L
Journal of Experimental Marine Biology and Ecology 249 (2000) 77–88
www.elsevier.nl / locate / jembe
Pigment signatures associated with an anoxic coastal zone:
Bahia Concepcion, Gulf of California
*
´ ´ ´
J. Bustillos-Guzman , D. Lopez-Cortes, F. Hernandez, I. Murillo
Northwest Center for Biological Research, POB 128, La Paz, B.C.S. Mexico Received 14 April 1999; received in revised form 29 February 2000; accepted 2 March 2000
Abstract
Bahia Concepcion is a coastal lagoon that has bottom anoxic conditions and high pigment concentrations during the summer. The phytoplankton responsible for this pigment increase is enigmatic, therefore we sampled the lagoon to analyze the pigment with a C8-HPLC system to look for signatures of phytoplankton groups. Analysis reveals a low pigment concentration in the mixed layer with a higher concentration of zeaxanthin and increasing values of chlorophyll a, peridinin, and fucoxanthin below, which peaked at the depth where oxygen dramatically decreases and H S increases. Below this depth, a high pigmentation was recorded and the most important2 signatures were six chlorophyll-like pigments that eluted between the fucoxanthin and the chlorophyll a, and one carotenoid that eluted just after the chlorophyll a. Spectral characteristics of these last pigments are very similar to pigments present in the Chlorobiales group. These results suggest that cyanobacteria, diatoms, and dinoflagellates are responsible for the chlorophyll a increases, though in highly pigmented samples, anoxygenic phototrophic bacteria are probably the main contributors to the increase in pigments. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Bacteriochlorophylls; Coastal lagoons; Gulf of California; HPLC; Pigments
1. Introduction
Bahia Concepcion is one of the largest bays in the Gulf of California with about 400
2
km of surface and a maximum depth of 30 m (Lechuga-Deveze, 1994). Human influence is small because it is in a zone of low precipitation (between 150 and 250 mm per year), with practically no important towns (only small tourist settlements), and little maritime navigation. These characteristics allow the study of events (i.e. red tides,
*Corresponding author. Tel.: 133-112-536-33; fax: 133-112-536-25. ´
E-mail address: [email protected] (J. Bustillos-Guzman)
biogeochemical cycles, etc.) in an unperturbed environment. Phytoplankton in the area is dominated by diatoms, but red tide species are occasionally present (Martinez-Lopez and Garate-Lizarraga, 1994). For summer, Lechuga-Deveze (1994) reported bottom pigment
21
concentrations of chlorophyll a (Chl a) between 7.1 and 45.5 mg l and chlorophyll b 21
(Chl b) values between 10.4 and 99.1mg l in more than a half of the area bay. These values are high when compared with normal stations where Chl a and Chl b
21
concentration maximum values, in general, are lower than 2mg l . Although there are several reports on the phytoplankton composition and abundance (Kiefer and Laker, 1975; Gilmartin and Revelante, 1978; Martinez-Lopez and Garate-Lizarraga, 1994; Lopez-Cortez, 1999; Lechuga-Deveze and Morquecho-Escamilla, 1999; Lechuga-De-veze et al., 2000), these high pigment concentrations are enigmatic because no one has reported species or groups of species that can be related to such an increase of biomass. Recently, Lechuga-Deveze (1994) analyzed the spectra of acetone extracts and sug-gested a population of prochlorophytes may be responsible for the high biomass. His reason is that highly pigmented samples have red-shift spectra when compared with normally pigmented ones, similar to that existing between the Chl a and divinyl chlorophyll a (DvChl a). Because of the difficulties of identifying and quantifying this picoplanktonic group with traditional methods of microscopy, this hypothesis has not been tested.
We have used high-performance liquid chromatography (HPLC) to analyze the phytoplankton pigment and look for markers of phytoplankton groups in these samples. Specifically, we tested for the existence of DvChl a, a marker of prochlorophytes (Goericke and Repeta, 1993), or other phytoplankton group fingerprints that may be responsible for this increase of biomass.
2. Material and methods
In August 1997, we sampled at several stations throughout the bay (see Lechuga-Deveze, 1994) to detect the ‘anomalous’ highly pigmented area. Once the area was located, one station was selected (Fig. 1) as representative of the ‘anomalous’ area and a detailed profile (at 0, 2, 4, 6, 8, 10, 12, 13, 15, 17, 19, 21, 22, 25, 26 and 27-m depth) was obtained for pigment, temperature, nitrates, oxygen and H S analysis. Additionally,2
´
J. Bustillos-Guzman et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 77 –88 79
Fig. 1. Vertical variation of physical and chemical variables in Bahia Concepcion, Gulf of California: (A) temperature; (B) irradiance (I ) as a percentage of incident irradiance (I ), and dissolved oxygen (O ); (C)0 2
(1; 50, 50), (15; 0, 100), (18.5; 0, 100) and (19; 75, 25). Pigments were detected with the diode-array absorbance signal set at 440 nm. Identification was made by comparing retention time and spectral characteristics with those of commercial pigment standards
14
(International Agency for C determinations, Denmark) and a rich prochlorophyte sample collected at the deep chlorophyll maximum from the Mexican Pacific Ocean (19852.2 N, 113811.99W) during the USA METOX01 cruise for the DvChl a. Quantification was done with the pigment response factor (HPLC peak area / pigment mass) obtained with the commercial pigment standards according to Mantoura and Repeta (1997). Nitrate and dissolved oxygen were measured according to Strickland and Parson (1972), sulfide by the iodometric method (Clesceri et al., 1989), and temperature and salinity with a Kahlsico salinometer model 140.
3. Results
3.1. Physical and chemical conditions
Using the physical and chemical variables (Fig. 1), three zones can be distinguished: (i) a surface mixed layer (upper 12 m) with the highest temperature (32.58C) and
21
dissolved oxygen (.6 ml l ), and relatively low nitrate concentration (lower than 0.4 21
mg at l ); (ii) an intermediate layer with temperature between 32.5 and 298C, 21
decreasing dissolved oxygen (from 6.6 to 1.0 ml l ) and stable nitrate concentration 21
(0.4–0.42mg at l ); and (iii) a bottom layer with temperature decreasing to 228C, very 21
low or undetectable dissolved oxygen (,1.0 ml l ), and increasing nitrate and H S2
concentration. In this last layer, irradiance was between 4 and 10% of the incident irradiance at the surface [Fig. 1(B)].
3.2. Pigments
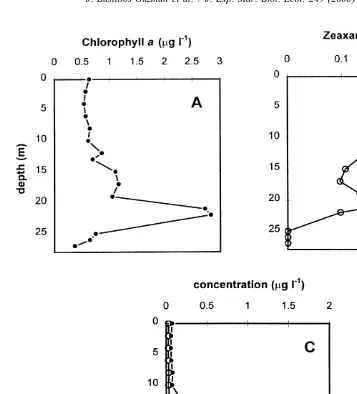
Peridinin and fucoxanthin, pigment signatures of dinoflagellates (Jeffrey, 1974) and of diatoms (Liaaen-Jensen, 1985; Wright and Jeffrey, 1987) had a low concentration in the mixed layer, a slight increase between 12 and 19 m, and a sharp increase at 21 m [Fig. 2(C)]. A second increase of peridinin occurs at the bottom. Zeaxanthin concentration, an indicator of cyanobacteria and prochlorophyte abundance (Guillard et al., 1985; Goericke and Repeta, 1993) is relatively low and homogeneous in the upper 20-m layer
21
´
J. Bustillos-Guzman et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 77 –88 81
Fig. 2. Vertical variation of pigment concentration in Bahia Concepcion, Gulf of California: (A) chlorophyll a; (B) zeaxanthin; (C) fucoxanthin and peridinin.
3.3. Unidentified pigments
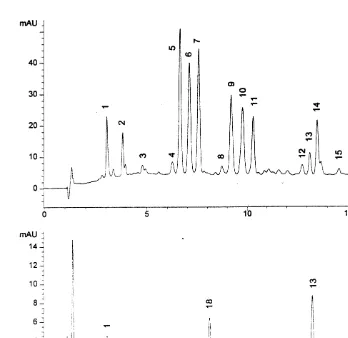
Fig. 3. Typical chromatogram (440 nm) from highly pigmented (A) and normal samples (B). Peak identification: 1, chlorophyll c1 and c2; 2, peridinin; 3, fucoxanthin-like pigment; 4, diadinoxanthin; 5–11, unknown chlorophyll-like pigments; 12, chlorophyll b; 13, chlorophyll a; 14, unknown carotene; 15, unknown pigment; 16,b-carotene; 17, fucoxanthin; 18, zeaxanthin; 19 and 20, unknown pigments. X-axis units are min.
Y-axis units are milliabsorbance units.
´
J. Bustillos-Guzman et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 77 –88 83
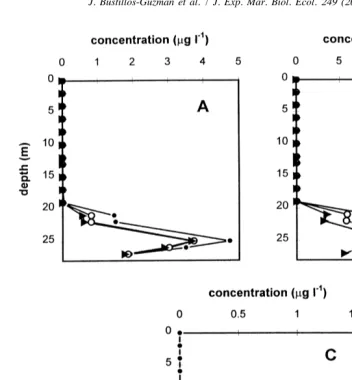
Fig. 4. Vertical variation of chlorophyll-like pigments in Bahia Concepcion, Gulf of California: (A) pigments 5, 6 and 7; (B) pigments 9, 10 and 11; (C) carotenoid 14.
4. Discussion
´
J. Bustillos-Guzman et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 77 –88 85
a-containing algae. It is also evident from the spectral characteristics of the unknown pigments that their presence in samples will interfere with the calculation of Chl a and Chl b. This will result in an overestimation of chlorophylls when using spectro-photometric measurements at some specific wavelengths and explains the high con-centration of Chl a and Chl b reported by Lechuga-Deveze (1994). We can now examine in detail these unknowns and other pigments and the physical and chemical conditions. The hydrographic structure corresponds well with the one previously described when high pigmented samples were recorded by Lechuga-Deveze (1994). According to Lechuga-Deveze et al. (2000), these conditions are caused because a change of wind direction and the warming of air temperature during the end of winter and beginning of spring induce the formation of a sharp thermocline that insulates the surface layer from bottom waters. Phytoplankton and zooplankton biomass develop profusely during the transition from mixed to stratified water column and supply organic matter that, when it sinks, exhausts the dissolved oxygen during the summer. The recurrence of anoxic water near the bottom and highly pigmented samples suggest that these conditions are a summer feature of this bay.
Distribution of pigments in samples seems to be associated with the physical and chemical structure of the water column. Thus, pigment signatures of cyanobacteria (zeaxanthin) were in the oxygenic, nitrate-poor, warmer surface water whereas di-noflagellate and diatom pigment (peridinin and fucoxanthin) maxima were at the base of the oxycline. The increase of cyanobacteria in surface waters has been related to their capacity to photosynthesize at high light intensities (Kana and Glibert, 1987), or to absorb photons in the green domain (Ong et al., 1986). Diatom and dinoflagellate pigments allow absorption in the spectral blue band (Bricaud et al., 1988; Antoine, 1995) and high nutrient levels are a prerequisite for proliferation (Cortes-Altamirano and
˜
Nunez-Pasten, 1986; Goldman, 1993; Bustillos-Guzman et al., 1995). Even though we did not record the highest nitrate concentration with the highest fucoxanthin and peridinin concentration, a mechanism of upwelling flux of nutrients at the base of the oxycline may be acting as has been suggested for other zones (Jorgensen, 1989; Goldman, 1993).
At the bottom, Chl-like and the carotenoid were dominant and were in a zone where oxygen concentration was very low or undetectable and H S appeared. The origin of2
intensities are too low to support growth of other phototrophic organisms (Pfennig and Truper, 1981; Schlegel and Jannasch, 1981; Schaub and van Gemerden, 1996). These conditions correspond well with that described here and others for the bay (Lechuga-Deveze, 1994; Lechuga-Deveze et al., 2000).
A second group of bacteriochlorophylls (BChls) or secondary homologues are also present in the Chlorobiaceae group and elute after the main set of BChls (Borrego and Garcia-Gil, 1994; Borrego et al., 1999). These could be the pigments eluting before the Chl a [peaks 8–11; Fig. 3(A)], but spectral characteristics are different to the similar BChl e set [peaks 6–7; Fig. 3(A)]. According to Otte et al. (1993) and Borrego et al. (1999), BChl e and their homologues have similar spectra. The absorption spectra of peaks 8–11, however, resemble the spectrum of bacteriopheophytin e (BPhtin e) and homologues (Gloe et al., 1975; Simpson and Smith, 1988; Borrego and Garcia-Gil, 1994). These pigments differ from BChl e in that they are Mg-free derivatives and their absorption maxima are at shorter wavelengths (Gloe et al., 1975). In our highly pigmented extracts, maximum absorption of Bphtin e is 32 nm lower than its parents. Bacteriopheophytins and homologues form an important part of the bacteriochlorophyll– protein complexes (Oelze, 1985). However, the high content of Bphtin e in our samples has to be considered with caution because Bphtin e can be an artifact of extraction as shown by Gloe et al. (1975) who observed that acetone extracts from Chlorobium cell material can contain up to 50% of this pigment and the response factor used to calculate the Bphtin e concentration is not correct.
In summary, the use of HPLC to analyze pigments in this research has helped us to advance the identification of the cause of the high pigmentation of the bottom of more than half the bay. Our analysis suggests that an important population of autotrophic anoxygenic bacteria is the main cause of the general pigment increase whereas Chl a increase is mainly linked to dinoflagellates and diatoms in Bahia Concepcion during summer. This bacterial population may play an important role in the nutrient reminerali-zation and anoxygenic photosynthesis in this and other coastal lagoons with similar physical and chemical characteristics.
Acknowledgements
This study was part of the IAYCG-11 project from CIBNOR and partially supported ´
by CONACYT (Mexico). We wish to thank Robert Blankenship, Daniel Brune and Michael Lince from Arizona State University and C. Borrego (Institute of Aquatic Ecology, Spain) and N.U. Triqaard for helpful discussions, the two referees for their critical review of the manuscript, and Ellis Glazier who edited this English-language paper. [RW]
References
´
J. Bustillos-Guzman et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 77 –88 87 Barlow, R.G., Mantoura, R.F., Gough, M.A., Fileman, T.W., 1993. Pigment signatures of the phytoplankton
composition in the northeastern Atlantic during the 1990 spring bloom. Deep Sea Res. 40, 459–477. Borrego, C.M., Garcia-Gil, L.J., 1994. Separation of bacteriochlorophyll homologues from green
photo-synthetic sulfur bacteria by reverse-phase HPLC. Photosynth. Res. 41, 157–163. ˜
Borrego, C.M., Baneras, M., Garcia-Gil, J., 1999. Temporal variability of Chlorobium phaeobacteroides antenna pigments in a meromictic karstic lake. Aquat. Microb. Ecol. 17, 121–129.
Bricaud, A., Bedhomme, A.L., Morel, A., 1988. Optical properties of diverse phytoplanktonic species: experimental results and theoretical interpretations. J. Plankton Res. 10, 851–873.
´
Bustillos-Guzman, J., Claustre, H., Marty, J.-C., 1995. Specific phytoplankton signatures and their relationship to hydrographic conditions in the coastal northwestern Mediterranean Sea. Mar. Ecol. Progr. Ser. 124, 247–258.
Clesceri, L.S., Greenberg, A.E., Trussell, R.R., 1989. Standard Methods for the Examination of Water and Wastewater, APHA, AWWA and WPCF.
˜
Cortes-Altamirano, R., Nunez-Pasten, A., 1986. Twelve years (1979–1990) of red tide records in Mazatlan Bay, Sinaloa, Mexico. Anal. Inst. Cienc. Mar Limnol. UNAM. 19, 113–121.
Frigaard, N.-U., Larsen, K.L., Cox, R.P., 1996. Spectrochromatography of photosynthetic pigments as a fingerprinting technique for microbial phototrophs. FEMS Microbiol. Ecol. 20, 69–77.
Gilmartin, J.Y., Revelante, N., 1978. The phytoplankton characteristics of the barrier island lagoons of the Gulf of California. Est. Coast. Mar. Sci. 7, 29–47.
Gloe, A., Pfennig, N., Brockmann, Jr. H., Trowitzsch, W., 1975. A new bacteriochlorophyll from brown-colored Chlorobiaceae. Arch. Microbiol. 102, 103–109.
Goericke, R., Repeta, D., 1993. The pigments of Prochlorococcus marinus: the presence of divinyl chlorophyll a and b in a marine prokaryote. Limnol. Oceanogr. 37, 426–433.
Goldman, J.C., 1993. Potential role of large oceanic diatoms in new primary production. Deep Sea Res. 40, 159–168.
Guillard, R.R.L., Murphy, L.S., Foss, P., Liaaen-Jensen, S., 1985. Synechococcus spp. as likely zeaxanthin-dominant ultraplankton in the North Atlantic. Limnol. Oceanogr. 30, 412–414.
Jeffrey, S.W., 1974. Profiles of photosynthetic pigments in the central North Pacific Ocean. Mar. Biol. 34, 101–110.
Jeffrey, S.W., 1997. Preparation of chlorophyll standards. In: Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W. (Eds.), Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods, Monographs on Oceanographic Methodology, Vol. 10, UNESCO Publishing, pp. 207–238.
Jorgensen, B.B., 1989. Biogeochemistry of chemoautotrophic bacteria. In: Schlegel, H.G., Bowien, B. (Eds.), Autotrophic Bacteria, Science Tech Publishers, USA, pp. 117–146.
22 21
Kana, T.M., Glibert, P.M., 1987. Effect of irradiance up to 2000 mE m s on marine Synechococcus WH7803-I. Deep Sea Res. 34, 179–195.
Kiefer, D.A., Laker, R., 1975. Two blooms of Gimnodinium splendens, an unarmored dinoflagellate. Fish. Bull. 73, 675–678.
Lechuga-Deveze, C.H., 1994. Shift of acetone extracted pigments due to unknown natural phytoplankton populations from Conception Bay, Gulf of California. Bull. Mar. Sci. 55, 248–255.
Lechuga-Deveze, C.H., Morquecho-Escamilla, L., 1999. Early spring potentially harmful phytoplankton in Bahia Concepcion, Gulf of California. Bull. Mar. Sci. 64, 1–10.
Lechuga-Deveze, C.H., Morquecho-Escamilla, M.L., Reyes-Salinas, A., Hernandez-Alfonso, J.R., 2000. Environmental natural disturbances at Bahia Concepcion, Gulf of California. In: Munawar, M., Laurence, S.G., Munawar, I.F., and Mallay, D.F. (Eds.) Ecosystem of Mexico: Status and scope. Backhuys Publishers B.V., Netherlands. 435 pp., In press.
Liaaen-Jensen, S., 1985. Carotenoids of lower plants — recent progress. Pure Appl. Chem. 57, 649–658. Liaaen-Jensen, S., Andrewes, A.G., 1972. Microbial carotenoids. Ann. Rev. Microbiol. 26, 227–247. Lopez-Cortez, A., 1999. Marine cyanobacteria from Bahia Concepcion, B.C.S. Mexico. In: Charpy, L.,
Larkum, A.W.D. (Eds.), Marine Cyanobacteria, Bulletin de l’Institute Oceanographique, Monaco, No. 19, 87–93.
Martinez-Lopez, A., Garate-Lizarraga, I., 1994. Quantity and quality of the particulate organic matter in Conception Bay, during the spawning season of the scallop Argopecten circularis (Sowerby, 1835). Cienc. Marinas 20, 301–320.
Oelze, J., 1985. Analysis of bacteriochlorophylls. Methods Microbiol. 18, 257–284.
Ong, L.J., Glazer, A.N., Waterbury, J.B., 1986. An unusual phycoerythrin from a marine cyanobacterium. Science 224, 80–82.
Otte, S.C.M., van de Meent, E.J., van Veelen, P.A., Pundsnes, A., Amesz, J., 1993. Identification of the major chlorosomal bacteriochlorophylls of the green sulfur bacteria Chlorobium vibriforme and chlorobium
phaeobacteroides: their functions in lateral energy transfer. Photosynth. Res. 35, 159–169.
Pfennig, N., 1989. Ecology of phototrophic purple and green sulfur bacteria. In: Schlegel, H.G., Bowien, B. (Eds.), Autotrophic Bacteria, Science Tech Publishers, USA, pp. 97–116.
Pfennig, N., Truper, H.G., 1981. Isolation of member of the families Chromatiacea and Chlorobiaceae. In: Starr, P.M., Stolp, H., Truper, H.G., Balows, A., Schlegel, H.G. (Eds.), The Prokaryotes, Springer Verlag, NY, pp. 279–289.
Schaub, B.E.M., van Gemerden, H., 1996. Sulfur bacteria in sediments of two coastal ecosystems: the Bassin ´
d’Arcachon and the Etang du Prevost, France. Hydrobiologia 329, 199–210.
Schlegel, H.G., Jannasch, H.W., 1981. Prokaryotes and their habitats. In: Starr, P.M., Stolp, H., Truper, H.G., Balows, A., Schlegel, H.G. (Eds.), The Prokaryotes, Springer Verlag, NY, pp. 43–82.
Simpson, D.J., Smith, K.M., 1988. Structures and transformation of the bacteriochlorophylls e and their bacteriopheophorbides. J. Am. Chem. Soc. 110, 1753–1758.
Strickland, J.D.H., Parson, T.R., 1972. A practical handbook of seawater analysis. Fisheries Res. Board of Canada. Bull. 310 pp.
Truper, H.G., 1970. Culture and isolation of phototrophic sulfur bacteria from the marine environment. Helgolander Wiss. Meeresunters 20, 6–16.
Vidussi, F., Claustre, H., Bustillos-Guzman, J., Cailleau, C., Marty, J.-C., 1996. Determination of chlorophylls and carotenoids of marine phytoplankton: separation of chlorophyll a from divinyl-chlorophyll a and zeaxanthin from lutein. J. Plankton Res. 18, 2377–2382.