PENGARUH DELIGNIFIKASI MENGGUNAKAN
Phanerochaete chrysosporium DAN
HIDROLISIS OLEH KAPANG SELULOLITIK TERHADAP
KUALITAS TONGKOL JAGUNG SEBAGAI PAKAN TERNAK
SKRIPSI
JULIANDO SARAGIH
F34060005
DEPARTEMEN TEKNOLOGI INDUSTRI PERTANIAN
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
THE EFFECT OF DELIGNIFICATION USING Phanerochaete chrysosporium AND HYDROLYSIS BY CELLULOLYTIC FUNGI TO CONCORB QUALITY AS LIVESTOCK FEED
Juliando Saragih, Liesbetini Hartoto, and Titi Candra Sunarti
Department of Agricultural Engineering, Faculty of Agricultural Technology,
Bogor Agricultural University, IPB Darmaga Campus, PO Box 220, Bogor, West Java, Indonesia Phone 62 251 8624622, e-mail: [email protected]
ABSTRACT
Concorb was one of the agricultural waste that has potency for ruminant feed. Concorb has high fiber content, but it has limited application since its low digestibility and protein content. Lignin in corncob should be liberate from cellulose and hemicellulose in the fiber substances. This research used white-root fungus of Phanerochaete chrysosporium for delignification and fungi Aspergillus niger and Trichoderma viride for hydrolysis the delignified corncob to increase its digestibility as feed. Urea and ammonium sulphate were added to enrich the corncob during hydrolysis in solid-state cultivation. After 20 days of biodelignification, 17.76 % of lignin was removed. Cultivation of delignified corncob by cellulolytic fungi increased the total and reducing sugar contents. The best hydrolysis treatment obtained from T. viride enriched with ammonium sulphate while A. niger with urea.T. viride increased 14.11 % of dry matter and 13.76 % of organic matter digestibilities. A. niger increased 15.03 % of dry matter and 8.11 % of organic matter digestibilities. Increasing of oligosacaride content made hydrolized corncob as prebiotic for better feed quality. Mixing isolates between A. niger and T. viride hydrolysis is expected to give better results in next research.
Keywords : Concorb, Biodelignification, hydrolysis, Phanerochaete chrysosporium, Aspergillus niger, Trichoderma viride
Juliando Saragih. F34060005. Pengaruh Delignifikasi Menggunakan Phanerochaete
chrysosporium dan Hidrolisis oleh Kapang Selulolitik terhadap Kualitas Tongkol Jagung
sebagai Pakan Ternak. Di bawah bimbingan Liesbetini Hartoto dan Titi Candra Sunarti. 2010
RINGKASAN
Tongkol jagung merupakan salah satu limbah pertanian yang potensial untuk dimanfaatkan. Berdasarkan data Badan Pusat Statistik, produksi tongkol jagung di Indonesia pada tahun 2009 mencapai 17,5 juta ton. Menurut Irawadi (1990), limbah pertanian (termasuk tongkol jagung) mengandung selulosa (40-60 %), hemiselulosa (20-30 %), dan lignin (15-30 %). Sampai saat ini, tongkol jagung sudah dimanfaatkan sebagai pakan tambahan pada hewan ruminansia. Kandungan selulosa tongkol jagung yang tinggi dapat dijadikan sumber energi utama yang menyokong pertumbuhan, reproduksi, dan produksi hewan ruminansia. Akan tetapi, pemanfaatan tongkol jagung sebagai pakan tidak dapat dimaksimalkan karena kandungan protein dan tingkat kecernaan yang rendah. Rendahnya kemampuan mikroba dalam rumen untuk merombak lapisan polisakarida dinding sel diakibatkan oleh keberadaan lapisan lignin yang membungkus selulosa dan hemiselulosa. Hidrolisis selulosa maupun hemiselulosa oleh rumen akan berjalan dengan lebih baik jika sebelumnya lapisan lignin telah didegradasi.
Penelitian ini bertujuan untuk mengkaji pengaruh delignifikasi tongkol jagung menggunakan kapang Phanerochaete chrysosporium dan memanfaatkannya sebagai bahan pakan ternak melalui proses hidrolisis oleh kapang selulolitik. Kapang selulolitik yang digunakan dalam penelitian ini yaitu Trichoderma viride dan Aspergillus niger.
Penelitian ini terbagi menjadi tiga tahap, yaitu persiapan tongkol jagung, delignifikasi, dan hidrolisis. Pada tahap persiapan dilakukan analisa komposisi bahan awal (proksimat, lignin, selulosa, hemiselulosa, gula pereduksi, dan total gula). Tahap delignifikasi oleh P. chrysosporium dilakukan untuk melepaskan komponen lignin pada tongkol jagung dan memudahkan hidrolisis selulosa. Tahap hidrolisis dilakukan untuk mengetahui kemampuan kapang selulolitik Aspergillus niger dengan Trichoderma viride dalam menghidrolisis selulosa. Selain itu dikaji juga kemampuan kedua kapang tersebut untuk memanfaatkan sumber nitrogen sebagai komponen media pertumbuhan. Sumber nitrogen yang digunakan yaitu urea dan amonium sulfat (ZA).
Hasil penelitian yang diperoleh menunjukkan kadar lignin pada bahan awal sebesar 10.97 % (b.k) menurun menjadi 9.02 % (b.k) pada bahan hasil delignifikasi atau terjadi penurunan sebesar 17.76 % (b.k). Selain lignin, kadar hemiselulosa juga menurun sebesar 51.41 % (b.k) pada bahan awal menjadi 44.61% (b.k) pada bahan hasil delignifikasi atau menurun sebesar 13.22 % (b.k). Dari hasil yang diperoleh dapat diketahui bahwa kapang P. chrysosporium tidak hanya mendegradasi lignin, tetapi juga sekaligus memproduksi enzim xilanase untuk memecah hemiselulosa menjadi senyawa lebih sederhana.
Hidrolisis tongkol jagung terdelignifikasi oleh T. viride dengan penambahan ZA menunjukkan hasil penurunan kandungan selulosa, peningkatan total gula, dan peningkatan gula pereduksi yang paling baik dibandingkan penambahan urea. Perlakuan ini dapat meningkatkan kandungan gula pereduksi dari 0,25 % sampai 0.52 % dan total gula dari 0.52 % sampai 1.08 %. Hidrolisis tongkol jagung terdelignifikasi oleh A. niger dengan penambahan urea juga juga menunjukkan penurunan kandungan selulosa, serta peningkatan gula pereduksi dan total gula. Perlakuan ini meningkatkan gula pereduksi dari 0.16 % menjadi 0.35 % dan total gula dari 0.57 % sampai 0.62 %. Secara keseluruhan, hidrolisis T. viride dengan penambahan amonium sulfat menunjukkan peningkatan total gula dan gula pereduksi yang lebih baik dibandingkan hasil kultivasi A. niger dengan penambahan urea.
Analisa daya cerna tongkol jagung hasil perlakuan kultivasi terbaik secara in vitro menunjukkan peningkatan kecernaan bahan kering dan bahan organik. Hidrolisis tongkol jagung terdelignifikasi oleh T. viride dengan penambahan amonium sulfat meningkatkan kecernaan bahan kering sebesar 14.11 % dan kecernaan bahan organik sebesar 13.76 %. Sedangkan hidrolisis tongkol jagung terdelignifikasi oleh A. niger dengan penambahan urea meningkatkan kecernaan bahan kering sebesar 15.03 % dan kecernaan bahan organik sebesar 8.11 %. Secara keseluruhan, tongkol jagung
hasil delignifikasi dan hidrolisis belum dapat memenuhi kelayakan sebagai pakan ternak berbentuk konsentrat. Namun demikian, kerusakan pada ikatan lignoselulosa akan memudahkan degradasi selulosa oleh mikroba rumen. Selain itu, peningkatan komponen oligosakarida dalam bahan memungkinkan tongkol jagung hasil hidrolisis dapat berperan sebagai prebiotik yang dapat meningkatkan kecernaan bahan pakan.
PENGARUH DELIGNIFIKASI MENGGUNAKAN
Phanerochaete chrysosporium DAN
HIDROLISIS OLEH KAPANG SELULOLITIK TERHADAP
KUALITAS TONGKOL JAGUNG SEBAGAI PAKAN TERNAK
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar
SARJANA TEKNOLOGI PERTANIAN
pada Departemen Teknologi Industri Pertanian
Fakultas Teknologi Pertanian
Institut Pertanian Bogor
Oleh
JULIANDO SARAGIH
F34060005
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2010
Judul Skripsi : Pengaruh Delignifikasi Menggunakan Phanerochaete chrysosporium dan Hidrolisis oleh Kapang Selulolitik terhadap Kualitas Tongkol Jagung sebagai Pakan Ternak
Nama : Juliando Saragih
NIM : F34060005
Menyetujui,
Pembimbing I, Pembimbing II,
(Dr. Ir. Liesbetini Hartoto, MS) (Dr. Ir. Titi Candra Sunarti, MSi.)
NIP : 19550904 198003 2 001 NIP : 19661219 199103 2 001
Mengetahui :
Ketua Departemen Teknologi Industri Pertanian,
(Prof. Dr. Ir. Nastiti Siswi Indrasti) NIP : 19621009 198903 2 001
PERNYATAAN MENGENAI SKRIPSI
DAN SUMBER INFORMASI
Saya yang bertanda tangan di bawah ini menyatakan dengan sebenar-benarnya bahwa skripsi dengan judul “Pengaruh Delignifikasi Menggunakan Phanerochaete chrysosporium dan Hidrolisis oleh Kapang Selulolitik terhadap Kualitas Tongkol Jagung sebagai Pakan Ternak”
adalah hasil karya saya sendiri dengan arahan dosen pembimbing akademik. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir skripsi ini.
Bogor, November 2010 Yang membuat pernyataan
Juliando Saragih
RIWAYAT HIDUP
Juliando Saragih dilahirkan di Pematang Raya, Sumatera Utara pada tanggal 16 Juli 1988 dari ayah bernama Judiaman Saragih dan ibu yang bernama Demianna Purba. Penulis merupakan anak kedua dari tiga bersaudara. Riwayat pendidikan penulis dimulai dari tahun 1993 di Taman Kanak-kanak Bina Insani. Pada tahun 1994 sampai tahun 2000 penulis mengikuti pendidikan di Sekolah Dasar Inpres 097800 Kampung Jawa, kemudian melanjutkan ke Sekolah Lanjutan Tingkat Pertama Negeri 1 Pematang Raya dari tahun 2000 sampai tahun 2003. Tahun 2003 sampai tahun 2006 penulis menanjutkan pendidikan ke Sekolah Menengah Umum Negeri 1 Pematang Raya.Tahun 2003 penulis melanjutkan pendidikan di Tingkat Persiapan Bersama Kampus Institut Pertanian Bogor melalui jalur Ujian Seleksi Masuk IPB (USMI). Penulis melanjutkan pendidikan di Departemen Teknologi Industri Pertanian, Fakultas Teknologi Pertanian IPB dari tahun 2004. Penulis telah melakukan praktek lapang di PT. PG. Madukismo, Bantul, Yogyakarta. Penulis mengakhiri masa studi di IPB setelah menyelesaikan skripsi yang berjudul “Pengaruh Delignifikasi Menggunakan Phanerochaete
chrysosporium dan Hidrolisis oleh Kapang Selulolitik terhadap Kualitas Tongkol Jagung
KATA PENGANTAR
Puji syukur penulis panjatkan kepada Tuhan Yesus Kristus yang telah memberikan segala berkat dan anugrah, sehingga skripsi ini berhasil diselesaikan. Penelitian dengan judul “Pengaruh Delignifikasi Menggunakan Phanerochaete chrysosporium dan Hidrolisis oleh Kapang Selulolitik terhadap Kualitas Tongkol Jagung sebagai Pakan Ternak” dilaksanakan di Laboratorium Bioindustri, Departemen Teknologi Industri Pertanian, Fakultas Teknologi Pertanian, Institut Pertanian Bogor sejak bulan Maret hingga Agustus 2010.
Penulis menyadari bahwa penulisan skripsi ini tidak terlepas dari adanya bimbingan dan dukungan dari banyak pihak. Oleh karena itu, dengan segala kerendahan hati, penulis mengucapkan terima kasih kepada :
1. Ibu Dr. Ir. Liesbetini Hartoto, MS selaku dosen pembimbing I dan Dr. Ir. Titi Candra Sunarti, MSi selaku dosen pembimbing II yang telah sabar membimbing, memberikan pengarahan, kritik, saran, dan informasi untuk mendukung pendidikan dan penyelesaian skripsi penulis di Departemen Teknologi Industri Pertanian, FATETA, IPB
2. Bapak Ir. Andes Ismayana, MT selaku dosen penguji yang telah memberi saran dan kritikan kepada penulis untuk menyelesaikan skripsi ini
3. Seluruh keluarga yang senantiasa mendukung penulis dalam doa dan motivasi untuk manjalani hari-hari dengan penuh rasa syukur
4. Rekan-rekan di Persekutuan Mahasiswa Kristen IPB, terkhusus Kopelkhuers yang mendukung penulis dalam setiap kesulitan dan masalah
5. Seluruh dosen yang telah membimbing penulis selama menempuh pendidikan di Departemen Teknologi Pertanian, Fakultas Teknologi Pertanian IPB
6. Bu Ega, Pak Edy, Pak Darwan, Pak Gun, dan seluruh karyawan Departemen Teknologi Industri Pertanian, FATETA yang telah banyak memberikan bantuan
7. Pangeran, Yulia, Cucu, Winda, Yana, Martin, Yuli, Sarfat, Siska, Sely, Laura, Mumun, dan teman-teman TIN 43 yang menjadi rekan seperjuangan di kampus selama menempuh studi
8. Rano, Dwiko, Rio, Rudi, Juan, dan seluruh penghuni Malea Putra atas kebersamaan dan saling mendukung selama masa studi.
9. Seluruh pihak yang terlibat secara langsung maupun tidak langsung dalam pembuatan skripsi ini. Terimakasih semuanya.
Tentunya skripsi ini belum sempurna, tetapi penulis berharap semoga tulisan ini bermanfaat bagi yang menyusun maupun yang membaca.
Bogor, November 2010
DAFTAR ISI
Halaman KATA PENGANTAR ... i DAFTAR ISI ... ii DAFTAR TABEL ... iv DAFTAR GAMBAR ... v DAFTAR LAMPIRAN ... vi I. PENDAHULUAN A. LATAR BELAKANG ... 1 B. TUJUAN ... 2II. TINJAUAN PUSTAKA A. TONGKOL JAGUNG ... 3 B. Phanerochaete chrysosporium ... 4 C. Trichoderma viride ... 6 D. Aspergillus niger ... 7 E. DELIGNIFIKASI BIOLOGIS ... 8 F. HIDROLISIS SELULOLITIK ... 9
G. PAKAN TERNAK RUMINANSIA ... 10
III. METODOLOGI A. BAHAN DAN ALAT ... 12
1. Bahan ... 12
2. Alat ... 12
B. METODE PENELITIAN ... 12
1. Karakterisasi Bahan Baku ... 12
2. Delignifikasi Biologis ... 13
3. Hidrolisis Oleh Kapang Selulolitik ... 14
IV. HASIL DAN PEMBAHASAN
A. DELIGNIFIKASI TONGKOL JAGUNG ... 18
B. HIDROLISIS TONGKOL JAGUNG ... 22
1. Perlakuan Hidrolisis ... 22
2. Karakteristik Produk Hasil Hidrolisis... 23
a. Komponen Proksimat ... 23
b. Komponen Serat ... 27
c. Struktur Mikroskopik Tongkol Jagung Hasil Hidrolisis ... 30
d. Pembentukan Oligosakarida ... 33
e. Kecernaan Bahan Kering dan Bahan Organik (in vitro) ... 37
V. KESIMPULAN DAN SARAN A. KESIMPULAN ... 40
B. SARAN ... 40
DAFTAR PUSTAKA ... 41
DAFTAR TABEL
Halaman
1. Komposisi tongkol jagung (Johnson 1991) ... 4
2. Komponen mineral pertumbuhan kapang P. chrysosporium ... 13
3. Perubahan komposisi tongkol jagung sebelum dan setelah delignifikasi ... 19
4. Komposisi kimia bekatul (substrat inokulum) ... 23
5. Hasil analisia proksimat tongkol jagung sebelum dan setelah hidrolisis ... 25
6. Hasil analisa komponen serat tongkol jagung sebelum dan setelah hidrolisis ... 29
7. Perubahan gula pereduksi, total gula, dan derajat polimerisasi produk sebelum dan setelah hidrolisis ... 34
8. Peningkatan kecernaan bahan kering dan bahan organik tongkol jagung terhidrolisis ... 38
DAFTAR GAMBAR
Halaman
1. Konfigurasi dinding sel tanaman (Perez et al. 2002) ... 3
2. Skema sistem degradasi lignin oleh P. chrysosporium ... 5
3. Struktur mikroskopik miselia P. chrysosporium ... 5
4. T. viride di tanah (a) dan struktur mikroskopis (b) ... 7
5. A. niger dalam bentuk spora (a) dan struktur mikroskopis (b) ... 8
6. Hidrolisis selulosa menjadi glukosa (Lynd et al. 2002) ... 10
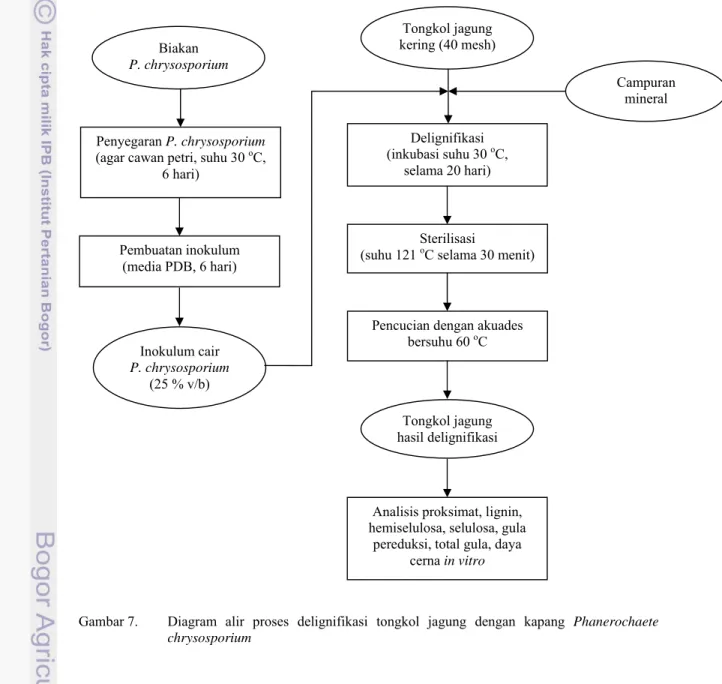
7. Diagram alir proses delignifikasi tongkol jagung ... 16
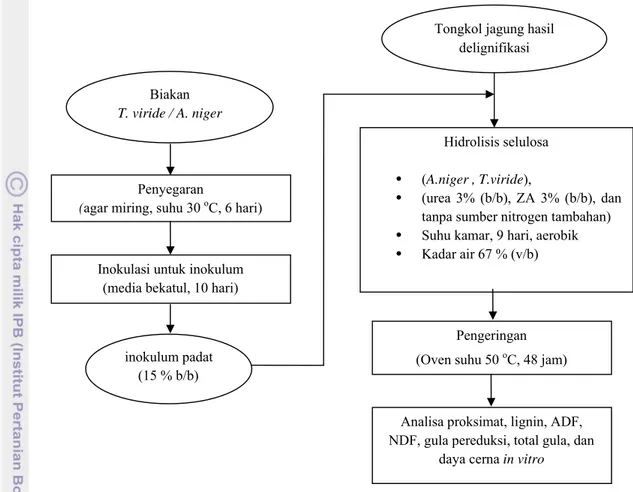
8. Diagram alir proses hidrolisis tongkol jagung terdelignifikasi ... 17
9. Struktur mikroskopis tongkol jagung awal dan terdelignifikasi ... 21
10. Perubahan nilai protein kasar bahan pada perlakuan berbeda ... 26
11. Struktur tongkol jagung (hidrolisis Aspergillus niger) ... 31
12. Struktur tongkol jagung (hidrolisis Trichoderma viride)... 33
13. Perubahan nilai kandungan total gula pada perlakuan hidrolisis ... 35
DAFTAR LAMPIRAN
Halaman
1. Prosedur analisis kimia ... 47
2. Foto aliran proses delignifikasi ... 55
3. Foto aliran proses Hidrolisis ... 56
4. Foto bahan sebelum dan setelah 20 hari delignifikasi ... 57
5. Data Analisa Proksimat Hidrolisis Tongkol Jagung ... 58
6. Dokumentasi sampel hasil hidrolisis ... 61
7. Analisa Statistik Terhadap Kadar Protein ... 62
8. Data Analisa Komponen Serat Hidrolisis Tongkol Jagung ... 64
9. Data Analisa Pembentukan Gula Hidrolisis Tongkol Jagung ... 66
10. Analisa Statistik Terhadap Pembentukan Gula ... 67
I. PENDAHULUAN
A.
LATAR BELAKANG
Tongkol jagung merupakan salah satu bagian terbesar dari buah jagung yang menjadi tempat menempelnya seluruh biji jagung. Tongkol jagung umumnya menjadi limbah yang dibuang begitu saja di lingkungan dan terurai secara alami di alam. Berdasarkan data Badan Pusat Statistik, produksi jagung di Indonesia pada tahun 2009 adalah 17.6 juta ton pipilan kering dan diperkirakan akan meningkat setiap tahunnya, sedangkan produksi tongkol jagung pada tahun yang sama mencapai 17.5 juta ton. Demikian halnya peningkatan produksi jagung akan diikuti peningkatan produksi tongkol jagung setiap tahunnya.
Tongkol jagung sebagai salah satu limbah sektor pertanian memiliki kandungan serat yang terdiri dari komponen lignoselulosa yaitu selulosa, hemiselulosa, dan lignin. Keberadaan komponen serat ini terutama selulosa sangat memungkinkan pemanfaatan tongkol sebagai pakan ternak ruminansia. Namun demikian, lapisan komponen lignoselulosa yang kuat dapat menyulitkan mikroba rumen ternak untuk mencerna tongkol jagung sebagai pakan. Dengan demikian dibutuhkan proses penguraian komponen lignoselulosa untuk meningkatkan kemampuan rumen mencerna tongkol jagung.
Kebutuhan akan pakan ruminansia yang berkualitas merupakan suatu masalah yang penting untuk dipenuhi pada saat ini. Secara umum ruminansia diberi pakan rumput segar yang dapat diperoleh dengan mudah dari alam. Namun kebutuhan pakan di saat padang rumput berkurang dan musim kemarau tetap harus disediakan. Menurut Djajanegara (1999), ada beberapa kendala penyediaan pakan hijauan yaitu, terjadinya perubahan fungsi lahan sumber hijauan menjadi pemukiman, lahan tanaman pangan, dan tanaman industri. Di sisi lain, limbah pertanian seperti tongkol jagung, jerami, ampas tebu, ampas tebu, dan bahan sejenis lain dapat dijadikan sumber pakan bagi ternak ruminansia. Akan tetapi limbah pertanian dan perkebunan ini umumnya mempunyai kandungan protein dan kecernaan yang rendah. Rendahnya kecernaan ini diakibatkan keberadaan lignin yang menghalangi proses perombakan polisakarida dinding sel oleh mikroba yang terdapat pada rumen.
Lapisan lignin dan hemiselulosa yang terikat merupakan bagian yang penting untuk dipisahkan dari tongkol jagung sehingga selulosa dapat dihidrolisa. Menurut Suparjo (2008), struktur berkristal serta adanya lignin dan hemiselulosa disekeliling selulosa merupakan hambatan utama dalam menghidrolisis selulosa. Kristalisasi selulosa dan pengerasan fibril selulosa oleh lignin membentuk suatu senyawa lignoselulosa yang keras. Efisiensi pemanfaatan selulosa sebagai sumber energi bagi ternak ruminansia sangat tergantung pada kemampuan ternak untuk memutus ikatan yang memproteksi selulosa dari serangan enzim selulase. Selulosa dan hemiselulosa pada lignoselulosa tidak dapat dihidrolisis oleh enzim selulase dan hemiselulase kecuali lignin yang ada pada substrat dilepaskan terlebih dahulu.
Pemisahan lignin dan penguraian ikatan hemiselulosa dapat dilakukan dengan proses delignifikasi menggunakan bahan kimia, enzim, maupun pemanfaatan mikroba (Martina 2002). Delignifikasi dilakukan sebagai proses pendahuluan untuk memudahkan pelepasan selulosa dan menghilangkan lignin yang terkandung dalam tongkol jagung. Hidrolisis dilakukan untuk memutus sisa ikatan lignin dan hemiselulosa di sekitar selulosa sekaligus menguraikan selulosa menjadi glukosa.
Pemanfaatan mikroba merupakan salah satu alternatif yang diharapkan akan menguntungkan dibandingkan pemanfaatan bahan kimia ataupun enzim secara langsung. Penggunaan bahan kimia dalam untuk menghidrolisis selulosa sulit diterapkan karena membutuhkan energi yang tinggi, bahan kimia yang tidak sedikit, dan menghasilkan limbah yang berbahaya (Martina 2002). Hidrolisis dengan menggunakan enzim secara langsung juga baik untuk dilakukan, namun biaya yang sangat tinggi dapat menghalangi terlaksananya penelitian. Dengan kondisi tersebut, pemanfaatan mikroba untuk menghilangkan lignin dan hidrolisis selulosa yang terkandung dalam tongkol jagung merupakan salah satu cara yang baik dilakukan untuk mengubah tongkol menjadi pakan yang baik bagi ruminansia.
Kapang Phanerochaete chrysosporium merupakan salah satu jenis kapang pelapuk putih yang baik untuk mendegradasi lignin sekaligus menguraikan ikatan polimer hemiselulosa dan selulosa. Menurut Hataka (2001), kapang pelapuk putih P. chrysosporium mampu mendegradasi lignin dan hemiselulosa lebih banyak dibandingkan selulosa, sehingga baik dimanfaatkan untuk proses delignifikasi. Bahan selanjutnya dihidrolisis dengan memanfaatkan kapang selulolitik. Trichoderma viride dan Aspergillus niger merupakan dua jenis kapang diharapkan dapat menghidrolisis selulosa sehingga daya cerna terhadap tongkol jagung dapat ditingkatkan. Penambahan sumber nitrogen dalam substrat pertumbuhan kapang juga diharapkan dapat meningkatkan kandungan protein tongkol jagung sebagai pakan ternak ruminansia.
B.
TUJUAN
Penelitian ini bertujuan untuk mengkaji pengaruh delignifikasi menggunakan kapang Phanerochaete chrysosporium dan hidrolisis oleh kapang selulolitik (Trichoderma viride dan Aspergillus niger) terhadap kualitas tongkol jagung yang akan digunakan sebagai bahan pakan ternak.
II. TINJAUAN PUSTAKA
A.
TONGKOL JAGUNG
Tongkol jagung merupakan salah satu limbah hasil pertanian dari buah jagung. Secara keseluruhan total bobot total, jagung terdiri dari 30 % bagian berupa tongkol jagung, sedangkan sisanya adalah biji dan kulit (Koswara 1991). Menurut Irawadi (1990), limbah pertanian (termasuk tongkol jagung) mengandung selulosa (40-60%), hemiselulosa (20-30%), dan lignin (15-30%).
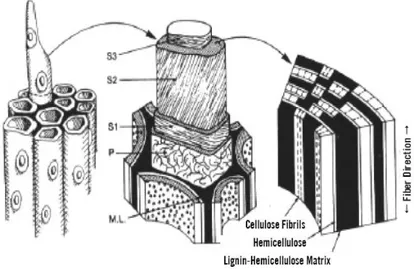
Komponen selulosa, hemiselulosa, dan lignin dalam tongkol jagung terikat satu sama lain dengan ikatan yang kompleks seperti terlihat pada Gambar 1. Menurut Perez et al. (2002), lapisan dinding sekunder terluar (S1) mempunyai struktur serat menyilang, lapisan S2 mempunyai mikrofibril yang paralel terhadap poros lumen, dan lapisan S3 mempunyai mikrofibril yang berbentuk heliks. Mikrofibril dikelilingi oleh hemiselulosa dan lignin. Bagian antara dua dinding sel disebut lamela tengah (M) dan diisi oleh hemiselulosa dan lignin. Selulosa secara alami terproteksi dari degradasi dengan adanya hemiselulosa dan lignin.
Gambar 1. Konfigurasi dinding sel tanaman (Perez et al. 2002)
Sampai saat ini tongkol jagung sudah dimanfaatkan untuk menghasilkan beberapa produk bernilai tambah seperti bioetanol, pakan ternak, maupun pengganti bahan bakar secara langsung. Sebagai pakan ternak, bahan yang mengandung komponen lignoselulosa (termasuk tongkol jagung) memiliki dua kelemahan yaitu tingkat kecernaan yang rendah dan kandungan protein yang rendah (Artika 2000). Karakteristik dan dan komposisi tongkol jagung dapat dilihat di Tabel 1.
Serat kasar yang tinggi yang terkandung di dalam tongkol jagung merupakan bahan yang penting yang memungkinkan tongkol jagung dimanfaatkan sebagai pakan ternak. Selulosa yang
tinggi dalam serat dapat dicerna oleh ternak ruminansia. Menurut Lynd et al. (2002), ternak ruminansia dapat memanfaatkan selulosa sebagai sumber energi utama dalam menyokong pertumbuhan, produksi, dan reproduksi. Selulosa merupakan komponen penyusun dinding sel tanaman yang tidak pernah ditemukan secara murni di alam, tetapi berikatan dengan bahan lain, yaitu lignin dan hemiselulosa.
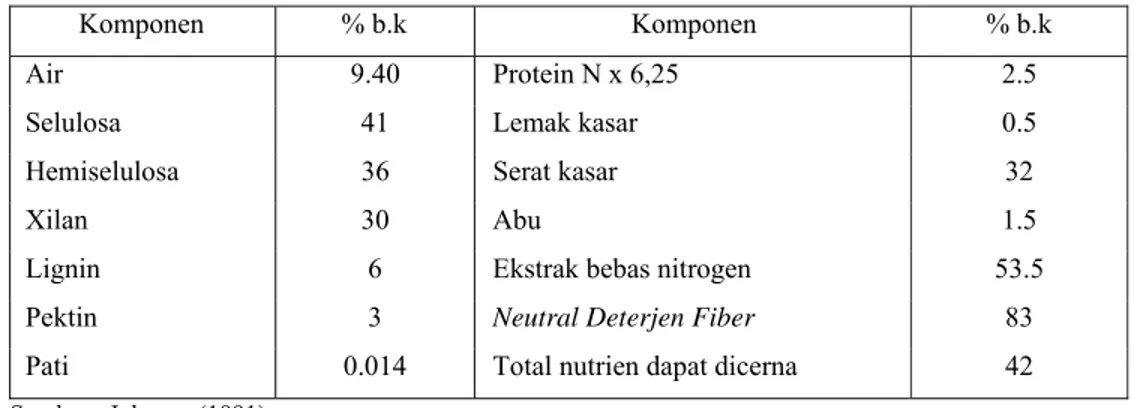
Tabel 1. Komposisi Kimia Tongkol Jagung
Komponen % b.k Komponen % b.k
Air 9.40 Protein N x 6,25 2.5
Selulosa 41 Lemak kasar 0.5
Hemiselulosa 36 Serat kasar 32
Xilan 30 Abu 1.5
Lignin 6 Ekstrak bebas nitrogen 53.5
Pektin 3 Neutral Deterjen Fiber 83
Pati 0.014 Total nutrien dapat dicerna 42
Sumber : Johnson (1991)
Struktur berkristal serta adanya lignin dan hemiselulosa disekeliling selulosa merupakan hambatan utama dalam menghidrolisis selulosa. Pemanfaatan selulosa oleh ternak sangat bergantung pada kemampuan ternak untuk memutus ikatan yang memproteksi selulosa. Pemutusan ikatan lignin yang memproteksi selulosa inilah yang disebut sebagai delignifikasi. Menurut Hofrichter (2002), lignin merupakan senyawa yang heterogen dengan berbagai tipe ikatan sehingga tidak dapat diuraikan oleh enzim hidrolisis. Namun lignin dapat didegradasi oleh kapang pelapuk kayu dan secara sempurna hanya oleh kapang pelapuk putih. Kapang Phanerochaete chrysosporium merupakan salah satu kapang yang dapat menguraikan ikatan dan mendegradasi lignin dengan bantuan enzim pendegradasi lignin.
B.
Phanerochaete chrysosporium
Phanerochaete chrysosporium adalah kapang pendegradasi lignin dari kelas Basidiomycetes yang tumbuh dalam bentuk sekumpulan miselia dan berkembang biak secara
aseksual (melalui spora) atau secara seksual (Dhawale dan Katrina 1993). Kapang P. chrysosporium dapat mendegradasi lignin dan senyawa turunannya secara efektif dengan cara
menghasilkan enzim peroksidase ekstraseluler yang berupa lignin peroksidase (LiP) dan mangan peroksidase (MnP) yang sama efektifitasnya dengan H2O2. Menurut Johjima et al. (1999), kapang P. chrysosporium dapat mendegradasi lignin dan berbagai polutan aromatik selama fase pertumbuhan stasioner yang dipacu oleh kekurangan nutrisi pada substrat. Skema sistem degradasi lignin oleh kapang tersebut dapat dilihat pada Gambar 2.
Gambar 2. Skema sistem degradasi lignin oleh Phanerochaete chrysosporium (Akhtar et al., 1997)
Menurut Fadillah et al. (2008), kapang P. chrysosporium dapat mendegradasi lignin pada
batang jagung sebesar 81.4 % setelah inkubasi pada suhu 38 oC selama 30 hari. Degradasi lignin juga diikuti dengan degradasi selulosa walaupun jumlahnya relatif lebih sedikit yaitu sebesar 22.3 % setelah 30 hari inkubasi. P. chrysosporium adalah jenis kapang yang termasuk kelas Basidiomycetes tingkat tinggi yang dapat membentuk badan buah. Badan buah kelas Basidiomycetes memiliki ukuran yang beraneka ragam, mulai dari yang berukuran mikroskopik sampai dengan yang bergaris tengah sepanjang 3 kaki (Alexopoulus dan Mims 1979). Umumnya pada media dengan kandungan nitrogen yang rendah, P. chrysosporium mampu mendegradasi sebagian besar zat warna. Pada media dengan konsentrasi nitrogen yang cukup tinggi, fraksi zat
warna yang terserap oleh biomassa akan meningkat. Struktur mikroskopis miselia P. chrysosporium dapat dilihat pada Gambar 3.
Untuk tumbuh dan berkembang dengan baik P. chrysosporium membutuhkan lingkungan dengan kondisi yang sesuai dengan karakteristiknya. Beberapa faktor fisiologis yang dibutuhkan untuk pertumbuhan kapang adalah suhu, oksigen, konsentrasi ion hidrogen (pH), dan nutrisi. Lingkungan fisik lain yaitu cahaya dan aerasi serta lingkungan biologis seperti interaksi dengan mikroba lain, juga akan mempengaruhi laju degradasi oleh kapang. Kisaran suhu optimum untuk
pertumbuhan kapang P. chrysosporium adalah antara 20-30 oC. Kapang ini dapat mengambil
oksigen secara bebas dari udara untuk keperluan respirasi agar pertumbuhannya optimum. Kekurangan oksigen dapat menyebabkan terjadinya akumulasi karbon dioksida (CO2), sehingga pertumbuhan kapang terhambat. Umumnya kapang lebih menyukai suasana asam untuk pertumbuhannya. Kisaran pH optimum untuk kapang dari kelas Basidiomycetes adalah pH 4.5 – 5.5 (Chang dan Hayes 1978).
Proses delignifikasi diharapkan dapat mendegradasi dan melepaskan ikatan lignin yang melapisi selulosa. Menurut Adaskaveg et al. (1995), P. chrysosporium mendegradasi komponen lignoselulosa secara selektif yaitu mendegradasi lignin substrat yang berwarna coklat dan meninggalkan selulosa yang berwarna putih. Dengan demikian lepasnya lignin tanpa terurainya selulosa akan memudahkan hidrolisis selulosa oleh kapang selulolitik.
C.
Trichoderma viride
Trichoderma viride adalah salah satu jenis kapang yang bersifat selulolitik karena dapat menghasilkan selulase (Wood 1985). Kapang ini mampu menghidrolisis selulosa tingkat tinggi dan memiliki kemampuan mensintesis beberapa komponen lignoselulosa yang terikat kuat dengan ikatan hidrogen. T. viride mampu menghasilkan selulase yang menghidrolisis selulosa pada biji-bijian sehingga dapat dicerna ternak. Selain itu T. viride mempunyai kemampuan meningkatkan protein bahan pakan dan pada bahan berselulosa mampu menghasilkan enzim selulase (Poesponegoro 1976). Kusumaningtyas dan Djaenudin (2006), menambahkan bahwa T. viride merupakan kapang yang bersifat antagonis terhadap berbagai jenis kapang patogen pada tanaman sehingga dapat dijadikan agen kontrol biologi.
Trichoderma adalah kapang penghuni tanah yang dapat diisolasi dari perakaran tanaman lapangan (Gambar 4). Spesies Trichoderma di samping sebagai organisme pengurai, dapat juga berfungsi sebagai agen hayati dan stimulator pertumbuhan tanaman. Beberapa spesies Trichoderma telah dilaporkan sebagai agen hayati adalah T. harzianum, T. viride, dan T. konigii yang berspektrum luas pada berbagai tanaman pertanian. Biakan Trichoderma dalam media aplikatif seperti dedak dapat diberikan ke areal pertanaman dan berlaku sebagai biodekomposer. Dengan demikian Trichoderma viride memiliki kemampuan mendekomposisi limbah organik (rontokan dedaunan dan ranting tua) menjadi kompos yang bermutu. Selain itu, Trichoderma dapat juga digunakan sebagai biofungisida karena mampu menghambat pertumbuhan beberapa kapang penyebab penyakit pada tanaman antara lain Rigidiforus lignosus, Fusarium oxysporum, Rizoctonia solani, dan Sclerotium rolfsii (Ramada 2008).
Ramada (2008) menyatakan bahwa pupuk biologis Trichoderma dapat dibuat dengan inokulasi biakan murni pada media aplikatif seperti dedak, sedangkan biakan murni dapat dibuat melalui isolasi dari perakaran tanaman, serta dapat diperbanyak dan diremajakan kembali pada media PDA (Potato Dextrose Agar).
(a) (b) Gambar 4. T. viride di tanah (a) dan struktur mikroskopis (b)
(Volk 2006)
Kapang Trichoderma viride juga digunakan untuk meningkatkan nilai manfaat jerami
padi melalui kultivasi, karena kapang ini mempunyai sifat selulolitik dan mengeluarkan enzim selulase yang dapat merombak selulosa menjadi selobiosa hingga akhirnya menjadi glukosa. Proses yang terjadi ketika jerami padi dikultivasi menggunakan Trichoderma viride adalah terjadinya degradasi terhadap dinding sel yang diselaputi oleh lignin, selulosa, dan hemiselulosa. Akibat degradasi ini, sebagian lignin akan terurai, sedangkan selulosa dan hemiselulosa juga akan terurai menjadi glukosa (Mandels dan Reese 1957).
D.
Aspergillus niger
Aspergillus niger termasuk kapang selulolitik yang menghasilkan selulase untuk menghidrolisis selulosa (Usama et al. 2008). Frazier dan Westhoff (1981), menyatakan bahwa Aspergillus niger merupakan kapang yang termasuk dalam genus Aspergillus, famili Moniliaceae, ordo Monoliales, dan kelas Fungi imperfecti. Aspergillus adalah kapang yang
hifanya berseptat dan sporanya bersifat aseksual. Spora aspergillus berbentuk globula dan
konidianya kasar dengan beberapa pita yang berpigmen. Ada dua macam pita pada Aspergillus, yaitu hifa yang terletak pada bagian terendam dari substrat, berfungsi untuk menyerap zat hara, sedangkan bagian yang menghadap ke permukaan berfungsi sebagai alat reproduksi.
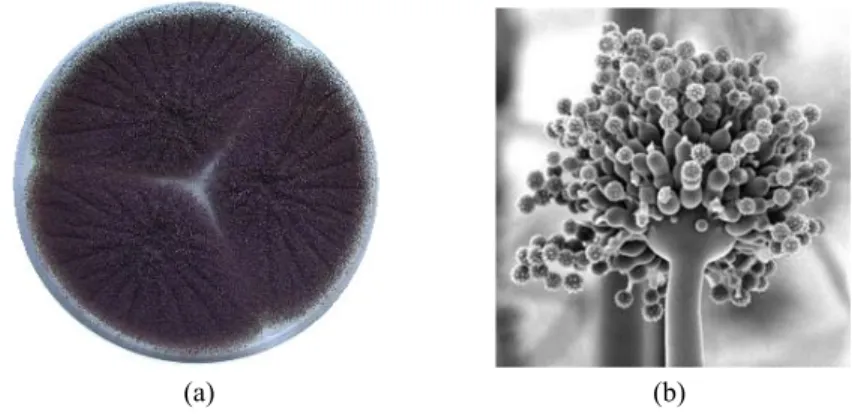
A. niger dapat tumbuh dengan cepat, sehingga dapat digunakan secara komersial dalam produksi asam sitrat, asam glukonat, dan pembuatan beberapa enzim seperti amilase, pektinase, amiloglukosidase, dan selulase. Menurut Villena (2007), A. niger dapat memproduksi enzim lignoselulotik seperti enzim selulase dan enzim xilanase. A. niger dapat tumbuh pada suhu 35 ºC-37 ºC (optimum), 6 ºC-8 ºC (minimum), 45 ºC-47 ºC (maksimum), dan memerlukan oksigen yang cukup (Frazier dan Westhoff 1981).
A. niger memiliki bulu dasar berwarna putih atau kuning dengan lapisan konidiospora tebal berwarna coklat gelap sampai hitam. Kepala konidia berwarna hitam, bulat, cenderung memisah menjadi bagian-bagian yang lebih longgar dengan bertambahnya umur. Konidiospora memiliki dinding yang halus, tetapi juga berwarna coklat. A. niger mempunyai koloni pada medium Czapek’s Dox mencapai diameter 4-5 cm dalam 7 hari dan terdiri dari suatu lapisan yang kompak berwarna putih hingga kuning dan suatu lapisan konidiofor yang lebat yang berwarna coklat tua hingga hitam. Konidia berbentuk bulat hingga semibulat, berwarna coklat, dan memiliki ornamentasi berupa tonjolan dan duri-duri yang tidak beraturan (Gambar 5).
A. niger memerlukan mineral (NH4)2SO4, KH2PO4, MgSO4, urea, CaCl2.7H2O, FeSO4, dan MnSO4.H2O untuk dapat tumbuh dengan baik. Bahan organik dengan kandungan nitrogen tinggi dapat dikomposisi lebih cepat dari pada bahan organik yang rendah kandungan nitrogennya pada tahap awal dekomposisi. Penurunan bahan organik disebabkan pemanfaatan karbon dan nitrogen sebagai sumber energi oleh A. niger untuk bahan penunjang pertumbuhan. A. niger dalam pertumbuhannya dipengaruhi langsung oleh zat makanan yang terdapat dalam substrat. Dalam hal ini molekul sederhana yang terdapat di sekeliling hifa dapat langsung diserap sedangkan molekul yang lebih kompleks harus dipecah dahulu sebelum diserap ke dalam sel.
(a) (b)
Gambar 5. A. niger dalam bentuk spora (a) dan struktur mikroskopis (b) (Stajich 2007)
Berdasarkan penelitian Sumangat (2003), kapang A. niger memiliki kemampuan untuk meningkatkan kadar protein pada kultivasi bahan berserat tinggi dengan adanya penambahan amonium sulfat (ZA) sebagai sumber nitrogen. Penelitian Sumangat (2003) menunjukkan adanya peningkatan kandungan protein pada serat jambu mete sampai 17 % melalui kultivasi dengan penambahan amonium sulfat sebanyak 3 % bahan. Menurut Garaway dan Evans (1984) dan Cochrane (1958) di dalam Risfaheri (1998), A. niger menghasilkan enzim urease yang dapat
digunakan untuk menghidrolisis urea menjadi ion NH4+ dan CO2. Selanjutnya ion NH4+ ini
digunakan untuk pembentukan asam amino yang merupakan penyusun protein.
E.
DELIGNIFIKASI BIOLOGIS
Lignin merupakan polimer tiga dimensi yang terdiri dari unit fenil propan yang diikat dengan ikatan eter (C-O-C) dan ikatan karbon (C-C). Lignin bersifat tahan terhadap hidrolisis karena adanya ikatan arilalkil dan ikatan eter. Lignin dengan struktur tiga dimensi merupakan material yang paling kuat dalam biomassa, serta sangat resisten terhadap degradasi, baik secara biologi, enzimatis, maupun kimia (Judoamidjojo et al. 1989).
Menurut Fridia (1989), secara umum proses delignifikasi biologis merupakan perlakuan pendahuluan terhadap bahan baku, sehingga mempermudah pelepasan hemiselulosa, dan proses ini berfungsi untuk menghilangkan lignin. Pada proses delignifikasi, lignin dihilangkan, sehingga hemiselulosa dan selulosa mudah dihidrolisis. Menurut Anggraini (2003) delignifikasi dilakukan
dengan merendam serbuk tongkol jagung dalam larutan NaOCl 1 % selama 5 jam pada suhu 28 oC. Lignin akan terlarut pada fraksi cairan, sedangkan fraksi padatan merupakan hemiselulosa dan selulosa.
Menurut Fengel dan Wegener (1995), bahan yang dikenai proses delignifikasi selain mengalami penurunan kandungan ligninnya juga mengalami penurunan selulosa dan hemiselulosa. Proses delignifikasi yang baik adalah yang menghasilkan holoselulosa dengan kandungan sisa lignin yang rendah, hilangnya polisakarida minimal, serta terjadi degradasi oksidatif dan hidrolitik selulosa minimal. Johjima et al. (1999) menambahkan bahwa delignifikasi atau degradasi lignin dilakukan oleh kapang Phanerochaete chrysosporium karena mampu menghasilkan lignin peroksidase (LiP) dan manganese peroksidase (MnP) sebagai produk metabolit sekunder. Kedua jenis enzim ini diproduksi untuk memecah lignin sekaligus memenuhi kebutuhan karbon
Selulosa diekstraksi dengan cara merendam bahan hasil proses delignifikasi dalam larutan
NaOH 15 % selama 24 jam pada suhu 28 oC (Anggraini 2003). Hemiselulosa yang terkandung
pada bahan akan terlarut pada bahan tersebut. Fraksi padatan merupakan selulosa tongkol jagung. Pemisahan dilakukan menggunakan kain saring dengan dibilas dengan air berulang kali hingga pH 8. Kemudian dilakukan pencucian dan pengeringan. Pengeringan dilakukan pada suhu 50 oC selama 48 jam.
F.
HIDROLISIS SELULOLITIK
Hidrolisis adalah kelanjutan proses delignifikasi yang dilakukan untuk memecah selulosa menjadi monomer glukosa yang mudah dicerna oleh hewan ruminansia. Degradasi selulosa terjadi akibat adanya produksi enzim selulase yang yang dihasilkan oleh kapang selulolitik. Enzim merupakan katalis yang spesifik, seperti halnya enzim selulase dapat menghidrolisis selulosa dengan sedikit hasil samping.
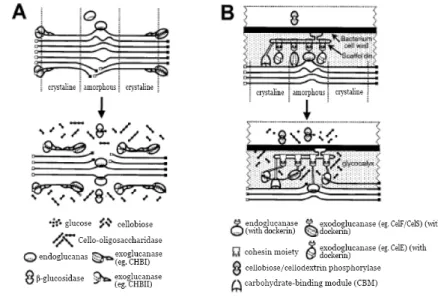
Enzim selulase yang berasal dari kapang merupakan suatu campuran yang terdiri atas tiga enzim yaitu endo β-glukonase, selobiohidrolase, dan β-glukosidase yang bekerja secara sinergi dalam menghidrolisis selulosa berkristal menjadi glukosa. Selobiohidrolase menyerang struktur berkristal selulosa dan menghasilkan selobiosa (disakarida). Endo beta glukonase menghidrolisis bagian amorf selulosa menjadi senyawa–senyawa dengan bobot molekul yang lebih kecil (beta– oligomer menjadi glukosa) (Sasaki 1982).
Menurut Wasserman (1984), hidrolisis selulosa dengan enzim dapat dilakukan dengan dua cara yaitu dengan menggunakan enzim yang telah diisolasi dari sumbernya atau dengan menggunakan mikroba penghasil enzim selulase. Enzim selulase pada umumnya tidak tahan terhadap panas. Aktivitasnya akan hilang sebanyak 50 % dengan pemanasan selama dua jam pada suhu 60 oC. Selulosa merupakan polimer yang tersusun atas unit-unit glukosa melalui ikatan
α-1-4-glikosida dengan bentuk yang menumpuk dan terikat menjadi serat yang sangat kuat.
Selulosa mempunyai dua macam ikatan hidrogen, yaitu hidrogen intramolekul dan ikatan hidrogen intermolekul. Struktur fibril dan kuatnya ikatan hidrogen menyebabkan selulosa bersifat tidak larut dalam berbagai pelarut (Achmadi 1989). Pemutusan ikatan pada selulosa untuk menghasilkan monomer glukosa disajikan pada Gambar 6.
Gambar 6 menunjukkan sistem kerja enzim menghidrolisis selulosa menjadi glukosa dan gula sederhana lainnya. Enzim endoglukanase menghidrolisis secara acak bagian amorf selulosa serat menghasilkan oligosakarida dengan panjang yang berbeda dan terbentuknya ujung rantai
baru. Enzim eksoglukanase bekerja terhadap ujung pereduksi dan non pereduksi rantai polisakarida selulosa dan membebaskan glukosa yang dilakukan oleh enzim glukanohidrolase atau selobiosa yang dilakukan oleh enzim selobiohidrolase. Hidrolisis bagian berkristal hanya dapat dilakukan secara efisien oleh enzim eksoglukanase. Hasil kerja sinergis endoglukanase dan eksoglukanase menghasilkan molekul selobiosa. Hidrolisis selulosa secara efektif memerlukan
enzim β-glukanase yang memecah selobiosa menjadi dua molekul glukosa (Howard 2003: Lynd
et al. 2002).
Hidrolisis dapat terjadi baik pada selulosa maupun hemiselulosa. Hemiselulosa dapat didegradasi menjadi xilosa dengan menggunakan asam dan enzim. Hidrolisis hemiselulosa secara asam dapat dilakukan dengan dua cara yaitu, hidrolisis dengan menggunakan suhu tinggi atau menggunakan asam konsentrasi tinggi (Kosaric et al. 1983). Selain penggunaan asam dan panas, hidrolisis selulosa dan hemiselulosa juga dapat dilakukan dengan pemanfaatan enzim. Menurut Saddler (1993), hidrolisis sempurna xilan memerlukan aktivitas sinergis kelompok enzim hemiselulosa (hidrolitik), di antaranya adalah enzim endo-β-xilanase dan exo-β-xilosidase.
Gambar 6. Hidrolisis selulosa menjadi glukosa (Lynd et al. 2002)
G.
PAKAN TERNAK RUMINANSIA
Pakan ternak ruminansia dapat digolongkan menjadi dua bagian yaitu golongan pakan hijauan dan golongan pakan tambahan berupa konsentrat. Pakan hijauan dapat diperoleh dari jenis rumput, kacang-kacangan, daun pisang, daun nangka, dan hijauan lain. Pakan konsentrat adalah pakan tambahan yang kandungan proteinnya tinggi, diusahakan murah harganya, serta tidak bersaing dengan kebutuhan manusia. Jenis pakan ini dapat diperoleh dari limbah pertanian seperti tongkol jagung, ampas bir, bekatul, onggok, dan tetes. Untuk memenuhi kebutuhan pakan ternak, dikenal istilah complete feed atau pakan lengkap yang berasal dari bahan bukan hijauan. Complete feed merupakan pakan yang terbuat dari limbah pertanian yang diformulasikan sedemikian sehingga semua nutrisi kebutuhan ternak ruminansia bisa dipenuhi. Kandungan
protein dan serat kasar yang dapat memenuhi kebutuhan ternak ruminansia masing-masing sebesar > 8 % dan > 15 % (Riwantoro 2005).
Riwantoro (2005) juga menyatakan bahwa bahan-bahan yang biasa digunakan untuk pembuatan complete feed antara lain : (1) sumber serat kasar (jerami kedelai, tongkol jagung, pucuk tebu dan lain), (2) sumber energi (pollard, dedak padi, bungkil tapioka, tetes dan lain-lain), (3) sumber protein (bungkil kopra, bungkil sawit, bungkil minyak biji kapok atau klenteng, kulit kopi, kulit kakao, urea dan lain-lain), dan (4) sumber mineral (tepung tulang, campuran mineral, garam dapur dan lain-lain).
III. METODOLOGI PENELITIAN
A.
BAHAN DAN ALAT
1.
Bahan
Bahan baku utama dalam penelitian ini adalah tongkol jagung manis kering yang diperoleh dari daerah Leuwiliang, Bogor. Kapang yang digunakan untuk proses delignifikasi adalah fungi pelapuk putih jenis Phanerochaete chrysosporium yang diperoleh dari Laboratorium Patologi, Departemen Silvikultur, Fakultas Kehutanan IPB. Kapang lain yang digunakan adalah kapang selulolitik Aspergillus niger dan Trichoderma viride yang diperoleh dari Laboratorium Bioindustri, Departemen Teknologi Industri Pertanian, Fakultas Teknologi Pertanian IPB.
Bahan-bahan lain yang digunakan dalam proses delignifikasi dan hidrolisis tongkol jagung antara lain, media PDA (Potato Dextrose Agar), PDB (Potato Dextrose Broth), bekatul, spiritus, alkohol, akuades, glukosa, dan sumber mineral untuk media yang meliputi KH2PO4, MgSO4.7H2O, CaCl2.H2O, FeCl3.6H2O, ZnSO4.7H2O, CuSO4.5H2O. Bahan untuk analisis hasil delignifikasi dan hidrolisis adalah air destilata,heksan, H2SO4, NaOH, akuades, fenol, DNS (Dinitrosalisilat), natrium sulfit, Cetyl Trimethyl Amonium Bromida, pereaksi Neutral Detergent Fiber (NDF), enzim α-Amilase, bufer fosfat, dan bahan-bahan lain.
2.
Alat
Peralatan yang digunakan dalam penelitian ini antara lain hammer mills, disc milling cutter, neraca analitik, penyaring 40-60 mesh, oven, otoklaf, inkubator, jarum Ose, plastik wrap, tabung reaksi, cawan petri, labu ukur, labu Erlenmeyer, aluminium foil, desikator, labu didih, tabung Soxhlet, kertas saring, sudip, penjepit, pengaduk kaca, gelas penyaring, kain
saring, magnetic stirrer, gelas kimia, pipet, sumbat kapas, ruang asap, pompa vakum,
pembakar Bunsen, clean bench, dan orbital shaker.
B.
METODE PENELITIAN
1.
Karakterisasi Bahan Baku
Bahan baku berupa tongkol jagung dijemur dan dikeringkan untuk memudahkan proses pengolahan. Selanjutnya tongkol jagung kering dikecilkan ukurannya menggunakan hammer mill hingga ukuran ± 20 mesh dan dilanjutkan dengan disc milling cutter hingga ± 40 mesh. Pada tahap awal ini dilakukan uji proksimat, yaitu uji kadar air, protein kasar, lemak kasar, abu, serat kasar, serta penghitungan karbohidrat (by difference). Selain itu juga dilakukan analisis komponen serat yaitu selulosa, hemiselulosa, dan lignin. Prosedur analisis untuk karakterisasi bahan baku disajikan pada Lampiran 1.
2.
Delignifikasi Secara Biologis
a. Persiapan Substrat
Tongkol jagung berukuran 40 mesh dimasukkan ke dalam labu Erlenmeyer dan
selanjutnya diberi tambahan mineral (Tabel 2) untuk persiapan pertumbuhan P. chysosporium.
Tabel 2. Komponen mineral untuk pertumbuhan P.chrysosporium
Komponen Jumlah (g/150 ml) KH2PO4 7.20 MgSO4.7H2O 1.50 CaCl2.H2O 0.30 FeCl3.6H2O 0.045 ZnSO4.7H2O 0.023 CuSO4.5H2O 0.015 MnSO4.H2O 0.03 Sumber : Fadillah et al., (2008)
Larutan mineral selanjutnya ditambahkan ke dalam bahan dengan perbandingan 1.5 ml/g bahan padatan tongkol jagung. Campuran tongkol jagung dan mineral disterilisasi menggunakan otoklaf dengan suhu 121 oC selama 15 menit.
b. Penyiapan Inokulum
Kultur yang baik dijadikan inokulum adalah kultur yang segar. Untuk menyegarkan dan menumbuhkan kultur kapang yang telah disimpan sebagai stok kultur dalam cawan petri maka perlu dilakukan peremajaan kultur. Biakan Phanerochaete chrysosporium diremajakan dengan cara menginokulasikan kultur pada media agar miring Potato Dextrose Agar (PDA) yang sebelumnya telah disterilkan pada otoklaf selama 30 menit pada suhu 121oC. Selanjutnya kultur diinkubasi selama 7 hari pada suhu ruang (30 oC). Untuk melihat pertumbuhan kapang pada media dilakukan pengamatan secara visual karena penampakan miselia pada media terlihat dengan jelas.
Media yang digunakan untuk perbanyakan kultur adalah media cair Potato Dextrose Broth. Medium sebanyak 225 ml disiapkan, disterilisasi pada suhu 121oC selama 15 menit dalam otoklaf, dan diinokulasi kapang segar setelah suhunya < 30 oC. Inokulasi dilakukan dengan mengambil 1-2 Ose isolat agar miring dan dicampurkan ke dalam media Potato Dextrose Broth (PDB). Inkubasi dilakukan pada shaker dengan kecepatan 140 rpm selama 6 hari.
c. Proses Kultivasi
Sebanyak 150 g tongkol jagung yang telah tercampur dengan 225 ml larutan mineral disiapkan dalam keadaan steril untuk kultivasi. Media steril diinokulasi setelah suhunya di bawah 38 oC. Inokulasi inokulum cair dilakukan dengan perbandingan 0.25 ml/g tongkol jagung kering, sehingga suspensi kapang P. chrysosporium yang diinokulasi adalah sebanyak 37.5 ml untuk 150 g bahan kering. Proses kultivasi dilakukan pada suhu 30 oC selama 20 hari. Pertumbuhan kapang dihentikan dengan sterilisasi bahan dalam otoklaf bersuhu 121 oC selama 30 menit dan sterilisasi sinar ultraviolet selama 2 jam.
d. Proses Hilir
Tongkol jagung hasil delignifikasi diberi perlakuan pencucian dengan akuades
bersuhu 60 oC untuk melarutkan lignin yang terlepas dan meregangkan ikatan
lignoselulosa. Volume air yang digunakan untuk mencuci lignin adalah 500 ml/150 g tongkol awal. Tongkol jagung terdelignifikasi selanjutnya dikeringkan dalam oven pada
suhu 50 oC selama 48 jam. Tongkol jagung hasil pencucian dan pengeringan disebut
sebagai tongkol jagung terdelignifikasi. Bahan ini selanjutnya dianalisis untuk mengetahui perubahan yang terjadi terhadap kadar air, protein kasar, lemak kasar, abu, serat kasar, total gula, gula pereduksi, lignin, selulosa, dan hemiselulosa. Prosedur analisis tongkol jagung terdelignifikasi disajikan pada Lampiran 1. Diagram alir proses delignifikasi dapat dilihat pada Gambar 7 dan Lampiran 2.
3.
Hidrolisis Oleh Kapang Selulolitik
a. Persiapan Substrat
Bahan yang digunakan untuk proses hidrolisis selulolitik adalah tongkol jagung yang sebelumnya telah melalui proses delignifikasi sebanyak 75 g. Serbuk limbah tongkol jagung tersebut dimasukkan dalam wadah plastik, ditambah air sampai kadar 67 %, diberi perlakuan penambahan urea atau penambahan amonium sulfat 3 % (b/b). Selanjutnya bahan disterilisasi dalam otoklaf dengan suhu 121 oC selama 15 menit.
b. Penyiapan Inokulum
Biakan Aspergillus niger dan Trichoderma viride segar diperoleh dengan peremajaan pada media Potato Dextrose Agar (PDA) yang sebelumnya telah disterilkan di otoklaf selama 15 menit pada suhu 121oC. Selanjutnya kultur diinkubasi selama 7 hari pada suhu ruang (30oC).
Media yang digunakan pada perbanyakan kultur untuk penyiapan inokulum adalah media bekatul. Sebanyak 67.5 g bekatul kering direndam dalam 200 ml akuades selama 24 jam dan ditiriskan kemudian dimasukkan ke dalam labu Erlenmeyer dan disumbat kapas. Media disterilisasi pada suhu 121oC selama 15 menit dalam otoklaf selanjutnya diinkubasi pada suhu ruang (30oC) selama beberapa jam sampai siap diinokulasi kapang. Inokulasi kultur A. niger dan T. viride diawali dengan menambahkan 10 ml akuades steril ke dalam biakan (dalam PDA). Selanjutnya miselia dan spora tercampur dalam akuades
dan dituang merata ke dalam media bekatul. Inokulum selanjutnya diinkubasi selama 10 hari pada suhu ruang. Pada kondisi tersebut terlihat adanya pertumbuhan spora pada permukaan bekatul.
c. Proses Kultivasi
Inokulasi dilakukan setelah suhu media tongkol jagung steril berada di bawah 30 oC. Inokulum yang akan digunakan sebelumnya diaduk terlebih dahulu untuk meratakan kultur kapang yang tumbuh di dalam media. Inokulum T. viride dan A. niger diinokulasi masing-masing pada media yang berbeda sesuai dengan perlakuan yang digunakan. Inokulum yang diinokulasi adalah 15 % (b/b) dari bobot tongkol jagung hasil delignifikasi yang digunakan. Kultivasi dilakukan pada inkubator suhu ruang (± 25 oC) selama 9 hari
d. Proses Hilir
Tongkol jagung yang telah dikultivasi selama 9 hari selanjutnya dikeringkan untuk menghindari terjadinya perubahan komposisi bahan akibat aktivitas kapang. Selanjutnya tongkol jagung dianalisis kadar air, protein kasar, abu, lemak kasar, serat kasar, total gula, gula pereduksi, lignin, selulosa, hemiselulosa, dan daya cerna bahan secara in vitro. Prosedur analisis bahan hasil hidrolisis disajikan pada Lampiran 1. Diagram alir proses hidrolisis dapat dilihat pada Gambar 8 dan Lampiran 3.
4.
Rancangan Percobaan
Penelitian ini menggunakan Rancangan Acak Lengkap Faktorial 3 x 2 dengan dua ulangan. Faktor pertama menunjukkan jenis sumber nitrogen, yaitu perlakuan penambahan urea [(CO(NH2)2], perlakuan penambahan amonium sulfat (ZA=[(NH4)2SO4]), dan perlakuan tanpa penambahan sumber nitrogen. Faktor kedua adalah waktu pengambilan sampel dalam kultivasi yaitu hari ke-0 dan hari ke-9. Model Rancangan Percobaan Acak Kelompok adalah sebagai berikut (Mattjik et al. 2006) :
Yijk = u + Ai + Bj + ABij + €k(ij)
Yijk = variable respon dari hasil observasi ke-k yang terjadi karena pengaruh taraf faktor ke-i faktor jenis sumber nitrogen dan tarat ke-j waktu
u = nilai tengah populasi
Ai = efek dari taraf ke-i faktor jenis sumber nitrogen Bj = efek dari taraf ke-j faktor waktu
ABij = efek interaksi antara taraf ke-i faktor jenis sumber nitrogen dan taraf ke-j faktor waktu
€k(ij) = galat percobaan dari perlakuan jenis sumber nitrogen ke-i dan perlakuan waktu ke-j pada pengamatan ke-k.
Perlakuan rancangan percobaan dilakukan pada tahap hidrolisis tongkol jagung dengan pemanfaatan dua jenis kapang yang berbeda. Kapang yang pertama adalah Aspergillus niger
dan yang kedua adalah Trichoderma viride. Pengamatan dilakukan terhadap pembentukan oligosakarida (total gula) dan perubahan kadar protein. Data tersebut kemudian diolah dengan menggunakan program SPSS 15.0 untuk melihat keragaman yang terjadi pada setiap perlakuan dan interaksi antara perlakuan. Uji lanjut Duncan dilakukan jika terjadi pengaruh yang signifikan antara bahan pada perlakuan yang berbeda, sehingga dapat diketahui perbedaan yang terjadi antar level.
Gambar 7. Diagram alir proses delignifikasi tongkol jagung dengan kapang Phanerochaete
chrysosporium Tongkol jagung kering (40 mesh) Campuran mineral Delignifikasi (inkubasi suhu 30 oC, selama 20 hari) Sterilisasi
(suhu 121 oC selama 30 menit)
Inokulum cair P. chrysosporium (25 % v/b) Biakan P. chrysosporium Penyegaran P. chrysosporium (agar cawan petri, suhu 30 oC,
6 hari)
Pembuatan inokulum (media PDB, 6 hari)
Pencucian dengan akuades bersuhu 60 oC
Analisis proksimat, lignin, hemiselulosa, selulosa, gula
pereduksi, total gula, daya cerna in vitro Tongkol jagung hasil delignifikasi
Gambar 8. Diagram alir proses hidrolisis tongkol jagung terdelignifikasi dengan kapang selulolitik.
Inokulasi untuk inokulum (media bekatul, 10 hari)
Penyegaran
(agar miring, suhu 30 oC, 6 hari) Biakan
T. viride / A. niger
Hidrolisis selulosa • (A.niger , T.viride),
• (urea 3% (b/b), ZA 3% (b/b), dan
tanpa sumber nitrogen tambahan) • Suhu kamar, 9 hari, aerobik
• Kadar air 67 % (v/b)
Tongkol jagung hasil delignifikasi
inokulum padat (15 % b/b)
Pengeringan (Oven suhu 50 oC, 48 jam)
Analisa proksimat, lignin, ADF, NDF, gula pereduksi, total gula, dan
IV. HASIL DAN PEMBAHASAN
Tongkol jagung merupakan bahan yang mengandung komponen lignoselulosa yaitu lignin, selulosa, hemiselulosa, dan bahan ekstraktif lainnya. Seluruh komponen lignoselulosa ini merupakan sumber bahan organik yang dapat diperbaharui. Dalam hal produksi bahan pakan, selulosa merupakan komponen dalam tongkol jagung yang dapat dicerna oleh ternak ruminansia. Namun, pemanfaatannya terbatas karena adanya lignin yang merupakan komponen terikat pada selulosa dan tidak dapat dicerna oleh ternak. Proses delignifikasi diharapkan dapat melepaskan lignin, agar hidrolisis kapang selulolitik dapat berlangsung lebih baik. Dengan demikian kandungan selulosa dalam bahan menurun dan dihasilkan gula sederhana yang lebih mudah dicerna.
A.
DELIGNIFIKASI TONGKOL JAGUNG
Bahan baku tongkol jagung merupakan bahan dengan karakteristik tertentu seperti kadar air, abu, serat kasar, protein kasar, dan lemak kasar. Kadar serat yang tinggi pada tongkol jagung terdiri dari lignin, hemiselulosa, dan selulosa. Hasil hidrolisis terhadap selulosa akan menghasilkan gula sederhana. Selain itu tongkol jagung yang dipersiapkan untuk menjadi pakan harus diketahui daya cernanya. Hasil analisis komponen kimia tongkol jagung awal disajikan pada Tabel 3.
Karakteristik tongkol jagung dipengaruhi oleh jenis jagung, lahan penanaman, dan perlakuan yang diberikan selama jagung sebelum dan setelah dipanen. Hasil analisis tongkol jagung yang diperoleh dari penelitian ini menunjukkan adanya perbedaan dengan hasil analisis tongkol jagung yang dilakukan oleh Johnson (1991) pada Tabel 1. Seperti halnya pada pengujian diperoleh kadar serat yang lebih tinggi dan protein kasar yang lebih rendah dibandingkan data pada Tabel 1. Perbedaan ini dapat disebabkan oleh umur jagung yang lebih tua, sehingga menghasilkan kadar serat yang lebih tinggi.
Kadar air merupakan faktor yang sangat berpengaruh terhadap mutu dan daya simpan bahan. Tongkol jagung mempunyai kadar air sebesar 10.71 % (Tabel 3), yang berarti bahwa bahan dalam keadaan relatif kering dan dapat disimpan dalam waktu yang lama. Hasil penelitian juga menunjukkan kadar serat yang tinggi dan protein yang rendah pada tongkol jagung. Keadaan inilah yang menyebabkan rendahnya mutu dan daya cerna ternak terhadap tongkol jagung. Kandungan selulosa sebesar 34.90 % dalam serat menunjukkan pentingnya proses degradasi selulosa. Selain itu, komponen lignin sebesar 10.97 % juga menjadi suatu alasan pentingnya dilakukan proses delignifikasi.
Tongkol jagung merupakan bahan dengan penampang dinding sel yang sangat kompleks dengan lapisan lignin, hemiselulosa, dan selulosa. Karakteristik ini menyebabkan tongkol jagung lebih sulit untuk diuraikan dibandingkan bahan berserat lainnya. Untuk memudahkan pemanfaatan tongkol jagung, pada pendahuluan penelitian dilakukan proses pengecilan ukuran tongkol jagung menjadi ukuran tepung ± 40 mesh. Ukuran yang lebih halus juga bermanfaat untuk memudahkan kapang pendegradasi lignin untuk melepaskan kandungan lignin yang terkandung di dalam tongkol jagung. Keberadaan tongkol jagung dalam keadaan kering bermanfaat untuk memudahkan pengecilan ukuran tongkol jagung sekaligus mencegah tumbuhnya mikroba jenis tertentu sebelum tongkol dimanfaatkan untuk proses penelitian.
Konfigurasi dinding sel tanaman yang terdiri dari lapisan-lapisan lignin, hemiselulosa, dan selulosa (Gambar 1). Lapisan lignin merupakan lapisan paling luar yang membungkus selulosa dan hemiselulosa. Lapisan lignin tidak hanya membungkus selulosa dari satu sisi saja, tetapi secara keseluruhan lignin membungkus selulosa baik dari bagian luar maupun bagian dalam dinding sel tanaman. Pencacahan tongkol jagung akan menyebabkan luas permukaan tumbuh yang lebih besar dan diharapkan meningkatkan kinerja kapang P. chrysosporium untuk mendegradasi lignin dari dinding sel tanaman.
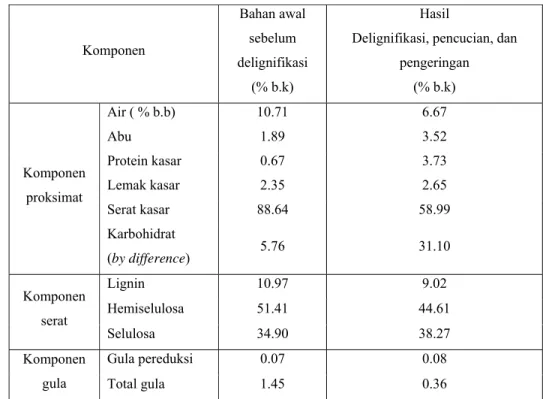
Tabel 3. Perubahan komposisi tongkol jagung sebelum dan setelah delignifikasi
Komponen Bahan awal sebelum delignifikasi (% b.k) Hasil
Delignifikasi, pencucian, dan pengeringan (% b.k) Komponen proksimat Air ( % b.b) 10.71 6.67 Abu 1.89 3.52 Protein kasar 0.67 3.73 Lemak kasar 2.35 2.65 Serat kasar 88.64 58.99 Karbohidrat (by difference) 5.76 31.10 Komponen serat Lignin 10.97 9.02 Hemiselulosa 51.41 44.61 Selulosa 34.90 38.27 Komponen gula Gula pereduksi 0.07 0.08 Total gula 1.45 0.36
Proses delignifikasi dalam penelitian ini dilakukan dengan memanfaatkan kapang pendegradasi lignin jenis pelapuk putih yaitu P. chrysosporium. Kapang P. chrysosporium memiliki kemampuan degradasi lignin yang baik dan hanya menguraikan komponen serat lain dalam jumlah yang kecil. Kapang akan mendegradasi lignin menjadi produk yang larut dalam air dan CO2 (Boyle et al. 1992), sehingga bahan hasil delignifikasi diberi perlakuan pencucian dengan air. Suhu air yang digunakan untuk melarutkan bahan hasil degradasi lignin adalah 60 oC dengan tujuan agar lignin dapat terlarut lebih sempurna dan ikatan lignoselulosa dapat direnggangkan. Renggangnya ikatan lignoselulosa akan memudahkan hidrolisis tongkol jagung. Namun demikian, penggunaan suhu yang terlalu tinggi dapat menyebabkan lignin akan terurai menjadi asam format, metanol, asam asetat, aseton, dan vanillin (Jujoamidjoyo et al. 1989)
Pengaruh degradasi kapang P. chrysosporium terhadap kandungan lignin dalam tongkol jagung dapat diketahui dari analisis lignin yang dilakukan terhadap bahan. Hasil penelitian memperlihatkan adanya penurunan lignin dari bahan awal terhadap bahan hasil delignifikasi (Tabel 3). Tongkol jagung yang pada awalnya mengandung lignin sampai 10.97 % mengalami penurunan sebesar 17.76 % sehingga kadar lignin tongkol jagung setelah delignifikasi menjadi
9.02 %. Selain lignin, hemiselulosa juga mengalami penurunan sampai 13.22 % terhadap kadar hemiselulosa awal. Penurunan kandungan hemiselulosa disebabkan oleh adanya penguraian hemiselulosa oleh kapang P. chrysosporium menjadi ikatan-ikatan yang lebih sederhana. Menurut Perez et al. (2002), kapang P. chrysosporium menghasilkan enzim endoxilanase yang berperan dalam pemecahan xilan menjadi oligosakarida. Xilan adalah karbohidrat utama yang menyusun hemiselulosa.
Hasil analisis menunjukan adanya peningkatan kadar protein kasar dari hasil proses delignifikasi terhadap tongkol jagung. Kadar protein kasar sebesar 0.67 % pada bahan sebelum delignifikasi meningkat menjadi 3.73 % setelah delignifikasi. Adanya miselium kapang yang tersisa di dalam bahan hasil delignifikasi dapat meningkatkan kandungan protein dalam tongkol jagung terdelignifikasi. Menurut Darma (2002), kandungan protein dalam biomassa miselium atau tubuh buah jamur adalah ± 9.25 %.
Berbeda dengan protein, kandungan lemak kasar dalam tongkol jagung terdelignifikasi tidak menunjukkan peningkatan. Dengan demikian dapat dikatakan proses delignifikasi tidak berpengaruh terhadap kadar lemak bahan. Kadar serat kasar mengalami penurunan sebagai akibat dari berkurangnya kandungan lignin dan hemiselulosa dalam bahan. Selain itu penurunan serat juga dapat diperngaruhi oleh terlarutnya komponen serat lain seperti pektin dan gum oleh proses pencucian. Penguraian hemiselulosa menjadi xilan dan gula sederhana lainnya juga menyebabkan turunnya kandungan serat kasar dalam bahan. Gula sederhana dan komponen lain akan terhitung sebagai komponen karbohidrat (by difference). Keadaan inilah yang menyebabkan meningkatnya karbohidrat (by difference) dalam bahan.
Data hasil analisis gula pereduksi pada tongkol jagung sebelum dan setelah delignifikasi tidak menunjukkan perubahan yang cukup signifikan, sehingga dapat diketahui bahwa perlakuan delignifikasi tidak memberi pengaruh yang nyata terhadap peningkatan kadar gula pereduksi pada bahan. Hal tersebut memperlihatkan rendahnya kemampuan kapang P. chrysosporium untuk menghasilkan enzim pendegradasi selulosa menjadi gula sederhana. Berbeda dengan gula pereduksi, analisis total gula menunjukkan penurunan total gula pada tongkol jagung yaitu 1.45 % sebelum delignifikasi dan 0.36 % setelah delignifikasi. Sebenarnya penurunan ini tidak disebabkan oleh proses delignifikasi, tetapi adanya penurunan total gula diakibatkan proses pencucian tongkol jagung terdelignifikasi dengan air panas setelah delignifikasi. Pencucian tongkol jagung hasil delignifikasi melarutkan glukosa, xilan, gula sederhana, serta monosakarida dan disakarida lain yang larut dalam air.
Kapang P. chrysosporium mendegradasi lignin pada fase stasioner yang merangsang
pembentukan enzim. Fase ini diindikasi dengan terlihatnya spora pada media pertumbuhan kapang sebagai akibat kondisi kekurangan nutrisi. Menurut Johjima et al. (1999), kapang P. chrysosporium dapat mendegradasi lignin dan berbagai polutan aromatik selama fase pertumbuhan stasioner yang dipacu oleh kekurangan nutrisi dalam substrat, sehingga menghasilkan dua enzim peroksidase yaitu lignin peroksidase (LiP) dan mangan peroksidase (MnP). Penanaman kapang P. chrysosporium dilakukan dengan pemberian substrat cair untuk mendukung pertumbuhan kapang menjadi lebih baik seperti digambarkan foto pertumbuhan kapang pada Lampiran 4. Dengan lapisan dinding struktur tongkol jagung yang sulit diuraikan, maka penambahan substrat cair sampai 150 % (substrat cair/tongkol jagung) dimaksudkan untuk menumbuhkan kapang secara maksimum di tahap awal, sehingga dihasilkan enzim yang tinggi pada fase stasioner. Selama fase stasioner, enzim LiP akan memecah unit non fenolik yang menyusun 90 % lignin, sedangkan enzim MnP memutus unit fenolik yang menyusun lignin (Johjima 1999).
Gambar 9 memperlihatkan adanya perbedaan struktur mikroskopis tongkol jagung sebelum didelignifikasi dengan tongkol jagung hasil delignifikasi. Gambar (a) merupakan struktur tongkol jagung berukuran 40 mesh yang belum diberi perlakuan apa pun. Struktur yang terlihat terputus-putus adalah akibat pengecilan ukuran tongkol jagung, namun dalam hal ini struktur bahan masih kompleks terdiri dari lapisan lignin, hemiselulosa, dan selulosa. Menurut Lynd (2002), delignifikasi dapat dilakukan dengan pengecilan ukuran bahan, akan tetapi hal tersebut tidak dapat mengubah struktur molekul tongkol jagung secara keseluruhan.
(a) (b)
(c) (d) Gambar 9. Pengamatan mikroskopis (a) tongkol jagung awal, (b) tongkol jagung hasil
delignifikasi, (c) tongkol jagung awal (terpolarisasi), (d) tongkol jagung hasil delignifikasi (terpolarisasi). Perbesaran 400 x.
Gambar 9 (b) merupakan struktur tongkol jagung setelah proses delignifikasi. Struktur yang terlihat pada gambar ini lebih acak dan berongga dibandingkan dengan struktur tongkol jagung awal yang tersusun rapat. Bagian yang berongga adalah struktur tongkol jagung yang telah terpisahkan dari lapisan lignin. Walaupun lapisan lignin tidak terlepas seluruhnya oleh proses delignifikasi, namun ikatan lignoselulosa yang lebih berrongga oleh enzim pendegradasi lignin akan memudahkan degradasi selulosa dan hemiselulosa di lapisan dalam tongkol jagung.
Gambar 9 (c dan d) merupakan penampakan struktur mikroskopis tongkol jagung dengan bantuan cahaya (polarisasi). Kedua gambar ini juga menunjukkan adanya perbedaan yang jelas dari struktur tongkol jagung sebelum dan setelah delignifikasi. Gambar (c) yang merupakan struktur tongkol jagung awal memperlihatkan struktur yang rapat. Hal ini disebabkan oleh tidak adanya rongga yang memperlihatkan struktur penampang yang lebih jelas. Bagian penampang
yang rapat dan berwarna kecoklatan merupakan struktur penyusun bahan yang belum terhidrolisis. Gambar (d) memperlihatkan adanya pantulan cahaya sebagai akibat dari struktur yang longgar dan memungkinkan cahaya tembus dan ditunjukkan bagian berwarna putih. Bagian berongga ini adalah struktur yang awalnya dilapisi oleh lignin, tetapi menjadi longgar karena terlepasnya lignin oleh proses delignifikasi. Selain itu struktur yang kosong disebabkan terlepasnya fraksi terlarut tertentu dalam bahan, baik oleh proses delignifikasi maupun proses pencucian.
B.
HIDROLISIS TONGKOL JAGUNG
Hidrolisis tongkol jagung dilakukan terhadap tongkol jagung hasil delignifikasi dengan tujuan untuk menguraikan komponen selulosa yang terdapat dalam tongkol jagung menjadi gula sederhana. Keberadaan kandungan gula yang meningkat dan komponen selulosa yang berkurang menjadi parameter yang baik untuk pemanfaatan tongkol jagung sebagai pakan ternak.
1.
Perlakuan Hidrolisis
Penelitian ini memanfaatkan dua jenis kapang selulolitik yaitu Aspergillus niger dan Trichoderma viride. Kedua jenis kapang ini memiliki kemampuan untuk menghasilkan enzim selulolitik untuk mendegradasi selulosa ataupun hemiselulosa yang terkandung di dalam tongkol jagung. Karakteristik tongkol jagung dengan kandungan serat yang tinggi membutuhkan kultur kapang yang proporsional dan aktif untuk mendegradasi selulosa. Dengan demikian, diperlukan perbanyakan kultur dalam bentuk inokulum. Substrat inokulum yang digunakan dalam penelitian ini adalah bekatul dengan karakteristik kandungan nitrogen (N) yang tinggi. Kandungan protein dan karbohidrat yang tinggi dalam bekatul diharapkan menjadi subsrat penyedia karbon (C) dan nitrogen (N) untuk perbanyakan inokulum lebih cepat. Komponen kimia dalam bekatul yang digunakan sebagai substrat inokulum disajikan pada Tabel 4.
Kandungan nitrogen yang tinggi dalam substrat inokulum diharapkan dapat mempercepat pertumbuhan kapang selulolitik, sehingga diperoleh inokulum dengan konsentrasi miselium kapang yang baik untuk menghidrolisis tongkol jagung secara langsung. Inokulum yang siap dan baik ditanam ke tongkol jagung adalah inokulum yang pada berada pada pertumbuhan kapang fase stasioner yang ditunjukkan dengan munculnya spora. Menurut Suhartono (1989), organisme pembentuk spora biasanya memproduksi enzim tertentu pada fase pasca eksponensial. Jadi dapat diduga bahwa pada saat aktivitas enzim yang dihasilkan tinggi, maka kapang telah berada pada fase stasioner. Rendahnya sumber C dan N dalam inokulum selama fase stasioner akan mengarahkan kapang untuk menghidrolisis tongkol jagung secara langsung. Dengan demikian dapat diperoleh degradasi komponen lignoselulosa yang lebih baik.
Hidrolisis tongkol jagung ataupun hidrolisis selulosa dapat dilakukan dengan memanfaatkan beberapa cara seperti hidrolisis enzimatik dan hidrolisis asam. Menurut Subekti (2006), hidrolisis enzim lebih banyak menghasilkan gula dibandingkan hidrolisis asam. Hal ini terjadi karena enzim selulase yang dihasilkan mikroba merupakan enzim kompleks, sehingga tongkol jagung dapat dihidrolisis dengan sempurna. Howard (2003) menyatakan
bahwa degradasi selulosa oleh fungi merupakan hasil kerja sekelompok enzim selulolitik yang bekerja secara sinergis. Enzim selulolitik terdiri dari enzim endoglukanase, eksoglukanase, dan β-glukanase. Produksi enzim oleh kapang sebenarnya dipengaruhi oleh pola pertumbuhan mikroba yaitu fase adaptasi, fase eksponensial, dan fase stasioner. Menurut Darwis et al. (1995), pada awal kultivasi aktivitas enzim selulase masih sangat rendah. Aktivitas enzim akan meningkat sejalan dengan bertambahnya waktu kultivasi dan menurun pada hari ke-10.
Tabel 4. Komposisi kimia bekatul (substrat inokulum)
Komponen % b.k Air (% b.b) 9.26 Lemak 16.34 Serat 13.42 Protein 14.56 Abu 12.19 Karbohidrat (by difference) 39.10
2.
Karakteristik Produk Hasil Hidrolisis
Proses kultivasi tongkol jagung terdelignifikasi dengan memanfaatkan kapang selulolitik akan menyebabkan adanya perubahan komposisi bahan selama kultivasi. Perubahan dapat terjadi karena penggunaan nutrisi yang terkandung dalam bahan untuk pertumbuhan kapang. Selain itu kapang juga dapat memanfaatkan menghasilkan enzim yang mendegradasi kandungan serat dalam bahan sebagai sumber nutrisi. Dengan demikian perubahan dapat terjadi pada seluruh komponen yang terkandung dalam tongkol jagung yang dikultivasi.
a. Komponen Proksimat
Analisis proksimat tongkol jagung hasil hidrolisis dilakukan untuk mengetahui adanya perubahan komposisi yang terjadi pada tongkol jagung sebelum dan setelah hidrolisis. Pengamatan yang dilakukan adalah terhadap kadar air, abu, lemak kasar, protein kasar, dan serat kasar. Hasil analisis proksimat terhadap bahan dengan perlakuan sumber nitrogen dan jenis kapang yang berbeda dapat dilihat pada Tabel 5 dan Lampiran 5.
Bahan yang digunakan dalam penelitian dikeringkan, sehingga kadar air bahan berada pada kisaran 3 % sampai 10 %. Bahan dalam keadaan kering bermanfaat untuk menghindari perubahan komposisi jika disimpan dalam jangka waktu yang lebih lama. Keberadaan kapang pada substrat dengan kadar air yang rendah juga dapat menghambat pertumbuhan spora kapang yang tercampur di dalam bahan kering siap uji. Dengan demikian terjadinya hidrolisis lanjut yang berpengaruh terhadap komposisi bahan dapat diminimalkan.
Secara umum kultivasi tongkol jagung memanfaatkan kapang selulolitik menyebabkan terjadinya penurunan kandungan protein dan lemak dalam bahan. Penurunan ini dapat disebabkan oleh pemanfaatan komponen protein dan lemak sebagai nutrisi yang mendukung pertumbuhan kapang selama fase eksponensial. Menurut Fardiaz (1988),