BAB II
TINJAUAN PUSTAKA
2.1 Epidemi HIV/AIDS
Infeksi Human Immunodeficiency Virus (HIV) dan Acquired Immune Deficiency Syndrome (AIDS) telah menjadi pandemi di tingkat global dengan berbagai dampak yang merugikan, baik dampak kesehatan, sosial ekonomi maupun politik. Menurut laporan United Nations Programme on HIV/AIDS (UNAIDS) pada akhir tahun 2009, jumlah penduduk yang hidup dengan HIV/AIDS hingga Desember 2008 diperkirakan 33,4 juta (31,1 juta – 35,8 juta). Pasien dewasa berjumlah 31,3 juta (29,2 juta – 33,7 juta) dan 15,7 juta adalah wanita. Sedangkan anakanak diperkirakan 2,1 juta (1,2 juta -2,9 juta) hidup dengan HIV. Kasus terbanyak ditemukan di Afrika Sub-Sahara yaitu 22,4 juta sedangkan di Asia Selatan dan Tenggara didapatkan 3,8 juta orang (Anonim, 2009a). Secara kumulatif jumlah kasus AIDS di Indonesia menurut Departemen Kesehatan RI, hingga 30 September 2009 mencapai 18.442 kasus, sedangkan kasus kumulatif infeksi HIV hingga Juni 2009 mencapai 28.260 orang di Indonesia. Berdasarkan provinsi yang melaporkan, kasus AIDS lebih banyak di Jawa Barat, Jawa Timur, DKI Jakarta, Papua, Bali, Kalimantan Barat, Jawa Tengah, Sumatera Utara, Riau dan Kepulauan Riau (Anonim, 2009b). Di Bali, jumlah kasus AIDS sampai September 2009 ditemukan 1.508 kasus AIDS dan kumulatif infeksi HIV sebanyak 3.047 orang (Anonim, 2009c).
Berdasarkan laporan UNAIDS (2009), perbaikan akses pengobatan ARV yang pada tahun 2003 sebanyak 7% menjadi 42% pada tahun 2008. Hal ini menyebabkan terjadinya penurunan jumlah kematian odha di dunia tetapi angka prevalensi penderita HIV menjadi meningkat (Anonim, 2009a). Di Indonesia, antiretrovirus mulai digunakan dengan subsidi pemerintah pada tahun 2004. Angka kematian odha menurun dari 46% pada tahun 2006 menjadi 17% pada tahun 2008 (Anonim, 2009b).
2.2 HIV sebagai penyebab AIDS
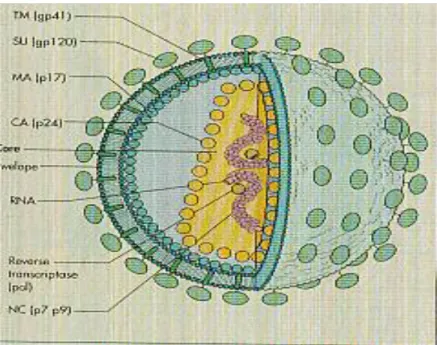
Virus HIV-1 pertama kali diisolasi dari penderita AIDS oleh Luc Montagnier dan Barre-Sinoussi di Pasteur Institute Paris pada tahun 1983 yang kemudian secara cepat dikonfirmasi oleh Robert Gallo (Collier dan Oxford, 2000). Virus HIV termasuk dalam famili Retroviridae, genus Lentivirus. Partikel virus berbentuk spheris, berselubung (envelope), dengan diameter 100 nm. Pada lapisan selubung luar lemak terdapat 72 penetrasi tonjolan glikoprotein yang disebut dengan protein envelope (env). Polipeptida env terdiri dari 2 sub unit yaitu tonjolan glikoprotein luar (gp 120) dan bagian transmembran (gp 41) yang menghubungkan tonjolan keselubung lemak. Lapisan dalam selubung lemak virus tersusun dari protein matrix (p17). Selubung lemak HIV menutupi icosahedral shell protein (p24) yang didalamnya terdapat protein inti (P7 dan P9) yang mengandung 2 untai single strand RNA (ssRNA) (Murray, et al. 2002).
Gambar 2.1 : Struktur HIV (dikutip dari Murray et al, 2002)
Pengaturan genom ssRNA yang berukuran 10 kb pada HIV-1, cukup kompleks. HIV tidak memiliki gen onc yang bersifat onkogenik seperti virus lain dalam famili yang sama. Tetapi HIV memiliki gambaran gen unik seperti gen rev yang meningkatkan replikasi virus, gen tat yang mengatur transaktivasi dan gen vif yang menentukan infektivitas virus. Gen pol menyandi sedikitnya tiga jenis protein. Protein terbesar yang disandi oleh gen pol adalah reverse transcriptase yaitu enzim yang mampu mengubah RNA menjadi DNA. Protein yang lain adalah integrase yaitu enzim yang berfungsi untuk menggabungkan gen proviral HIV kedalam DNA sel inang. Protease adalah protein ketiga yang disandi oleh gen pol yang berfungsi untuk memotong protein yang terbentuk sesaat setelah virus dilepas dari inti sel inang. Ketiga enzim ini merupakan target kerja dari ARV (Murray, et al. 2002; Freed dan Martin, 2007)
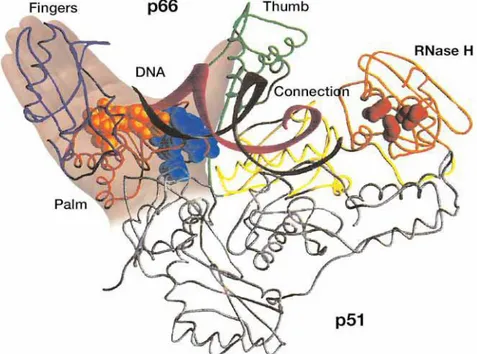
Reverse transcripase (RT) merupakan enzim multifungsi. Enzim ini mengkatalisis sedikitnya tiga reaksi, yaitu: reaksi RNA-dependent DNA polimerase, DNA-dependent DNA polymerase dan ribonuclease H (RNase H). Reverse transcriptase dapat berikatan secara spesifik dengan tRNA yaitu tRNAlys3 yang digunakan sebagai primer untuk reaksi RNA-dependent DNA polymerase. Protein RT tersusun dari 560 residu asam amino dengan berat molekul 66 kDa (p66). Protein RT rekombinan HIV-1 dapat membentuk suatu homodimer (p66/p66) yang selanjutnya oleh protease akan dipecah pada satu subunit diantara residu 440 dan 441 (Tyr-Phe). Akibat pemecahan oleh protease, mengakibatkan pengeluaran bagian C-terminal domain RNase H (p51) sehingga membentuk suatu heterodimer (p66/p51) (Stammers dan Ren, 2006).
Struktur kristal RT HIV-1 menggambarkan arsitektur dasar dari heterodimer p66/p51. Bagian N-terminal dari subunit p66 disusun dalam suatu struktur yang analog dengan gambaran seperti tangan kanan terbuka yang mengandung 3 domain seperti jari (fingers), telapak tangan (palm) dan ibu jari (thumb). Domain penghubung mengikuti domain ibu jari dan berakhir pada domain C-terminal RNase H(Stammers dan Ren, 2006).
Gambar 2.2 : Struktur kristal RT HIV (dikutip dari Stammers dan Ren, 2006)
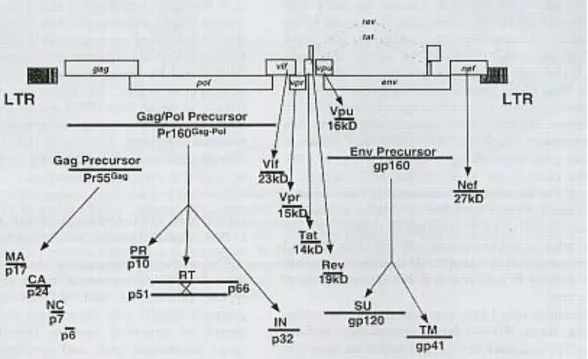
Terdapat tiga produk translasi utama pada HIV yang bereplikasi secara utuh. Ketiga produk translasi ini akan menyandi protein struktural. Pada awalnya protein ini disintesa sebagai prekusor poliprotein yang kemudian diproses oleh protease virus atau sel inang menjadi protein matur. Prekusor Gag 55-kd (Pr55Gag) akan dipecah menjadi matrik (MA), kapsid (CA), nukleokapsid (NC) dan protein p6. Autokatalisis dari poliprotein Gag-Pol 160-kd (Pr 160Gag-Pol) akan menjadi protein protease (PR), reverse transcriptase heterodimer (RT) dan integrase (IN). Digesti proteolitik oleh enzim seluler akan merubah prekusor env gp 160 terglikosilasi menjadi surface gp 120 (SU) dan transmembrane gp 41 (TM). Sedangkan 6 protein HIV lainnya (Vif, Vpr, Tat, Rev, Vpu dan Nef) merupakan produk utama dari spliced mRNA (Freed dan Martin, 2007).
Gambar 2.3: Skema protein HIV, lokasi gen HIV dan ukuran produk translasi utama (dikutip dari Freed dan Martin, 2007)
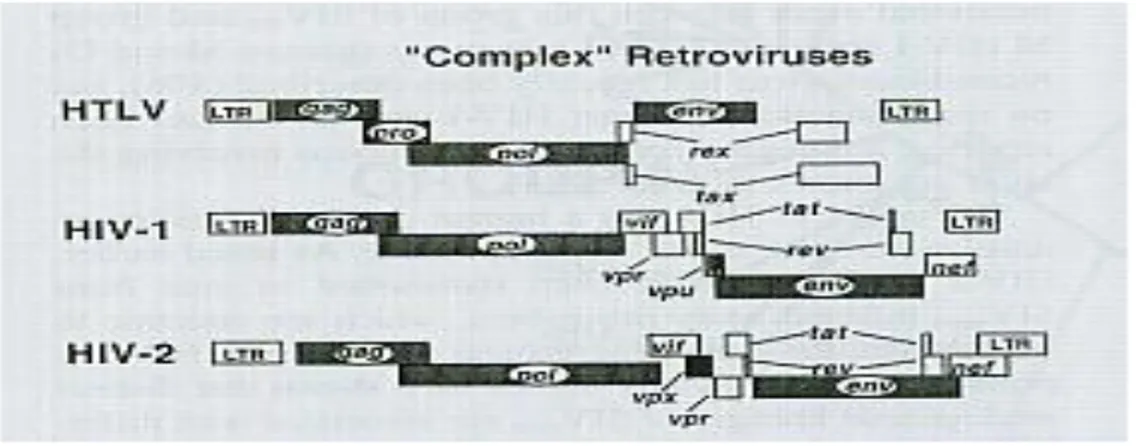
Didalam famili Retroviridae, genus Lentivirus, terdapat dua tipe penyebab AIDS, yaitu HIV-1 dan HIV-2. Pada umumnya sebagai penyebab AIDS terbanyak adalah HIV-1. Kedua jenis virus merupakan virus yang berbeda. Struktur genetiknya hampir sama, dimana HIV-1 mempunyai gen vpu tetapi tidak memiliki gen vpx, sedangkan sebaliknya HIV-2 mempunyai vpx tetapi tidak mempunyai vpu. Perbedaan struktur gen ini diperkirakan memiliki peranan dalam menentukan perbedaan patogenitas dan perjalanan penyakit kedua tipe virus ini (Freed dan Martin, 2007). Selain perbedaan genom, saat ini telah diketahui adanya perbedaan protein struktural pada HIV-1 dan HIV-2. Pada HIV-2, gen env akan menghasilkan protein gp125, gp 105
dan gp 36 yang analog dengan produk gen env pada HIV-1 yaitu gp 160, gp 120 dan gp 41 (Griffith, et al.2007).
Gambar 2.3 : Perbedaan susunan gen HIV-1 dan HIV-2 (dikutip dari Freed dan Martin. 2007)
2.3 Siklus replikasi HIV
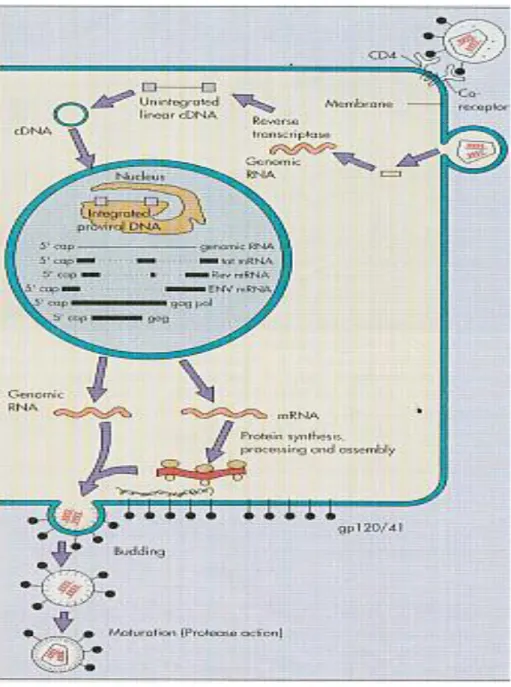
Siklus replikasi HIV-1 secara in vivo diperkirakan membutuhkan waktu selama 24 jam. Proses replikasi dimulai dengan melekatnya gp 120 pada reseptor CD4 sel inang. Perlekatan ini menyebabkan perubahan yang memungkinkan virus berinteraksi dengan tempat koreseptor sel lainnya, yaitu CXCR4 dan CCR5. Setelah virus masuk kedalam sel, maka RNA virus akan dilepaskan. Partikel HIV mengandung reverse transcriptase (RT) yaitu enzim yang berperanan penting dalam proses replikasi. RT memiliki tiga fungsi yang berbeda: RNA-dependent DNA polymerase, yang berperan sebagai sintesa cDNA; RNase H, yang menghancurkan RNA dari kompleks cDNA-RNA dan DNA-dependent DNA polymerase, yang menyalin untai cDNA. Integrase adalah enzim yang akan memasukkan cDNA kedalam genom sel inang. Selanjutnya
cDNA akan berfungsi sebagai cetakan bagi RNA virus. Pengaktifan transkripsi HIV dan ekspresi gen dipercepat oleh faktor transkripsi seluler dan oleh protein pengatur virus, seperti Tat, Rev, Nef dan Vrp. Pengaturan gen tat dan rev sangat mempengaruhi kecepatan replikasi virus. Protein Tat akan meningkatkan kecepatan replikasi sedangkan protein Rev akan memperantarai pengeluaran unspliced atau partially spliced RNA yang menyandi protein struktural virus. Pada akhir siklus, virion akan disusun dan menonjol keluar melalui membran plasma. RT pada HIV memiliki proof reading yang jelek (infidelity), sehingga banyak kesalahan saat proses replikasi terjadi. Hal ini menyebabkan seseorang yang terinfeksi HIV, dalam tubuhnya memiliki keragaman virus HIV yang dikenal dengan istilah quasispecies (Griffith, et al. 2007; Freed dan Martin, 2007).
2.4 Penyebaran HIV
Virus HIV-1 dan HIV-2 memiliki pola penyebaran yang sama. Pola penyebaran terbanyak dari HIV adalah penyebaran melalui hubungan seksual melalui kontak langsung mukosa genital dengan cairan tubuh terinfeksi seperti darah, semen sperma dan sekresi vagina. Infeksi juga dapat terjadi melalui inokulasi darah terinfeksi lewat transfusi darah, transplantasi jaringan atau penggunaan ulang jarum terkontaminasi. Sebagian besar penyebaran HIV dari ibu kepada anaknya terjadi di negara-negara berkembang. Adanya RNA HIV-1 ibu di darah dan cairan genital telah terbukti erat hubungannya dengan penyebaran dari ibu kepada anak. Penyebaran HIV dapat terjadi saat dalam kandungan, saat proses melahirkan atau saat proses menyusui (Griffith, et al.2007; Kuritzkes dan Walker, 2007).
2.5 Subtipe HIV-1
Pandemi HIV-1 disebabkan oleh penyebaran lentivirus antar spesies mahluk hidup. Cikal bakal virus penyebab infeksi adalah virus HIV-1 group M (main). Selama penyebarannya diantara manusia, virus HIV-1 group M berkembang pesat dengan keragaman genetik yang sangat bervariasi dan sebagian besar terpisah menjadi subtipe murni dan beberapa bentuk gabungan atau rekombinan. Virus HIV memiliki beberapa mekanisme intrinsik yang menyebabkan cepatnya evolusi virus. Enzim RT memiliki kekurangan proofreading yang bertanggung jawab atas keakuratan kode RNA serta angka mutasi 3,4 x 10-5 mutasi pasang basa disetiap siklus replikasi. Diperkirakan panjang genom HIV adalah 104 bp, maka akan terjadi jutaan variasi yang dihasilkan pada seseorang terinfeksi virus HIV setiap harinya (Taylor et all, 2008)
Kelompok M adalah kelompok virus HIVyang bersirkulasi paling luas. Kelompok ini dibagi menjadi subtipe yang ditandai dengan huruf serta sub-subtipe yang ditandai dengan angka. Saat ini dikenal subtipe dan sub-subtipe A1, A2, A3, A4, B, C, D, F1, F2, G, H, J dan K. Untuk memudahkan pembagian kelompok subtipe maka HIV secara umum akan dikelompokkan menjadi kelompok subtipe B dan subtipe non B. Yang termasuk dalam kelompok subtipe non B adalah seluruh subtipe atau bentuk rekombinan selain subtipe B. Variasi genetik diantara satu jenis subtipe berkisar antara 15-20%, sedangkan variasi diantara subtipe sebesar 25-35% (Taylor et al, 2008). Seiring dengan berkembangnya teknologi skuensing genom HIV, maka telah diketahui adanya bentuk gabungan bersirkulasi (Circulating recombinant form = CRF) dan bentuk gabungan unik (Unique recombinant form = URF). Satu jenis rekombinan terbentuk bila 2 jenis subtipe menginfeksi seseorang dalam waktu bersamaan dan rekombinan terbentuk disebarkan kepada penderita berikutnya. Progeni rekombinan akan ditentukan sebagai bentuk CRF baru bila bentuk rekombinan tersebut ditemukan pada tiga atau lebih penderita yang tidak memiliki hubungan langsung secara epidemiologis. Beberapa bentuk rekombinan yang telah ditentukan antara lain : CRF01_AE dominan ditemukan di Asia Tenggara, CRF03_AB banyak ditemukan pada negara bekas Uni Soviet, CRF14_BG ditemukan di Spanyol dan Portugal sedangkan CRF terbaru adalah CRF35_AD di Afganistan dan Iran (Taylor et al, 2008; Kantor et al, 2005)
Bila dilihat susunan RT dan protease diantara masing-masing subtipe, maka akan terdapat perbedaan nukleotida sebesar 10-12% dan perbedaan asam amino sebesar
5-6% (Kantor et al, 2005). Perbedaan diantara subtipe ini dapat mempengaruhi variasi mutasi yang terjadi karena tekanan obat secara selektif. Perbedaan susunan asam amino diantara subtipe akan mempengaruhi ketahanan virus terhadap ARV. Beberapa mutasi pada subtipe B akan mengkoding kekebalan terhadap ARV tetapi mutasi yang terjadi pada asam amino sama pada subtipe non B dapat berfungsi sebagai variasi genetik atau polimorfisme tanpa mengkoding kekebalan terhadap ARV. Polimorfisme ditentukan bila mutasi yang terjadi lebih dari 1% pada skuensing dari virus penderita yang belum diterapi. Polimorfisme spesifik subtipe adalah mutasi yang secara signifikan lebih banyak ditemukan pada setiap subtipe non B daripada subtipe B pada penderita yang tidak diterapi. Mutasi berhubungan dengan terapi yang subtipe spesifik terjadi bila mutasi signifikan lebih banyak pada isolat HIV-1 dari penderita yang diterapi daripada penderita yang tidak diterapi yang terinfeksi oleh subtipe yang sama.
Kantor et al (2005) mengembangkan suatu model respon binomial menggunakan subtipe dan riwayat pengobatan sebagai variabel untuk memprediksi apakah suatu posisi asam amino merupakan posisi sesuai virus standard (wild-tipe) atau merupakan mutasi. Model ini mengidentifikasi 3 jenis posisi, yaitu: 1) posisi pada skuensing dari penderita tidak diterapi lebih sering mutasi pada subtipe non B daripada subtipe B disebut dengan subtype-specific polymorphism. 2) posisi pada skuensing pada subtipe sama lebih sering bermutasi pada penderita yang diobati daripada yang tidak, disebut dengan subtype-specific treatment-related positions. 3) posisi dimana efek pengobatan berbeda secara signifikan diantara subtipe B dan non B, disebut dengan subtype-treatment interaction.
2.6 Gambaran klinis HIV/AIDS
Gejala klinis HIV/AIDS tergantung dari fase infeksi yang terjadi. Pada awal infeksi, 2 atau 4 minggu setelah terinfeksi HIV, penderita akan mengalami gejala seperti flu, antara lain : demam, sakit kepala, sakit tenggorokan, pembesaran kelenjar getah bening dan kemerahan pada kulit. Pada infeksi lanjut, gejala timbul setelah 8 atau 9 tahun terinfeksi. Gejala pada fase lanjut antara lain pembesaran kelenjar getah bening, diare, penurunan berat badan, demam, batuk dan sesak napas. Fase terakhir dari infeksi yang terjadi 10 tahun sejak terinfeksi, menunjukkan gambaran klinis yang berat yang sering disebut dengan AIDS. Gejala AIDS antara lain: berkeringat malam, demam tinggi selama beberapa minggu, batuk kering, sesak napas, diare kronis, adanya lesi keputihan menetap pada lidah atau mulut, sakit kepala, gangguan penglihatan, penurunan berat badan (Anonim, 2010)
Centers for Disease Control and Prevention (CDC) melakukan revisi klasifikasi derajat penyakit infeksi HIV pada tahun 1993. Klasifikasi dibuat berdasarkan gejala klinik dan jumlah sel limfosit CD4. Berdasarkan gejala klinis, dibagi atas katagori A, B dan C. Kategori A adalah asimtomatik, persisten generalized lymphadenopathia (PGL) dan acut symptomatic primary infection. Kategori B adalah adanya gejala klinik yang tidak termasuk kategori C, namun berkaitan dengan infeksi HIV atau akibat defisiensi imun. Kategori C: merupakan penyakit petunjuk AIDS. Berdasarkan jumlah CD4, terdapat kategori 1, 2 dan 3. Kategori 1 bila CD4 ≥ 500 /µl darah. Kategori 2 bila CD4 berjumlah 200 – 499 /µl darah sedangkan kategori 3 bila CD4 < 200 /µl darah. Berdasarkan kombinasi kedua kategori tersebut, maka penderita infeksi HIV akan
masuk kedalam derajat 1 adalah A1, B1 dan A2; derajat 2 adalah B2,, A3 dan B3. Sedangkan derajat 3 adalah C1, C2 dan C3 (Anonim, 1992)
2.7 Terapi AIDS
Menghilangkan infeksi HIV tidak dapat dicapai dengan menggunakan ARV yang tersedia saat ini. Hal ini terjadi karena virus telah berada di dalam CD4 sel T sejak fase awal infeksi akut dan dapat menetap dalam jangka panjang. Menurut Panel on ARV Guideline (2009), tujuan pemberian terapi ARV pada penderita infeksi HIV/ AIDS adalah: menekan sebanyak dan selama mungkin viral load HIV dalam plasma; menurunkan kematian yang berhubungan dengan HIV dan memperpanjang harapan hidup penderita; meningkatkan kualitas hidup penderita; meningkatkan fungsi imunologis penderita dan mencegah penyebaran HIV (Anonim, 2009e)
Program penggunaan ARV pada penderita AIDS memberikan manfaat yang besar, antara lain: menyebabkan harapan hidup penderita AIDS meningkat dari 4 bulan sejak terdeteksi menjadi 50 bulan (Kilsztajn, 2007). Kombinasi ARV dengan kotrimoksasol berhasil menurunkan kematian di Uganda (Mermin, 2008) dan di Zambia (Walker, 2007). Di Botswana, penderita yang mendapat ARV hidup 5 tahun lebih lama daripada yang tidak mendapat ARV (Bussmann, 2008).
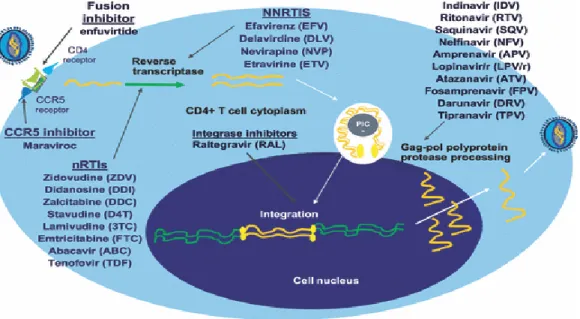
Antiretroviral terdiri dari 5 golongan obat berbeda yang digunakan secara kombinasi. Metode pengobatan kombinasi ini disebut dengan highly active antiretroviral therapy (HAART). Secara umum HAART terdiri dari 3 jenis golongan ARV. Kombinasi yang biasa digunakan adalah dua jenis obat golongan nucleoside
analogues atau nucleoside reverse-transcriptase inhibitor (NRTI) dengan satu golongan protease inhibitor (PI) atau golongan non nucleoside reverse-transcriptase inhibitor (NNRTI). Penggunaan secara kombinasi bertujuan untuk mencegah timbulnya virus resisten, tetapi beberapa kombinasi memberi efek antagonis dan tidak ada bukti kombinasi yang bersinergis baik secara in vitro (Clavel dan Hance, 2004).
Gambar 2.5 : Golongan ARV dan cara kerja masing-masing obat (dikutip dari Kozal,2009)
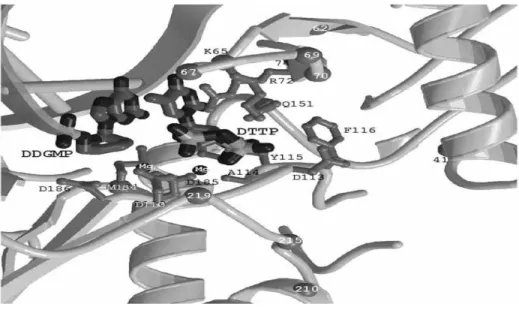
Nucleotide akan berikatan pada bagian kantong katalitik kompleks RT. Didalam stuktur RT terdapat struktur khusus tempat terjadinya katalitik kompleks enzim dengan ikatan dTTP dan ikatan kovalen, dsDNA, yang disebut dengan celah aktif atau kantong (pocket). Basa guanosine termodifikasi mengandung grup sulfhydryl akan bergabung kedalam untaian template, menyebabkan ikatan pada cysteine diposisi 258 pada domain ibu jari. Komplek RT dengan ikatan kovalen dsDNA memiliki suatu dideoxyguanylate terminating untai primer. Basa berikutnya yang tersedia didalam template adalah
adenine. Menyebabkan site dNTP akan diisi oleh dTTP, tetapi tanpa reaksi selanjutnya. Pada tempat ini terjadi pendekatan domain ibu jari dan jari-jari, mengakibatkan bagian tengah celah semakin mengkerut dan protein terdekat kontak dengan site dNTP. Template berada pada posisi tergantung diluar bagian tengah celah. dTTP terdapat pada akhir primer dengan basa thymin yang tersangkut, sedangkan bagian untaian dari Lys65 dan Arg72 akan bereaksi dengan bagian permukaan luar. Penutupan domain jari-jari menyebabkan bagian cincin bereaksi dengan grup trifosfat dTTP. Interaksi tambahan dapat terjadi melalui residu cincin utama grup –NH, 113 dan 114, dan melalui dua ion magnesium, satu akan berikatan dengan dua dari katalitik kunci aspartat, 110 dan 185. 3’hidroxyl dari dTTP akan menonjol kedalam suatu kantong yang mengandung bagian cincin dari Asp113, Tyr115, Phe116 dan Gln151. Kantong ini memberi ruang untuk grup 3’-azido dari zidovudine (Stammers dan Ren, 2006).
Gambar 2.6 : Struktur kristal katalitik kompleks RT (dikutip dari Stammers dan Ren, 2006)
Nucleotide dan nucleoside analogues menghentikan sintesa DNA virus pada proses transkripsi balik. Setelah difosforilasi oleh enzim kinase seluler, komponen ini akan berikatan dengan cincin nascent DNA virus melalui proses transkripsi balik. Karena obat ini tidak memiliki group hidroksi 3’ maka tidak ada nukleotida yang dapat ditambahkan pada obat ini sehingga proses berhenti (Clavel dan Hance, 2004). Zidovudin trifosfat bekerja sebagai suatu penghambat kompetitif dari substrat thymidine trifosfat (dTTP). Obat ini dapat bergabung sendiri kedalam untai primer dan berfungsi sebagai cincin penghenti karena grup azido tidak memiliki posisi 3’ ribose (Stammers dan Ren, 2006)
Yang termasuk dalam kelompok nucleoside analogues lainnya adalah: lamivudine (Epivir), didanosine (Videx), Stavudine (Zerit) dan abacavir (Ziagen)dan
emtricitabine (Emtriva). Sedangkan yang termasuk dalam nucleotide analogues hanya tenofovir (Viread) (Anonim, 2010)
Tempat ikatan NNRTI berada diantara dua three-stranded β-sheets dan sebagian besar berada pada subunit p66 kecuali bagian cincin Glu138 terbentuk dari subunit p51. Tempat ini berupa kantong besar pada molekul RT. Permukaan dalam kantong secara natural bersifat hidrofobik. Ikatan dengan NNRTI melibatkan residu asam amino Leu100, Try181 dan Tyr188. Bagian atas kantong dibentuk oleh Try181, Try188 dan variant Trp229. Residu asam amino penting lainnya yang kontak dengan NNRTI antara lain: Val106, Val179, Phe227 dan Try318. Residu polar yang juga merupakan bagian dari kantong antara lain Lys101, Lys103 dan Glu138 (p51) (Stammers dan Ren, 2006).
Antiretroviral kelompok NNRTI merupakan molekul kecil yang memiliki afinitas kuat terhadap kantong hidrofobik yang berlokasi pada domain katalitik reverse-transcriptase. Inhibitor akan berefek terhadap kelenturan enzim yang akan menghambat kemampuan enzim untuk mensintesa DNA (Clavel dan Hance, 2004) Terdapat 4 macam NNRTI yang sudah mendapat persetujuan untuk digunakan, yaitu: nevirapine (Viramune), delavirdine (Rescriptor), efavirenz (Sustiva) dan etravirine (Intelence) (Anonim, 2010)
Protease HIV memecah prekusor poliprotein besar pada tempat yang spesifik, melepas protein struktural dan enzim yang dibutuhkan untuk menyusun partikel virus. Bila protease tidak ada maka akan terbentuk partikel virus tetapi imatur dan tidak infeksius (Clavel dan Hance, 2004). Yang termasuk dalam kelompok protease inhibitor (PI) antara lain: saquinavir (Invirase), ritonavir (Norvir), indinavir (Crixivan), nelfinavir (Viracept), amprenavir (Agenerase), lopinavir/ritonavir (Kaletra), natazanavir (Reyataz) dan tipranavir (Aptivus). Danuravir (Prezista) cenderung diberikan pada penderita yang tidak berespon terhadap pengobatan dengan golongan obat lain. Kelompok PI biasanya diberikan secara kombinasi dengan obat lain untuk mencegah terjadinya resistensi (Anonim, 2010)
Salah satu masalah yang sedang dihadapi dalam pengobatan AIDS adalah timbulnya HIVDR. Di seluruh dunia, sebagian besar penduduk yang menerima pengobatan HIV akan resisten minimal dengan satu jenis obat dan banyak yang tidak berespon dengan kombinasi tiga jenis obat. Saat ini terdapat satu kelompok obat yang dapat mengatasi masalah HIVDR yaitu kelompok fusion inhibitor. Enfurvirtide
(Fuzeon) merupakan satu-satunya obat yang termasuk dalam kelompok fusion inhibitor yang bekerja dengan cara memhambat fusi virus dengan sel inang yang sehat sehingga proses replikasi tidak terjadi. Fuzeon biasa digunakan dalam bentuk kombinasi dengan golongan obat ARV lainnya (Anonim, 2010)
Integrase inhibitor dikembangkan juga untuk mengobati HIVDR. Yang termasuk dalam kelompok ini adalah raltegravir (Isentress). Obat ini akan menghambat kerja enzim integrase sehingga DNA virus tidak dapat menyatu dengan DNA sel inang. Efek samping yang sering dilaporkan dalam penggunaan obat ini adalah diare, mual, sakit kepala dan demam (Anonim, 2010)
2.8 HIV drug resistance (HIVDR)
Terdapat dua hal penting mendasari timbulnya HIVDR. Pertama, infeksi HIV secara khas ditandai dengan siklus virus yang cepat serta produksi virus yang tinggi. Pada penderita yang tidak diobati, total sel produktif yang terinfeksi HIV pada jaringan limfoid berkisar antara 107 hingga 108 sel. Pada fase kronis infeksi HIV, jumlah ini relatif stabil, yang menunjukkan keseimbangan antara sel yang baru terinfeksi dengan sel yang mati. Disebabkan masa hidup sel terinfeksi cukup singkat maka untuk mempertahankan keseimbangan ini diperlukan kecepatan tinggi dari virus untuk membentuk virus baru. Kedua, populasi virus pada penderita HIV sangat heterogen. Reaksi balik virus dari RNA menjadi DNA cenderung sering salah menyebabkan kesalahan rata-rata satu mutasi pada setiap transkripsi gen virus. Sebagian besar kesalahan adalah salah mengganti basa (base substitution) tetapi kadang-kadang juga
terjadi duplikasi, insersi dan rekombinasi. Tingginya kecepatan replikasi disertai dengan tingginya angka mutasi pada setiap siklus infeksi menyebabkan penderita memiliki suatu squasi species (Clavel dan Hance, 2004)
2.8.1 Mekanisme resisten 2.8.1.1 Resisten terhadap NRTI
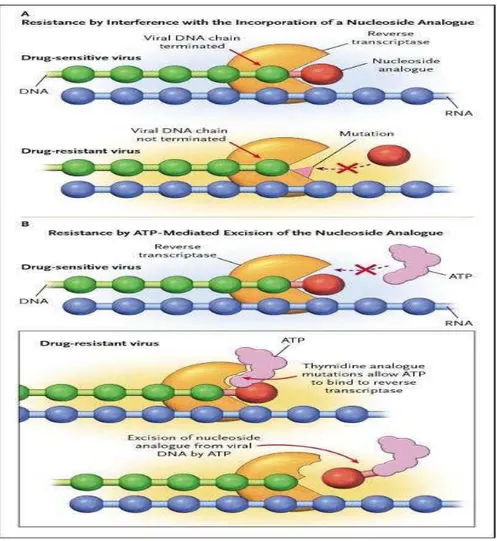
Terdapat 2 mekanisme resisten yaitu ; kerusakan gabungan analog dengan DNA atau menghilangkan analog dari cincin DNA yang dihentikan lebih awal (the prematurely terminated DNA chain) (gambar 2.9a). Pada mekanisme resisten karena kerusakan gabungan analog, dapat terjadi pada mutasi M184V, Q151M mutasi kompleks dan mutasi K65R. Mutasi M184V merubah methionin menjadi valin pada posisi 184. Mutasi ini menyebabkan resisten terhadap l amivudine dengan sifat resisten tinggi (Boucher, et al. 1993; Fumero dan Podzamczer, 2003). Mutasi Q151M kompleks berhubungan dengan kegagalan terapi terhadap stavudine dan didanosin. Jalur ini selalu didahului oleh substitusi Q151M, yang diikuti oleh akumulasi mutasi sekunder bertahap yang akan memperkuat resistensi dan meningkatkan aktivitas enzim. Kompleks Q151M relatif jarang pada 1 (<5%) lebih sering terjadi pada tipe HIV-2. Sedangkan mutasi K65R sering pada terapi dengan tenofovir dan abacavir (Fumero dan Podzamczer, 2003). Pola mutasi yang ditemukan pada HIV-1 subtipe CRF01_AE, terhadap kelompok NRTI antara lain: M41L, A62V, K70R, L74I, V75M, M184I/V, T215Y dan K219E/Q (gambar 2.8) (Chan dan Kantor, 2009).
Gambar 2.8 : Pola mutasi berhubungan dengan resisten NRTI (dikutip dari Johnson, 2004)
Pada mekanisme resisten karena menghilangnya analog dari cincin DNA terminal disebut dengan thymidine analog mutasi. Sering ditemukan setelah terapi dengan thymin analog seperti zidovudine dan stavudine tetapi dapat juga merangsang resisten terhadap semua golongan nucleoside dan nucleoside analogues. Resisten timbul melalui fostering ATP atau penghilangan pyrophosphat-mediated nucleoside analog dari ujung 3’ untai DNA terminal. Phyrophosphat dan ATP secara normal terdapat dalam limfosit, tidak berperanan dalam reaksi DNA polimerasi tetapi mutasi yang terjadi menyebabkan ATP dapat masuk kedalam menggantikan analog. Pada posisi ini ATP atau pirofosfat dapat menyerang ikatan fosfodiesterase yang mengikat analog ke DNA (gambar 2.9b). Proses ini dikenal dengan istilah ‘primer rescue (Clavel dan Hance, 2004)
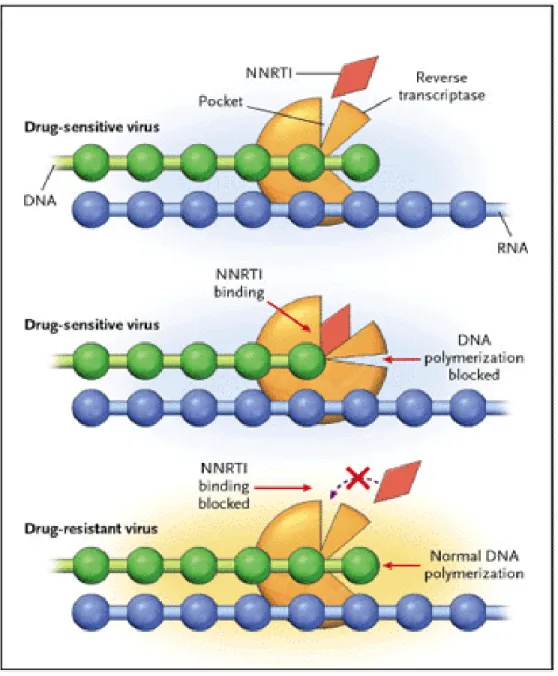
Gambar 2.9. Skema mekanisme resistensi NRTI (dikutip dari Clavel dan Hance, 2004) 2.8.1.2 Resisten terhadap NNRTI
Mutasi yang timbul sebagian besar bersifat drug-dependent. Resisten terhadap nevirapine berhubungan dengan mutasi Y181C, tetapi mutasi yang lain seperti Y188C, G190A dan V106A juga dapat terjadi. Mutasi K103N sering berhubungan dengan resistan terhadap efavirens. Terdapat 2 kelompok pola mutasi yang dominan yaitu: kelompok K103N, V106M, Y188Y dan kelompok L100I, V106A, Y181C/I, G190S/A, M230L (Clavel dan Hance, 2004; Hirsch, et al. 2008) (gambar 2.11). Pada subtipe
CRF01_AE, hanya 4 mutasi yang ditemukan terhadap NNRTI, yaitu: K103N, Y181C, Y188L dan G190E (Chan dan Kantor, 2009)
Gambar 2.10. Skema mekanisme resisten terhadap NNRTI (dikutip dari Clavel dan Hance, 2004)
Gambar 2.11 : Pola mutasi berhubungan dengan resisten NNRTI (dikutip dari Johnson, 2004)
2.8.1.3 Resisten terhadap PI
Resisten terhadap protease inhibitor merupakan akibat dari pergantian asam amino yang terjadi didalam domain ikatan substrat enzim atau pada tempat distant. Mutasi yang menyandi perubahan tempat aktif protease menghambat ikatan PI disebut
dengan major mutasi. Mutasi D30N akan menyebabkan resisten terhadap nelfinavir. Mutasi V82A/F/T/S, I84V dan L90M menyebabkan mutasi terhadap sebagian besar kelompok PI(Clavel dan Hance, 2004; Hirsch, et al. 2008). Pada subtipe CRF01_AE, terdapat 4 mutasi yang ditemukan terhadap PI, yaitu: D30N, L33F, M46I, F53L, G73S, N88D dan L90M (Chan dan Kantor, 2009)
2.8.2 Jenis HIVDR
Terdapat 2 jenis HIVDR yaitu primary/transmitted yaitu HIVDR resisten terhadap satu atau lebih ARV yang ditemukan pada penderita yang belum pernah mendapat ARV (naïve patient). Pada umumnya transmitted HIVDR didapat pada penyebaran langsung dari penderita HIV yang telah diobati. Sedangkan acquired/secondary yaitu HIVDR yang timbul saat penderita mendapat terapi ARV (Vella dan Palmisano, 2005; Shafer, et al. 2007).
Terapi ARV akan dimulai berdasarkan penurunan jumlah CD4+ sel T. Pada umumnya penurunan jumlah CD4+ hingga level membutuhkan terapi terjadi dalam waktu 10-12 tahun sejak terinfeksi. Pada negara maju, terapi ARV telah dimulai sejak tahun 1996 tetapi pada negara berkembang seperti Indonesia, ARV baru tersedia pada tahun 2004. Oleh karena itu, para ahli menyimpulkan bahwa HIVDR yang terjadi pada negara berkembang pada umumnya disebabkan karena aquired HIVDR bukan karena transmitted HIVDR, sehingga angka prevalen transmitted HIVDR pada negara berkembang lebih rendah daripada di negara maju (Bennett, et al. 2008).
2.8.3 Epidemi HIVDR
Prevalensi HIVDR bervariasi tergantung dari populasi penelitian. Huang (2008) melaporkan hasil penelitian di 10 kota besar di Amerika Serikat pada penderita na ïve HIV yang dilakukan pada bulan Agustus 2005 hingga Januari 2007. Didapatkan prevalensi resistan ARV dari 228 penderita yang diperiksa adalah 12,1% (9,8% resisten terhadap NNRTI, 4,5% resisten terhadap NRTI dan 1,8% resisten terhadap PI). Diantara 228 penderita, 8,9%, 2,2% dan 1,0% galur HIV yang ditemukan resistan terhadap satu, dua dan tiga kelas ARV (Huang et al, 2008). Pada tahun 1997 hingga 2001, Weinstock (2004) terlebih dahulu melakukan penelitian pada 10 kota besar di AS. Dari 1311 penderita naïve HIV yang memenuhi kriteria inklusi hanya 1082 pasien yang berhasil dilakukan pemeriksaan genotyping dan didapatkan 8,3% dari subyek penelitian telah resisten terhadap RT dan PI. Sedangkan di Israel, didapatkan 14,8% penderita yang baru terdiagnosa terinfeksi HIV telah resisten terhadap ARV (Grossman, et al. 2004). Penelitian penderita naïve HIV di penjara di Massachusetts mendapatkan 16% penderita mengandung virus HIV yang telah resisten terhadap ARV (Stone et al. 2002).
Prevalensi dari Transmitted Drug Resistance (TDR) HIV-1 bervariasi tergantung dari lokasi penelitian, kelompok resiko tinggi yang diteliti dan waktu pengambilan spesimen setelah terinfeksi virus HIV-1. Di New York, dilaporkan peningkatan TDR secara tajam dari 13,2% pada periode waktu 1995-1998 menjadi 24,1% pada periode waktu 2003-2004. Kelompok peneliti di Inggris juga melaporkan tingginya TDR pada tahun 2003 yaitu: 19,2% terhadap semua obat, 12,4% terhadap NRTI, 8,1% terhadap NNRTI dan 6,6% terhadap PI (Hirsh, et al. 2008). Prevalensi
TDR pada HIV subtipe non B sulit untuk ditentukan karena kurangnya standar definisi dan klasifikasi resisten ARV pada kelompok tersebut. Prevalensi TDR HIV subtipe non B bervariasi di Asia. Penelitian di China mendapatkan angka resisten yang rendah, < 6%. Hasil ini hampir sama dengan yang dilaporkan dari Malaysia, Singapura, Kamboja, Vietnam. Sedangkan di Thailand, hasil resistensi TDR bervariasi dari 0 hingga 24%. Walaupun di Taiwan subtipe B yang dominan, tetapi angka resisten ARV cukup rendah yaitu 8%. Di India, virus HIV-1 yang dominan adalah subtipe C, dilaporkan angka resisten ARV kurang dari 5%. (Chan and Kantor, 2009)
Pada penderita yang sedang dalam pengobatan dengan ARV atau mengalami gagal terapi dengan ARV, maka angka resisten virus terhadap ARV lebih tinggi daripada angka yang ditemukan pada penderita naïve HIV. Penelitian resisten ARV diantara penderita HIV-1 di Soweto Afrika Selatan yang sedang dalam pengobatan ARV didapatkan 78% penderita viremia sedikitnya telah resisten terhadap satu jenis ARV dan 52% resisten terhadap NNRTI dan NRTI. Dari 94 penderita yang mengalami viremia pada pengobatan first-line, ditemukan 64%, 81% dan 2% telah resisten terhadap NRTI, NNRTI dan PI secara berturutan (El-Khatib et al. 2010). Hasil penelitian yang menarik dilaporkan oleh Eyer-Silva dan Morgado. Peneliti ini melakukan tes genotyping pada populasi penderita HIV yang pernah atau sedang menggunakan terapi ARV (treatment-experienced patient) pada kota kecil di Brazil. Dari 27 pasien yang diperiksa, tidak ada penderita yang menunjukkan resisten terhadap ARV. Tetapi 33% (9/27) dari pasien telah menunjukkan penurunan kepekaan terhadap ARV, 6 orang menunjukkan penurunan kepekaan terhadap RT dan 3 orang menunjukkan penurunan
kepekaan terhadap RT dan PI (Eyer-Silva dan Morgado, 2005). Di Amerika Serikat, >60% dari penderita HIV yang mendapat terapi ARV dalam kurun waktu tahun 1996-1998 memiliki viral load >500 kopi/ml. Dari kelompok tersebut 76% telah resisten terhadap sedikitnya 1 jenis ARV (Richman, et al. 2004)
Resisten terhadap ARV juga ditemukan pada anak-anak yang menderita HIV. Di Tanzania, lebih dari 50% anak-anak yang menerima pengobatan ARV dalam jangka waktu panjang menunjukkan adanya resistensi terhadap ARV yang digunakan (Bratholm, et al. 2010). Almeida (2009) di dalam Jornal de Pediatria menulis bahwa diantara 23 anak-anak yang mendapat terapi ARV di Sao Paulo Brazil, telah resisten terhadap NRTI, NNRTI dan PI masing-masing 96%, 61% dan 17% berturut-turut. Tetapi dari 24 orang anak naïve HIV, tidak ada yang resisten terhadap ARV (Almeida, 2009). Delaugerre (2007) menganalisa sekuens RT dan protease dari 119 anak-anak yang mendapat terapi ARV di Paris antara tahun 2001-2003. Didapatkan bahwa 82%, 32% dan 27% virus telah resisten terhadap satu, dua dan tiga jenis ARV secara berturut-turut (Delaugerre et al, 2007). Sedangkan Machado (2005) mendapatkan dari 37 anak yang mendapat terapi di Sao Paulo Hospital, 84%, 35% dan 14% telah resisten terhadap satu, dua dan tiga jenis ARV (Machado et al, 2005).
2.8.4 Pemeriksaan HIVDR
Pengawasan keberadaan dan perkembangan transmitted HIVDR yang sering disebut dengan transmitted drug resistance (TDR), sangat penting dilakukan. Hal ini terutama penting bagi daerah yang memiliki persediaan obat lini kedua terbatas dan
pemilihan obat lini pertama merupakan hal penting dalam pengobatan. Kelompok kerja HIV pada WHO menyarankan untuk melakukan uji resistensi pada awal terapi pada negara-negara yang telah berkembang. Sedangkan pada negara-negara sedang berkembang, uji resistensi dilakukan sebagai bagian pendekatan komprehensif pada pengobatan HIV(Anonim, 2007)
Faktor-faktor yang mempengaruhi deteksi TDR, antara lain: durasi infeksi dan sensitivitas metoda yang digunakan untuk mendeteksi. HIV secara alami memiliki tingkat replikasi dan mutasi yang tinggi. Oleh karena itu, mutasi pada TDR dapat menghilang seiring dengan lamanya proses infeksi terjadi. Saat ini, uji resistensi secara umum hanya mampu mendeteksi 15-30% mutasi yang terjadi pada populasi quasi spesies. Kesulitan deteksi TDR semakin meningkat karena HIV memiliki keragaman berdasarkan type (HIV-1 atau HIV-2), group (group M, O atau N), subtype dan recombinant forms. Group M dilaporkan sebagai penyebab terbanyak infeksi HIV di dunia yang terdiri dari beberapa macam subtype seperti A,B,C,D,F,G,H atau J dan bentuk recombinant forms lainnya. Subtipe non B diperkirakan 90% sebagai penyebab infeksi HIV, tetapi sebagian besar infeksi HIV subtipe non B terjadi pada negara-negara yang memiliki infrastuktur dan keuangan yang terbatas. Sedangkan infeksi HIV subtipe B lebih sering terjadi pada negara-negara maju. Beberapa mutasi yang berhubungan dengan resistensi obat yang terjadi pada subtipe B merupakan mutasi normal atau variasi normal pada subtipe non B. Penelitian TDR lebih banyak dilaporkan dari negara-negara maju yang pada umumnya memiliki subtipe B sehingga mutasi yang dilaporkan belum tentu sebagai mutasi resistansi obat pada subtipe non B. Oleh karena
itu dibutuhkan metoda spesifik untuk membedakan mutasi resistensi obat dengan mutasi variasi normal pada subtipe yang berbeda (Chan dan Kantor, 2009).
Saat ini disarankan untuk melakukan pemeriksaan HIVDR secara rutin terhadap seluruh penderita infeksi HIV yang baru terdiagnosis, karena transmitted HIVDR akan menetap dalam periode waktu yang tidak dapat ditentukan sejak terinfeksi (Hirsch, et al. 2003; Litte, et al. 2008) dan berkembang berlebihan bila terapi dihentikan (Barbour, et al. 2004). Sedangkan aquired HIVDR akan menghilang secara bertahap dari sirkulasi virus dalam beberapa bulan bahkan tahun bila penderita tidak mendapat ARV (Vella dan Palmisano, 2005) tetapi pendapat lain menyatakan bahwa aquired HIVDR akan tetap ada sebagai kelompok kecil dalam sirkulasi virus yang mungkin tidak terdeteksi menggunakan pemeriksaan yang tersedia (Barbour, et al. 2004). Sax (2008) melaporkan bahwa pasien yang menunjukan kegagalan secara virologi terhadap terapi yang mengandung golongan obat NNRTI karena adanya mutasi yang kemudian terapi pengganti mengalami keterlambatan perubahan jenis obat akan meningkatkan resiko kematian dan kegagalan imunologis. Tetapi hal ini tidak terjadi pada kegagalan terapi menggunakan golongan PI (Sax, 2008). Sehingga untuk mendeteksi mutasi yang terjadi, tes resistensi sangat diperlukan.
Pada negara berkembang, kegagalan terapi pada penderita HIV didasari oleh pengamatan secara klinis dan pemeriksaan immunologi saja (Anonim. 2007). Pemeriksaan viral load dan tes resistensi yang dapat mendeteksi kegagalan terapi lebih awal sulit dilakukan karena biaya pemeriksaan yang tinggi dan tidak semua negara memiliki fasilitas pemeriksaan tersebut. Untuk fasilitas penunjang pemeriksaan
resistensi menggunakan tes genotipik dibutuhkan biaya minimal Rp 9.000.000.000,- per lab (Shepherd, et al, 2008). Keadaan ini menyebabkan kesulitan mendeteksi kegagalan terapi terutama karena infeksi oleh HIVDR sehingga akan lebih banyak penderita yang terinfeksi HIVDR multi drug (Kozal, 2009).
Terdapat 2 jenis pemeriksaan resistensi yaitu tes resisten fenotipik (phenotypic resistance test) dan tes resisten genotipik (genotypic resistance test). Tes yang sering digunakan dan lebih murah (berkisar Rp 2.700.000,- hingga Rp 4.500.000,- per tes) adalah tes genotipik. Tes genotipik dilakukan dengan cara mengamplifikasi gen RT dan protease menggunakan teknik RT-PCR dari RNA virus yang bersirkulasi dan dilanjutkan dengan sekuensing produk DNA. Mutasi dilaporkan sebagai perubahan residu asam amino pada kodon spesifik dan diinterpretasikan berdasarkan alogaritma komputer menggunakan data sebelumnya yang sebagian besar data diambil dari virus subtipe B (Shepherd, et al, 2008). Tes genotipik memiliki beberapa keuntungan dibandingkan dengan tes fenotipik, antara lain: adanya standardisasi pemeriksaan tes genotipik, lebih sensitif untuk mendeteksi resisten obat pada penderita yang belum pernah mendapat terapi ARV dan hasil tes dapat digunakan sebagai data epidemiologi molekuler (Shafer, et al. 2008). Selain itu, tes genotipik lebih murah dan lebih cepat daripada tes fenotipik. Tes genotipik memungkinkan untuk menentukan subtipe dari virus HIV dan melihat apakah terjadi kontaminasi pada sampel pemeriksaan (Kuritzkes, 2007b). Kelemahan tes genotipik adalah hanya mampu mendeteksi virus bila viral load virus lebih dari 500-1000 kopi/ml (Parkin, et al. 2000; Shepherd, et al, 2008) dan interpretasi hasil tes genotipik sangat tergantung dari alogaritma yang dibuat
berdasarkan opini para ahli yang lambat diperbaharui. Kesulitan lain adalah sulit untuk memperkirakan efek total interaksi mutasi bila hanya berdasarkan hasil genotipik saja (Kuritzkes, 2007b)
Tes fenotipik dilakukan dengan mengamplifikasi gen RT dan protease yang selanjutnya diklon kedalam suatu vektor pseudotyping HIV dan ditumbuhkan pada jaringan kultur sel. Hasil lebih mudah diinterpretasikan untuk satu jenis obat tetapi akan sulit bila akan menentukan kombinasi obat. Keuntungan tes fenotipik adalah dapat menentukan dosis obat secara spesifik yang diperlukan untuk menghambat virus. Tes fenotipik sangat baik digunakan untuk menentukan resistensi terhadap PI karena dapat menunjukkan akumulasi mutasi yang terjadi (Kuritzkes, 2007b). Kelemahan tes fenotipik adalah waktu tes yang lama dan mahal, tidak menunjukkan subtipe virus, sulit mendeteksi kontaminasi pada spesimen dan nilai standard tes sulit ditentukan (Kuritzkes, 2007b; Shepherd, et al, 2008).
Tes fenotipik virtual merupakan kombinasi dari tes genotipik dan fenotifik. Tes ini menggunakan pendekatan tes genotipik yang disesuaikan dengan data dasar korelasi genotifik dengan hasil tes fenotifik sebelumnya. Tes ini lebih murah dan lebih singkat daripada tes fenotifik dan juga menyediakan nilai standard untuk beberapa jenis obat. Kelemahan dari tes ini adalah versi terakhir belum tervalidasi dalam percobaan klinik (Kuritzkes, 2007b).