristem, yaitu meristem akar
dan
meristem tunas.Dengan memiliki struktur tersebut maka perbanyakan melalui embrio somatik lebih menguntungkan daripada pembentukan tunas adventif yang unipolar. Disarnping stdturnya, tahap perkembangan embrio somatik menyerupai embrio zigotik. Secara spesifik tahap perkembangan tersebut dimulai
dari
fase globular, fase hati, fase torpedo, dan planlet (Henry et.al., 1998 dalam Gaj, 2001).proses munculnya embrio somatik adalah kandungan auksin pada media, campuran nitrogen yang ditambahkan sebagai nutrisi. Faktor fisik seperti temperatur, intensitas cahaya, fotoperiode, udara, keadaan media dan kecepatan pengocokan juga telah dilaporkan mempengaruhi embrio somatik. Temperatur optimum adalah spesifik untuk setiap spesies clan tahap perkembangan. Perlakuan panas atau dingin pada tahap tertentu dapat meningkatkan embriogenesis
dan
perkecambahan dari embrio somatik dan propagul lain untuk perkembangan yang lebih lengkap (Vajrabhaya, 1 988).
Analisis Tanaman Transgenik
Analisis tanaman transgenik dalam proses transformasi genetik tanaman dapat dilakukan pada berbagai tahapan, yaitu : introduksi gen, transkripsi, translasi,
dan
pengujian efektifitas protein yang dihasilkan. Dewasa ini telah dikenal berbagai teknik analisis tanaman transgenik seperti : Uji histokirniaP-
glucuronidase (uji Gus), analisis Polymerase Chain Reaction (PCR), analisis Sothern Blot, Northern, Western, ELISA dan lain-lain. Untuk gen ketahanan terhadap serangga, pengujian efektifitas protein yang dihasilkan terhadap serangga target dapat d i l w a n melalui uji hayati, uji pakan dan sebagainya.Uji GUS
Uji histokimia P-glucuronidase (uji GUS) dapat dilakukan pada tahap awal segera setelah ko-kultivasi maupun setelah gen terintegrasi dengan stabil di kromosom. Warna biru yang muncul pada sel atau jaringan menunjukkan hasil transformasi positif. Warna biru disebabkan oleh reaksi substrat X-gluc (5-bromo-
4-chrorno-3-irtdoZyZ-~-D-glucuronide) dengan enzim P-glucuronidase menjadi
suatu senyawa perantara yang kemudian melalui reaksi dimerisasi oksidatif membentuk senyawa dichloro-dibromoindigo (CIBr-Indigo) yang berwarna biru (Stomp, 1992). Warna biru tersebut menunjukkan telah terekspresinya gen gus-A.
dipergunakan sebagai penanda pada sistem transformasi tanaman dan seringkali digunakan untuk mempelajari fimgsi promoter (Jefferson, 1987).
Analisis Polymerase Chain Reaction (PCR)
PCR merupakan teknik analisis tingkat DNA, yang menggunakan penggandaan urutan basa DNA spesifik secara in vitro, seperti pada cara replikasi DNA, dengan bantuan enzirn polimerase
dan
pemanfaatan perubahan sifat fisikDNA terhadap suhu (Davis et al., 1994). Keuntungan teknik PCR diantamnya adalah analisisnya cepat, tidak diperlukan DNA dalam jumlah banyak, dapat dilakukan pada fase awal pertumbuhan dan metode ekstraksi DNAnya relatif sederhana. Dalam transformasi genetik, teknik PCR dapat dipergunakan untuk mengamplifikasikan gen yang telah diintroduksi ke sel tanaman target untuk membuktikan keberadaannya.
Dengan reaksi PCR, DNA dapat diperbanyak dengan menggunakan enzim polymerase yang dihasilkan oleh bakteri termofilik melalui serangkaian pengaturan suhu yang berbeda selama waktu tertentu pada satu siklus perbanyakan. Dalam satu siklus perbanyakan, terjadi penggandaan urutan basa cetakan. Masing-masing siklus terdiri dari tiga tahap yaitu tahap denaturasi DNA, penempelan (annealing) dan sintesis DNA (Krawetz, 1989). Pada tahap pertama DNA didenaturasi dengan meningkatkan suhu sehingga 95 O C selama 60-90 detik. Tahap berikutnya suhu diturunkan hingga 55 "C atau antara 40-60 "C tergantung panjang primer selama 30-60 detik untuk penempelan primer ke DNA target secara spesifik. Pada tahap terakhir suhu dinaikkan kembali sekitar 72 OC untuk sintesis DNA yang dimulai dari ujung 3' hidroksil pada masing-masing primer (Krawetz, 1989; Cha dun Thilly, 1993). PCR merupakan metode yang sangat sensitif sehingga dengan hanya satu molekul DNA dapat memperbanyak DNA jutaan kali, sehingga sangat bermanfaat baik pada penelitian maupun penggunaan
komersial (Promega, 1996). Southern Blot
pemotongan DNA tanaman transgenik dengan enzim restriksi tertentu yang memotong pada situs tunggal dalam DNA plasmid. Produk hibridisasi berasal dari gen yang diintroduksikan dari hasil pemotongan DNA genomik sehingga polimorfisme yang terbentuk menunjukkan sisi integrasi yang berbeda (Casas et
al., 1995). Selain dipergunakan untuk membedakan kejadian transformasi, analisis Southern blot dapat membedakan integrasi ekstra kromosomal dan kromosomal (Davis et al., 1994). Kelemahan penerapan metode Southern blot adalah memerlukan sejumlah DNA yang relatif banyak dengan kemurnian tinggi dan waktu pelaksanaan relatif lama (Sambrook et al., 1989).
Northern Blot
Hibridisasi Northern merupakan suatu prosedur yang dipergunakan untuk identifikasi dan analisis transkip RNA (Kafatos et al., 1979). RNA tidak dapat berikatan secara efisien pada membran, sehingga dalam analisis northern dipergunakan suatu membran spesifik dimana RNA dapat berikatan secara kovalen. Ikatan RNA tersebut dapat dihibridisasi dengan menggunakan probe RNA radioaktif atau DNA utas tunggal (Freifelder, 1995).
Dalam analisis Northern, sekuen RNA spesifik dideteksi menggunakan teknik bloting yaitu RNA ditransfer
dari
agarose ke membran. Hasil bloting dianalisis melalui proses hibridisasi dengan probe RNA. RNA merupakan bentuk utas tunggal, sehingga dapat membentuk struktur sekunder melalui pasangan basa intramolekul, sehingga hams dielektroforasi di bawah kondisi denaturasi. Denaturasi dilakukan dengan penambahan formaldehid ke gel maupun loadingbuHer, atau perlakuan glyoxal dan dimethyl sulfoxide (DMSO) pada loading
bufler. Berbagai bahan untuk denaturasi gel RNA telah dipergunakan termasuk
formaldehid, glyoxal dan methilmercuri klorida yang sangat toksik. Total RNA dapat dipergunakan untuk proses hibridisasi northern, akan tetapi total RNA biasanya memberikan hasil yang kurang memuaskan sebab terjadi hibridisasi nonspesifik. Meskipun sedikit, molekul rRNA akan menghasilkan signal hibridisasi yang lebih kuat (Ausubel, 1995).
Western Blot
elektroforesis dan ditransfer ke suatu membran sehingga protein akan berikatan secara kovalen. Sebagai probe dipergunakan utas ganda DNA radioaktif dengan cara penggabungan dari radioaktivitas dengan pita-pita protein yang menunjukkan bahwa protein tertentu merupakan DNA binding protein (Dale 1995; Freifelder, 1995).
Prinsip dasar Western blot adalah identifikasi pemisahan protein yang tidak terlabel dengan SDS gel elektroforesis polyacrilamide (PAGE) yang didasarkan pada immunoradioaktivitasnya dengan menggunakan antibodi monoklonal. Kemudian protein ditransfer ke membran dan diberi perlakuan awal untuk mereduksi ikatan nonspesifik dari antiserum ke membran. Inkubasi membran dilakukan dengan antiserum spesifik, kemudian diinkubasi dengan antibodi yang berkonjugasi dengan reagen pendeteksi dan berikatan pada antiserum primer. Setelah itu diikuti deteksi dari immunoreaksi diantara antiserum primer dan target protein spesifik (Davis et al., 1994).
Park et. al. (2004) telah melakukan uji ekspresi pada tanaman poplar yang telah ditransformasi dengan menggunakan teknik Western blot. Analisis Polymerase Chain Reaction (PCR) juga telah dilakukan sebelum Western blot. Hasil uji ekspresi memberikan nilai positif dengan munculnya pita-pita. Western blot adalah teknik yang paling tepat untuk uji ekspresi protein pada tanaman yang telah ditransformasi.
Uji Hayati
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilaksanakan di laboratorium Biologi Molekuler Tanaman, Pusat Penelitian Bioteknologi
-
LIPI, Cibinong, mulai bulan Agustus 2006 sarnpai dengan Agustus 2007.Bahan dan Alat
Eksplan yang digunakan pada transformasi adalah kalus clan tunas majemuk yang berasal dari embrio. Sedangkan pucuk, batang dan daun berasal dari biji Acacia mangium Willd yang telah dikecambahkan pada media MS. Biji berasal dari Kebun Botani PUSPIPTEK, Serpong.
Vektor yang digunakan dalam transformasi adalah Agrobacterium tumefaciens LBA4404 yang membawa gen xyloglucanase, gen nptII, dan promotor 35s pada plasmid pAaXEG300. Vektor ini diperoleh Pusat Penelitian Bioteknologi-LIP1 dari RISH (Research Institute for Sustainable Huymanosphere), Kyoto university, Jepang dalam rangka kerjasama Bioteknologi Kehutanan.
Media yang digunakan yaitu media MS (Murashige Skoog), media ?4 MS dengan penambahan Kanarnisin, Carbenicillin, TDZ (Thidiamron), dan IAA. Bahan kimia yang digunakan pada kegiatan sterilisasi adalah deterjen, fungisida Dithane M-45, Masalgin, Ethanol 70%, Byclin, dan akuades steril. Bahan kimia lain yang juga digunakan adalah Yeast Extract 1 grtl, Saytone 4 grll, Peptone 5 grll, Sucrose 5 grll, MgS04
.
7 H20 0.246 grll, dan Bacto Agar 14 grll.Alat dan barang aus yang digunakan pada penelitian ini antara lain aluminium foil, botol kultur, otoklaf, petridish, botol alkohol, corong, gelas piala, gelas ukur, hot plate magnetic, stirer, inkubator, laminar airflow, bunsen, pH meter, shaker, spatula, mikro pipet, dan timbangan analitik.
Metode Penelitian Persiapan eksplan
Bahan tanaman yang digunakan untuk menyediakan eksplan yang akan ditransformasi adalah biji Acacia mangium Willd. Prosedur sterilisasi yang digunakan adalah yang telah dikembangkan oleh Pusat Penelitian Bioteknologi -
LIPI, Cibinong. Sterilisasi dilakukan dengan cara biji Acacia mangium Willd diberi deterjen (Sunlight) kemudian diletakkan pada air mengalir selama 10 menit. Kemudian, biji Acacia mangium Willd direndam pada air panas (80 OC) selama 30 menit, rendaman biji Acacia mangium Willd dikocok dengan menggunakan shaker selama 30 menit dengan menambahkan Dithane (4 g/100 ml), dibilas 3 kali dengan menggunakan akuades. Tahap selanjutnya biji Acacia mangium Willd direndam pada larutan Masalgin (4 g1100 ml), dikocok dengan menggunakan shaker selama 30 menit, kemudian dibilas 3 kali dengan akuades steril di dalam
laminar airflow. Setelah itu, biji Acacia mangium Willd direndam pada larutan
Bayclin yang telah diencerkan 1.5 kaIi selama 10 menit, kemudian dibilas 3 kali dengan akuades steril. Selanjutnya, biji Acacia mangium Willd direndam pada Ethanol 70% selama 5 menit, lalu dibilas dengan akuades steril3 kali. Biji Acacia
mangium Willd dipindahkan ke botol steril, kemudian ditutup rapat dengan
aluminium foil dan dikocok dengan menggunakan shaker selama 24 jam.
telah steril, dengan cara biji dikupas untuk diambil embrionya cialam laminar airflow. Untuk mendapatkan kalus yang embriogenik, embrio dikecambahkan pada media MS yang dilengkapi dengan 0,25 mg/l IAA, 1 mg/l TDZ, 20 grA
sukrosa, dan 2 g/l gelrite selama 3-4 bulan. Biji Acacia mangium Willd yang tidak dikupas kulit bijinya, dikecambahkan pada media MS tanpa zat pengatur tumbuh selarna 3-4 bulan.
Optimasi media seleksi
Untuk mendapatkan konsentrasi antibiotik Kanamisin
dan
Carbenicillinyang tepat dalam menyeleksi tanaman Acacia mangium Willd transgenik, dilakukan uji efektifitas konsentrasi antibiotik pada media MS yang dilengkapi dengan 0,25 mg/l IAA, 1 mg/l TDZ, 20 gr/l sukrosa, dan 2 grA gelrite. Kanamisin dan carbenicillin ditarnbahkan pada media seleksi setelah media diotoklaf dan dibiarkan dingin sampai suhu sekitar 50 OC. Konsentrasi Kanamisin yang diuji adalah 0, 100, 200, dan 400 mg/l. Sementara konsentrasi Carbenicillin yaitu 225 mg/l.
Kultur Agrobacterium tumefaciens
Agrobacferium tumefacciem yang mengandung pAaXEG300 ditumbuhkan pada media YES-cair yang telah ditarnbah 50 mg/l Kanamisin. Diinkubasi dan dikocok dengan menggunakan shaker (1 50 rpm) pada suhu 28 OC selama 24 jam. Selanjutnya diremajakan pada media YES-cair baru, diinkubasi dan dikocok kembali dengan menggunakan shaker (150 rpm) pada suhu 28 OC selama 24 jam. Suspensi bakteri ini disentrifbgasi dengan kecepatan 3000 rpm selama 10 menit pada suhu 4 OC untuk diambil peletnya. Kemudian pelet dibilas dengan air steril clan disentrifbgasi kembali (3000 rpm) selama 10 menit pada suhu 4 OC. Pelet bakteri kemudian dilarutkan dengan aquades steril, diukur pada
selanjutnya siap digunakan untuk transformasi.
Transformasi dan regenerasi
hari pada media ?4 MS eksplan dipindah ke media seleksi dengan konsentrasi kanamisin 0, 100, 200, dan 400. Pengamatan dilakukan terhadap eksplan yang tumbuh pada tiap media seleksi.
Eksplan yang tumbuh pada media seleksi selanjutnya dipindah pada media regenerasi
yang
terdiridari
media MS yang dilengkapi dengan 0,25 mg/l IAA, 1 mg/l TDZ, 20 grll sukrosa, dan 2 g/l gelrite. Pengamatan dilakukan terhadap persentase eksplan kalus yang membentuk somatik embrio, jumlah daun yang tumbuh pada eksplan pucuk dan batang. Untuk menumbuhkanakar
pada eksplan pucuk dan batang, eksplan dipindah ke media MS yang dilengkapi den- 0,25 mg/l IAA, 20 grll sukrosa, dan 2 g/l gelrite.Uji ekspresi Western blot
Untuk mengetahui keberhasilan transformasi, pada eksplan hasil kokultivasi dan eksplan yang tumbuh di media seleksi dilakukan uji ekspresi Western blot. Metoda yang digunakan adalah metoda yang telah dilakukan di Pusat Penelitian Bioteknologi-LIPI, Cibinong sesuai dengan metoda yang telah dilakukan Park et al. (2004) pada tanaman poplar yang telah ditransformasi.
Ekstraksi protein. Bahan tanaman (2-3 tangkai daun) yang diduga transgenik berdasarkan uji media seleksi yang mengandung Kanamisin dirnasukkan ke tabung Eppendorf yang berisi 200 p1 buffer pengekstrak (Sodium acetat
pH
5,5), digerus dengan penggerus plastik biru sampai dengan homogen. Kemudian dibiarkan selama 30 menit di dalam es. Setelah disentrifbgasi dengan kecepatan 12.000 rpm selama 5 menit pada suhu 4 "C, kemudian supernatan diambil dan disentrifugasi kembali dengan kecepatan 12.000 rpm selama 5 menit pada 4 OC.HASIL
DAN
PEMBAHASAN
Sterilisasi dan penanaman biji Acacia mcmgium Willd
Metoda sterilisasi yang digunakan pada penelitian ini berdasarkan atas metoda standar Pusat Penelitian Bioteknologi-LIPI, Cibinong. Biji Acacia
mangium Wiild berasal dari Kebun Botani PUSPIPTEK, Serpong. Pada Tabel 1
dapat dilihat persentase biji yang berkecambah pada media MS.
Tabel 1 Persentase biji Acacia mangium Willd yang berkecambah pada media MS tanpa zat pengatur tumbuh
Percobaan Jumlah biji yang Persentase biji yang
dikecambahkan berkecambah
1 150 12011 50 (80%)
Tabel 1 menunjukkan bahwa rata-rata persentase biji Acacia mangium
Willd yang berkecambah sebesar 86,67%. Persentase kecambah ini dipengaruhi oleh kematangan biji secara fisiologis, kesehatan biji, dan perlakuan pemecahan dormansi. Biji yang telah matang secara fisiologis akan cepat berkecambah. Secara visual biji yang telah matang secara fisiologis mempunyai ciri-ciri yaitu berkulit hitam mengkilat, ukuran lebih besar dibanding biji yang lain,
dan
biji lebih bersih. Selain itu, Acacia mangium Willd memiliki kulit biji yang sangat keras, sehingga perlu perlakuan pemecahan dormansi. Pemecahan dormansi dapat dilakukan dengan 3 cara yaitu, menggunakan bahan kirnia seperti perendaman pada HzS04, perlakuan secara biologi seperti penjemuran biji dibawah sinar matahari, dan perlakuan secara mekanis seperti pengamplasan clan perendaman pada air 80 "C.
Pada penelitian ini, pemecahan dormansi dilakukan secara mekanis yaitu dengan cara perendarnan pada air 80 OC selama 30 menit. Biji mulai berkecambah pada hari ke 3 setelah tanam pada media MS.permukaan kulit biji seperti debu, cendawan, dan bakteri. Semakin besar ukuran eksplan maka semakin luas permukaannya dan kemungkinan adanya kontaminan juga semakin besar. Selain itu, kontaminan juga dapat berasal dari alat diseksi
yang digunakan.
Gambar 3 Eksplan Acacia mangium Willd berumur 3 bulan pada media MS (material untuk transformasi)
Gambar 3 menunjukkan pertumbuhan biji Acacia mangium Willd pada media MS dan diinkubasi pada suhu 20-22 "C. Pertumbuhan tanaman secara in vitro dipengaruhi oleh jenis eksplan, media, kondisi fisik kultur, dan kondisi lingkungan kultur seperti cahaya dan suhu ruangan kultur. Kualitas cahaya yang paling baik untuk pertumbuhan kultur adalah put& sedangkan suhu optimum untuk pertumbuhan kultur tergantung dari jenis tanaman dan tempat tumbuh alami
dari
tanaman tersebut, namun umumnyauntuk
tanaman tropis berkisar 25 "C.Induksi embrio somatik Acacia mangium Willd
Tabel 2 Persentase embrio dari biji Acacia mangium Willd yang berkalus dan tumbuh tunas majemuk pada media MS
+
0,25 mg/l IAA+
1 mg/l TDZPercobaan Jumlah embrio Persentase embrio Persentase tumbuh yang diinduksi yang mengkalus tunas majemuk
1 100 9011 00 (90%) 151100 (15%)
Tabel 2 menunjukkan bahwa rata-rata persentase embrio dari biji Acacia mangium Willd yang mernproduksi kalus sebesar 92,33%. Pertumbuhan kalus sangat dipengaruhi oleh media tanam yang dilengkapi dengan unsur hara
dan
zat pengatur tumbuh. Persentasetunas
majemuk yang tumbuh sebesar 1 1,67%. Tunas majemuk ini muncul karena tidak sempurnanya pemisahan yang dilakukan antara embrio dengan titik tumbuh, sehingga pertumbuhantunas
majemuk yang tidak diharapkan terjadi.Gambar 4 Kalus yang berasal dari embrio Acacia mangium Willd pada media MS
+
0,25 mg/l IAA+
1 mg/l TDZ(A. Embrio berumur 12 rninggu, B. Kalus , dan C. tunas majemuk)
Pengamatan yang dilakukan pada pembentukan kalus diawali dengan terjadinya pemanjangan dan pembengkakan pada hari ke 3 setelah tanam pada media induksi dan terjadi perubahan warna, yaitu warna embrio sebelum tanam adalah kuning, setelah tanam berubah menjadi kuning keputihan. Kalus mulai muncul pada hari ke 12. Zat pengatur tumbuh yang digunakan pada penelitian ini adalah IAA dari golongan auksin dan TDZ dari golongan sitokinin. TDZ merupakan jenis sitokinin yang paling kuat untuk pembelahan sel. Menurut Sulistiani (1997) bahwa keberhasilan TDZ dalam menghasilkan embrio somatik dari biji sengon adalah dengan menggunaan TDZ pada konsentrasi 1,s mg/l dengan lama induksi 8 minggu.
Mekanisme perubahan ekspresi gen tersebut ternyata berhubungan dengan banyaknya auksin eksogen. Dengan demikian, penggunaan auksin atau senyawa mirip auksin pada metode induksi embriogenesis somatik sangat diperlukan. Pada beberapa jenis tanaman, sitokinin digunakan dan dikombinasikan dengan auksin untuk induksi embriogenesis somatik.
Pengamatan yang dilakukan pada rninggu ke 12, dapat dilihat bahwa kalus berubah warm menjadi coklat. Siswanto (2000) menyatakan bahwa fenolik yang menyebabkan pencoklatan pada kopi arabika terlihat menghambat embriogenesis somatik. Menurut Figueora
et
al. (2001) pencoklatan pada jaringan yang disebabkan oleh akumulasi senyawa fenolik yang berlebihan penting untuk proses embriogenesis somatik pada tanaman kopi. Kemungkinan senyawa fenolik ini berperan sebagai signal untuk induksi diferensiasi. Kemungkinan lain, senyawa ini juga berperan sebagai faktor yang menginaktifkan senyawa-senyawa penghambat yang terdapat dalam kultur embriogenik.Kendala dalam penerapan embriogenesis yaitu peluang terjadinya mutasi lebih tin@ apabila menggunakan auksin dengan konsentrasi tinggi dalam jangka waktu lama, metode lebih sulit, ada p e n m a n daya morfogenesis dari kalus embriogenik karena sub kultur berulang serta memerlukan penanganan yang lebih intensif untuk memelihara kemampuan embriogenesis. Apabila tidak secara kontinyu dipelihara maka akan kehilangan kemampuan embriogenesis karena tertutupi kalus fiiabel yang tidak diinginkan.
Seleksi terhadap resistensi Kanamisin pada eksplan yang belum ditransformasi
Seleksi eksplan terhadap resistensi Kanamisin bertujuan untuk mengetahui konsentrasi Kanamisin yang menghambat pertumbuhan dan bahkan dapat menyebabkan eksplan tersebut mati. Eksplan yang digunakan pada penelitian ini yaitu; pucuk, batang, daun, kalus, dan tunas majemuk. Media seleksi yang digunakan diberi zat pengatur turnbuh 0,25 mg/l IAA dan 1 mg/l
TDZ.
Sedangkan antibiotik yang digunakan adalah 225 mg/l Carbenicillin
clan
perbedaan ketahanan terhadap Kanamisin dipengaruhi oleh adanya perbedaan genotip tanaman, umur jaringan, serta kondisi kultur jaringan.
Salah satu tahapan penting yang harus dilalui dalam proses transformasi genetik untuk memperoleh tanaman transgenik adalah seleksi. Tersedianya metode seleksi awal pada tanarnan transgenik sangat membantu dalam menyeleksi tanaman transforman. Tahap awal yang dapat dilakukan adalah dengan menumbuhkan tanaman hasil transformasi pada media seleksi yang mengandung antibiotik tertentu tergantung gen penanda yang dipakai.
Gen penanda yang digunakan pada penelitian
ini
adalah Kanamisin, sehingga perlu dilakukan uji coba ketahanan terhadap Kanamisin yang dilakukan hingga 8 minggu. Kanamisin merupakan kelompok antibiotika aminoglikosida yang efektif menghambat sintesis protein melalui aksinya terhadap ribosom sub- unit 30s dan bekerja efektif baik pada organisme prokariot maupun eukariot.Media seleksi yang digunakan diberi zat pengatur tumbuh 0,25 mg/l IAA dan 1 mg/l TDZ. Sedangkan antibiotik yang digunakan adalah 225 mg/l Carbenicillin dan Kanamisin dengan konsentrasi yang berbeda yaitu 0, 100, 200, dan 400 mg/l. Eksplan yang digunakan pada penelitian ini ada lima jenis, yaitu; pucuk, batang, daun, kalus, dan tunas majernuk. Eksplan yang mengalami nekrosis hingga rninggu ke 8 pengamatan pada media yang mengandung Kanarnisin dapat dilihat pada Tabel 3.
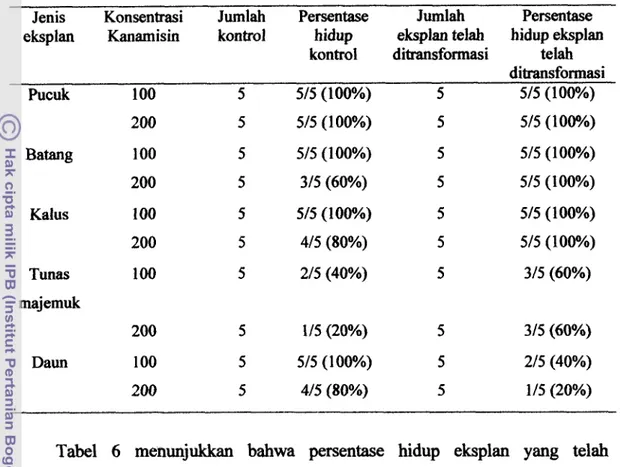
Tabel 3 Uji konsentrasi Kanamisin pada berbagai eksplan Acacia mangium
Willd yang tidak ditransformasi
Konsentrasi Persentase eksplan yang nekrosis
kanamisin pucuk batang Daun tunas kalus
(m@J1)
majemuk
0 015 (0) 015 (0) 015 (0) 015 (0) 015 (0)
membentuk kalus. Begitu juga pada eksplan yang ditumbuhkan pada media seleksi 100 mgh Kanamisin, tetapi eksplan tidak membentuk kalus. Konsentrasi Kanamisin 200 mg/l dan 400 mg/l menyebabkan nekrosis dan eksplan mengalami browning dan akhirnya mati. Eksplan tampak kering dan mengalami browning dan mati. Eksplan yang mengalami neksrosis tertinggi terlihat pada eksplan dam, sebesar 80% dan yang terendah adalah pada eksplan pucuk dan
tunas
majemuk sebesar 40%. Sebelumnya telah dilakukan pengujian yaitu eksplan pucuk ditumbuhkan pada konsentrasi Kanamisin 600 mg/l, pada minggu ke 2 tanaman mengalami nekrosis dan pada minggu ke 4 mengalami kematian.Penggunaan sistem seleksi antibiotik dilaporkan sering menyebabkan sebagian besar sel yang tertransformasi tidak atau sulit beregenerasi, diduga karena adanya penghambat pertumbuhan atau toksin yang dikeluarkan dari sel nontrmsgenik yang mati atau karena terganggunya transportasi senyawa esensial melalui jaringan mati tersebut (Haldrup et al., 200 1).
Seleksi tahap awal dengan menggunakan media yang mengandung antibiotik Kanamisin dan menentukan konsentrasi Kanamisin yang tepat sangat diperlukan, mengingat respon setiap tanaman terhadap antibiotik K a n d s i n berbeda-beda. Sel atau kalus yang tidak dilengkapi dengan sifat ketahanan Kanamisin akan berhenti tumbuh atau mati bila ditumbuhkan pada media seleksi yang mengandung Kanamisin. Dari penelitian optirnasi media seleksi ini, dihasilkan bahwa penambahan Kanamisin 400 mgll ke dalam media seleksi dapat digumkan sebagai konsentrasi yang menghambat pertumbuhan. Dosis ini digunakan pada media seleksi pada tahapan percobaan selanjutnya.
Penentuali trilai OD6o0 Agrubacterim tuntefens
Konsentrasi bakteri yang dapat diketahui dengan mengukur nilai ODm juga memberi pengaruh terhadap keberhasilan transformasi. Bakteri yang digunakan untuk menginfeksi sel tanaman sebaiknya bekteri yang sedang tumbuh aktif (fase logaritmik) (Rahmawati, 2006). Pucuk yang direndam pada suspensi bakteri
Tabel 4 Persentase eksplan Acacia mangium Willd yang hidup setelah direndam pada Agrobacterium tumefaciens selama 5 menit pada beberapa nilai OD600
Nilai ODm Jumlah eksplan Persentase eksplan yang hidup 0,4 4 1 41/41 (100%)
0 4 87 24/87 (27,58%)
Tabel 4 menunjukkan bahwa pada OD600 sebesar 0,6 eksplan pucuk yang telah direndam pada Agrobacterium tumefaciens selama 5 menit hanya 27,58% yang tumbuh tanpa mengalami gangguan pertumbuhan berlebihan dari
Agrobacterium tumefaciens, pengulangan yang dilakukan bahwa setelah nilai ODm diturunkan menjadi 0,4 terlihat bahwa 100% eksplan dapat tumbuh tanpa ada gangguan dari pertumbuhan Agrobacterium tumefaciens. Berdasarkan hasil inilah, maka nilai OD6()() yang digunakan untuk transforrnasi berikutnya adalah 0,4.
Masing-masing strain Agrobacterium mempunyai sensitivitas yang berbeda-beda terhadap antibiotik, sehingga perlu pengujian awal
untuk
menentukan jenis dan konsentrasi antibiotik yang sesuai untuk menghambat pertumbuhan Agrobacterium yang digunakan. Antibiotik yang
umum
digunakan untuk mengeliminasi Agrobacterium adalah Carbenicillin dan Cefotaxim. Keduanya temasuk kelompok P-laktam yang menghambat pembentukan dinding sel balcteri. Narnun, Carbenicillin sensitif terhadap enzim fl-laktamase yang dihasilkan oleh bakteri sehingga kurang efektif dalam mengeliminasiAgrobacterium pasca kokultivasi (Rahmawati, 2006).
Transforrtlasi berbagai eksplan Acacia msn&rn WilM
Keberhasilan proses infeksi melalui Agrobacterium tergantung kompatibilitas antara kultivar tanaman dengan isolat Agrobacterium yang digunakan. Isolat Agrobacterium yang kompatibel akan mampu menangkap signal
dari
tanaman yang terluka untuk memulai proses infeksi, sebaliknyatanaman yang kompatibel
akan
mampu memberi signal kepada AgrobacteriumHasil pengamatan hingga minggu ke 8 menunjukkan bahwa eksplan Acacia
mangium Willd yang telah ditransformasi dapat bertahan pada media seleksi. Pada
Tabel 5 dapat dilihat persentase hidup eksplan yang telah ditransformasi clan ditumbuhkan pada media seleksi dengan konsentrasi Kanamisin 400 mg/l sebesar.
Tabel 5 Persentase hidup eksplan Acacia mangium Willd yang telah ditransforrnasi pada minggu ke 8 pada media seleksi (0,25 mg/l IAA
+
1 mg/l TDZ
+
225 Carbenicillin+
400 mgfl Kanamisin)Jenis eksplan Jumlah Persentase Jumlah Persentase eksplan eksplan hidup eksplan eksplan hidup kontrol kontrol
Pucuk 5 40% 3 6 63,89%
Batang 5 60% 24 95,83%
Tunas majemuk 5 40% 26 48,39%
Daun 5 80% 40 2,5%
Hasil pengamatan pada berbagai eksplan Acacia mangium Willd hingga minggu ke 8 dapat dilihat pada Tabel 5. Secara
umum
eksplan yang telah ditransformasi memiliki persentase hidup yang tinggi dibandingkan dengan kontrol, kecuali pada eksplan daun, persentase hidup eksplan daun yang telah ditransformasi hingga minggu ke delapan lebih rendah yaitu sebesar 2,5% dibandingkan dengan kontrol sebesar 80%. Sedangkan persentase hidup eksplan yang telah ditransformasi tertinggi adalah pada eksplan batang sebesar 95,83%.Rendahnya persentase hidup eksplan daun yang telah ditransformasi diduga karena belum sesuainya metoda transformasi yang dilakukan. Nilai kerapatan bakteri juga mempengaruhi keberhasilan transformasi. Pada eksplan daun, kerapatan bakteri dapat d i m a n . Hasil transformasi yang telah dilakukan oleh Park et.al(2004) pada eksplan daun poplar menunjukkan adanya regenerasi dari eksplan dam yang telah ditransformasi, berupa munculnya tunas adventif pada bagian daun yang telah mengalami perlukaan.
bakteri, lama inokulasi, dan lama kokultivasi (Hinchee et al., 1988). Untuk jenis eksplan, semakin muda jaringan eksplan akan semakin mudah diinfeksi oleh bakteri. Pada penelitian ini, eksplan yang digunakan sebagai bahan transformasi adalah eksplan yang telah dikecambahkan pada media MS selama tiga bulan.
Persentase hidup eksplan yang telah ditransformasi hingga minggu ke 8 dipengaruhi oleh tumbuhnya bakteri pada media seleksi (MS +0,25 mg/l IAA
+ 1
mg/l TDZ+
225 mg/l Carbenicillin+
400 mg/l Kanamisin). Siswanto et al.(1997) menyatakan bahwa jumlah bakteri yang diperlukan dalarn proses infeksi suatu eksplan h m tepat. Jika jurnlahnya kurang, proses infeksi tidak efektif, sebaliknya jika jumlah bakteri terlalu banyak akan terjadi pertumbuhan bakteri yang berlebih (overgrowth). Akibatnya tingkat kompetisi bakteri sangat tinggi dan pertumbuhan eksplan terhambat atau mati sehingga proses infeksi tidak efektif
Lama inokulasi juga menentukan keberhasilan infeksi bakteri. Semakin lama waktu inokulasi, peluang infeksi semakin tinggi. Xnokulasi dilakukan selama 5 menit. Untuk eksplan yang berukuran besar dan tebal seperti pada penelitian ini adalah kalus dan
tunas
majemuk diperlukan waktu inokulasi yang lebih lama. Seperti yang telah dinyatakan Park et al. (2004) bahwa lamanya inokulasi eksplanpada suspensi bakteri untuk tanaman poplar adalah 5 menit. Begitu juga yang dilaporkan Hartati et al. (2005) bahwa waktu inokulasi yang efisien untuk pucuk
Acacia mangium Willd adalah selama 5 menit. Sementara untuk eksplan dam chrysanthemum, Boase et al. (1998) menyatakan bahwa perendaman eksplan pada suspensi bakteri selama 5 menit lebih baik daripada 1 menit.
Lama kokultivasi (inkubasi) antara bakteri dan eksplan juga sangat mempengaruhi keefektifan infeksi bakteri. Inkubasi yang terlalu cepat menyebabkan pertumbuhan bakteri kurang baik, sehingga belum marnpu menginfeksi sel-sel eksplan dengan sempurna. Sebaliknya, jika inkubasi terlalu lama akan terjadi pertumbuhan koloni bakteri yang berlebihan sehingga menghambat pertumbuhan atau bahkan mematikan eksplan. Hartati et al. (2004) menyatakan bahwa pada Acacia mangium Willd masa kokultivasi yang efektif adalah 1 hari. Begitu juga dengan Park et al. (2004) yang menyatakan bahwa
Xie dan Hong (2002), bahwa masa kokultivasi untuk tanaman Acacia mangium
Willd adalah 3 hari.
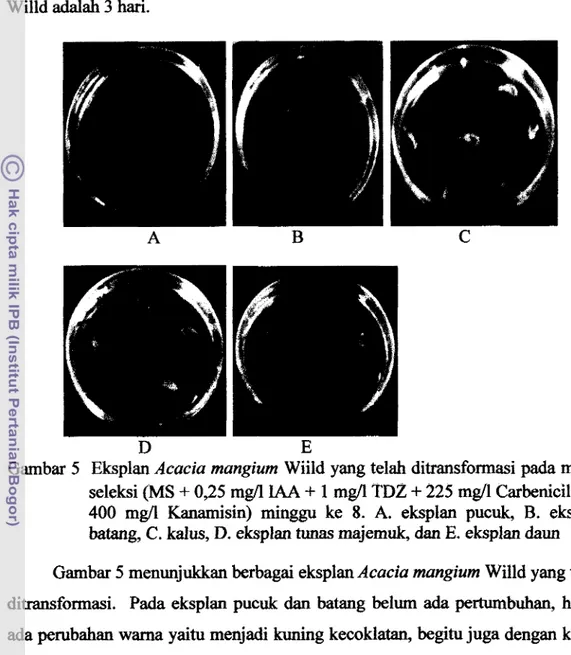
Gambar 5 Eksplan Acacia mangium Wiild yang telah ditransformasi pada media seleksi (MS
+
0,25 mgA LAA+
1 mg/l TDZ+
225 mgll Carbenicillin+
400 mgll Kanamisin) rninggu ke 8. A. eksplan pucuk, B. eksplan batang, C. kalus, D. eksplan tunas majemuk, dan E. eksplan daun Gambar 5 menunjukkan berbagai eksplan Acacia mangium Willd yang telah ditransformasi. Pada eksplan pucuk dm batang belum ada pertumbuhan, hanya ada perubahan warna yaitu menjadi kuning kecoklatan, begitu juga dengan kalus. Sementara pada eksplan tunas majemuk dan eksplan dam berwarna coklat, bahkan masih ada bakteri yang tumbuh pada bagian eksplan yang bersentuhan dengan media. Metoda yang digunakan untuk transformasitunas
majemuk clan daun diduga belum sesuai, perlu dilakukan percobaan selanjutnya dengan metoda transformasi yang lebih bervariasi.Regenerasi tanaman Acacia
mangium
Willd yang telah ditransformasiKeberhasilan penelitian transformasi sangat ditentukan oleh metoda regenerasi dan transformasi yang digunakan. Setiap tanaman memiliki tingkat resistensi yang berbeda terhadap berbagai mekanisme ketahanan, bahkan satu
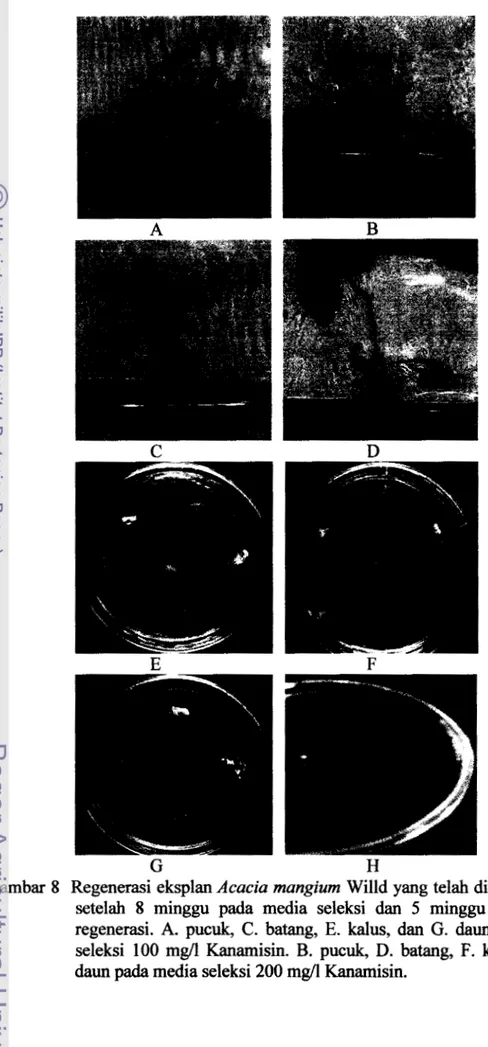
Tabel 7 Persentase regenerasi eksplan pucuk dan batang yang telah ditransformasi setelah 8 minggu pada media seleksi dan 5 minggu pada media regenerasi
Jenis eksplan - Jumlah eksplan yang telah - - Persentase regenerasi eksplan ditr&sformasi yang telah ditransformasi
Pucuk 3 6 2/36 (53%)
batang 24 2/24 (8,3%)
Tabel 7 menunjukkan bahwa persentase regenerasi eksplan batang yang telah ditransformasi lebih tinggi dibandingkan dengan eksplan pucuk yang telah ditransformasi yaitu sebesar 8,3% dibanding 53%. Komposisi media seleksi dapat juga digunakan sebagai media regenerasi selanjutnya. Pada penelitian
ini,
komposisi media seleksi dan media regenerasi adalah sama yaitu MS
+
0,25 mg/l IAA+
1 mg/lTDZ
+
225 mg/l Carbenicillin, tetapi konsentrasi Kanamisin berbeda, yaitu pada media seleksi sebesar 400 mg/l sedangkan pada media regenerasi sebesar 100 mg/l. Kanarnisin masih tetap digunakan pada media regenerasi untuk menghindari tumbuhnya bakteri pada media regenerasi. Setelah rninggu ke 8 pada media seleksi, eksplan pucuk dan batang yang telah ditnmsforrnasi dipindahkan pada media regenerasi yang tidak mengandung 1 mg/l TDZ, untuk merangsang pertumbuhan akar.Garnbar 6 Eksplan yang telah ditransformasi pada media regenerasi
(A. eksplan pucuk, B. eksplan yang telah ditransforrnasi pada media MS +
0,25 mg/l IAA
+ 225
m&/l Carbenicillin+ 100
mg/l Kanamisin. C. kalus, D.tunas majemuk, dan E. daun, pada media MS