L
Journal of Experimental Marine Biology and Ecology 246 (2000) 1–29
www.elsevier.nl / locate / jembe
The feeding behaviour and competition for carrion between
two sympatric scavengers on a sandy shore in Hong Kong:
the gastropod, Nassarius festivus (Powys) and the hermit
crab, Diogenes edwardsii (De Haan)
*
Brian Morton , W.Y. Yuen
The Swire Institute of Marine Science and Department of Ecology and Biodiversity, The University of Hong Kong, Cape d’Aguilar, Shek O, Hong Kong, Hong Kong Received 21 May 1999; received in revised form 11 November 1999; accepted 18 November 1999
Abstract
Field and laboratory experiments compared the feeding behaviours of two marine scavengers, the gastropod, Nassarius festivus and the hermit crab, Diogenes edwardsii, sympatric on the lower intertidal shore at Starfish Bay, Hong Kong. Field trials demonstrated that they both preferred bivalve and fish to other carrion. N. festivus arrived at bait in greater numbers and more quickly
3
than D. edwardsii. Numbers of the former species attracted to fish bait (1 cm ) peaked at a mean of 41 after 30 min, whereas the latter peaked at a mean of only 2.3 after 55 min. Laboratory investigations revealed that N. festivus was able to detect and move fast towards carrion from a distance of .80 cm, feed quickly (x513.20 min) and, after 14 days starvation, had a high
21 21
consumption rate (0.124 mg wet weight or 0.034 mg dry weight of food individual min , or
21
1.4% and 1.3% of its wet and dry body weights min , respectively). In contrast, D. edwardsii had a shorter food detection distance (,10 cm), took longer to arrive at bait, a variable feeding time ranging from 0.28 to .119.75 min and a lower consumption rate (0.003 mg wet weight and
21 21 21
0.001 mg dry weight of food individual min , or 0.1% of its wet and dry body weights min , respectively). Manipulative experiments provided evidence for interspecific competition for carrion between the two species. A morphological advantage, i.e. an extendible proboscis allowing feeding at a distance, and chemoreceptors that permit long distance food detection, combined with numerical superiority on the shore, are mechanisms by which N. festivus outcompetes D. edwardsii and dominates feeding clusters. Interactive behaviour studies revealed the occurrence of interspecific interactions between the two species and intraspecific interactions among D. edwardsii but not N. festivus. The number of interspecific interactions fell when the numbers of N. festivus increased and those of D. edwardsii decreased, and vice versa. The aggressive D. edwardsii did not significantly affect the feeding behaviour of N. festivus. When the number of D.
*Corresponding author. Tel.: 1852-2809-2179; fax: 1852-2809-2197. E-mail address: [email protected] (B. Morton)
edwardsii increased and the number of N. festivus decreased, the number of intraspecific interactions in the former increased. The feeding time of N. festivus, however, decreased with increasing numbers of conspecifics and a decreasing number of D. edwardsii. This suggests the possibility of intraspecific competition in N. festivus. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Competition; Diogenes edwardsii; Feeding behaviour; Interactions; Nassarius festivus; Scavengers
1. Introduction
Marine scavengers feed on ephemeral carrion (Britton and Morton, 1994a). Studies on the feeding behaviour of scavengers on sandy shores in Hong Kong have focused on the Nassariidae (Gastropoda). A well-studied representative of this family is Nassarius
festivus (Powys) (Morton, 1990; Britton and Morton, 1992, 1993, 1994a; Cheung, 1994,
1997) which feeds on moribund bivalves, fish and decapod crustaceans on the sandy shore at Starfish Bay, Hong Kong (Britton and Morton, 1992). Its opportunistic scavenging behaviour includes long distance food detection, fast locomotion towards it, and rapid consumption of an enormous amount relative to its body weight (Morton, 1990; Britton and Morton, 1992).
In addition to nassariid gastropods, many other representatives of different phyla are scavengers of marine carrion, and intra- and interspecific competition can occur between them (Britton and Morton, 1993, 1994a). The feeding behaviours of other potential intertidal scavengers on sandy shores in Hong Kong and interactions, or competition, between them, if any, have received less attention. On the soft shore at Shiu Hau, hermit crabs, including Clibanarius infraspinatus and Diogenes edwardsii, were considered to constitute a scavenging force as important as nassariid gastropods (Morton and Morton, 1983). Hermit crabs and Nassarius festivus were attracted to bivalve and soldier crab (Mictyris longicarpus) baits put out on the shore at Starfish Bay, but the former were attracted in far fewer numbers than the latter (Britton and Morton, 1992). Apart from these observations, the scavenging behaviour of local hermit crabs, as well as their interactions with other scavengers, have not hitherto been studied.
Conversely, a number of studies on the scavenging behaviour of hermit crabs have been conducted elsewhere, but are confined to sublittoral species. Studies on the behaviour of hermit crabs from Otago, New Zealand, demonstrated that 12 out of 15 species used scavenging as a subsidiary feeding mechanism and consumed all sorts of carrion when available (Schembri, 1982). Several studies have identified the hermit crab
Pagurus bernhardus as one of the most important and abundant benthic scavengers that
aggregate in areas of fishing activity in the Irish Sea. Together with starfish, whelks, swimming crabs and fish, they feed upon fisheries discards which fall to the sea bed (Ramsay et al., 1997a,b, 1998). In addition, P. bernhardus was shown to outcompete
Pagurus prideauxi in areas of intense trawling activity (Ramsay et al., 1996, 1998).
numbers of individuals (Ramsay et al., 1997a). Laboratory studies of P. bernhardus and other epibenthic scavenging invertebrates collected from the Clyde Sea showed that the hermit crab was attracted to bait before the others (Nickell and Moore, 1992a,b). Confrontations in the form of short-term approach–retreat cycles were observed frequently among P. bernhardus around the bait (Nickell and Moore, 1992b).
The counterparts of these crabs on intertidal sand flats in Hong Kong are Diogenes
edwardsii and Clibanarius infraspinatus, and both are common at Starfish Bay where
they live sympatrically with Nassarius festivus. This study investigated and compared their food preferences and their sequence of arrival at and departure from bait at this site. The more numerous D. edwardsii was chosen for further investigations of feeding behaviour, in comparison with that of N. festivus, in laboratory experiments. It was hypothesized that N. festivus possesses certain characteristics which confer upon it an advantage in competition with D. edwardsii for carrion, resulting in the dominance of the former over the latter in feeding clusters on the shore. Laboratory experiments involving an investigation of the respective abilities of the two species to detect carrion and their consumption profiles were therefore carried out, and the results compared. Finally, this study also aimed to provide evidence for interspecific competition. This is best studied by altering the densities of both species and monitoring their resource utilization patterns (Underwood, 1986; Begon et al., 1990). The ratio of the numbers of
N. festivus to D. edwardsii in feeding assemblages was manipulated, accordingly, and any changes in feeding patterns monitored.
2. Materials and methods
2.1. Food preferences
Preliminary food preference experiments for Nassarius festivus and hermit crabs were conducted on the sandy shore at Starfish Bay, New Territories, Hong Kong. Fresh carrion, including fish (Lateolabrax sp.), bivalve (Tapes philippinarum), soldier crab (Mictyris longicarpus) and mud-shrimp (Upogebia major) were offered as bait. The four
3
different food items of |2 cm were placed at 2-m intervals across the lower intertidal
shore at a depth of |5 cm during an ebbing tide for 60 min. Scavengers which came to
the carrion were collected by hand and the numbers of each species recorded. The experiments were repeated five times more on different days in October 1998 at different times and locations on the shore. N. festivus, Diogenes edwardsii and Clibanarius
infraspinatus were the commonest scavengers attracted to the bait. The results of the six
experiments upon these three species were pooled so as to assess the attractiveness of the four different baits using a chi-square test for goodness of fit. A significance level of a 50.05 was used for this and all the other subsequent statistical analyses.
2.2. Sequence of arrival and departure
3
Fifteen pieces of |1 cm of fish carrion were placed on the lower intertidal shore at
1.5-m intervals during an ebbing tide. After 5 min, one piece of fish carrion was recovered and the numbers of Nassarius festivus, Diogenes edwardsii and Clibanarius
infraspinatus which approached it, recorded. Other pieces of fish carrion and feeding
clusters were recovered, and the numbers of individuals of each species recorded every 5 min throughout the 75-min experiment. The experiment was repeated six times more on different days in October 1998 and at different times and locations on the shore. The mean numbers of individuals of each species which approached the bait in every 5-min interval were calculated using the pooled data from the seven experiments.
Since the two hermit crabs, Diogenes edwardsii and Clibanarius infraspinatus, were observed to behave similarly in the field, only the former was chosen for further investigations of feeding behaviour in the laboratory.
2.3. Maintenance of animals
Nassarius festivus and Diogenes edwardsii collected from Starfish Bay were kept
separately in aquaria with flow-through seawater at The Swire Institute of Marine Science. Ambient seawater was used to ensure that salinity (29‰) and temperature (22628C) matched those values in the field. Prior to each laboratory experiment, the hunger levels of the animals were standardized by allowing them to feed on fish carrion to satiation and then starving them for 2 weeks.
2.4. Food detection distance
A long aquarium (90330330 cm) was used to investigate the chemoreception ability of Nassarius festivus and Diogenes edwardsii. Seawater in the aquarium flowed
21
unidirectionally at a speed of 3 mm s . Thirty-two similar-sized N. festivus (shell length512.0–15.0 mm) were selected and divided randomly into eight groups of four individuals. Spots of different coloured rapid-drying enamel paints unique to each of the eight groups were marked on the shells facilitating easy identification. Individuals belonging to the same group were arrayed in a line at distances of 10, 20, 30, 40, 50, 60,
3
70 and 80 cm from a piece of fish carrion of |2 cm placed at one end of the aquarium
where the seawater flowed in. The times at which each individual reached the bait and began feeding were recorded. Individuals were, however, restrained from eating the bait by removing them immediately and placing them in a tray without food. Animals that could not locate the bait within 30 min were discounted. Five replicate experiments were conducted using the 32 individuals. The distance of each group of animals from the bait was, however, selected randomly for each trial. The aquarium was also cleaned between each trial. The whole procedure was repeated using Diogenes edwardsii (home shell length512.0–15.0 mm).
positioned closer to the bait were also found to be within detection distance (Britton and Morton, 1994b).
The data collected from the five trials were pooled for each species. The cumulative numbers of individuals arriving at food from each distance were compared by a chi-square test for goodness of fit for each species. The arrival times to food for individuals at each distance were compared by one-way analysis of variance (ANOVA) for each species using the computer software, JMP (Sall and Lehman, 1996).
2.5. Time spent feeding and consumption
Twenty individuals of Nassarius festivus (shell length512.0–15.0 mm) were used in the consumption experiments. Their shells were also marked using different-coloured paints. They were then placed in a 1731437 cm plastic tray with running seawater at
21
a flow rate of 3 mm s and allowed to feed upon a piece of fish carrion of |0.6 g. The
time each individual spent feeding was recorded. N. festivus began feeding when its proboscis was extended to the bait, i.e. the proboscis search reaction (Kohn, 1983). Feeding ended when the individual abandoned the bait. The experiment was terminated when all individuals abandoned the bait.
Prior to each experiment, the bait and similar-sized controls were immersed in seawater for 40 min in order to standardize the amount of contained tissue water. The controls, which were placed in a tray with running seawater in the absence of Nassarius
festivus, were used to determine the error due to natural weight loss to or gain from the
water. They were blotted dry, using standard procedures and weighed to the nearest 0.00001 g. The bait and the controls were also blotted and weighed immediately after the experiment. Shell lengths of those individuals which fed on the bait were measured to the nearest 0.5 mm. Their wet body weights were obtained after removal from the shells and standard blotting. The dry weights of the food and the dead N. festivus were obtained after drying in an oven at 808C for 2 days. This experiment was repeated four times.
The initial wet and dry body weights of Nassarius festivus were obtained from calibration graphs of wet and dry body weights (log ) against shell length. Similarly, thee
initial dry tissue weights of the fish carrion were determined from a calibration graph of dry tissue weight against wet tissue weight. The former were constructed from the wet and dry body weights of 23 N. festivus of known shell lengths (between 9.7 and 15.4 mm) whereas the latter was constructed from the wet and dry weights of 39 pieces of fish carrion (wet weight between 0.01 and 1.97 g).
The difference in the weights of the food before and after feeding by Nassarius
festivus, after adjustment for either weight loss or gain, represented consumption. The
consumption rate was then calculated and expressed in terms of mg wet (or dry) weight 21 21
of food consumed individual min . The amount of food eaten relative to wet (or dry) body weight was expressed as the percentage wet (or dry) body weight consumed
21
min .
h, the experiment was terminated even though some of the individuals had not abandoned the bait but continued to eat. Instead of shell length, the shield length of each individual was measured to the nearest 0.1 mm with an ocular micrometer under a stereomicroscope, after removal from its shell. Shield length is defined as the carapace from the rostrum tip to the cervical suture which separates it from the abdomen (Gherardi et al., 1994). The initial body weights of D. edwardsii were deduced from the calibration graphs of wet and dry body weights (log ) against shield length. Such graphse
were plotted by recording shield lengths and the wet and dry body weights of 20 D.
edwardsii (shield length51.27–2.23 mm).
2.6. Behaviour when feeding together
Twenty individuals of Nassarius festivus to Diogenes edwardsii (shell length512.0– 15.0 mm) were used in each trial of this experiment. The ratio of the numbers of the two species varied in each trial. The ratios used were 20:0, 19:1, 15:5, 10:10, 5:15, 1:19 and 0:20, and the shells of the animals marked with different colours using paint. They were placed at the end of a 1731437 cm plastic tray with running seawater maintained at 3
21 3
mm s and allowed to feed upon |1 cm of fish carrion which was placed on the other
side of the tray at a distance of 12 cm. The seawater flowed in the direction from the carrion to the scavengers. After 5 min, the numbers of individuals which showed any of the behaviours and the numbers of intra- and interspecific interactions described in Tables 1–3 at that particular instance were recorded. Behaviours continued to be recorded at 5-min intervals for 30 min. Three replicate experiments were undertaken for each ratio, but each individual was used only once.
The percentages of the total numbers of individuals of both species showing a certain behaviour at each 5-min interval were calculated for each ratio, using the pooled data from the three replicate experiments. A bar chart showing the changes in the behaviour of the animals over time for each species at each ratio was plotted. Graphs showing the variation in the total numbers of intraspecific interactions among Diogenes edwardsii and interspecific interactions between the two species with changes in the ratios, using the pooled data, were constructed.
2.7. Percentage feeding, arrival time and time spent feeding when together
The experimental procedure identified in Section 2.6 above was repeated. Instead of recording the numbers of individuals showing a particular behaviour and the numbers of
Table 1
Types of behaviour shown by Nassarius festivus
Behaviour Description
Moving in Moving towards the food
Feeding Proboscis extends and penetrates the food
Table 2
Types of behaviour shown by Diogenes edwardsii
Behaviour Description
Moving in Moving towards the food
Feeding It uses its left (larger) cheliped to hold the food and right (smaller) cheliped (occasionally left cheliped) to tear pieces of it and convey them to its mouth
Grasping food Grasps the food with its chelipeds without feeding Moving out Abandons the food and moves away from it (,5 cm) Stationary near food Remains stationary at a distance of ,2 cm from the food
Moving around Moves around
Stationary away from food Remains stationary at a distance of .2 cm from the food Intraspecific interactions
a
Manipulation The aggressor holds the shell of the other crab, the recipient, with its chelipeds and removes it from blocking its way to the food. Recipients are usually individuals which are feeding, grasping the food or near the food a
Cheliped strike Two individuals extend their chelipeds towards each other and make physical contact. Such aggressive behaviour usually occurs between two individuals around the food, between two individuals feeding or grasping the food, or between two individuals feeding / grasping and around the food
a
Ambulatory display Two individuals extend their legs towards each other and make physical contact. Such behaviour occurs between two feeding individuals or between a feeding individual and an individual around the food. This usually results in one of them being driven away
a
Descriptions modified from Ramsay et al. (1997a).
interactions at 5-min intervals, however, the total numbers of individuals of each species feeding upon the fish carrion, the arrival time at food and the time spent feeding by each individual were recorded. The experiment was terminated after 30 min. The time spent feeding by each individual was calculated by adding up all its feeding times during the 30-min trial. A total of five replicate experiments was conducted for each of the seven ratios.
The percentages of the total numbers of individuals of each species feeding was calculated when the frequency data for the experiments corresponding to each ratio were pooled. One-way ANOVA was carried out to test for any significant difference in the percentages of the total number of Diogenes edwardsii feeding upon the food among the
Table 3
Types of behaviour associated with the interspecific interactions between Nassarius festivus and Diogenes edwardsii
Behaviour Description
Cheliped contact A D. edwardsii attacks a feeding or moving-out N. festivus from the front with its chelipeds Grasping A D. edwardsii climbs over and grasps the back
of a feeding or moving-out N. festivus
trials using the ratios of 19:1, 15:5, 10:10, 5:15, 1:19 and 0:20. The Tukey–Kramer test for multiple comparisons of mean percentages was performed because a significant difference was indicated by the ANOVA test (Sokal and Rohlf, 1995).
Mean arrival and mean feeding times were calculated for each species at each ratio and were plotted on graphs showing their variation with the changing ratio of the numbers of Nassarius festivus to Diogenes edwardsii. A Kruskal–Wallis test was performed to compare the arrival times among the different ratios for each species. A nonparametric test was used because of the non-normal distribution of the data (Sokal and Rohlf, 1995). Feeding times were also analyzed in the same way. Tests that indicated significant differences were further analyzed by a Mann–Whitney U-test for pairwise comparisons (Sokal and Rohlf, 1995). The computer software JMP was used for all statistical analyses except the Mann–Whitney U-test (Sall and Lehman, 1996).
3. Results
3.1. Food preferences
All the four baits, i.e. fish, bivalve, soldier crab and shrimp, attracted far greater numbers of Nassarius festivus than Diogenes edwardsii and Clibanarius infraspinatus (Table 4). The attractiveness of the four different baits to each of the three species was analyzed by a chi-square test for goodness of fit. When the data from the six experiments for each of the species were pooled, the chi-square test showed that, for all three, the numbers of individuals attracted to the four baits were significantly different (N. festivus:
2 2
x 5317.43, df53, P,0.001; D. edwardsii: x 536.37, df53, P,0.001; C.
2
infraspinatus: x 525.20, df53, P,0.001). Fish and bivalve baits attracted greater numbers of all three species. The chi-square test for goodness of fit comparing the numbers of N. festivus attracted to their two favourite food items, i.e. fish and bivalve, suggested that the latter was preferred by a significantly higher number of N. festivus
2
than the former (x 553.66, df51, P,0.001). Conversely, the same statistical tests demonstrated that the numbers of hermit crabs attracted to their two favourite baits, fish
Table 4
Cumulative numbers (six experiments) of Nassarius festivus, Diogenes edwardsii and Clibanarius infras-pinatus attracted to four different baits at Starfish Bay in October 1998. The attractiveness of the four different
a baits to each of the three species was analyzed by a chi-square test for goodness of fit
2
Species Bait df x P
Fish Bivalve Soldier crab Shrimp
Nassarius festivus 1333 1739 1114 878 3 317.43 ,0.001
(222.2) (289.8) (185.7) (146.3)
Diogenes edwardsii 25 30 2 6 3 36.37 ,0.001
(4.2) (5.0) (0.3) (1.0)
Clibanarius infraspinatus 20 21 1 6 3 25.20 ,0.001
(3.3) (3.5) (0.2) (1.0)
a
and bivalve, were insignificantly different for both D. edwardsii and C. infraspinatus (D.
2 2
edwardsii: x 50.45, df51, P.0.05; C. infraspinatus: x 5 0.02, df51, P.0.05).
3.2. Sequence of arrival and departure
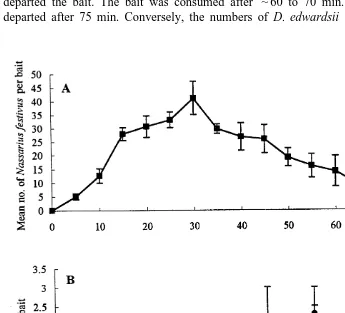
The results of the seven feeding experiments conducted in the field are illustrated in
3
Fig. 1. After 5 min, fish baits of 1 cm each had attracted a mean of five Nassarius
festivus. The mean number of Clibanarius infraspinatus arriving at the bait was,
however, less than 1. No Diogenes edwardsii arrived within 5 min. The numbers of N.
festivus attracted to the bait increased rapidly up to a mean of about 28 after 15 min and
peaked at a mean of 41 after 30 min. Thereafter, an increasing number of N. festivus departed the bait. The bait was consumed after |60 to 70 min. All individuals had
departed after 75 min. Conversely, the numbers of D. edwardsii and C. infraspinatus
attracted to the bait remained low, and were far less than N. festivus throughout the experiment. The mean numbers of the two species of hermit crabs did not exceed three. Both showed a similar pattern of arrival and departure at the bait. Numbers for both species increased slowly until they peaked at 55 min, with a mean of 2.3 and 1.7 for D.
edwardsii and C. infraspinatus, respectively. Individual numbers reached a maximum, 25 min later than N. festivus. The times at which all individuals departed the bait were 70 and 75 min for C. infraspinatus and D. edwardsii, respectively.
It should be noted that, although both species of hermit crabs were attracted to the bait, some which arrived late were unable to find their way through the large congregation of Nassarius festivus surrounding the fish so as to feed. They either grasped some of the feeding N. festivus or remained stationary nearby. Those which arrived early, however, and managed to climb over the N. festivus did eat.
3.3. Food detection distance
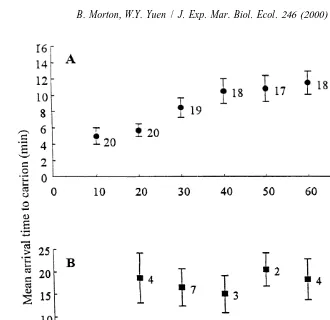
Nassarius festivus was able to locate the fish carrion from all distances up to 80 cm
(Table 5). When the data from the five experiments were pooled, over 50% of the total number of N. festivus from each of the tested distances reached the food. They were considered to be within the detection distance of the bait by application of the 50% rule. The chi-square test for goodness of fit showed no significant difference in the numbers
2
of N. festivus arriving at the bait from any of the tested distances (x 52.0, df57,
P.0.05). Although it can be suggested that such an insignificant difference was due to random encounters of N. festivus with the bait, it is more likely that 80 cm is less than the maximum food detection distance of |2.5 m (Britton and Morton, 1992). This is
reflected by the significant difference in the arrival times to food by individuals from different distances according to one-way ANOVA using the pooled data from the five experiments (F54.90, P,0.0001). There was a trend of increasing arrival time at fish carrion with distance (Fig. 2A).
Conversely, the percentages of the total number of Diogenes edwardsii reaching the carrion from all the tested distances were less than Nassarius festivus (Table 5). Since all percentages were ,50%, none of the tested distances was considered to be within the detection range of D. edwardsii. When the data from the five experiments were
Table 5
Cumulative numbers (five experiments) of Nassarius festivus and Diogenes edwardsii arriving at fish carrion
21
from various distances in seawater flowing at 3 mm s . The numbers of arrivals from different distances were a
analyzed by a chi-square test for goodness of fit for each species
2
Species Distance from carrion (cm) df x P
10 20 30 40 50 60 70 80
Fig. 2. The mean arrival times at fish carrion by (A) Nassarius festivus and (B) Diogenes edwardsii positioned
21
at various distances from it in seawater flowing at 3 mm s . Numerals indicate the numbers of individuals represented by each data point. Bars indicate the standard error of the mean.
pooled, a chi-square test for goodness of fit demonstrated that there was no significant
2
difference in the numbers of D. edwardsii arriving from different distances (x 57.0, df57, P.0.05). No significant difference in arrival times from different distances was detected by one-way ANOVA using the pooled data (F51.71, P50.1524). No clear relationship between arrival time and distance was, therefore, revealed (Fig. 2B). These data suggest that the food detection distance of D. edwardsii was ,10 cm and it appeared to move randomly in the tank until it was in close proximity to the food and reached its food detection distance. It then moved towards it.
When the mean arrival times at food by the two species were compared, Nassarius
3.4. Time spent feeding and consumption
Correlation coefficients for the data related to wet and dry body weights (log ) againste
shell length of Nassarius festivus and Diogenes edwardsii were highly significant (N.
festivus: r50.9416 and 0.9376, respectively; D. edwardsii: r50.9761 and 0.9760, respectively) where P,0.0001. The regression equations obtained were used to estimate the initial wet and dry body weights of the animals before they fed upon the fish carrion. Similarly, the correlation coefficient for the data related to dry tissue weight against wet tissue weight of fish carrion was highly significant (r50.9975, P,0.0001). The regression equation obtained was used to estimate the dry weights of fish carrion prior to feeding by Nassarius festivus and Diogenes edwardsii.
The results of the five consumption experiments on Nassarius festivus and Diogenes
edwardsii are summarized in Table 6. Data from the five experiments were pooled. The
percentage of the total numbers of N. festivus feeding on the carrion was very high (96%) and was greater than D. edwardsii (69%). The mean time spent feeding on a single meal by N. festivus was 13.20 min (Table 6). The equivalent figure for D.
edwardsii could not, however, be estimated as some individuals continued to eat at the
end of the experimental time. The time spent feeding by D. edwardsii thus varied considerably from 0.28 min to 119.75 min during the 2-h experiments (Table 6).
The mean consumption rates for Nassarius festivus were calculated to be 0.124 mg 21 21
wet weight and 0.034 mg dry weight of food individual min . The equivalent figures for Diogenes edwardsii were 0.003 mg and 0.001 mg, respectively (Table 6). When these figures were compared between the two species, the amount of food eaten
21 21
individual min by N. festivus was over 30 times that of D. edwardsii.
Table 6
Summary of the results of the five consumption experiments using Nassarius festivus and Diogenes edwardsii a
feeding on fish carrion
Nassarius festivus Diogenes edwardsii
Total number feeding 96 69
Percentage of animals feeding 96% 69%
Mean time spent feeding on a 13.2064.73 –
single meal (min) (5.45, 28.50) (0.28, .119.75)
Mean time spent feeding during 13.20 60.72
the experiment (min) Mean consumption rate
21 21
(mg wet weight ind min ) 0.123560.0432 0.003460.0014 (0.0642, 0.1850) (0.0012, 0.0045)
21 21
(mg dry weight ind min ) 0.033660.0126 0.001060.0004 (0.0166, 0.0486) (0.0007, 0.0015)
21
Mean % wet body weight min 1.4 0.1
(with shell removed)
21
Mean % dry body weight min 1.3 0.1
(with shell removed) a
When the consumption rates were expressed in terms of the percentage wet and dry 21
body weights consumed min , Nassarius festivus consumed a mean of 1.4% of its own
21 21
wet body weight min (18.5% average feeding bout ) and a mean of 1.3% of its own
21 21
dry body weight min (17.2% average feeding bout ). Conversely, Diogenes 21 edwardsii only consumed a mean of 0.1% of its own wet and dry body weights min
(Table 6). N. festivus thus ate 13 to 14 times faster than D. edwardsii. The mean consumption rates, which were expressed in terms of percentage body weight consumed
21
min , were calculated based on a reduced data set. This was because a few percentage values were negative, and eliminated from the analysis. As a result, the size of the final data set was reduced to 88 N. festivus individuals and 61 D. edwardsii individuals.
3.5. Behaviour when feeding together
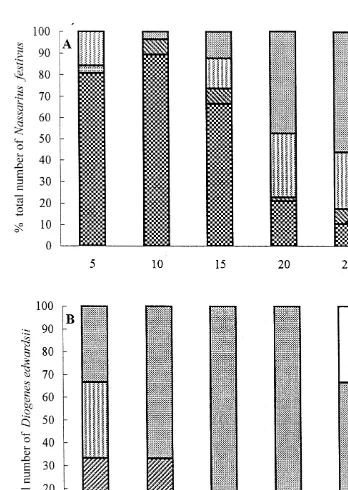
A clear sequence of behaviour associated with feeding by Nassarius festivus was shown. Figs. 3, 4A, 5A, 6A, 7A and 8A correspond to the ratios of the numbers of N.
festivus to Diogenes edwardsii, i.e. 20:0, 19:1, 15:5, 10:10, 5:15 and 1:19, respectively.
The patterns of behaviour were similar for all these ratios. They first moved randomly in the tray, followed by directed movement towards the carrion and commenced feeding. At 5 min, .60% of N. festivus had fed upon the food in all cases (Figs. 3–8). The
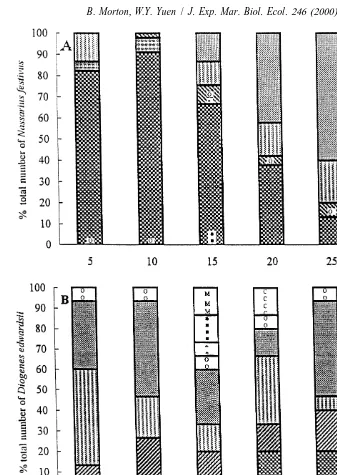
Fig. 5. Behaviour of (A) Nassarius festivus and (B) Diogenes edwardsii when they were allowed to feed together in the ratio of 15:5. The percentage of the total number of individuals showing a certain behaviour was calculated using the pooled data from three experiments, i.e. a total of 45 N. festivus and 15 D. edwardsii were used (interspecific interaction* — cheliped contact of moving-out N. festivus by D. edwardsii; interspecific interaction** — grasping of feeding / moving-out N. festivus by D. edwardsii; interspecific interaction*** — manipulation of feeding N. festivus by D. edwardsii; intraspecific interaction among D.
[ [[
Fig. 6. Behaviour of (A) Nassarius festivus and (B) Diogenes edwardsii when they were allowed to feed together in the ratio of 10:10. The percentage of the total number of individuals showing a certain behaviour was calculated using the pooled data from three experiments, i.e. a total of 30 N. festivus and 30 D. edwardsii were used (interspecific interaction** — grasping of feeding / moving-out N. festivus by D. edwardsii; interspecific interaction*** — manipulation of feeding N. festivus by D. edwardsii; intraspecific interaction
[ [[
Fig. 7. Behaviour of (A) Nassarius festivus and (B) Diogenes edwardsii when they were allowed to feed together in the ratio of 5:15. The percentage of the total number of individuals showing a certain behaviour was calculated using the pooled data from three experiments, i.e. a total of 15 N. festivus and 45 D. edwardsii were used (interspecific interaction* — cheliped contact of feeding / moving-out N. festivus by D. edwardsii; interspecific interaction** — grasping of feeding N. festivus by D. edwardsii; interspecific interaction*** — manipulation of feeding / moving-out N. festivus by D. edwardsii; intraspecific interaction among D.
[ [[
edwardsii — manipulation; intraspecific interaction among D. edwardsii — cheliped strike; intraspecific
[[[
Fig. 8. Behaviour of (A) Nassarius festivus and (B) Diogenes edwardsii when they were allowed to feed together in the ratio of 1:19. The percentage of the total number of individuals showing a certain behaviour was calculated using the pooled data from three experiments, i.e. a total of three N. festivus and 57 D. edwardsii were used (interspecific interaction*** — manipulation of feeding N. festivus by D. edwardsii;
[ [[
intraspecific interaction among D. edwardsii — manipulation; intraspecific interaction among D. edwardsii
[[[
percentage could be as high as 100%, as in the experiment using the ratio of 1:19 (Fig. 8). For all the six ratios, the percentage of N. festivus feeding peaked at a value of about or .90% at 5 to 10 min. Feeding was observed to be the dominant behaviour between 5 to 15 min. After feeding, they moved away from the food. Subsequently, they either moved around in the tray or remained stationary. The latter behaviour was observed to be the most common (.46%) at 25 to 30 min.
Unlike Nassarius festivus, the sequence of behaviour associated with feeding by
Diogenes edwardsii is less clearly shown in Figs. 4B, 5B, 6B, 7B, 8B and 9, which
correspond to the ratios of 19:1, 15:5, 10:10, 5:15, 1:19 and 0:20, respectively. It was, however, similar to that of N. festivus, i.e. moving randomly, moving towards, feeding, moving away and either moving or remaining stationary away from food, except that D.
edwardsii often stopped feeding and grasped the food with their chelipeds for some time
before resuming. Some also remained stationary near the food before approaching it; such behaviour was recorded during the 5 to 20 min period in Fig. 5B. Some approached and ate more than once. Some D. edwardsii, usually those some distance from the food,
Fig. 9. Behaviour of 20 Diogenes edwardsii when they were allowed to feed together in the absence of Nassarius festivus. The percentage of the total number of individuals showing a certain behaviour was calculated using the pooled data from three experiments, i.e. a total of 60 D. edwardsii were used (intraspecific
[ [[ [[[
either moved or remained stationary showing no other behaviours. That is, not all individuals fed during the experiments.
The patterns of behaviour of Diogenes edwardsii were not similar in the experiments using different ratios. The remaining stationary behaviour away from the food was common when the ratios of 19:1 and 15:5 were used (Figs. 4B and 5B). For the ratios of 10:10, 5:15, 1:19 and 0:20, the behaviour shown by a majority of D. edwardsii (35–70%) was moving randomly in the tray (Figs. 6 to 9). The percentage of D.
edwardsii feeding varied with changes in the ratio. None of the individuals fed at the
ratio of 19:1 (Fig. 4B). At the ratio of 15:5, D. edwardsii began to feed after 10 min (Fig. 5B). As the number of Nassarius festivus declined and the numbers of D.
edwardsii increased, an increasing percentage of the hermit crab either fed or grasped
the food at an earlier time. At the ratio of 0:20, .13% of them either fed or grasped the food throughout the 30-min experiment (Fig. 9). Unlike N. festivus, for which the percentage of feeding individuals always peaked in the early stages of the experiment, the percentage of D. edwardsii which either fed or grasped the food increased slowly. Intra- and interspecific interactions were also investigated. No intraspecific interaction among Nassarius festivus individuals was detected. Conversely, intraspecific interactions among Diogenes edwardsii, including manipulation, cheliped strike and ambulatory display, were observed (Figs. 5–9). The intraspecific interactions increased when numbers increased and the numbers of N. festivus decreased, but remained at 12 after the ratio of 1:19 was reached (Fig. 10A).
Interspecific interactions between Nassarius festivus and Diogenes edwardsii were detected. Such interactions involved cheliped contact, grasping and manipulation of the former by the latter (Figs. 4–8). Numbers of interspecific interactions were greater at the ratios of 15:5, 10:10 and 5:15 (Fig. 10B), i.e. they fell when the numbers of N. festivus increased and those of D. edwardsii decreased, and vice versa.
3.6. Percentage feeding, arrival time and time spent feeding when together
All nassariids fed upon the carrion in all the experimental trials at ratios of Nassarius
festivus to Diogenes edwardsii of 20:0, 19:1, 15:5, 10:10, 5:15 and 1:19 (Fig. 11). Even
though the numbers of D. edwardsii increased from 0 to 19, all N. festivus were able to feed. Conversely, the percentages of the total number of D. edwardsii feeding upon the carrion were never .65% (Fig. 11). One-way ANOVA showed a significant difference in the percentages of the total number of D. edwardsii feeding among the experiments using the ratios of 19:1, 15:5, 10:10, 5:15, 1:19 and 0:20 (F524.29, P,0.0001). The Tukey–Kramer test for multiple comparisons showed that the mean percentages of the total numbers of D. edwardsii feeding at the ratio of 19:1 was significantly lower than that for the other five ratios and that the ratio of 15:5 was also significantly lower than that of the other ratios, except 19:1 (P,0.05). This means that the percentage number of D. edwardsii feeding increased with a decreasing number of N. festivus and an increasing number of D. edwardsii, but such a percentage levelled off when the numbers of the former fell to 10 and the latter rose to 10 (Fig. 11).
The Kruskal–Wallis test detected no significant difference in the arrival times of
Fig. 10. The total numbers of (A) intraspecific interactions among D. edwardsii and (B) interspecific interactions between N. festivus and D. edwardsii recorded during the 30-min experiment, using the pooled data from three trials, when different ratios of the number of N. festivus to D. edwardsii were used. The numbers of these two kinds of interactions were recorded at 5-min intervals.
Similarly, the arrival times of Diogenes edwardsii differed insignificantly (H53.27, df54, P50.5133), i.e. there was no obvious trend in the mean arrival time with changes in the ratio of the number of N. festivus to D. edwardsii for each species (Fig. 12).
The feeding times of both Nassarius festivus and Diogenes edwardsii were shown to differ significantly among the trials using different ratios, as analyzed by the Kruskal– Wallis tests (N. festivus: H544.19, df55, P,0.0001; D. edwardsii: H59.60, df54,
Fig. 11. The percentages of the total numbers of Nassarius festivus and Diogenes edwardsii feeding upon fish carrion when they were allowed to feed together in different ratios.
significantly lower than those at the ratios of 15:5 and 10:10 (Mann–Whitney U-test for pairwise comparisons: z54.30 for 20:0 vs. 15:5; z55.35 for 20:0 vs. 10:10; z53.15 for 19:1 vs. 15:5; z54.66 for 19:1 vs. 10:10; P,0.0001 for all the above four pairs). The Mann–Whitney U-test for pairwise comparisons showed that N. festivus, at the ratio of 5:15, fed for a significantly shorter time than those at a ratio of 10:10, but significantly longer than those at a ratio of 20:0 (z5 22.84, P50.0023 and z52.55,
P50.0054, respectively). These observations indicate that the mean feeding time of N.
festivus increased when its number fell from 20 to 10 and the numbers of D. edwardsii increased from 0 to 10, but decreased slightly when numbers dropped further to 1 and the numbers of the hermit crab increased further to 19 (Fig. 13A).
The Mann–Whitney U-test for pairwise comparisons suggested that the feeding times of Diogenes edwardsii at the ratios of 15:5 and 10:10 were both significantly shorter than those at the ratio of 0:20 (z5 22.20, P50.0139 and z5 22.75, P50.0030, respectively). These data imply that the mean feeding time of D. edwardsii increased with an increasing number of conspecifics and a decreasing number of Nassarius festivus (Fig. 13B).
4. Discussion
Nassarius festivus and the hermit crabs Diogenes edwardsii and Clibanarius infraspinatus approached different carrion baits on the shore at Starfish Bay and fed.
Fig. 12. The mean arrival time at fish carrion by (A) Nassarius festivus and (B) Diogenes edwardsii when different ratios of the number of N. festivus to D. edwardsii were used. Only the time taken for an individual to arrive at the food the first time was used to calculate the mean arrival time. Numerals indicate the numbers of individuals represented by each data point. Bars indicate the standard error of the mean.
(1992). Both species of hermit crabs preferred bivalve and fish to other baits. Tapes
philippinarum is the dominant bivalve species on the lower intertidal shore at Starfish
Bay (Morton and Chiu, 1990) and represents autochthonous, or endemic, carrion (Britton and Morton, 1992). Fish carrion was the second most preferred bait by N.
festivus and a great number of them were also attracted to it. It represents allochthonous
carrion which is washed in by the tide (Britton and Morton, 1992). N. festivus, D.
edwardsii and C. infraspinatus, therefore, exploit both autochthonous and allochthonous carrion on the shore.
Fig. 13. The mean time spent feeding upon fish carrion by (A) Nassarius festivus and (B) Diogenes edwardsii when different ratios of the number of N. festivus to D. edwardsii were used. The time spent on food by each individual was recorded within 30 min from the beginning of the experiments. Numerals indicate the numbers of individuals represented by each data point. Bars indicate the standard error of the mean.
showed that a decrease in the former and an increase in the latter led to an increase in the percentage of hermit crabs either feeding or grasping the food and an increase in feeding time. That is, the opportunity to access the carrion resource by D. edwardsii increased. The time spent feeding by N. festivus also fell after a loss of dominance at ratios of 5:15 and 1:19. These experimental results suggest that the two sympatric scavengers compete with each other for the limited carrion resources on sandy shores in Hong Kong, as suggested by Britton and Morton (1993, 1994a).
Field experiments showed that Nassarius festivus was the dominant scavenger which arrived at the bait in far greater numbers and more quickly than Diogenes edwardsii. The former is more dominant at lower tidal levels but less so higher on the shore. In the manipulative laboratory experiments, however, regardless of what ratios of N. festivus to
D. edwardsii were used, all the former fed and the pattern of their feeding behaviour changed little. These results collectively indicate that N. festivus outcompetes D.
edwardsii for carrion which is, thus, a resource distributed unequally between them.
causing dominance by the former over the latter (Kaiser et al., 1998). Examples of this can be found in the interspecific competition for fisheries discards between sympatric species of hermit crabs, Pagurus bernhardus and P. prideauxi, in the North Sea. The former outcompeted the latter because of larger chelae and more aggressive behaviour (Kaiser et al., 1998). Similarly, the greater mechanical and muscular strength, as well as aggressiveness, of the red form of Carcinus maenas, an intertidal crab in the Menai Straits, North Wales, place them at a competitive advantage over the green form when predating mussels (Kaiser et al., 1990). On Starfish Bay, Hong Kong, Nassarius festivus possesses a number of morphological and behavioural traits that confer upon it a competitive advantage over Diogenes edwardsii (and Clibanarius infraspinatus) in exploiting carrion resources. N. festivus sweeps its siphon across the direction of the water flow on the shore and moves towards the food (Britton and Morton, 1992). Its siphon possibly bears chemoreceptor cells, as in N. reticulatus (Crisp, 1971), thus enhancing its ability to detect chemical stimuli at a distance and orient itself towards the carrion. In contrast, aquatic crabs, including intertidal hermit crabs, locate food through less sensitive antennular chemoreceptors (Dunham and Gilchrist, 1988).
The long proboscis of Nassarius festivus allows it to feed through a large number of conspecifics crowded around a bait. The morphology of Diogenes edwardsii does not allow this and in order to feed it must have close contact with the food. This accounts for the failure of some D. edwardsii to feed upon bait surrounded by large numbers of N.
festivus in the field.
The food detection distance of Nassarius festivus exceeded the maximum tested distance of 80 cm and it can reportedly arrive at fish carrion from |2.5 m (Britton and
Morton, 1992). Conversely, the maximum food detection distance of Diogenes
edwar-dsii was much shorter at | ,10 cm. Unlike N. festivus, which moved purposely
towards the bait from a long distance, D. edwardsii moved randomly. Only after it came in close proximity (|6 cm) to the food, did it move towards it. In studies conducted by
Nickell and Moore (1992a,b) on epibenthic scavenging invertebrates collected from the Clyde Sea, the hermit crab Pagurus bernhardus was shown to be more effective at locating bait than other invertebrates, including the gastropod Buccinum undatum, in both static and moving water. The subtidal Babylonia lutosa (Buccinidae) has a poorer ability for distance chemoreception than the intertidal nassariid, N. festivus (Morton, 1990) so that both scavenging gastropods and hermit crabs living in the intertidal and subtidal may have different chemoreception abilities.
Nassarius festivus and Diogenes edwardsii differed in the speed they moved towards
food, the former moving faster. This is reflected in the shorter mean arrival time at food by the former than the latter from all of the tested distances, except 80 cm. The different result obtained for 80 cm can be explained by the small sample size of only two D.
edwardsii arriving at the bait.
Nassarius festivus fed more quickly on carrion than Diogenes edwardsii. N. festivus spent a mean time of 13.20 min on a single meal after 14 days starvation. The value obtained for the same species by Morton (1990) was 7.97 min after 10 days starvation. In contrast, the time spent feeding on a single meal by D. edwardsii varied greatly from 0.28 to .119.75 min. The consumption rate of N. festivus far exceeded that of D.
21 21 21
individual min or percentage wet (or dry) body weight min . The mean time spent feeding by D. edwardsii could, however, have been overestimated because they ate intermittently, grasped the bait and either remained stationary or interacted with conspecifics before feeding resumed. Hence, although consumption rates for this species could have been underestimated it did feed more slowly than N. festivus.
Nassarius festivus not only has morphological and behavioural competitive superiority
over Diogenes edwardsii, it is also dominant numerically on the Starfish Bay shore. The 22
population of N. festivus has, moreover, increased dramatically from ,4 m in 1973 22
to .60 m in 1994 (Britton and Morton, 1994a) owing to its resistance to pollution (Britton and Morton, 1992; Morton, 1992).
The possession of a siphon and proboscis, better distance chemoreception, faster locomotion, faster consumption and numerical dominance on the shore by Nassarius
festivus gives it greater efficiency at exploiting carrion resources and, thus, an advantage
over Diogenes edwardsii in interspecific competitions for food. This results in the dominance of N. festivus over D. edwardsii in feeding clusters on the shore. It is predicted that another common species of hermit crab, Clibanarius infraspinatus, is also less dominant in feeding clusters on the shore for similar reasons. N. festivus and hermit crabs account for 96.3% and 3.6% of all individuals, respectively, in carrion feeding clusters at Starfish Bay (Britton and Morton, 1992).
Interspecific competition varies from slight to intense depending on how much overlap there is in the resource requirements of the species involved and how much similarity in their morphology (Branch, 1984; Kaiser et al., 1998). Laboratory experi-ments involving the manipulation of the ratio of the numbers of Nassarius festivus to
Diogenes edwardsii demonstrated that the percentage number of the latter feeding upon
carrion in the absence of the former was not very high (63%) when compared with that of N. festivus feeding in either the absence or presence of D. edwardsii (100%). This suggests their feeding niches do not overlap greatly and, thus, that the intensity of interspecific competition is not high.
Moribund tissue is not the primary source of food for many marine scavengers and scavenging serves only to supplement diets when carrion is available (Britton and Morton, 1994a). Most hermit crab species are omnivorous microphagous detritivores (Hazlett, 1981; Wight et al., 1990; Britton and Morton, 1994a; Tudge, 1995) and detritus is their primary source of nutrients (Hazlett, 1981). They spend a great deal of time dredging up sediments with their chelipeds and sorting out organic material with their mouthparts (Jensen, 1995). Macrophagous carrion scavenging is only a subsidiary feeding mechanism for many hermit crabs (Schembri, 1982). Diogenes edwardsii can, therefore, be regarded as a facultative, opportunistic, scavenger which supplements its diet with carrion when it is available (Britton and Morton, 1994a). Conversely, species of Nassariidae, including Nassarius festivus, along with lysianassid amphipods, represent the closest attempt at an obligate scavenging lifestyle (Britton and Morton, 1993, 1994a).
Nassarius pyrrhus and Cominella eburnea were dominant scavengers on Bramble
Beach in Western Australia (Morton and Britton, 1991). The additional predatory habits of C. eburnea played an important role in supporting the nassariid scavengers there, the carrion supply being insufficient to account for their high densities and diversity. This is similar to the situation at Starfish Bay where the additional detritivory of Diogenes
edwardsii permits the existence of large populations of scavengers, including N. festivus and other hermit crabs, on the shore. This agrees with the views held by Branch (1984) and Keddy (1989) that niche differences between competitors, as described above, allow them to coexist. Coexistence is the usual consequence of competition for food among marine organisms and accounted for 90% of all such cases reviewed by Branch (1984). Interspecific interactions between Nassarius festivus and Diogenes edwardsii occur, with the latter taking the role of aggressor and the former the role of recipient. The feeding behaviour of N. festivus was not, however, affected significantly by interference from D. edwardsii. Only when the numbers of N. festivus decreased from 10 to 5 and those of D. edwardsii increased from 10 to 15, i.e. N. festivus lost its numerical dominance, did the feeding time of the nassariid shorten. This can be partly attributed to the increase in the incidence of attacks on feeding N. festivus by D. edwardsii. The situation in the field may, however, be different from that in the laboratory. It is likely that, because of the numerical dominance of N. festivus on the lower intertidal shore of Starfish Bay, any attack from D. edwardsii may not significantly reduce the feeding time of the former.
edwardsii would be less intense than that observed in the laboratory because such
feeding clusters are dominated by N. festivus (Britton and Morton, 1992).
Though no obvious intraspecific interactions among Nassarius festivus were detected and there is no published literature on such competition, it is unreasonable to exclude the possibility of its existence. In this study, feeding time was reduced significantly when numbers of N. festivus increased from 10 to 20 and numbers of Diogenes edwardsii fell from 10 to 0. This implies that N. festivus tended to finish feeding and leave the bait earlier when crowding around it increased. Some N. festivus, unable to penetrate the cluster of feeding individuals gathering on dying clams in the field, were observed to turn away (Cheung, 1994). A possible mechanism of intraspecific competition could involve exploitation. Individuals of N. festivus do not directly, but do indirectly affect each other through a reduction in the available pool of a common carrion resource (Branch, 1984; Keddy, 1989). Exploitation competition is common among marine animals which compete for food (Branch, 1984; Wolcott, 1988). In contrast, intraspecific competition for carrion among D. edwardsii is interference competition as it involves physical attack which can either hinder or deny another’s access to the carrion resource (Branch, 1984; Keddy, 1989). [RW]
References
Begon, M., Harper, J.L., Townsend, C.R., 1990. In: 2nd Edition, Ecology: Individuals, Populations and Communities, Blackwell Scientific, Oxford.
Branch, G.M., 1984. Competition between marine organisms: ecological and evolutionary implications. Oceanogr. Mar. Biol. Annu. Rev. 22, 429–593.
Britton, J.C., Morton, B., 1992. The ecology and feeding behaviour of Nassarius festivus (Prosobranchia: Nassariidae) from two Hong Kong Bays. In: Morton, B. (Ed.), The Marine Flora and Fauna of Hong Kong and Southern China III. Proceedings of the Fourth International Marine Biological Workshop: The Marine Flora and Fauna of Hong Kong and Southern China, Hong Kong, 1989, Hong Kong University Press, Hong Kong, pp. 395–416.
Britton, J.C., Morton, B., 1993. Are there obligate marine scavengers? In: Morton, B. (Ed.), The Marine Biology of the South China Sea. Proceedings of the International Conference on the Marine Biology of Hong Kong and the South China Sea, Hong Kong, 1990, Hong Kong University Press,, Hong Kong, pp. 357–391.
Britton, J.C., Morton, B., 1994a. Marine carrion and scavengers. Oceanogr. Mar. Biol. Annu. Rev. 32, 369–434.
Britton, J.C., Morton, B., 1994b. Food choice, detection, time spent feeding, and consumption by two species of subtidal Nassariidae from Monterey Bay, California. Veliger 37, 81–92.
Cheung, S.G., 1994. Feeding behaviour and activity of the scavenging gastropod Nassarius festivus (Powys). In: Morton, B. (Ed.), The Malacofauna of Hong Kong and Southern China III. Proceedings of the Third International Workshop on the Malacofauna of Hong Kong and Southern China, Hong Kong, 1992, Hong Kong University Press, Hong Kong, pp. 327–338.
Cheung, S.G., 1997. Physiological and behavioural responses of the intertidal scavenging gastropod Nassarius festivus to salinity changes. Mar. Biol. 129, 301–307.
Crisp, M., 1971. Structure and abundance of receptors of the unspecialized external epithelium of Nassarius reticulatus (Gastropoda, Prosobranchia). J. Mar. Biol. Assoc. UK 51, 865–890.
Dunham, D.W., Gilchrist, S.L., 1988. Behavior. In: Burggren, W.W., McMahon, B.R. (Eds.), Biology of the Land Crabs, Cambridge University Press, Cambridge, pp. 97–138.
Hazlett, B.A., 1981. The behavioral ecology of hermit crabs. Annu. Rev. Ecol. Syst. 12, 1–22. Jensen, G.C., 1995. Pacific Coast Crabs and Shrimps, Sea Challengers, California.
Kaiser, M.J., Hughes, R.N., Reid, D.G., 1990. Chelal morphometry, prey-size selection and aggressive competition in green and red forms of Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 140, 121–134. Kaiser, M.J., Ramsay, K., Hughes, R.N., 1998. Can fisheries influence interspecific competition in sympatric
populations of hermit crabs? J. Nat. Hist. 32, 521–531. Keddy, P.A., 1989. Competition, Chapman and Hall, London.
Kohn, A.J., 1983. Feeding biology of gastropods. In: Saleuddin, A.S.M., Wilbur, K.M. (Eds.), Physiology, Part 2, The Mollusca, Vol. 5, Academic Press, New York, pp. 1–63.
Morton, B., 1990. The physiology and feeding behaviour of two marine scavenging gastropods in Hong Kong: the subtidal Babylonia lutosa (Lamarck) and the intertidal Nassarius festivus (Powys). J. Moll. Stud. 56, 275–288.
Morton, B., 1992. Hong Kong’s marine scavengers. In: Ades, G., Reels, G.T. (Eds.), Porcupine No. 2, Newsletter of The Hong Kong University Ecology Research Group, Kadoorie Agriculture Research Centre, University of Hong Kong, Hong Kong, p. 7.
Morton, B., Britton, J.C., 1991. Resource partitioning strategies of two sympatric scavenging snails on a sandy beach in Western Australia. In: Wells, F.E., Walker, D.I., Kirkman, H., Lethbridge, R. (Eds.), Proceedings of the Third International Marine Biological Workshop: The Marine Flora and Fauna of Albany, Western Australia, 1988, Western Australian Museum, Perth, pp. 579–595.
Morton, B., Chiu, S.T., 1990. The diet, prey size and consumption of Philine orientalis (Opisthobranchia: Philinidae) in Hong Kong. J. Moll. Stud. 56, 289–299.
Morton, B., Morton, J., 1983. The Sea Shore Ecology of Hong Kong, Hong Kong University Press, Hong Kong.
Nickell, T.D., Moore, P.G., 1992a. The behavioural ecology of epibenthic scavenging invertebrates in the Clyde Sea area: laboratory experiments on attractions to bait in static water. J. Exp. Mar. Biol. Ecol. 156, 217–224.
Nickell, T.D., Moore, P.G., 1992b. The behavioural ecology of epibenthic scavenging invertebrates in the Clyde Sea area: laboratory experiments on attractions to bait in moving water, underwater TV observations in situ and general conclusions. J. Exp. Mar. Biol. Ecol. 159, 15–35.
Ramsay, K., Kaiser, M.J., Hughes, R.N., 1996. Changes in hermit crab feeding patterns in response to trawling disturbance. Mar. Ecol. Prog. Ser. 144, 63–72.
Ramsay, K., Kaiser, M.J., Hughes, R.N., 1997a. A field study of intraspecific competition for food in hermit crabs (Pagurus bernhardus). Estuar. Coast. Shelf Sci. 44, 213–220.
Ramsay, K., Kaiser, M.J., Moore, P.G., Hughes, R.N., 1997b. Consumption of fisheries discards by benthic scavengers: utilization of energy subsidies in different marine habitats. J. Anim. Ecol. 66, 884–896. Ramsay, K., Kaiser, M.J., Hughes, R.N., 1998. Responses of benthic scavengers to fishing disturbance towed
gears in different habitats. J. Exp. Mar. Biol. Ecol. 224, 73–89.
Sall, J., Lehman, A., 1996. JMP Start Statistics: A Guide to Statistics and Data Analysis Using JMP and JMP in Software, SAS Institute Inc, California.
Schembri, P.J., 1982. Feeding behaviour of fifteen species of hermit crabs (Crustacea: Decapoda: Anomura) from the Otago region, southeastern New Zealand. J. Nat. Hist. 16, 859–878.
Sokal, R.R., Rohlf, F.J., 1995. In: 3rd Edition, Biometry: The Principles and Practice of Statistics in Biological Research, W.H. Freeman, New York.
Tudge, C.C., 1995. Hermit Crabs of the Great Barrier Reef & Coastal Queensland, Backhuys, Brisbane. Underwood, T., 1986. The analysis of competition by field experiments. In: Kikkawa, J., Anderson, D.J. (Eds.),
Community Ecology: Pattern and Process, Blackwell Scientific, Oxford, pp. 240–268.
Wight, K., Francis, L., Eldridge, D., 1990. Food aversion learning by the hermit crab Pagurus granosimanus. Biol. Bull. 178, 205–209.