POLYMERASE PADA VIRUS DENGUE SECARA IN SILICO
WAHYU RONGGO WIMAG BIAN 0305037055
UNIVERSITAS INDONESIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM DEPARTEMEN KIMIA
DEPOK
2009
POLYMERASE PADA VIRUS DENGUE SECARA IN SILICO
Skripsi diajukan sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
WAHYU RONGGO WIMAG BIAN 0305037055
UNIVERSITAS INDONESIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM DEPARTEMEN KIMIA
DEPOK
2009
POLYMERASE PADA VIRUS DENGUE SECARA IN SILICO
NAMA : WAHYU RONGGO WIMAG BIAN
NPM : 0305037055
SKRIPSI INI TELAH DIPERIKSA DAN DISETUJUI DEPOK, 7 JULI 2009
PROF. DR. USMAN SUMO FRIEND TAMBUNAN, M.Sc PEMBIMBING
Tanggal lulus ujian sarjana : ... Penguji I : Prof. Dr. Soleh Kosela,.M.Sc... Penguji II : Dra. Sri Handayani,.M.Biomed... Penguji III : Dr. Amarila Malik, Apt,.M.Si...
i
Abadi Yang Maha Baik, yang selama ini telah mencurahkan segala kasih sayang, anugerah serta rahmat-Nya yang tak terkira terhadap penulis, serta pelajaran hidup yang membuat penulis mencoba terus belajar untuk mengerti tentang makna hidup, agar penulis menjadi manusia yang lebih baik.
Skripsi ini didedikasikan untuk Bapak dan Ibu. Terima kasih untuk segala kasih sayang,dorongan, nasehat, dan pengorbanan yang telah diberikan kepada penulis selama ini. Tak lupa penulis ucapkan terima kasih kepada saudara penulis, Febri, yang memberikan dorongan dan semangat selama ini.
Penulis ingin mengucapkan terima kasih kepada Prof. Dr. Usman.S.F. Tambunan, selaku pembimbing penelitian yang telah meluangkan waktunya tidak hanya membimbing penulis mengenai penelitian, namun juga
mengajarkan tentang kemampuan soft skill, dan pelajaran hidup. Mohon maaf jika selama ini banyak mengecewakan, tapi yang pasti penulis mencoba untuk selalu melakukan yang terbaik yang penulis bisa. Tak lupa penulis ingin mengucapkan terima kasih yang sebesar-besarnya kepada:
1. Dr. Ridla Bakri, selaku Ketua Departemen Kimia UI
2. Dra. Sri Wardhani, M.Si dan Ir. Widyastuti Samadi, selaku pembimbing akademik, yang telah membimbing penulis dalam bidang akademik selama perkuliahan
ii
hidup kepada penulis
5. Mba Fitri Amelia atas diskusinya
6. Sahabat penulis yang selalu mendukung, menyemangati, dan
memberi hiburan di kala susah: Santi dan Simas (semangat Simas!!!) 7. Rekan selama penelitian: Danang, Ramdhan, dan khususnya Samira
yang ikut jatuh bangun dan merasakan terang gelapnya penelitian, terima kasih banyak
8. Teman-teman 2005 serta teman-teman penelitian lantai 3 dan 4, khususnya Lusi,Angel,Purnama, Lulu, Meli, Lila,Faroq, dan Yusni. 9. Catherine dan Desti Far’05 atas diskusi dan penjelasannya
10. Seluruh karyawan Departemen Kimia,khususnya Babe,Mba Ina, dan Mba Cucu
11. Adik-adik angkatan 2006, 2007, dan 2008, kakak-kakak angkatan 2003 dan 2004
Terakhir, terima kasih yang sebesar-besarnya kepada semua pihak yang membantu penulis selama ini, baik langsung maupun tidak langsung, yang namanya tidak dapat disebut satu-persatu,TERIMA KASIH SEMUA
Semoga penelitian ini bermanfaat di kemudian hari. Amin.
Penulis 2009
Penyakit demam berdarah dengue yang disebabkan virus dengue
merupakan penyakit yang menjadi risiko pada negara-negara di daerah tropis dan subtropis, dengan kejadian tiap tahunnya mencapai 100 juta kasus. Sampai saat ini belum ditemukan adanya vaksin yang dapat mencegah terjadinya infeksi. Oleh karena itu diperlukan suatu upaya untuk menemukan obat berupa inhibitor yang dapat menghambat enzim-enzim yang berperan pada replikasi virus dengue, salah satunya enzim RNA-dependent RNA
polymerase (RdRp), yang berperan dalam penggandaan RNA virus dengue.
Peptida dipilih menjadi inhibitor yang potensial karena memiliki spesifitas dan aktivitas yang tinggi. Untuk meningkatkan kestabilan, peptida dirancang siklik dengan adanya jembatan disulfida. Peptida yang dirancang menggunakan kombinasi aspartat dan glutamat. Berdasar hasil docking diketahui bahwa peptida siklik dengan kombinasi residu CDEEC mempunyai nilai energi ikat yang terendah, yaitu sekitar -10,04 kkal/mol dan nilai Ki sebesar 43,44 nM yang mengindikasikan konformasi terstabil ligan-enzim, serta memiliki kontak dengan residu enzim dengan jumlah terbanyak yaitu 13 residu.
Kata kunci : Virus Dengue, Enzim RdRp, Inhibitor, Peptida Siklik, Docking ix+63 hlm; gbr; tab; lamp.
Bibliografi; 33 (1991-2009)
iii
v
KATA PENGANTAR………...i
ABSTRAK……….iii
DAFTAR ISI ………v
DAFTAR GAMBAR ………vii
DAFTAR TABEL……….viii
DAFTAR LAMPIRAN………..ix
BAB I. PENDAHULUAN……….1
1.1 Latar Belakang ………1
1.2 Tujuan………...3
BAB II. TINJAUAN PUSTAKA………...5
2.1 Sejarah Dengue………..5
2.2 Virus Dengue………...6
2.3 Daur Hidup Virus Dengue………...9
2.4 Enzim RNA-dependent RNA polymerase……….10
2.5 Inhibitor Enzim……….14
2.6 Peptida……….17
2.7 Bioinformatika………....19 2.7.1 Peran Bioinformatika dalam Proses Pencarian Obat…19
vi
2.10 Docking……….22
BAB III. METODE PENELITIAN……….23
3.1 Pencarian Data Enzim RdRp Virus Dengue Protein Data Bank (PDB) ……….23
3.2 Preparasi Ligan Peptida………24
3.3 Preparasi Enzim RdRp………..24
3.4 Docking Ligan Peptida dengan Enzim………...25
3.5 Analisis Hasil Docking………25
BAB IV. HASIL DAN PEMBAHASAN……….27
4.1 Pencarian Data Enzim RdRp Virus Dengue………..27
4.2 Preparasi Ligan Peptida………29
4.3 Analisis Hasil Docking………30
BAB V. KESIMPULAN DAN SARAN……….41
5.1. Kesimpulan ………..41
5.2. Saran ……….42
DAFTAR PUSTAKA………..43
LAMPIRAN………..47
vii
1. Morfologi virus dengue………..7
2. Pemetaan genom dari virus dengue………...8
3. Daur hidup virus dengue………...9
4. Struktur enzim RdRp virus dengue……….12
5. Mekanisme katalisis………..13
6. Mekanisme inisiasi………13
7. Interaksi jembatan garam ligan peptida DEE dengan residu Asp-663………37
8. Letak ikatan ligan peptida DEE pada binding site……….38
viii
1. Tipe inhibitor reversibel………...16 2. Kelebihan dan kekurangan peptida sebagai obat………....18 3. Tabel data energi terendah, jumlah populasi dalam cluster dengan
populasi terbanyak dan cluster rank dari masing-masing ligan uji……….31
4. Tabel data hasil docking………....33 5. Tabel residu kontak dengan ligan uji………..35
ix
1. Bagan kerja………..47
2. Daftar asam amino dan kodenya……….48
3. Hasil RMSD antara protein 2J7U dan 2J7W………..49
4. Hasil docking………...50
5. Residu kontak………..61
1.1 Latar Belakang
Penyakit demam berdarah dengue merupakan masalah kesehatan penting di lebih besar 100 negara tropis dan subtropis dengan risiko pada 2 miliar penduduknya. Diperkirakan kurang lebih 100 juta kasus dari DF dan 500.000 kasus DHF terjadi tiap tahunnya. Dengue Hemmoragic Fever (DHF) menyebabkan penyakit dan kematian pada anak-anak di negara- negara Asia Tenggara selama dua dekade terakhir. Indonesia sendiri merupakan salah satu negara yang keseluruhan daerahnya merupakan daerah dengan epidemi dengue (Raekiansyah, et al, 2004).
Sebagian infeksi virus dengue menunjukkan gejala demam ringan yang disebut juga sebagai dengue fever (DF), sedang beberapa pasien menunjukkan tingkat keparahan yang berlanjut dan secara potensial dapat membahayakan kesehatan. Tingkat selanjutnya disebut dengan dengue
hemorrhagic fever (DHF) yang dikarakterisasikan dengan pecahnya
pembuluh kapiler darah dan thrombocytopenia. Pada kasus yang lebih ekstrem, pasien dapat mengalami shock hipovolemia dan disebut dengan
dengue shock syndrome (DSS) dengan tingkat kematian 5-10 % per kasus
(Raekiansyah, et al, 2004)
1
Sampai saat ini belum ditemukan adanya vaksin virus dengue guna mencegah terjadinya infeksi virus dengue, meski demikian beberapa kandidat vaksin sedang memasuki uji klinis. Terdapat kesulitan dalam pengembangan vaksin virus dengue dikarenakan mekanisme patogenasitasnya yang dikenal dengan antibody-dependent enhancement (ADE) (Raekiansyah, et al, 2004).
Kebanyakan infeksi virus akan menginduksi antibodi yang bisa
menahan tubuh terhadap serangan virus berikutnya. Hal ini berbeda dengan virus dengue, infeksi pertama (primary infection) malah mempermudah tubuh untuk mendapat serangan berikutnya (secondary infection). Serangan
berikutnya menimbulkan gejala yang lebih berat dan fatal, karena antibodi yang dibuat pada serangan pertama dapat membantu infeksi virus pada serangan selanjutnya (Raekiansyah, et al, 2004).
Oleh karena itu, diperlukan suatu alternatif pengobatan lain yang dapat menghambat perkembangan infeksi virus dengue. Diperlukan usaha pengembangan obat antiviral melalui penemuan inhibitor bagi enzim yang berperan dalam perkembangbiakan virus seperti enzim protease, helikase, RNA polimerase, dan lain-lain (Selisko,et al, 2006).
Peptida dilaporkan dapat dijadikan suatu inhibitor yang potensial dan nantinya dapat dikembangkan menjadi obat, dikarenakan mempunyai beberapa keunggulan, antara lain : memiliki aktivitas dan spesifitas yang tinggi, tidak terakumulasi pada organ, toksisitas yang rendah, minim interaksi dengan obat lain, dan tidak menghasilkan respon imunologi (Huther &
Dietrich, 2007).
Proses penemuan inhibitor pada metode konvensional dilakukan dengan cara menguji senyawa-senyawa yang dianggap potensial terhadap enzim yang akan dihambat. Kemudian dilakukan proses penyeleksian dari senyawa-senyawa tersebut sampai didapatkan senyawa yang benar-benar dapat memberikan efek inhibisi, sedangkan senyawa yang tidak memberikan efek inhibisi, akan dieliminasi. Proses tersebut sangatlah tidak efektif karena boros waktu, tenaga, dan dana. Proses pencarian inhibitor dengan bantuan komputer dapat membantu untuk mereduksi jumlah senyawa yang diusulkan secara rasional dan lebih efektif serta membantu mempelajari interaksi obat dengan targetnya bahkan kemungkinan sifat toksis senyawa tersebut dan metabolitnya (Larson, 2006).
Berdasarkan hal-hal tersebut di atas akan dilakukan proses
perancangan inhibitor berupa peptida dari salah satu enzim di atas secara in
silico.
1.2 Tujuan Penelitian
Tujuan penelitian ini adalah merancang inhibitor berupa peptida yang tepat bagi enzim RNA-dependent RNA polymerase (RdRp) pada virus dengue secara in silico.
2.1. Sejarah Dengue
Demam Berdarah Dengue ( DBD ) merupakan salah satu penyakit menular yang disebabkan oleh infeksi virus dengue. Penyakit ini sering menimbulkan wabah dan menyebabkan kematian terutama pada anak, khususnya di daerah tropis dan subtropis (Hiswani, 2003). Vektor pembantu penyebaran virus dengue di Indonesia ialah nyamuk Aedes aegypti dan mungkin juga Aedes alboictus, kedua jenis nyamuk ini terdapat hampir diseluruh pelosok Indonesia, kecuali ditempat-tempat yang mempunyai ketinggian lebih dari 1000 meter diatas permukaan laut (Hiswani, 2003).
Epidemi dengue pertama kali ditemukan pada tahun 1779-1780 di Asia, Afrika, dan Amerika Utara. Selama masa tersebut demam berdarah dengue dikategorikan sebagai penyakit yang tidak berbahaya karena hanya
menyebabkan demam ringan. Baru setelah 10 - 40 tahun berikutnya, penyakit ini dikategorikan menjadi sangat berbahaya, karena memakan korban jiwa. Penjelasan yang mungkin dari fenomena ini adalah karena serotipe virus dengue muncul secara bertahap. Di Asia Tenggara, epideminya muncul setelah Perang Dunia II dan kemudian menyebar ke seluruh dunia. Epidemi DHF pertama kali muncul pada tahun 1950-an. Pada tahun 1975 DHF
5
merupakan penyebab kematian anak-anak di berbagai negara Asia Tenggara (www.cdc.gov, 9 Februari 2009, 12.40). Kasus di Indonesia pertama kali dilaporkan terjadi di Surabaya dan Jakarta dengan jumlah kematian sebanyak 24 orang. Beberapa tahun kemudian penyakit ini menyebar ke beberapa propinsi di Indonesia
2.2. Virus Dengue
Virus dengue merupakan bagian dari genus flavivirus dari famili
flaviviridae (Burke et al,2005). Virus dengue merupakan jenis virus RNA untai
tunggal yang terdiri dari 4 serotipe yaitu DEN-1, DEN-2, DEN-3, dan DEN-4, yang struktur keempatnya sangat mirip satu sama lain namun antibodi terhadap masing-masing serotipe tidak dapat saling memberikan
perlindungan (Lindenbach, 2001). Bentuk morfologi dari virion virus dengue adalah bola (spherical) dengan diameter 40 - 60 nm
(http://athena.bioc.uvic.ca/, 9 Februari 2009, 12.48). Virus dengue merupakan jenis virus RNA (+), oleh karena itu dapat bermutasi lebih cepat karena RNA polimerase memiliki tingkat kesalahan lebih besar daripada DNA polimerase, yaitu terdapat satu kesalahan tiap 104 nukleotida yang ditranskripsi, sedang
pada transkripsi DNA hanya terdapat satu kesalahan tiap 106 nukleotida yang
digandakan oleh DNA polimerase (Alberts, et al, 1994).
Gambar 1. Morfologi virus dengue
Sumber: [http://biomarker.cdc.go.kr]
Genom virus dengue terdiri dari rantai tunggal RNA yang mempunyai berat molekul 11 Kb tersusun dari protein struktural dan protein non-struktural. Protein struktural terdiri dari protein core atau nukleokapsid (C), protein
envelope (E), dan protein pre-membran (prM atau preM). Protein struktural ini merupakan 25% dari total protein, sedangkan protein non-struktural virus terdiri dari tujuh macam yang dikode oleh gen terpisah. Protein tersebut
adalah : NS1, NS2a, NS2b, NS3, NS4a, NS4b, dan NS5 (Putnak, et al, 1990).
Gambar 2. Pemetaan genom dari virus dengue
Sumber: [http://carnot.utmb.edu]
Protein struktural merupakan protein yang berperan sebagai morfologi / bentuk dari virus, sedangkan protein non-struktural berperan sebagai enzim-enzim yang berperan dalam proses replikasi virus.
2.3. Daur Hidup Virus dengue
Gambar 3. Daur hidup virus dengue
Sumber: [www.dsimb.inserm.fr]
Virus dengue mempunyai genom RNA (+) sense dan bereplikasi pada sitoplasma sel inang. Genomnya menyerupai mRNA selular dalam segala aspek kecuali tidak adanya poly-adenilate (poly-A) tail. Hal ini menyebabkan
virus dapat menggunakan organel-organel dari sel inang untuk memproduksi protein-proteinnya, baik struktural maupun non-struktural. Ribosom sel inang mengambil peran penting untuk mensintesis poliprotein dari virus mengingat bahwa RNA virus menyerupai mRNA selular (Qi, et al, 2007)
Ketika ditranslasi, poliprotein yang terbentuk akan dipotong-potong oleh protease baik dari virus maupun dari inang, menghasilkan
produk-produk protein yang mempunyai fungsi masing-masing. Selain itu, poliprotein mempunyai aktifitas autokatalitik yang secara otomatis dapat menghasilkan peptida pertama, enzim virus yang spesifik. Enzim tersebut kemudian memotong bagian-bagian poliprotein yang lain. Salah satunya yaitu
RNA-dependent RNA polymerase yang berperan dalam sintesis sense (-), yang
kemudian nantinya akan dijadikan sebagai cetakan bagi terbentuknya RNA virus lainnya. Kemudian bagian-bagian dari virus dirakit dan akhirnya karena jumlahnya makin banyak menyebabkan lisisnya sel inang. (Qi, et al, 2007)
2.4. Enzim RNA-dependent RNA polymerase (RdRp)
RNA dependent RNA polymerase (RdRp) adalah enzim yang
mengkatalisis proses replikasi RNA dari template RNA dan berbeda dengan RNA polimerase, yang menggunakan DNA sebagai template-nya.
(http://encyclopedia.freedictionary.com, 8 Juni 2009, 10.50).
Protein NS5 merupakan protein terbesar dengan berat molekul mencapai 104kDa, memiliki kesamaan sekuens hampir 67 % di antara 4
serotipe, dan bertindak dwifungsi sebagai RNA- dependent RNA polymerase
dan metil transferase yang berperan dalam replikasi RNA virus. Protein NS5
memiliki kurang lebih 900 asam amino, dimana enzim S-adenosil metionin transferase terletak pada ujung N-terminal-nya, dan protein RdRp terletak pada residu 270 – 900. Replikasi dimulai dengan sintesis RNA strand ( - ) dari genom RNA ( + ) virus dengue, kemudian strand ( - ) tersebut dijadikan
template bagi terbentuknya genom RNA ( + ) virus dengue lainnya (Yap,et al,
2007).
Enzim RdRp terdiri dari beberapa bagian (domain), antara lain palm,
fingers, dan thumb yang antar bagiannya saling terhubung, sehingga
morfologi molekulnya terlihat tersusun rapat dan kompak, berbeda dengan DNA polimerase yang lebih terbuka (kurang tersusun rapat/kompak). Mempunyai bentuk seperti huruf U dan memiliki karakteristik right-handed (Ferrer, 2006).
Gambar 4. Struktur enzim RdRp virus dengue
Sumber: [Yap,et al, 2007]
Semua enzim RdRp dalam mekanisme katalisisnya melibatkan dua residu asam aspartat, dan dua ion logam divalen untuk pembentukan ikatan fosfodiester dengan substrat nukleotida (Joyce&Steitz, 1995 ; Steitz, 1998). Mempunyai dua jenis mekanisme inisiasi, antara lain inisiasi de novo
(primer-independent) dan inisiasi primer-dependent (Van Dijk, et al, 2004)
Gambar 5. Mekanisme katalisis
Sumber: [Steitz, 1998]
Gambar 6. Mekanisme inisiasi
Sumber: [van Dijk, 2004]
Yap, et al (2007) dalam jurnalnya menjelaskan bahwa terdapat tiga
residu asam amino yang amat berperan dalam proses inisiasi replikasi RNA virus dengan menggunakan mekanisme de novo, yaitu Ser-710, Arg- 729, dan Arg-737, dan ketiga residu ini keberadaannya pada enzim RdRp tidak berubah (conserved) baik pada tiap serotipe dari virus dengue (DEN1 - 4), namun juga pada enzim RdRp virus-virus famili flaviviridae lainnya misalnya
Yellow Fever Virus, West Nile Virus, Hepatitis C Virus, dan BVDV.
Berdasarkan uji mutasi yang telah dilakukan sebelumnya, yaitu dilakukan perubahan asam amino tunggal terhadap salah satu dari ketiga residu tersebut, didapatkan hasil bahwa proses sintesis RNA virus secara de
novo hampir tidak terjadi sama sekali, sedang untuk proses elongasi
berkurang kurang lebih dua sampai sembilan kali. Berdasarkan hasil-hasil tersebut, ketiga residu tersebut dapat dijadikan situs sasaran potensial penambatan/pelekatan ligan peptida pada enzim RdRp (Yap, et al, 2007).
2.5. Inhibitor Enzim
Inhibitor merupakan senyawa yang dapat menghambat kerja dari enzim. Inhibitor terbagi atas dua jenis, yaitu pertama yang menyebabkan inaktifasi enzim secara irreversibel (inhibitor irreversibel) dan yang kedua menyebabkan inhibisi secara reversibel (inhibitor reversibel)
(www.themedicalbiochemistrypage.org, 8 Juni 2009, 15.13)
Inhibitor irreversibel biasanya menyebabkan inaktifasi enzim,
dikarenakan adanya interaksi kovalen. Efek kinetik dari inhibitor irreversibel adalah menurunkan konsentrasi dari enzim aktif, sehingga kemungkinan tercapainya konsentrasi maksimum komplek enzim-substrat makin menurun. Inhibitor irreversibel biasanya bersifat toksik dan tidak dianjurkan untuk terapetik/ pengobatan (www.themedicalbiochemistrypage.org, 8 Juni 2009, 15.16)
Inhibitor reversibel terbagi atas tiga kategori, yaitu, inhibitor kompetitif, inhibitor non-kompetitif, dan inhibitor un-kompetitif. Biasanya inhibitor terikat secara non-kovalen, misalnya ikatan hidrogen, interaksi hidrofobik, dan ikatan ionik. Inhibitor dan enzim mempertahankan suatu kesetimbangan reversibel, sehingga mempunyai nilai konstanta kesetimbangan disosiasi dari komplek
enzim inhibitor yang disebut Ki (www.themedicalbiochemistrypage.org, 8 Juni
2009, 15.35).
E + I ↔ EI
Tabel 1. Tipe inhibitor reversibel
Tipe Inhibitor
Binding Site pada Enzim
Inhibitor Kompetitif
Berada pada sisi aktif secara spesifik, dimana terjadi persaingan dengan substrat. Inhibisi dapat dikembalikan dengan substrat.
Inhibitor Non-kompetitif
Berikatan dengan E atau ES pada tempat lain selain sisi aktif. Substrat tidak terganggu
ikatannya, tapi komplek ESI tidak menghasilkan produk. Inhibisi tidak dapat dikembalikan dengan substrat.
Inhibitor Un- Kompetitif
Hanya bisa berikatan dengan komplek ES pada tempat lain selain sisi aktif. Pelekatan substrat menyebabkan perubahan struktur enzim, sehingga inhibitor dapat terikat.
2.6. Peptida
Peptida adalah molekul organik / asam amino yang terikat secara kimia dengan ikatan peptida. Ikatan peptida selalu terdiri dari ikatan kovalen tunggal antara α-karboksil dari asam amino satu dengan gugus amino nitrogen dari asam amino kedua. Pada proses pembentukan ikatan peptida pada dua asam amino, akan melepaskan satu molekul air. Peptida kecil yang terdiri kurang dari 10 residu asam amino dinamakan oligopeptida, sedang yang terdiri lebih dari 10 residu asam amino dinamakan polipeptida
(Columbia Encyclopedia).
Didasarkan pada spesifisitas dan aktivitasnya dalam tubuh, peptida banyak dikembangkan sebagai molekul obat. Lebih dari 140 peptida telah digunakan pada saat ini sebagai obat, dan kurang lebih 400 lagi saat ini masuk dalam tahap pra-klinik, dengan pertumbuhan rata-rata tahunan mencapai 15%. Oleh karena itu peptida dapat dikembangkan sebagai obat, salah satunya dapat dijadikan sebagai inhibitor pada virus, contohnya VIRIP, yang merupakan peptida inhibitor pada HIV dengan mekanisme menghambat protein enveloped dari virus (Huther, 2007).
Berikut merupakan kelebihan dan kekurangan peptida jika digunakan sebagai molekul obat :
Tabel 2. Kelebihan dan kekurangan peptida sebagai obat
Kelebihan Kekurangan Ukurannya kecil
Aktivitas tinggi Spesifisitas tinggi
Tidak menimbulkan efek imunologi Toksisitas rendah
Diversitas tinggi
Minim interaksi dengan obat lain
Stabilitas rendah dalam tubuh Bioavailibilitas oral rendah Administrasi injeksi
Penghantaran ke sel yang kurang
Sumber: [Huther dan Dietrich, 2007]
Salah satu cara yang dapat dilakukan untuk meningkatkan kestabilan dari senyawa peptida adalah dengan melakukan siklisasi. Salah satu cara siklisasi peptida adalah dengan menambahkan residu sistein pada ujung-ujung peptida, yang nantinya gugus tiol dari sistein akan membentuk jembatan disulfida. Adanya jembatan disulfida pada peptida dapat memberikan kestabilan tambahan, karena ikatan disulfida dapat
mendestabilisasi bentuk unfolded (terbuka) dari peptida dengan menurunkan entropinya dan dapat membentuk interaksi hidrofobik (Futami, et al, 2000).
2.7. Bioinformatika
Bionformatika adalah suatu cabang ilmu yang menggabungkan ilmu komputer dan biologi. Bioinformatika meliputi pengelolaan informasi biologis yang diperoleh dari berbagai penelitian yang menghasilkan data dalam jumlah banyak dan komplek, seperti pemetaan genom manusia.
Bioinformatika memberikan prediksi maupun simulasi dengan
mempertimbangkan hubungan serta pola data biologis. Bioinformatika adalah suatu bidang ilmu baru yang menggunakan pendekatan komputasional untuk menyelesaikan persoalan biologis (Baxevanis dan Oullette, 2001).
2.7.1 Bioinformatika dalam Proses Pencarian Obat (Drug Discovery)
Komputer yang dilengkapi dengan aplikasi kimia komputasi yang memadai dapat menayangkan senyawa A secara tiga dimensi (3D) dan melakukan komparasi dengan senyawa lain yang sudah diketahui memiliki aktivitas tinggi, misal senyawa B. Berdasarkan komparasi 3D dilengkapi dengan perhitungan similaritas dan energi, memberikan gambaran bagian-bagian dan gugus-gugus potensial yang dapat dikembangkan dari senyawa A (pharmacophore query). Kemudian berbagai senyawa turunan dan analog disintesis secara in silico atau digambar sesuai persyaratan aplikasi komputer yang digunakan (untuk selanjutnya disebut senyawa hipotetik). Keberadaan data struktur 3D enzim X akan sangat membantu. Aplikasi komputer dapat melakukan studi interaksi antara senyawa-senyawa hipotetik dengan enzim X
secara in silico pula. Berdasar dari studi ini dapat diprediksi aktivitas
senyawa-senyawa hipotetik dan dapat dilakukan eliminasi senyawa-senyawa yang memiliki aktivitas rendah. Kemudian dari beberapa langkah in silico tersebut, dapat diusulkan beberapa senyawa analog dan turunan senyawa A yang memang potensial untuk disintesis dan dikembangkan, atau
mengusulkan untuk mengembangkan seri baru. Jumlah senyawa yang diusulkan biasanya jauh lebih sedikit dibandingkan penemuan obat secara konvensional. Dalam hal ini komputer membantu untuk mereduksi jumlah senyawa yang diusulkan secara rasional dan diharapkan lebih efektif serta, membantu mempelajari interaksi obat dengan targetnya (Galdenhuys, 2006).
2.8. Molecular Modelling
Molecular modelling adalah suatu teknik untuk memperoleh,
menampilkan, dan memanipulasi struktur, reaksi dari suatu molekul, serta sifat-sifat yang tergantung dari struktur tiga dimensi dari molekul tersebut. Teknik ini menggabungkan kimia komputasi, biologi komputasi, dan ilmu material dalam mempelajari sistem molekular dari sistem kimia yang kecil sampai molekul biologi yang besar dan material. Dewasa ini terdapat tiga jenis penggunaan yang paling sering digunakan dalam molecular modelling, yaitu analisis struktur, homology modelling,dan docking (www.ch.ic.ac.uk, diambil tanggal 4 Juni 2009, jam 12.08) .
2.9. Minimisasi
Merupakan proses pencarian konformasi yang ideal dari suatu molekul berdasarkan mekanika molekular, yang merupakan metode kalkulasi dari struktur molekul, energi konformasi, dan sifat-sifat molekular lainnya dengan menggunakan konsep mekanika klasik (Naz, et al, 2009). Sebuah molekul diibaratkan sebagai kumpulan atom yang disatukan oleh gaya energi klasik. Gaya-gaya ini dideskripsikan sebagai fungsi energi potensial dari segi struktural seperti panjang ikatan, sudut ikatan, dan sudut torsi. Energi dari suatu molekul dihitung sebagai penjumlahan dari suatu persamaan sebagai berikut :
E = E stretching + E bending + E torsion + E Vander Waals + E electrostatic + E hydrogen bond
+ cross term
Kumpulan dari fungsi serta parameter yang sesuai disebut sebagai force field (Naz, et al, 2009).
2.10. Docking
Docking merupakan suatu metode yang dapat memprediksikan
orientasi dari suatu molekul ke molekul yang lain ketika berikatan dalam membentuk komplek yang stabil (Rarey, 1996). Pengetahuan tentang orientasi tersebut dapat dijadikan sebagai prediksi tentang kekuatan atau
afinitas ikatan di antara dua molekul menggunakan fungsi scoring (Rarey, 1996).
Docking banyak digunakan untuk memprediksi orientasi ikatan antara
kandidat molekul kecil obat dengan protein target dalam rangka memprediksi afinitas dan aktifitas dari miolekul kecil tersebut. Oleh karena itu, docking mempunyai peran yang penting dalam desain obat baru (Bajorath, 2004)
Docking dapat diartikan sebagai solusi pemecahan tentang teori “lock
and key” , dimana dicari orientasi yang tepat dari kunci (key) yang dapat
membuka gembok (lock). Protein dapat dijadikan sebagai lock dan ligan / molekul obat dapat diartikan sebagai key. Docking mencerminkan proses optimasi dari orientasi yang sesuai antara ligan yang berikatan dengan protein target (Jorgensen, 1991). Untuk menjalankan docking, hal pertama yang perlu dibutuhkan adalah struktur dari protein target, yang biasanya ditentukan menggunakan teknik biofisika seperti kristalografi sinar-X atau spektroskopi NMR. Struktur protein ini dan database dari ligan yang potensial dijadikan input dalam program docking. Keberhasilan dari proses docking tergantung dari 2 komponen yaitu algoritma dan fungsi scoring (Bajorath, 2004).
3.1 Pencarian Data Enzim RdRp Virus dengue Protein Data Bank (PDB)
Pencarian struktur enzim RdRp dari Protein Data Bank (PDB) melalui situs Research Collaboratory for Structural Bioinformatics (RCSB) dengan alamat http://www.rcsb.org/pdb/ dengan menggunakan perangkat komputer yang terhubung dengan internet. Sistem operasi yang digunakan adalah
Microsoft Windows XP dengan browser Internet Explorer. Format data yang
digunakan adalah format PDB. Setelah struktur tersier protein RdRp virus dengue didapat, kemudian dilakukan analisis untuk menentukan binding site dari protein tersebut. Penentuan binding site dilakukan dengan
menggunakan software online molecular modelling. Input data dalam
program ini menggunakan format PDB dengan memasukkan PDB code atau PDB file
3.2 Preparasi Ligan Peptida
Ligan peptida yang akan diuji pada penelitian ini digambar secara 3 dimensi dengan menggunakan program ACDLabs. Peptida dimodelkan
23
sebagai peptida siklik, dengan masing-masing ujungnya ditambahkan residu sistein yang nantinya dapat membentuk jembatan disulfida. Asam amino yang akan dimodelkan adalah asama amino bermuatan negatif, yaitu asam glutamat dan asam aspartat. Peptida dimodelkan dengan deprotonasi pada gugus karboksilat, dan protonasi pada gugus amino. Gambar disimpan dalam format MDL Mol file.
NH3+- Cys- AAA –Cys- COO-
S - S
Proses optimasi dilakukan dengan menggunakan program VegaZZ.
Force field yang digunakan untuk ligan peptida adalah TRIPOS dan charges
Gasteigger. Pencarian konformasi dilakukan dengan metode steepest
descent dan conjugate gradient.
3.3. Preparasi Enzim RdRp
Molekul air, ion klorin, dan tryethylene glycol sebelumnya dihilangkan dengan menggunakan program PyMol.
Kemudian dilakukan optimasi dengan force field CHARMM22_PROT, dengan metode steepest descent dan conjugate gradient melalui program
VegaZZ. Hasil optimasi disimpan dalam format PDB file.
3.4. Docking antara ligan peptida dengan enzim
Berkas (file) parameter docking disiapkan dengan menggunakan
AutoDock Tools. Untuk molekul enzim, atom hidrogen polar ditambahkan dan
atom hidrogen non-polar disatukan, dan ditambahkan muatan Kollman. Untuk ligan, ditambahkan muatan Gasteiger, atom hidrogen non-polar disatukan, semua ikatan dibuat dapat berotasi. Penghitungan docking dilakukan dengan menggunakan program AutoDock4.0 menggunakan Lamarckian Genetic
Algorithm (LGA). Parameter yang digunakan antara lain; population sizes 150,
energy evaluations 2500000, dan dilakukan run sebanyak 50. Grid box
disiapkan dengan grid spacing sebesar 0,375Å. Nilai RMSD dari tiap cluster diatur tidak melebihi 1,0 Å.
3.5 Analisis Hasil Docking terhadap Kompleks Kandidat Peptida-Enzim RdRp
Analisis Docking dilakukan dengan menganalisis konformasi dengan nilai energi terendah dari cluster yang paling banyak mempunyai populasi. Kemudian dianalisis nilai ΔGbinding dan nilai Ki (konstanta inhibitor) antara
peptida-enzim. Mencari interaksi yang terjadi, dengan menganalisis ikatan hidrogen antara peptida dan enzim, serta analisis residu-residu enzim yang mangalami kontak dengan ligan peptida.
4.1. Pencarian Data Enzim RdRp Virus dengue Protein Data Bank (PDB)
Data struktur 3D protein dari enzim RdRp pada virus dengue dicari melalui Protein Data Bank melalui situs http://www.rcsb.org/pdb/, yang
dikelola oleh Research Collaboratory for Structural Biology. Proses pencarian dapat dilakukan baik dengan mengetikkan kata kunci atau PDB ID pada kolom search. PDB ID terdiri dari empat karakter, gabungan baik dari bilangan numerikal maupun alfabetikal. Pada file PDB berisi tentang nama protein, nama peneliti yang melakukan penelitian terhadap struktur kristal dari protein, jurnal dimana hasil penelitian ini dipublikasikan, metode yang
digunakan saat kristalisasi, geometri dari struktur, rincian sekuens, serta ligan yang terdapat pada struktur.
Proses pencarian dilakukan dengan melakukan pengetikkan kata kunci, sehingga didapatkan hasil 2 struktur enzim RdRp dari virus dengue, yang mempunyai kode PDB-ID 2J7U dan 2J7W. Perbedaan secara fisik antara 2 struktur tersebut terdapat pada ada tidaknya ligan pada masing-masing protein tersebut. Pada protein 2J7U tidak terdapat ligan, sedang pada 2J7W terdapat ligan yaitu 3'DGTP (3’-Deoxy Guanosine Triphospate), yang
diketahui dari jurnal merupakan suatu inhibitor dari enzim RdRp pada proses
27
elongasi. Perbedaan lainnya terletak pada resolusi kristalisasi, kristal 2J7U mempunyai nilai resolusi yang lebih kecil daripada krisatal 2J7W. Nilai resolusi sendiri mengacu pada kepadatan elektron. Pada kristalografi suatu makromolekul seringkali melibatkan ribuan atom dalam sel unit. Kristal makromolekul biasanya sulit untuk dibedakan antar atomnya, dengan kata lain biasanya atom-atom dan ikatan kimia biasanya terlihat masih bergabung, dan bukan terlihat sebagai atom-atom yang berdiri sendiri. Berdasarkan hal tersebut, dipilihlah kristal yang mempunyai resolusi yang terkecil dengan alasan bahwa makin kecil resolusi suatu kristal, maka tingkat kejelasan pemisahan dari atom-atom dalam suatu molekul makin terlihat jelas. Hal lainnya yang membedakan adalah adanya logam Mg pada kristal 2J7U yang berperan dalam proses katalisis, sedang pada kristal 2J7W tidak terdapat logam Mg.
Protein dengan PDB-ID 2J7U, strukturnya ditentukan dengan
menggunakan difraksi sinar-X dengan resolusi 1,85 Å oleh Yap, T.L., dkk dan dipublikasikan pada tanggal 13 Maret 2007. Dilihat dari susunan asam
aminonya, diketahui bahwa tidak terdapat asam amino yang hilang pada tengah rantai susunan asam aminonya, atau rantai proteinnya tidak ada yang terputus di tengah.
Selain itu dilakukan pula superimpose/tumpang tindih struktur enzim yang dipilih (2J7U) dengan struktur enzim lain pada Protein Data Bank yaitu 2J7W dengan menggunakan program PyMol, dan didapatkan hasil RMSD = 0,365. RMSD merupakan parameter kemiripan antara 2 struktur, makin kecil
nilainya, maka makin besar kemiripan strukturnya. Menurut Baxevanis dan Oullette yang menerangkan hubungan antara nilai RMSD dan hubungan antar struktur, jika nilai RMSD antar 2 struktur kurang dari 0.4, maka antara 2 struktur tersebut dapat dikatakan “essentially indistinguishable” atau dengan kata lain tidak dapat dibedakan satu sama lainnya.
4.2 Preparasi Ligan Peptida
Peptida yang akan dirancang susunan asam amino- asam aminonya bermuatan negatif /bersifat asam, yaitu asam aspartat (D) dan asam glutamat (E). Hal tersebut dikarenakan dengan merancang peptida yang terdiri dari asam amino-asam amino bermuatan negatif diharapkan nantinya dapat membentuk interaksi yang kuat antara ligan peptida dengan residu-residu penting pada enzim RdRp, antara lain Ser-710, Arg-729, dan Arg-737, yang diketahui lebih bersifat positif.
Pemilihan hanya 3 asam amino yang terdapat pada ligan peptida didasarkan bahwa jika asam amino yang terdapat dalam ligan terlalu banyak akan dapat membuat strukturnya menjadi lebih besar sehingga dapat
mempersulit melewati jalur paraselular.
Kombinasi dari asam aspartat dan asam glutamat yang digunakan dalam rancangan peptida menghasilkan ligan peptida siklik sebanyak 8 buah yang berbeda letak serta komposisi dari amino amino-asam amino
penyusunnya. Ligan-ligan peptida siklik yang dihasilkan dari kombinasi
tersebut antara lain: CDDDC, CDDEC, CDEDC, CEDDC, CDEEC, CEDEC, CEEDC, dan CEEEC.
Pemodelan ligan peptida dilakukan dengan memprotonasi gugus amino dan mendeprotonasi gugus karboksil pada ligan peptida. Hal ini dilakukan karena pada pH fisiologis gugus karboksil hampir seluruhnya
berada dalam bentuk R-COO-, sedang gugus amino berada pada bentuk R-
NH3+ . Rantai samping gugus karboksil pada residu asam aspartat dan asam
glutamat juga dideprotonasi, dikarenakan asam aspartat dan asam glutamat adalah asam amino bersifat asam yang diketahui bahwa titik isoelektrik-nya berada pada suasana asam sekitar pH 3, sedang pada pH 6-8 bentuk rantai samping gugus karboksilnya berada pada bentuk terdeprotonasi.
4.3 Analisis hasil docking
Proses docking/penambatan dilakukan sebanyak 50 kali untuk tiap-tiap ligan peptida, sehingga nantinya akan ada 50 bentuk konformasi berbeda ketika ligan peptida berikatan dengan enzim. Kenyataannya, tidak selalu terbentuk 50 konformasi ligan peptida yang berbeda-beda dan ada kalanya terdapat bentuk konformasi yang sama. Program AutoDock nantinya akan mengelompokkan konformasi-konformasi yang sama dalam sutu kelompok yang disebut sebagai cluster.
Tabel 3. Tabel data energi terendah, jumlah populasi dalam cluster dengan
populasi terbanyak dan cluster rank dari masing-masing ligan uji
Peptida ΔGbinding terendah dalam
cluster dengan populasi
terbanyak (kkal/mol)
Jumlah populasi dalam satu cluster*
Cluster Rank CEEEC - 9,03 5 3 CEEDC - 8,77 4 1 CEDEC -8,96 4 1 CDEEC -10,04 5 1 CDDEC -7,45 5 7 CDEDC -8,93 5 1 CEDDC -7,77 6 10 CDDDC -8,35 16 1
*) dengan jumlah populasi terbanyak dalam satu cluster
Pemilihan konformasi ligan peptida didasarkan banyaknya populasi dalam suatu cluster, jika dalam suatu cluster memiliki populasi (konformasi ligan yang sama) dengan jumlah yang paling banyak, maka bisa diartikan konformasi dalam cluster tersebut merupakan konformasi yang lebih ‘disenangi’ oleh ligan ketika berikatan dengan binding site. Hal tersebut dikarenakan dalam beberapa kali proses docking, konformasi itulah yang
berulangkali terjadi ketika berikatan dengan binding site, atau penyebaran bentuk konformasi ligan tertuju pada konformasi tersebut.
Cluster dengan jumlah populasi terbanyak seharusnya berada pada
cluster rank pertama. Cluster rank menunjukkan cluster/kelompok konformasi
ligan yang memiliki nilai ΔGbinding terendah selama beberapa kali proses
docking. Nilai ΔGbinding yang rendah mengindikasikan bahwa ligan peptida
berada pada konformasi yang paling stabil ketika berikatan dengan enzim (peluang terjadinya sekitar 80 – 90 %). Sehingga, jika cluster dengan jumlah populasi terbanyak berada pada cluster rank pertama, dapat diartikan bahwa konformasi ligan ketika berikatan dengan enzim merupakan konformasi yang paling stabil, dan hal ini diperkuat dengan beberapa kali proses docking, konformasi tersebut selalu dituju oleh ligan tersebut.
Berdasarkan data di atas, lebih dari sebagian ligan memenuhi hipotesis tentang konformasi terstabil ligan ketika berikatan dengan enzim (memiliki cluster dengan populasi terbanyak pada cluster rank pertama). Ada beberapa ligan yang tidak memenuhi hipotesis tersebut, sehingga konformasi dari ligan-ligan tersebut meragukan apakah benar-benar konformasi tersebut merupakan konformasi terstabil dari ligan tersebut ketika berikatan dengan
binding site. Dari data di atas, ligan-ligan yang memenuhi kriteria antara lain
CEEDC,CEDEC, CDEEC, CDEDC, dan CDDDC.
Setelah dilakukan pemilihan ligan-ligan mana saja yang akan dipilih berdasarkan konformasi terstabilnya, lalu dilakukan analisis nilai energi
berdasarkan hasil docking. Berikut merupakan data nilai energi hasil docking dari ligan-ligan yang dipilih:
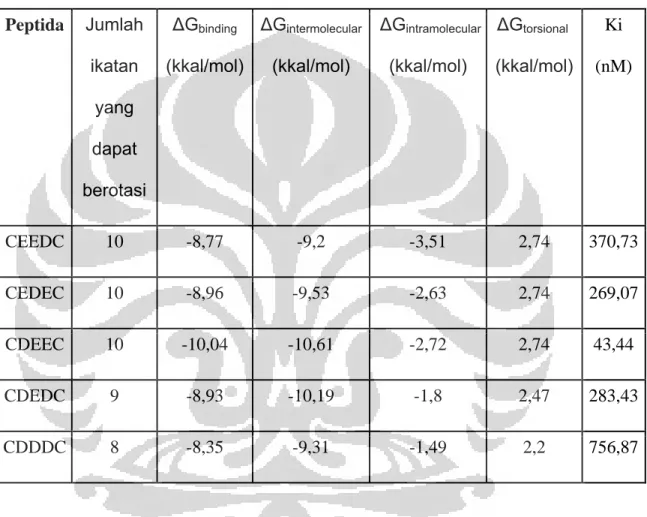
Tabel 4. Tabel data hasil docking Peptida Jumlah ikatan yang dapat berotasi ΔGbinding (kkal/mol) ΔGintermolecular (kkal/mol) ΔGintramolecular (kkal/mol) ΔGtorsional (kkal/mol) Ki (nM) CEEDC 10 -8,77 -9,2 -3,51 2,74 370,73 CEDEC 10 -8,96 -9,53 -2,63 2,74 269,07 CDEEC 10 -10,04 -10,61 -2,72 2,74 43,44 CDEDC 9 -8,93 -10,19 -1,8 2,47 283,43 CDDDC 8 -8,35 -9,31 -1,49 2,2 756,87
Berdasarkan data di atas dapat dilihat bahwa makin kecil jumlah ikatan yang dapat berotasi maka nilai ΔGtorsional juga makin berkurang. Pada ligan
yang mempunyai jumlah ikatan yang dapat berotasi sama maka nilai ΔGtorsional juga sama besar. Dari data di atas jika jumlah ikatan yang dapat
berotasi berkurang sebanyak 1 ikatan, maka nilai ΔGtorsional akan berkurang
dengan nilai konstan yaitu ~ 0,27 kkal/mol seiring dengan pengurangan
gugus metilen. Hal tersebut dapat diketahui dikarenakan perbedaan jumlah ikatan yang dapat berotasi berasal dari perbedaan jumlah antara residu asam amino aspartat dan asam glutamat pada satu peptida, seperti telah diketahui bahwa perbedaan mendasar antara asam glutamat dan asam aspartat terletak pada rantai sampingnya, yaitu asam glutamat memiliki rantai C yang lebih panjang (beda satu gugus metilen).
Nilai ΔGintramolecular dipengaruhi oleh panjang ikatan, sudut ikatan, dan
sudut dihedral pada molekul ligan. Berdasarkan data di atas, terdapat
kecenderungan bahwa nilai ΔGintramolecular makin besar (mendekati positif) jika
jumlah ikatan yang dapat berotasi makin kecil, walaupun tidak naik secara linier. Hal tersebut kemungkinan disebabkan berkurangnya perputaran ikatan pada ligan yang dapat menyebabkan peluang terjadinya perubahan jarak ikatan, sudut ikatan, atau sudut dihedral semakin berkurang, sehingga menyebabkan nilai ΔGintramolecular menjadi lebih besar (mendekati positif)
walaupun tidak secara signifikan.
Nilai konstanta inhibisi dari kelima ligan masuk dalam kisaran nM, mengindikasikan terbentuknya komplek ligan-enzim yang stabil, dikarenakan nilai Ki diperoleh dari persamaan : [E][I] / [EI], pada rekasi kesetimbangan pembentukan komplek enzim.
E + I ↔ EI
Sehingga makin kecil nilai Ki, maka reaksi lebih cenderung menuju pembentukan komplek EI daripada penguraian komplek EI. Berdasarkan data
hasil docking juga diketahui bahwa makin rendah nilai ΔGbinding, makin kecil
nilai Ki.
Selanjutnya dicari residu pada enzim yang mengalami kontak dengan ligan dengan menggunakan program Chimera, didapatkan hasil sebagai berikut:
Tabel 5. Residu kontak dengan ligan uji
Peptida Residu kontak
(residu dengan huruf tebal membentuk ikatan hidrogen dengan ligan)
CEEDC Tyr-606, Ser-710, Arg-729, Arg-737, Thr-794, Trp-795,
Ser-796, Ile-797
CEDEC Tyr-606, Cys-709, ser-710, His-711, Arg-729, Arg-737, Thr-794, Trp-795, Ser-796, Ile-797, His-798
CDEEC Leu-511, Ser-661, Asp-663, Asp-664, Cys-709, Ser-710,
His-711, Arg-729, Arg-737, Thr-794, Ser-796, Ile-797, His-798 CDEDC 710, His-711, Arg-729, Arg-737, Thr-794, Trp-795,
Ser-796, Ile-797, His-798
CDDDC Ser-661, Asp-663, Asp-664, Cys-709, Ser-710, His-711,
Arg-729, Arg-737, Ser-796, Ile-797, His-798
Berdasarkan hasil pencarian residu yang mengalami kontak dengan masing-masing ligan, diketahui bahwa kelima ligan mempunyai kontak
dengan 3 residu yang penting pada binding site, yaitu Ser-710, Arg-729, dan Arg-737. Hal ini menandakan bahwa hasil docking ligan peptida sesuai dengan yang diharapkan.
Diketahui berdasarkan hasil docking bahwa ligan CDEEC mempunyai nilai ΔGbinding terendah di antara ligan-ligan lainnya, selain itu pula diketahui
mempunyai kontak dengan jumlah residu yang banyak yaitu dengan 13 residu, di antaranya dengan Asp-663 dan Asp-664 yang merupakan residu yang terdapat pada catalytic site, sehingga diharapkan dapat mengganggu proses katalisis.
Peptida CDEEC juga mempunyai interaksi ikatan hidrogen dengan lima residu seperti yang tersebut di atas, di antaranya berikatan hidrogen dengan dua residu yang dianggap penting, yaitu Arg-729 dan Arg-737.
Selain mempunyai interaksi ikatan hidrogen dengan residu-residu yang tersebut di atas, ligan peptida CDEEC diketahui juga memiliki interaksi
jembatan garam dengan residu Asp-663, dimana ujung terminal N (-NH3+)
membentuk jembatan garam dengan rantai samping gugus COO- dari residu
Asp-663. Interaksi jembatan garam dianggap penting bagi sumbangsihnya pada nilai ΔGintermolecular, dikarenakan nilai stabilisasi dari interaksi jembatan
garam nilainya sama besarnya dengan ikatan hidrogen.
Gambar 7. Interaksi jembatan garam antara ligan peptida dengan residu
Asp-663.
Berdasarkan hasil visualisasi hasil docking, diketahui bahwa ligan peptida CDEEC berikatan dengan enzim RdRp pada suatu lubang yang terdapat pada enzim RdRp. Lubang tersebut diketahui merupakan jalur
masuknya RNA virus ketika ingin melakukan proses inisiasi dan elongasi atau biasa disebut NTP tunnel.
Gambar 8. Letak ikatan ligan peptida CDEEC pada binding site
Berdasarkan hasil docking dapat disimpulkan bahwa ligan peptida
siklik dengan kombinasi CDEEC (Cys-Asp-Glu-Glu-Cys) dapat dijadikan suatu inhibitor yang potensial guna menghambat aktivitas enzim RdRp, dikarenakan beberapa hal, antara lain:
1. Memiliki nilai energi ikat yang rendah (paling rendah di antara ligan peptida uji lainnya) ketika berikatan dengan enzim RdRp, yaitu sekitar -10,04 kkal/mol.
2. Mempunyai nilai Ki pada kisaran nM (43,44 nM), mengindikasikan terbentuknya komplek ligan peptida-enzim yang stabil
3. Memiliki kontak dengan residu yang terbanyak dibandingkan dengan peptida uji lainnya, diantaranya kontak dengan Asp-663 dan Asp-664 yang terdapat pada catalytic site, serta dengan Ser-710, Arg-729, dan
Arg-737, yang ketiganya diketahui berpengaruh terhadap proses inisiasi RNA virus
4. Memiliki ikatan hidrogen beberapa residu, diantarnya dengan residu penting yaitu Arg-729 dan Arg-737, serta membentuk interaksi jembatan garam dengan Asp-663.
5.1. Kesimpulan
Berdasarkan hasil docking dapat disimpulkan bahwa ligan peptida
siklik CDEEC dapat dijadikan suatu inhibitor yang potensial guna
menghambat aktivitas enzim RdRp, dikarenakan beberapa hal, antara lain memiliki nilai ΔGbinding yang rendah (paling rendah di antara ligan peptida uji
lainnya), yaitu sekitar -10,04 kkal/mol dan nilai Ki pada kisaran 43,44 nM, dan dari nilai-nilai tersebut mengindikasikan terbentuknya komplek ligan peptida-enzim yang stabil. Selain itu, ligan peptida siklik CDEEC memiliki kontak dengan residu yang terbanyak dibandingkan dengan peptida uji lainnya, diantaranya kontak dengan Asp-663 dan Asp-664 yang terdapat pada
catalytic site, serta dengan Ser-710, Arg-729, dan Arg-737, yang ketiganya
diketahui berpengaruh terhadap proses inisiasi RNA virus. Ligan peptida siklik CDEEC sendiri mempunyai ikatan hidrogen dengan beberapa residu penting seperti Arg-729 dan Arg-737, serta membentuk interaksi jembatan garam dengan residu Asp-663. Lain dari itu, berdasarkan visualisasi hasil
docking, diketahui bahwa letak terikatnya ligan peptida siklik CDEEC pada
enzim RdRp berada dalam NTP tunnel.
41
5.2. Saran
Diperlukan studi lanjutan dengan menggunakan metode simulasi dinamika molekular, dengan menyertakan keberadaan pelarut yang berfungsi untuk mengetahui apakah ikatan-ikatan antara enzim dengan ligan masih bertahan, jika ditambahkan adanya efek pelarut.
Alberts, B., A Jonson, J.Lewis, M. Raff, K.Roberts, P.Walter.2002. Molecular
Biology of the Cell.ed. ke-4,Garlang Science, New York.
Baxevanis, A.D, B.F.F. Oullete.2001. Bioinformatics, A Practical Guide to the
Analysis Genes and Protein. ed. ke-2, John Wiley & Sons, Inc, USA
Burke, D.S, T.P Monath. 2001. Fields Virology .ed ke-4, Lipincott Williams &Wilkins, Philadelphia
Christopher Smith .1998. Molecular Modelling - Seeing the Whole Picture with Modeling Software Packages.Scientist 12[17]:0, Aug. 31, 1998
Ferrer-Orta, C., A. Arias, C. Escarmis, N. Verdaguer. 2006. A Comparison of Viral RNA-dependent RNA polymerases, Current Opinion of
Structural Biology. 16:27-34.
Futami J, Tada H, Seno M, Ishikami S, Yamada H. 2000. Stabilization of Human RNase 1 by Introduction of a Disulfide Bond Between Residues 4 and 118. Journal of Biochemistry .128 (2): 245–50
Geldenhuys, W,J., et al. 2006. Optimizing the Use of Open-Source Software Applications in Drug Discovery. Drug Discovery Today (3/4): 127-132 Henchal, Erik A., J. Robert Putnak. 1990. The Dengue Virus. Clinical
Microbiology Reviews. Vol.3 No.4. p.376-396..
Hendra.2006. Karya Sarjana Utama Kimia: Analisis Rancangan vaksin Tetravalen Dengue Virus secara In Silico Menggunakan Protein E
43
DENV-3 Sebagai Backbone Melalui Homology Modelling. Departemen Kimia FMIPA –UI
Hidayah, Zulfa.2005. Karya sarjana Uatama Kimia : Studi Awal Bioinformatika dalam Perancangan Vaksin Dengue Tetravalen. Departemen Kimia FMIPA-UI
Hiswani. 2003. Pencegahan dan Pemberantasan Demam Berdarah Dengue. USU Digital Library
Huther, Alexandra & Ursula Dietrich. 2007. The Emergence of Peptides as Therapeutics Drug for Inhibition of HIV-1. AIDS Review. 9:208-17 Jorgensen WL .1991. Rusting of the Lock and Key Model for Protein-Ligand
Binding. Science 254 (5034): 954–5
Kitchen DB, Decornez H, Furr JR, Bajorath J. 2004. Docking and Scoring in Virtual Screening for Drug Discovery: Methods and Applications.
Nature Reviews :Drug discovery 3 (11): 935–49
Larson, Richard.S.2006. Bioinformatics and Drug Discovery.Humana Press, New Jersey
Lengauer T, Rarey M .1996. Computational Methods for Biomolecular Docking. Current Opinion of Structural Biology. 6 (3): 402–6
Naz, Afshan, Khalida Bano, Farhat Bano, Najaf Abbas, Naheed Akhtar.2009. Conformational Analysis (Geometry Optimization) of Nucleosidic Anti-Tumor Antibiotic Showdomycin by Arguslab 4 Software. Pakistan
Journal of Pharmaceutical Sciences., Vol.22, pp.78-82
Patrick, Graham.L.2001. Introduction to Medicinal Chemistry. Oxford University Press
Qi,Rui-Feng, Ling Zhang, Cheng Wu Chi. 2007. Biological Characteristics of Dengue Virus and Potential Targets for Drug Design. Acta
Biochimica et Biophysica Sinica.2008; 40: 91-101
Raekiansyah,Muhareva, T.Mirawati Sudiro. 2004. Genetic Variation Among Dengue Virus that Correlate with Pathogenesis. Medical Journal of
Indonesia;13:190-4
Rao, Gitta Subba, Manoj .V Ramachandran, J.S. Bajaj.2006.In Silico Structure-Based Design of Potent and Selective Small Peptide
Inhibitor of Protein Tyrosine Phosphatase 1B. Journal of Biomolecular
Structure and Dynamic:23(4)
Selisko, Barbara, Jean-Claude Guillemot, Karine Alvarez, Bruno Canard. 2006. Opportunities in Development of Anti-Dengue Drug. Working
Paper for the Scientific Working Group on Dengue Research,
convened by the Special Programme for Research and
Training in Tropical Diseases
Steitz, T.A & Steitz, J.A.1993. A General Two Metal Ion Mechanism for Catalytic RNA. Proceeding of National Academy of Science ; 90,6498-6052
Taufik,Rizky Ichsan. 2006. Karya Sarjana Utama Kimia: Analisis Rancangan Vaksin Tetravalen Dengue Virus secara In Silico Menggunakan
Protein E DENV-2 Sebagai Backbone Melalui Homology Modelling. Departemen Kimia FMIPA –UI
Van Dijk, Alberdina, Eugene .V Makeyev, Dennis H Bamford.2004. Initiation of Viral RNA-dependent RNA Polymerase Polimerization. Journal of
General Virology; 85:1077-1093
Yap,Thai Leong, Ting Xu, Yen-Liang Chen, Helene Malet, Marie-Pierre Egloff, Bruna Cannard, Subash G Vasudevan, Julian Lescar. 2007. Crystal Structure of Dengue Virus RNA-dependent RNA Polymerase
Catalytic Domain at 1.85 A Resolition.Journal of Virology; 81(9):4753-4765 http://athena.bioc.uvic.ca/. http://carnot.utmb.edu http://encyclopedia.freedictionary.com www.cdc.gov www.ch.ic.ac.uk www.dsimb.inserm.fr www.themedicalbiochemistrypage.or
Pencarian Data Enzim RdRp Virus dengue Protein Data Bank (PDB)
Preparasi Ligan Peptida Preparasi enzim RdRp
Docking antara ligan peptida dengan enzim RdRp
Analisis hasil docking
RMSD = 0,365
EEE CLUSTERING HISTOGRAM ____________________ _______________________________________________________________________ _________ | | | | | Clus | Lowest | Run | Mean | Num | Histogram -ter | Binding | | Binding | in | Rank | Energy | | Energy | Clus| 5 10 15 20 25
30 35 _____|___________|_____|___________|_____|____:____|____:____|____:____ |____:___ 1 | -9.52 | 33 | -9.50 | 2 |## 2 | -9.14 | 10 | -9.14 | 1 |# 3 | -9.03 | 6 | -8.68 | 5 |##### 4 | -8.57 | 21 | -8.35 | 2 |## 5 | -8.55 | 39 | -8.37 | 2 |## 6 | -8.46 | 46 | -8.46 | 1 |# 7 | -8.38 | 40 | -8.38 | 1 |# 8 | -8.10 | 35 | -8.10 | 1 |# 9 | -8.04 | 7 | -7.32 | 2 |## 10 | -8.04 | 31 | -7.96 | 2 |## 11 | -7.89 | 11 | -7.89 | 1 |# 12 | -7.88 | 8 | -7.88 | 1 |# 13 | -7.74 | 36 | -7.74 | 1 |# 14 | -7.73 | 14 | -7.65 | 2 |## 15 | -7.71 | 45 | -7.71 | 1 |# 16 | -7.71 | 12 | -7.71 | 1 |# 17 | -7.65 | 44 | -7.65 | 1 |# 18 | -7.60 | 50 | -7.60 | 1 |# 19 | -7.58 | 24 | -7.58 | 1 |# 20 | -7.47 | 4 | -7.47 | 1 |# 21 | -7.43 | 47 | -7.43 | 1 |# 22 | -7.37 | 15 | -7.37 | 1 |# 23 | -7.11 | 16 | -7.11 | 1 |# 24 | -7.09 | 48 | -7.09 | 1 |# 25 | -6.95 | 41 | -6.95 | 1 |# 26 | -6.95 | 17 | -6.95 | 1 |# 27 | -6.91 | 20 | -6.91 | 1 |# 28 | -6.86 | 38 | -6.86 | 1 |# 29 | -6.85 | 34 | -6.85 | 1 |# 30 | -6.79 | 2 | -6.79 | 1 |# 31 | -6.74 | 13 | -6.74 | 1 |# 32 | -6.64 | 19 | -6.64 | 1 |# 33 | -6.63 | 42 | -6.63 | 1 |# 34 | -6.57 | 3 | -6.57 | 1 |# 35 | -6.46 | 28 | -6.46 | 1 |# 36 | -6.41 | 43 | -6.41 | 1 |# 37 | -6.37 | 1 | -6.37 | 1 |# 38 | -6.27 | 9 | -6.27 | 1 |#
40 | -6.08 | 18 | -6.08 | 1 |#
_____|___________|_____|___________|_____|_____________________________ _________
Number of multi-member conformational clusters found = 7, out of 50 runs.
Run = 6
Cluster Rank = 3
Number of conformations in this cluster = 5
RMSD from reference structure = 65.479 A
Estimated Free Energy of Binding = -9.03 kcal/mol [=(1)+(2)+(3)-(4)]
Estimated Inhibition Constant, Ki = 241.59 nM (nanomolar) [Temperature = 298.15 K]
USER (1) Final Intermolecular Energy = -9.33 kcal/mol USER vdW + Hbond + desolv Energy = -6.23 kcal/mol USER Electrostatic Energy = -3.10 kcal/mol USER (2) Final Total Internal Energy = -4.57 kcal/mol USER (3) Torsional Free Energy = +3.02 kcal/mol USER (4) Unbound System's Energy = -1.86 kcal/mol
DEE CLUSTERING HISTOGRAM ____________________ _______________________________________________________________________ _________ | | | | | Clus | Lowest | Run | Mean | Num | Histogram -ter | Binding | | Binding | in | Rank | Energy | | Energy | Clus| 5 10 15 20 25
30 35 _____|___________|_____|___________|_____|____:____|____:____|____:____ |____:___ 1 | -10.04 | 21 | -9.61 | 5 |##### 2 | -9.15 | 47 | -8.75 | 4 |#### 3 | -9.13 | 40 | -9.13 | 1 |# 4 | -8.95 | 24 | -8.95 | 1 |# 5 | -8.83 | 15 | -8.83 | 1 |# 6 | -8.57 | 45 | -8.57 | 1 |# 7 | -8.57 | 23 | -8.39 | 2 |## 8 | -8.42 | 8 | -8.42 | 1 |# 9 | -8.28 | 18 | -8.28 | 1 |# 10 | -8.26 | 41 | -8.02 | 4 |#### 11 | -8.00 | 1 | -8.00 | 1 |# 12 | -7.89 | 36 | -7.89 | 1 |# 13 | -7.78 | 9 | -7.78 | 1 |# 14 | -7.72 | 34 | -7.72 | 1 |#
16 | -7.64 | 49 | -7.64 | 1 |# 17 | -7.62 | 33 | -7.62 | 1 |# 18 | -7.60 | 35 | -7.60 | 1 |# 19 | -7.57 | 27 | -7.57 | 1 |# 20 | -7.56 | 11 | -7.56 | 1 |# 21 | -7.56 | 46 | -7.56 | 1 |# 22 | -7.54 | 19 | -7.54 | 1 |# 23 | -7.47 | 16 | -7.47 | 1 |# 24 | -7.46 | 50 | -7.46 | 1 |# 25 | -7.46 | 5 | -7.46 | 1 |# 26 | -7.46 | 44 | -7.46 | 1 |# 27 | -7.45 | 25 | -7.45 | 1 |# 28 | -7.27 | 14 | -7.27 | 1 |# 29 | -7.26 | 39 | -7.26 | 1 |# 30 | -7.04 | 3 | -7.04 | 1 |# 31 | -7.03 | 37 | -7.03 | 1 |# 32 | -6.77 | 31 | -6.77 | 1 |# 33 | -6.76 | 7 | -6.76 | 1 |# 34 | -6.74 | 2 | -6.74 | 1 |# 35 | -6.71 | 42 | -6.71 | 1 |# 36 | -6.68 | 6 | -6.68 | 1 |# 37 | -6.55 | 17 | -6.55 | 1 |# 38 | -6.48 | 43 | -6.48 | 1 |# 39 | -6.43 | 29 | -6.43 | 1 |# _____|___________|_____|___________|_____|_____________________________ _________
Number of multi-member conformational clusters found = 4, out of 50 runs.
Run = 21
USER Cluster Rank = 1
USER Number of conformations in this cluster = 5 USER
USER RMSD from reference structure = 63.792 A USER
USER Estimated Free Energy of Binding = -10.04 kcal/mol [=(1)+(2)+(3)-(4)]
USER Estimated Inhibition Constant, Ki = 43.44 nM (nanomolar) [Temperature = 298.15 K]
USER
USER (1) Final Intermolecular Energy = -10.61 kcal/mol USER vdW + Hbond + desolv Energy = -6.54 kcal/mol USER Electrostatic Energy = -4.07 kcal/mol USER (2) Final Total Internal Energy = -2.72 kcal/mol USER (3) Torsional Free Energy = +2.74 kcal/mol USER (4) Unbound System's Energy = -0.55 kcal/mol
EDE
CLUSTERING HISTOGRAM ____________________
_________
| | | | | Clus | Lowest | Run | Mean | Num | Histogram -ter | Binding | | Binding | in | Rank | Energy | | Energy | Clus| 5 10 15 20 25
30 35 _____|___________|_____|___________|_____|____:____|____:____|____:____ |____:___ 1 | -8.96 | 17 | -8.76 | 4 |#### 2 | -8.76 | 44 | -8.58 | 2 |## 3 | -8.61 | 35 | -8.44 | 3 |### 4 | -8.59 | 42 | -8.37 | 2 |## 5 | -8.57 | 3 | -8.57 | 1 |# 6 | -8.55 | 43 | -8.26 | 4 |#### 7 | -8.50 | 10 | -8.50 | 1 |# 8 | -8.45 | 49 | -8.07 | 3 |### 9 | -8.42 | 26 | -8.42 | 1 |# 10 | -8.19 | 12 | -8.19 | 1 |# 11 | -8.19 | 32 | -8.19 | 1 |# 12 | -8.17 | 6 | -8.17 | 1 |# 13 | -8.16 | 18 | -8.07 | 2 |## 14 | -8.14 | 8 | -8.14 | 1 |# 15 | -8.06 | 45 | -8.06 | 1 |# 16 | -8.04 | 16 | -8.04 | 1 |# 17 | -7.87 | 7 | -7.87 | 1 |# 18 | -7.82 | 20 | -7.53 | 2 |## 19 | -7.46 | 38 | -7.46 | 1 |# 20 | -7.41 | 23 | -7.41 | 1 |# 21 | -7.40 | 22 | -7.40 | 1 |# 22 | -7.28 | 36 | -7.28 | 1 |# 23 | -7.24 | 27 | -7.24 | 1 |# 24 | -7.09 | 15 | -7.09 | 1 |# 25 | -7.08 | 29 | -7.08 | 1 |# 26 | -7.03 | 2 | -7.03 | 1 |# 27 | -6.86 | 48 | -6.86 | 1 |# 28 | -6.82 | 21 | -6.82 | 1 |# 29 | -6.78 | 4 | -6.78 | 1 |# 30 | -6.77 | 47 | -6.77 | 1 |# 31 | -6.72 | 34 | -6.72 | 1 |# 32 | -6.66 | 50 | -6.66 | 1 |# 33 | -6.62 | 39 | -6.62 | 1 |# 34 | -6.61 | 40 | -6.61 | 1 |# 35 | -6.17 | 41 | -6.17 | 1 |# 36 | -5.96 | 33 | -5.96 | 1 |# _____|___________|_____|___________|_____|_____________________________ _________
Number of multi-member conformational clusters found = 8, out of 50 runs.
Run = 17
USER Cluster Rank = 1
USER Number of conformations in this cluster = 4 USER
USER RMSD from reference structure = 66.091 A
USER Estimated Free Energy of Binding = -8.96 kcal/mol [=(1)+(2)+(3)-(4)]
USER Estimated Inhibition Constant, Ki = 269.07 nM (nanomolar) [Temperature = 298.15 K]
USER
USER (1) Final Intermolecular Energy = -9.53 kcal/mol USER vdW + Hbond + desolv Energy = -5.99 kcal/mol USER Electrostatic Energy = -3.54 kcal/mol USER (2) Final Total Internal Energy = -2.63 kcal/mol USER (3) Torsional Free Energy = +2.74 kcal/mol USER (4) Unbound System's Energy = -0.46 kcal/mol
EED CLUSTERING HISTOGRAM ____________________ _______________________________________________________________________ _________ | | | | | Clus | Lowest | Run | Mean | Num | Histogram -ter | Binding | | Binding | in | Rank | Energy | | Energy | Clus| 5 10 15 20 25
30 35 _____|___________|_____|___________|_____|____:____|____:____|____:____ |____:___ 1 | -8.77 | 11 | -8.27 | 4 |#### 2 | -8.45 | 20 | -8.06 | 2 |## 3 | -8.42 | 13 | -8.23 | 2 |## 4 | -8.37 | 33 | -8.02 | 2 |## 5 | -8.30 | 44 | -8.22 | 2 |## 6 | -8.21 | 38 | -8.21 | 1 |# 7 | -7.95 | 27 | -7.95 | 1 |# 8 | -7.93 | 4 | -7.93 | 2 |## 9 | -7.92 | 49 | -7.92 | 1 |# 10 | -7.89 | 39 | -7.89 | 1 |# 11 | -7.87 | 28 | -7.87 | 1 |# 12 | -7.86 | 31 | -7.81 | 2 |## 13 | -7.86 | 12 | -7.86 | 1 |# 14 | -7.78 | 36 | -7.78 | 1 |# 15 | -7.75 | 32 | -7.75 | 1 |# 16 | -7.72 | 26 | -7.67 | 2 |## 17 | -7.71 | 43 | -7.71 | 1 |# 18 | -7.70 | 41 | -7.70 | 1 |# 19 | -7.55 | 17 | -7.55 | 1 |# 20 | -7.55 | 42 | -7.55 | 1 |# 21 | -7.52 | 29 | -7.13 | 2 |## 22 | -7.34 | 1 | -7.34 | 1 |# 23 | -7.34 | 16 | -7.34 | 1 |# 24 | -7.33 | 3 | -7.33 | 1 |# 25 | -7.21 | 40 | -7.21 | 1 |# 26 | -7.14 | 24 | -7.14 | 1 |# 27 | -7.07 | 47 | -7.07 | 1 |# 28 | -7.05 | 10 | -7.05 | 1 |#
30 | -7.01 | 22 | -7.01 | 1 |# 31 | -6.93 | 25 | -6.93 | 1 |# 32 | -6.92 | 19 | -6.92 | 1 |# 33 | -6.83 | 37 | -6.83 | 1 |# 34 | -6.81 | 45 | -6.81 | 1 |# 35 | -6.80 | 15 | -6.80 | 1 |# 36 | -6.75 | 23 | -6.75 | 1 |# 37 | -6.66 | 46 | -6.66 | 1 |# 38 | -6.61 | 6 | -6.61 | 1 |# 39 | -6.45 | 30 | -6.45 | 1 |# _____|___________|_____|___________|_____|_____________________________ _________
Number of multi-member conformational clusters found = 9, out of 50 runs.
USER Run = 11
USER Cluster Rank = 1
USER Number of conformations in this cluster = 4 USER
USER RMSD from reference structure = 67.341 A USER
USER Estimated Free Energy of Binding = -8.77 kcal/mol [=(1)+(2)+(3)-(4)]
USER Estimated Inhibition Constant, Ki = 370.73 nM (nanomolar) [Temperature = 298.15 K]
USER
USER (1) Final Intermolecular Energy = -9.20 kcal/mol USER vdW + Hbond + desolv Energy = -5.70 kcal/mol USER Electrostatic Energy = -3.51 kcal/mol USER (2) Final Total Internal Energy = -3.48 kcal/mol USER (3) Torsional Free Energy = +2.74 kcal/mol USER (4) Unbound System's Energy = -1.17 kcal/mol
DDE CLUSTERING HISTOGRAM ____________________ _______________________________________________________________________ _________ | | | | | Clus | Lowest | Run | Mean | Num | Histogram -ter | Binding | | Binding | in | Rank | Energy | | Energy | Clus| 5 10 15 20 25
30 35 _____|___________|_____|___________|_____|____:____|____:____|____:____ |____:___ 1 | -8.16 | 5 | -8.06 | 2 |## 2 | -8.08 | 7 | -8.04 | 4 |#### 3 | -7.82 | 14 | -7.82 | 1 |# 4 | -7.67 | 32 | -7.67 | 1 |# 5 | -7.57 | 11 | -7.57 | 1 |#
7 | -7.45 | 2 | -7.17 | 5 |##### 8 | -7.36 | 23 | -7.27 | 3 |### 9 | -7.34 | 1 | -7.34 | 1 |# 10 | -7.29 | 3 | -7.28 | 2 |## 11 | -7.28 | 15 | -7.20 | 2 |## 12 | -7.25 | 50 | -7.25 | 1 |# 13 | -7.23 | 19 | -7.22 | 2 |## 14 | -7.12 | 42 | -7.12 | 1 |# 15 | -7.04 | 10 | -7.04 | 1 |# 16 | -7.03 | 24 | -6.99 | 3 |### 17 | -7.03 | 17 | -7.03 | 1 |# 18 | -7.02 | 13 | -6.94 | 2 |## 19 | -6.98 | 35 | -6.94 | 2 |## 20 | -6.97 | 34 | -6.97 | 1 |# 21 | -6.97 | 45 | -6.97 | 1 |# 22 | -6.94 | 33 | -6.94 | 1 |# 23 | -6.88 | 43 | -6.88 | 1 |# 24 | -6.77 | 44 | -6.77 | 1 |# 25 | -6.71 | 31 | -6.71 | 1 |# 26 | -6.69 | 30 | -6.69 | 1 |# 27 | -6.66 | 8 | -6.66 | 1 |# 28 | -6.65 | 18 | -6.65 | 1 |# 29 | -6.65 | 6 | -6.65 | 1 |# 30 | -6.53 | 27 | -6.53 | 1 |# 31 | -6.51 | 47 | -6.51 | 1 |# 32 | -6.35 | 46 | -6.35 | 1 |# 33 | -5.99 | 38 | -5.99 | 1 |# _____|___________|_____|___________|_____|_____________________________ _________
Number of multi-member conformational clusters found = 10, out of 50 runs.
USER Run = 2
USER Cluster Rank = 7
USER Number of conformations in this cluster = 5 USER
USER RMSD from reference structure = 72.177 A USER
USER Estimated Free Energy of Binding = -7.45 kcal/mol [=(1)+(2)+(3)-(4)]
USER Estimated Inhibition Constant, Ki = 3.48 uM (micromolar) [Temperature = 298.15 K]
USER
USER (1) Final Intermolecular Energy = -7.55 kcal/mol USER vdW + Hbond + desolv Energy = -4.33 kcal/mol USER Electrostatic Energy = -3.22 kcal/mol USER (2) Final Total Internal Energy = -2.95 kcal/mol USER (3) Torsional Free Energy = +2.47 kcal/mol USER (4) Unbound System's Energy = -0.59 kcal/mol
DED
![Gambar 1. Morfologi virus dengue Sumber: [http://biomarker.cdc.go.kr]](https://thumb-ap.123doks.com/thumbv2/123dok/4253512.3129283/18.918.186.793.109.517/gambar-morfologi-virus-dengue-sumber-http-biomarker-cdc.webp)
![Gambar 2. Pemetaan genom dari virus dengue Sumber: [http://carnot.utmb.edu]](https://thumb-ap.123doks.com/thumbv2/123dok/4253512.3129283/19.918.125.757.131.846/gambar-pemetaan-genom-dari-virus-dengue-sumber-carnot.webp)
![Gambar 3. Daur hidup virus dengue Sumber: [www.dsimb.inserm.fr]](https://thumb-ap.123doks.com/thumbv2/123dok/4253512.3129283/20.918.170.810.208.864/gambar-daur-hidup-virus-dengue-sumber-dsimb-inserm.webp)
![Gambar 4. Struktur enzim RdRp virus dengue Sumber: [Yap,et al, 2007]](https://thumb-ap.123doks.com/thumbv2/123dok/4253512.3129283/23.918.183.728.117.644/gambar-struktur-enzim-rdrp-virus-dengue-sumber-yap.webp)
![Gambar 5. Mekanisme katalisis Sumber: [Steitz, 1998]](https://thumb-ap.123doks.com/thumbv2/123dok/4253512.3129283/24.918.190.762.121.897/gambar-mekanisme-katalisis-sumber-steitz.webp)