www.elsevier.com / locate / bres
Interactive report
Light-induced zif268 expression is dependent on noradrenergic input
1in rat visual cortex
a b ,
*
cRaphael Pinaud , Liisa A. Tremere
, Marsha R. Penner
a
Department of Anatomy and Neurobiology, Dalhousie University, Halifax, Canada B3H 4H7 b
Department of Physiology and Biophysics, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4H7 c
Department of Pharmacology, Dalhousie University, Halifax, Canada B3H 4H7
Accepted 10 September 2000
Abstract
In the present paper we investigated the role of the noradrenergic projection from the locus coeruleus on the expression of the immediate early gene zif268 in the visual cortex of rats exposed to ambient light stimulation. Local administrations of 6-hydroxy-dopamine (6-OHDA), a specific toxin directed against the catecholaminergic system, were performed in the locus coeruleus prior to visual stimulation. Animals were stimulated for 2 h by ambient light, after a 2-week dark adaptation period. Sham-operated controls displayed a massive increase in the number of zif268 positive cells after light stimulation. To the contrary, lesioned animals demonstrated a dramatic reduction in the number of zif268 positive nuclei across all cortical layers. A few scattered immunopositive nuclei were identified in cortical layer IV, however, this region also underwent a significant reduction in the number of zif268 immunopositive nuclei. Our results indicate that the noradrenergic system plays an important role in the expression of zif268 in the visual cortex of rats exposed to ambient light after dark isolation. 2000 Elsevier Science B.V. All rights reserved.
Theme: Cellular and molecular biology
Topic: Gene structure and function: general
Keywords: Noradrenaline; Immediate early gene; Zif268; Visual cortex; Plasticity
Over the past 10 years, the expression of immediate [10]. In fact, it has been previously demonstrated that early genes (IEGs) has become widely used as a marker Zif268 regulates the expression of the synapsin I gene, for cortical neuronal activation, in various sensory systems which is involved in neurite development and synaptic [3–5,9,11]. IEGs are characterized by rapid and transient maturation [18].
induction, without the requirement of new protein syn- In the rat brain, the major source of cortical norad-thesis. The transcription factors encoded by this class of renaline is the nucleus locus coeruleus (LC), located in the genes have been postulated to be correlated with long-term dorso-lateral pons [7,8]. It has been previously demon-changes in central nervous system function [1,14]. Of strated that activity in the noradrenergic LC varies with the particular interest is the protein encoded by the IEG zif268 level of arousal and also with the amount of activity within (also known as NGFI-A, egr-1, Krox-24 and ZENK). This a sensory pathway [7]. The sensitivity of noradrenergic LC zinc-finger transcription factor has a high affinity for a neurons and zif268 expressing cells to sensory stimulation specific DNA motif, found in the promoter region of a prompted us to evaluate the contribution of the noradrener-number of ‘late-response genes’, which are thought to be gic projections to the expression of zif268 in neurons in responsible for long-lasting changes in neuronal function the visual cortex of animals exposed to sensory
stimula-tion.
A total of 20 adult male Wistar rats, weighing between
1
Published on the World Wide Web on 29 September 2000.
200 and 250 g, were used in the present study. Animals
*Corresponding author. Tel.: 11-902-494-2651; fax: 1
1-902-494-were anesthetized with sodium pentobarbital (50 mg / kg,
1685.
E-mail address: [email protected] (L.A. Tremere). i.p.) prior to a single injection of 6-hydroxydopamine
(6-OHDA; 2.5 l / ml), a specific toxin directed against the hydrogen peroxide had been added. Sections were dehy-catecholaminergic system, into the LC. To avoid possible drated, defatted and coverslipped for analysis. In addition, influences of 6-OHDA on serotonergic projections, all the extent to which the noradrenergic system was impaired animals were pretreated with the serotonin uptake inhibitor was assessed by immunocytochemistry directed against the fluoxetine (10 mg / kg, i.p.), administered 40 min prior the final synthetic enzyme for noradrenaline, dopamine-b -hy-6-OHDA infusion. -hy-6-OHDA was injected into the right droxylase (DBH), as described elsewhere [13].
hemisphere through a Hamilton syringe. Two microliters Photomicrographs were taken with a digital camera of a solution of 6-OHDA, in saline containing ascorbic mounted on an Zeiss-Axioplan microscope coupled to a acid (1 mg / ml) was injected directly in the LC, which was Macintosh computer. The grayscale of pictures were located according to the rat brain atlas published by adjusted using Adobe Photoshop software, without modi-Paxinos and Watson [16]. After the infusion procedure, all fications to the original data. A representative section from rats were individually housed in their home cages, in total the visual cortex of 8 animals (4 sham-operated and 4 darkness, where they recovered for 2 weeks, with water 6-OHDA treated) was selected for cell counting. These and food ad libitum. To ensure post-operative health, sections were chosen at the same stereotaxic level. In animals were observed under infrared light, which is addition, an adjacent section was used to assess cyto-invisible to rats [15]; all animals fully recovered without architectonic features. The area in 6-OHDA treated ani-complications. Control animals (n512) consisted of rats mals that continued to display Zif268 immunoreactivity injected unilaterally in the LC with a saline solution was identified as corresponding to cortical layer IV. Cell containing ascorbic acid (1 mg / ml), after the pre-treatment counts were performed for an area of tissue of 300mm in
with fluoxetine. width and 150mm in height. The decrease in the number
We used a paradigm of light stimulation after dark of labeled nuclei was averaged and calculated as a adaptation, which has already been demonstrated to be percentage. The raw data (Fig. 3) were evaluated with a effective not only for increasing synaptic activity, but also Kolmogorov–Smirnov non-parametric statistic.
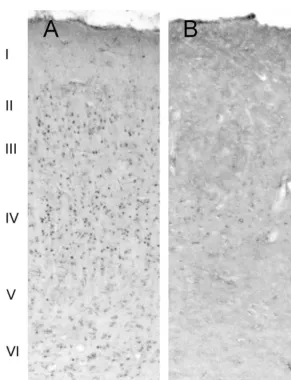
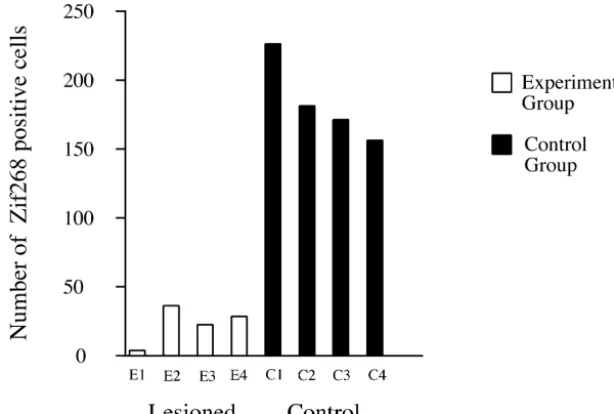
inducing plasticity in the visual cortex [11,12]. In our experimental animals, 6-OHDA injections were Following the injection of 6-OHDA, animals stayed in effective in severely depleting the majority of LC cells the dark-isolation room in order to minimize basal zif268 (Fig. 1). In accordance with previous studies performed in expression and to allow the toxin to affect noradrenergic rats, opossums and monkeys, light stimulation after dark cells in the LC. After 2 weeks of dark adaptation, rats were adaptation produced a pronounced increase in zif268 exposed to 2 h of ambient light. Animals that were immunoreactivity in the primary visual cortex [11,12,17,3]. spontaneously active for 80 min or longer of the stimula- This immunoreactivity was most prominent in layer IV, but tion period were considered in the present study. layers II, III, V and VI also displayed a large number of After stimulation, all animals were injected with an Zif268 positive nuclei, while no immunolabeling was overdose of sodium pentobarbital (70 mg / kg, i.p.). Rats identified in layer I (Fig. 2A). In contrast, animals subject-were perfused transcardially with 100 ml of 0.1 M PBS, ed to local administration of 6-OHDA in the LC displayed pH57.4, followed by 400 ml of 4% paraformaldehyde in a dramatic ipsilateral reduction of the number of Zif268 0.1 M PBS. Brains were removed, cryoprotected in a 30% positive nuclei. A few scattered labeled cell nuclei were sucrose solution and fast-frozen in a dry-ice / ethanol bath. identified in cortical layer IV of the primary visual cortex, Sections were cut at 20 mm in the coronal plane using a however, no immunostaining was found in other layers cryostat, and were directly mounted onto slides pre-coated (Fig. 2B). A comparison of the sham-operated control and with gelatin. All slides were stored at 2708C until further 6-OHDA lesioned groups revealed a dramatic reduction in
processing. the number of immunopositive nuclei in layer IV of the
We used the well-characterized polyclonal anti-egr-1 lesioned animals (Fig. 3); in layer IV the average reduction antibody raised against the carboxy terminus of the mouse in the number of labeled nuclei was 88%.
protein at 0.25 mg / ml (Santa Cruz Biotechnology, Santa Our work demonstrates that the LC noradrenergic Cruz, CA). Briefly, the immunocytochemical protocol projection plays a major role in the expression of zif268 in consisted of: (1) 2 h incubation at room temperature (RT) the primary visual cortex of rats exposed to ambient light. in a blocking buffer solution (0.5% albumin and 0.3% It has been previously demonstrated that zif268 expression Triton X-100 in 0.1 M PBS); (2) Overnight incubation in in the neocortex of rats is regulated by synaptic activity primary anti-egr-1 antibody at 48C; (3) 2 h at RT in [19], and that there are high basal levels of this protein in biotinylated goat-anti-rabbit secondary antibody (1:200 the rat visual cortex [9,11]. Moreover, Bhat and Baraban dilution in blocking buffer; Vector Laboratories); (4) demonstrated that the noradrenergic system is involved in Incubation in avidin–biotin–peroxidase complex for 2 h at the maintenance of the high zif268 basal levels in the rat RT (1:100, Vector Laboratories). All steps were separated neocortex [2]. In the present paper we demonstrate that by 30 min washes in 0.1 M PBS. Slides were incubated in noradrenaline also modulates the induction of genetic a solution of 0.03% diaminobenzidine and 0.15% nickel- mechanisms sensitive to sensory stimulation.
Fig. 1. (A) Photomicrograph depicting control levels of DBH immunostaining in the LC. (B) Lesion of LC is demonstrated as loss of DBH immunolabeling following 6-OHDA treatment. Scale bar550mm.
Fig. 3. Histograms representing cell counts (n58; 4 control and 4 lesioned) performed in individual animals, in a region selected from cortical layer IV. Note the pronounced difference between controls (black columns), which were injected with saline solution in the locus coeruleus, and experimental animals (white columns) which received an injection of 6-OHDA before light stimulation. The differences between values from control and lesioned animals were statistically significant (D510.00, df52, P,0.05).
strictly reflect neuronal activation, but rather is a com- References
ponent of a cascade of events that may lead to long-term
[1] W.C. Abraham, M. Dragunow, W.P. Tate, The role of immediate
changes in cortical circuitry [9–11]. There is considerable
early genes in the stabilization of long-term potentiation, Molec.
support for the hypothesis that zif268 is involved directly
Neurobiol. 5 (1991) 297–313.
or indirectly in the process of cortical plasticity. For [2] R.V. Bhat, J.M. Baraban, High basal expression of zif268 in cortex is example, it has been demonstrated that antagonism of the dependent on intact noradrenergic system, Eur. J. Pharmacol. 227 glutamatergic NMDA receptor, the activation of which is (1992) 447–448.
[3] A. Chaudhuri, M.S. Cynader, Activity-dependent expression of the
thought to be critical for inducing modifications in synaptic
transcription factor Zif268 reveals ocular dominance columns in
efficacy in the cortex, blocks zif268 expression [6]. In
monkey visual cortex, Brain Res. 605 (1993) 349–353.
addition, Thiel and colleagues showed that Zif268 reg- [4] A. Chaudhuri, J.A. Matsubara, M.S. Cynader, Neuronal activity in ulates the expression of the synapsin I gene, which is primate visual cortex assessed by immunostaining for the
transcrip-involved in synaptic maturation, neurotransmitter release tion factor Zif268, Vis. Neurosci. 12 (1995) 35–50.
[5] A. Chaudhuri, Neural activity mapping with inducible transcription
and neurite outgrowth [18]. In our work, we used light
factors, NeuroReport 8 (1997) 2–7.
stimulation after dark adaptation, a procedure that invokes
[6] A.J. Cole, D.W. Saffen, J.M. Baraban, P.F. Worley, Rapid increase of
neural plasticity [11,12]. The massive increase in Zif268 an immediate early gene messenger RNA in hippocampal neurons expression in our control animals supports the involvement by synaptic NMDA receptor activation, Nature 340 (1989) 474–
of this gene in the pathway that may lead to rearrangement 476.
[7] S.L. Foote, F.E. Bloom, G. Aston-Jones, Nucleus locus coeruleus:
of cortical circuits. In addition, the decrease of Zif268
new evidence of anatomical and physiological specificity, Physiol.
expression by depletion of the LC neurons indicates a role
Rev. 63 (1983) 844–914.
for noradrenaline in regulating the activation of this gene. [8] R. Grzanna, M.E. Molliver, The locus coeruleus in the rat: an
immunohistochemical delineation, Neuroscience 5 (1980) 21–40. [9] T. Herdegen, J.D. Leah, Inducible and constitutive transcription
factors in the mammalian nervous system: control of gene expres-Acknowledgements
sion by Jun, Fos and Krox, and CREB /ATF proteins, Brain Res. Rev. 28 (1998) 370–490.
This work was supported by the Medical Research [10] P. Hughes, M. Dragunow, Induction of immediate early genes and Council of Canada, CNPq, and an Izaak Walton Killam the control of neurotransmitter-regulated gene expression within the
Scholarship (to R.P.). The authors would like to thank Drs. nervous system, Pharmacol. Rev. 47 (1995) 133–178.
[11] L. Kaczmarek, A. Chaudhuri, Sensory regulation of immediate–
William Baldridge, Samuel Deurveilher and R. William
early gene expression in mammalian visual cortex: implications for
Currie for critically reading the manuscript and for useful
functional neural plasticity, Brain Res. Rev. 23 (1997) 237–256.
discussions. Raphael Pinaud would like to dedicate this [12] D. Konopka, A.W. Szklarczyk, R.K. Filipkowski, A. Trauzold, D. paper to Prof. (and mentor) Carlos Eduardo Rocha- Nowicka, M. Hetman, L. Kaczmarek, Plasticity- and
induc-ible cyclic-AMP early repressor messenger RNA expression in the [17] R. Pinaud, C.D. Vargas, M.V. Monteiro, S. Ribeiro, G. Cecchi, M.O. rat brain, Neuroscience 86 (1998) 499–510. Magnasco, C.V. Mello, C.E. Rocha-Miranda, E. Volchan, Regulation [13] C.V. Mello, R. Pinaud, S. Ribeiro, Noradrenergic system of the zebra of the expression of the early gene egr-1 in the opossum visual finch brain: immunocytochemical study of dopamine-b-hydroxylase, system by deprivation and exposure to light, Soc. Neurosci. 25 J. Comp. Neurol. 400 (1998) 207–228. (1999) 2190.
[14] D.V. Madison, R.C. Malenka, R.A. Nicoll, Mechanisms underlying [18] G. Thiel, S. Schoch, D. Petersohn, Regulation of synapsin I gene long-term potentiation of synaptic transmission, Annu. Rev. Neuro- expression by the zinc finger transcription factor zif-268 / egr-1, J.
sci. 14 (1991) 379–397. Biol. Chem. 269 (1994) 15294–15301.
[15] J. Neitz, G.H. Jacobs, Reexamination of spectral mechanisms in the [19] P.F. Worley, B.A. Christy, Y. Nakabeppu, R.V. Bhat, A.J. Cole, J. M rat (Rattus norvegicus), J. Comp. Psychol. 100 (1986) 21–29. Baraban, Constitutive expression of zif268 in neocortex is regulated [16] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates, by synaptic activity, Proc. Natl. Acad. Sci. USA 88 (1991) 5106–