Summary We studied the effects of apical maturation on the vascular cambium of juvenile and mature scions of Larix laricina (Du Roi) K. Koch that had been grafted on seedling rootstocks. Comparisons between the juvenile and mature cam-bium in nuclear genome size, nuclear shape, DNA concentra-tion, number and volume of nucleoli per nucleus, and concentration of extranuclear RNAs, proteins and insoluble carbohydrates were conducted on four occasions during the annual cycle of cambial activity and dormancy. All investigated variables exhibited strong annual oscillations, whereas differ-ences between the two maturation stages were less prominent. Many of the differences between the two phases could be explained by delayed spring reactivation and accelerated onset of dormancy in the mature cambium compared with the juve-nile cambium. At the time of reactivation and during activity, the mature cambium exhibited lower genome size, lower DNA concentration, fewer nucleoli per nucleus and a higher extranu-clear concentration of insoluble carbohydrates than the juve-nile cambium. The dormant mature cambium contained more extranuclear RNAs than the dormant juvenile cambium. The observed differences provide circumstantial evidence of changes in chromatin organization or functioning, or both, during maturation.
Keywords: conifers, cyclophysis, cytology, dormancy, eastern larch, genome size, phase change.
Introduction
Postembryonic ontogeny of both herbaceous and woody plants can be represented as a continuum of juvenile, mature vegeta-tive and reproducvegeta-tive phases. Each phase is characterized by more or less distinctive morphological, anatomical, biochemi-cal and physiologibiochemi-cal features (for recent reviews see Bonga 1982, Hackett 1985, Poething 1990, Greenwood and Hutchison 1993, Hackett and Murray 1993). Transition be-tween the juvenile and mature phases occurs in the apical meristem although it may be affected by factors produced in other plant parts (Wareing and Frydman 1976, Poething 1990). Once a particular phase is attained by the apical meristem, it is conserved during cell division. At present, neither the
induc-tion nor maintenance of phase change is understood. Two mechanisms operating at the chromatin or DNA level have been postulated to account for the conservation of gene expres-sion through successive cell cycles. The first is conservation of cytidine methylation, a well-known feature of the animal king-dom (Weissbach et al. 1989); however, no methylated, matura-tion-specific portions of the genome have been identified. A second mechanism, operating at the level of gene rearrange-ments, has been proposed for Hedera helix L. (Wareing and Frydman 1976, Kessler and Reches 1977, Schäffner and Nagl 1979, König et al. 1987); however, the evidence is contradic-tory and the hypothesis has not been tested in other species. Therefore, we have examined the cambial cells of juvenile and mature Larix laricina (Du Roi) K. Koch to determine whether nuclear DNA changes result in cytologically detectable shifts in nuclear genome size.
There is evidence that the secondary xylem, a direct product of the cambium, is affected by apical maturation (Rumball 1963, Sweet and Maddern Harris 1976, Olesen 1982, Dodd 1988, Dodd and Walker 1988, Takemoto and Greenwood 1993). Although effects of apical maturation on the cambium have not been studied in any species, it is known that the properties of cambial cells vary depending on position in the bole (Bailey 1920, Bannan 1964, Riding and Little 1986, Mellerowicz et al. 1990). For example, in Abies balsamea (L.) Mill., the size of the nuclear genome is 1.3 times larger in 20-year-old cambium from the trunk base than in 1-year-old cambium from the top of the crown (Mellerowicz et al. 1989). Because cambial cells at the top of the crown are derived from a more mature apex than cambial cells at the base of the tree, genome size might reflect the degree of apical maturation. Alternatively, differences in genome size might result from differences in internal conditions between the crown and the bole of the tree, or they might be associated with the course of cambial development with age at a specific location along the bole. To distinguish among these possibilities, we examined cambia produced by juvenile and mature apices of L. laricina following grafting of scions of the same age to seedling root-stocks. In this system, the cambial cells at the time of their formation were at a similar distance from the roots and foliage leaves. The samples collected were from internodes of the
Nuclear and cytoplasmic changes associated with maturation in the
vascular cambium of
Larix laricina
E. J. MELLEROWICZ,
1R. T. RIDING
1,3and M. S. GREENWOOD
21 Department of Biology, University of New Brunswick, Bag Service 45111, Fredericton, N.B. E3B 6E1, Canada 2 Department of Forest Ecosystem Science, 5755 Nutting Hall, University of Maine, Orono, ME 04469, USA 3 Author to whom correspondence should be addressed
Received May 31, 1994
same age. Thus, the major difference between the scions was the stage of maturity of the original donor scion. The nuclear DNA content was assessed at four different stages of cambial activity, because the size of the nuclear genome exhibits annual oscillations in other temperate species (Mellerowicz et al. 1989, 1993, Lloyd et al. 1994). In addition to nuclear DNA content, we investigated nuclear DNA concentration, number and volume of nucleoli, and extranuclear concentrations of RNAs, protein and insoluble carbohydrates.
Materials and methods
One-year-old scions of eastern larch were obtained from the upper crown of 19-year-old trees (mature material) and from their 1-year-old open-pollinated progeny (juvenile material) as described by Takemoto and Greenwood (1993). They were grafted on 2-year-old potted eastern larch rootstocks and grown in an unheated greenhouse for 4 years.
Two families, each comprising a grafted scion from the mother and one from its offspring, were sampled four times during the annual growth cycle: December (fully dormant cambium), March (cambial reactivation), May (fully active cambium) and August (onset of cambial dormancy). The up-permost section of the 2-year-old internode was selected for sampling at the first sampling occasion. At each subsequent sampling, an additional 5-cm long main stem segment was removed from below the most distal long shoot in the selected internode. Decapitation of the plant probably affected cambial behavior, but both juvenile and mature plants received the same treatment and the cambium continued to function. The segments were divided into 1-cm pieces or 0.3-cm slices that were further cut into 2--4 wedges. All pieces were fixed in FAA (formaldehyde/glacial acetic acid/95% ethanol/water, 1/1/14/4, v/v) (Johansen 1940) for 48 h and stored in 70% ethanol until used. The 1-cm pieces were used to prepare cambial cell isolates to examine nuclei, and the slices were used to determine cambial activity and for determination of cambial cytoplasmic RNAs, proteins and carbohydrates.
Nuclear observations
Cambial cell isolates were prepared as described by Mel-lerowicz et al. (1989). In brief, the sample was removed from ethanol, softened in pectinase, and cambial cells were isolated with a needle under a dissecting microscope, spread on a microscope slide and air-dried. For DNA determination, chicken erythrocytes (Sigma) were placed alongside to serve as an internal DNA standard (Dhillon et al. 1977). After rinsing in running, cold tap water to remove traces of formaldehyde, the slides were double stained in Feulgen and naphthol-yel-low S (Berlyn and Miksche 1976). The hydrolysis conditions were 5 N HCl at room temperature for 45 min. The relative DNA content of nuclei was determined cytophotometrically based on three plugs per nucleus to minimize the distributional error (Mellerowicz et al. 1989). Nuclei were treated as ellipses, and the plug diameter was between 50 and 100% of the nuclear width. The accuracy of this method is similar to that of the two-wavelength method (Mellerowicz et al. 1989) and image
analysis (Zhong et al. 1995). Twenty interphase nuclei were measured in each of the four trees per date. The relative DNA content of the cambial nuclei was converted to picograms assuming that each chicken erythrocyte nucleus contained 3.0 pg of DNA (Dhillon 1987).
Samples collected in December, May and August were used for nucleolar observations. Slides containing cambial cell iso-lates were stained with silver nitrate (Howell and Black 1980). To increase contrast, the slides were pre-incubated in 2× so-dium citrate saline at 60 °C for 15 min (Gosálvez et al. 1986). The number and total nucleolar volume per cell were deter-mined for 20 fusiform cells for each of four trees per date.
Cytoplasmic observations
Samples were removed from ethanol, dehydrated through a tertiary-butyl-alcohol series, embedded in Paraplast, sectioned transversely on a rotary microtome at 10 µm and mounted on chemically clean slides (Johansen 1940) without adhesive. The RNAs were stained with azure B (Jensen 1962), total proteins were stained with aniline blue-black (Riding and Little 1986), and total insoluble carbohydrates were stained with the peri-odic acid--Schiff’s (PAS) reaction (Johansen 1940). Concen-trations were measured in relative units of light absorption with a microspectrophotometer equipped with narrow-band light filters having transmission maxima of 600 nm for azure B, 450 nm for aniline blue-black, and 550 nm for PAS (Lillie 1969). Light absorption was measured in 10 plugs per tree per date on sections that did not include the nucleus. Plug size was selected to contain a cambial fusiform cell, including its cyto-plasm and cell walls.
Cambial phenology
The PAS-stained cambial cross sections were used to deter-mine cambial phenology. Presence of recent periclinal divi-sions and expanding cambial derivatives as well as the cambial cell shape and wall thickness were used to assess current cambial activity. An indication of past cambial activity was obtained by counting fully differentiated tracheids from the current growth increment along one radius per sample.
Statistical analysis
Data were subjected to analysis of variance with maturity stage, family and date as random factors, followed by Dun-can’s multiple range test when appropriate.
Results
Cambial phenology
families were active; fusiform cells were thin-walled, radially expanded, and numerous periclinal divisions and expanding derivatives were seen. In August, the juvenile cambium was active in Family 11 and dormant in Family 14, whereas the mature cambium was dormant in both families. In the previous annual increment, the number of tracheids produced per ring by the mature cambium was 63% of that produced by the juvenile cambium (data not shown).
Nuclear shape
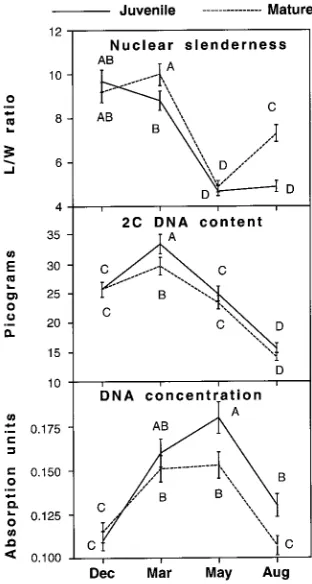
Nuclear length to width ratio (nuclear slenderness ratio) varied with maturation stage and date (Table 1), and there was a significant stage × date interaction. The nuclear slenderness ratio exhibited an annual oscillation of similar amplitude in both maturation stages, with a minimum in May and a maxi-mum in December (Figure 1); however, in the mature cam-bium, the ratio appeared to decrease later and to increase earlier than in the juvenile cambium.
Nuclear DNA content
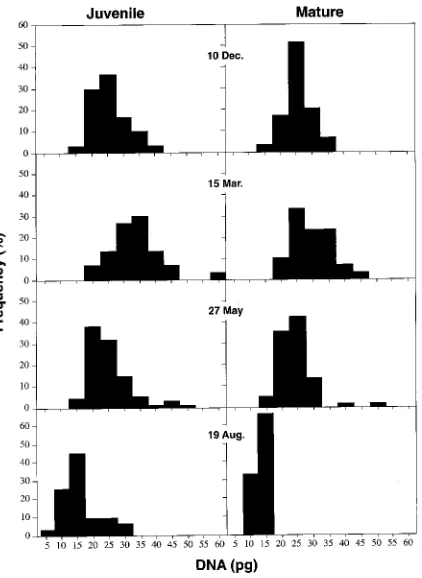
Distributions of nuclear interphase DNA contents were unimo-dal for the juvenile and mature cambia in December and August and for the mature cambium in March (Figure 2). Bimodal distributions were typical for the juvenile cambium in March and both stages of maturation in May. In the bimodal distributions, the larger population, which had the lower DNA values, represented nuclei with a 2C DNA content in the prereplicative phase (G1) of the cell cycle. This was confirmed
by measurements of DNA contents in telophase nuclei (data not shown). The smaller population represented nuclei with a 4C DNA content in a postreplicative phase (G2) of the cell
cycle. No nuclei had DNA contents higher than the 4C DNA value. In the juvenile and mature cambia, the modal values for both G1 and G2 populations showed distinct seasonal shifts,
with maxima on March 15 and minima on August 19. The average 2C DNA content of G1 nuclei varied with
maturation stage, date and family (Table 1 and Figure 1). The seasonal patterns of change in 2C DNA content in the juvenile and mature cambia were similar. The 2C DNA content in-creased from December (2C = 25 pg for both stages) to a maximum in March (2C = 29 pg for mature and 33 pg for juvenile) and then declined to a minimum in August (2C = 15 pg for both stages).
The DNA concentration of G1 nuclei depended on
matura-tion stage, date and family (Table 1). In both juvenile and mature cambia, the concentration was higher in March and May than in August and December (Figure 1). In May and August, the DNA concentration was lower in mature cambium than in juvenile cambium.
Nucleoli
The average number of nucleoli per nucleus was lower in mature cambium than in juvenile cambium as a result of the
Table 1. Probability values for type III analysis of variance.
Variable Factors
Stage Date Family Stage × date
Nuclear slenderness 0.0041 0.0001 0.1031 0.0036
2C DNA content 0.0100 0.0001 0.0165 0.4292
DNA concentration 0.0005 0.0001 0.0017 0.1983
Nucleolar number 0.0395 0.3987 0.8520 0.1998
Nucleolar volume 0.7062 0.0001 0.0782 0.7098
Cytoplasmic RNAs 0.0001 0.0001 0.7769 0.0001
Cytoplasmic proteins 0.2005 0.0001 0.8878 0.0001 Cytoplasmic carbohydrates 0.0003 0.0001 0.0001 0.0006
Figure 1. Seasonal changes in nuclear slenderness (length/width ratio), DNA content and DNA concentration in the G1 fusiform cells of the
low number of nucleoli observed in mature cambium in May (Table 1 and Figure 3). The maximum number of nucleoli per nucleus was five or six in both juvenile and mature cambia, depending on the date (data not shown).Total volume of nucle-oli per nucleus was identical for both maturation stages and varied with date (Table 1 and Figure 3). It was threefold higher in May than in August, with intermediate values observed in December.
Extranuclear RNAs
Concentrations of cytoplasmic and cell wall RNAs varied with maturation stage and date, and the stage × date interaction was significant (Table 1). For both stages, the concentration de-clined from a maximum in December to a minimum in May, and it started to increase again in August (Figure 4). The mature cambium had a higher RNA concentration then the juvenile cambium at all dates except May, when it was similar for both maturation stages.
Extranuclear proteins
Concentrations of cytoplasmic and cell wall proteins varied with date, and there was a significant stage × date interaction (Table 1). In juvenile cambium, protein concentrations de-clined from a high in December to a low in May--August (Figure 4). The protein concentrations in the mature cambium
were similar to those in the juvenile cambium for all dates except March.
Extranuclear insoluble carbohydrates
Concentrations of cytoplasmic and cell wall carbohydrates varied with maturation stage, date and family, and the stage × date interaction was significant (Table 1). The seasonal pattern was similar for both stages. The carbohydrate concentrations were highest in December and declined to a minimum in May (Figure 4). Mature cambium had higher carbohydrate concen-trations than juvenile cambium in May and August.
Discussion
Cambial phenology differed between maturation stages. Ma-ture cambium reactivated later in the year and entered dor-mancy earlier than juvenile cambium. As a result, seasonal changes in nuclear slenderness ratio and DNA concentration (Figure 1), and concentrations of cytoplasmic RNAs, proteins and carbohydrates (Figure 4) were delayed in the mature cambium during reactivation and accelerated during the onset of dormancy compared with seasonal changes in the juvenile cambium. These finding are in agreement with the patterns of seasonal variation in cytological features known to oscillate on an annual basis in the cambium (Bailey 1920, Riding and Little 1986, Mellerowicz et al. 1990). The observations are also consistent with a greater tendency for dormancy in mature apical meristems than in juvenile meristems (Borchert 1976, Franclet 1981, Greenwood 1984).
The nuclear DNA contents in the juvenile and mature cam-bia reflected camcam-bial activity. Thus, active cambium had cy-cling cells in the G1, S and G2 phases with DNA amounts
Figure 2. Distributions of interphase nuclear DNA contents in fusi-form cells of the vascular cambium produced by juvenile and mature scions, grafted on seedling rootstocks, at different times of the year (n = 40).
varying from 2C to 4C, whereas the dormant cambium had cells primarily in the G1 phase with a 2C DNA content
(Fig-ure 2) (cf. Mellerowicz et al. 1989, 1992). There was no evi-dence of DNA endoreduplication, that is, neither juvenile nor mature cambia contained nuclei with DNA higher than 4C. This contrasts with observations in Hedera helix where DNA endoreduplication in foliage and callus tissues is associated with a more mature state of the maternal apex (Kessler and Reches 1977, Polito and Alliata 1981). Thus, DNA endoredu-plication cannot be viewed as an attribute of maturation in general.
The seasonal pattern of changes in 2C DNA content of juvenile and mature cambia, with a maximum in March and a minimum in August (Figures 1 and 2), is similar to the patterns observed in Abies balsamea (Mellerowicz et al. 1989, 1992, Lloyd et al. 1994) and Fraxinus americana L. (Zhong et al. 1995), although the magnitude of the seasonal increase in larch (15 to 32 pg) is the highest observed. The nature of the fraction of the genome that increases is not known. In Abies balsamea, there is evidence that ribosomal DNA is transiently amplified in early spring concomitant with the resumption of cell cycling (Lloyd et al. 1994); however, a different fraction of the genome seems to contribute most to the annual oscillations (Lloyd, Mellerowicz, Riding and Little, unpublished data).
We observed a difference in genome size between the juve-nile and mature cambia during cambial reactivation (March) but not during subsequent activity and dormancy (May--De-cember) (Figure 1). However, because the mature cambium reactivated later than the juvenile cambium, the difference
might be due to a delay in amplification rather than to a difference in the degree of DNA amplification. A more fre-quent sampling schedule might resolve this question. Kessler and Reches (1977) reported that mature shoots of Hedera helix had a smaller genome than juvenile shoots; in addition, mature shoots had a smaller proportion of middle repetitive and AT-rich sequences but a larger proportion of highly repetitive DNA than juvenile shoots. Other researchers did not confirm these observations (Wareing and Frydman 1976, Schäffner and Nagl 1979, König et al. 1987). It is possible that the genome size of Hedera helix oscillates with season as we have observed in Larix laricina, Abies balsamea (Mellerowicz et al. 1989, 1992, Lloyd et al. 1994) and Fraxinus americana (Zhong et al. 1995), thereby confounding the effects of maturation.
The lower springtime DNA amplification and lower number of nucleoli per nucleus observed in mature cambium compared with juvenile cambium (Figures 1 and 3) suggest that a gross change in chromatin organization is involved in maturation (cf. Kessler and Reches 1977, Schäffner and Nagl 1979, Polito and Alliata 1981, Polito and Chang 1984). Such a change could be mediated through DNA methylation or activation of mobile DNA elements (Poething 1990). No major differences in DNA methylation of fully expanded leaves were observed between juvenile and mature larch scions (Greenwood et al. 1989); however, analysis of meristematic tissue and specific DNA fractions might be more informative.
Whatever the nature of the genomic change associated with maturation, it results in modifications of cellular metabolism by affecting gene transcription and protein synthesis. Extranu-clear RNAs, proteins and insoluble carbohydrates all varied with date (Table 1 and Figure 4), similar to trends described for Abies balsamea (Riding and Little 1986). Extranuclear RNAs and carbohydrates also differed between maturity stages (Ta-ble 1). Comparisons between juvenile and mature cambia made for December and May, when both cambia were at similar phases of their annual cycle, showed that mature cam-bium stored more extranuclear RNAs during dormancy and contained more insoluble carbohydrates during active growth than juvenile cambium (Figure 4). A similar difference in RNA content was found in leaves of a Pyrus species (Ali and West-wood 1966). The high carbohydrate content of the mature tissue is consistent with the proposed key role for carbohy-drates in the induction of maturation (Allsopp 1953, Trippi 1990, Materi and Cumming 1991).
A comparison of our findings with the positional effects on cambial cytology reported by Mellerowicz et al. (1993) indi-cated that most differences in the properties of cambial cells between the top and bottom of the bole may be attributed to internal conditions within the bole and not to the maturation stage of the apex at the time the cambium was formed. Thus, staining intensity for RNA, total carbohydrates and proteins was higher at the top (more mature) than at the bottom (more juvenile) of a tree (Riding and Little 1986). Our results suggest that differences in concentrations of RNA and carbohydrate, but not protein, might be due to maturation (Figure 4). Al-though the nuclear genome size was larger in the lower portion of the bole throughout the year (Mellerowicz et al. 1989), the
difference in nuclear genome size between juvenile and mature states was confined to the period of reactivation (Figure 1). The number of nucleoli was higher at the top (more mature) than at the bottom (more juvenile) of a tree (Mellerowicz et al. 1993), whereas the volume of nucleoli exhibited an opposite trend. In contrast, we observed no differences in nucleolar volume be-tween the juvenile and mature scions, although the juvenile material had more nucleoli per nucleus than the mature mate-rial (Figure 3). We conclude that cells of the vascular cambium incorporate changes in maturation state of the apical meristem at the time of cambium formation as well as in situ changes associated with the increase in distance from the apical mer-istem as the bole increases in length and diameter.
References
Ali, N. and M.N. Westwood. 1966. Nucleic acid as related to juvenility in several Pyrus species. Am. Soc. Hortic. Sci. 89:123--131. Allsopp, A. 1953. Experimental and analytical studies of
pterido-phytes. XIX. Investigations on Marsilea. 2. Induced reversion to juvenile stage. Ann. Bot. N.S. 17:37--56.
Bailey, I.W. 1920. The cambium and its derivative tissues. III. A reconnaissance of cytological phenomena in the cambium. Am. J. Bot. 7:417--434.
Bannan, M.W. 1964. Tracheid size and anticlinal divisions in the cambium of Pseudotsuga. Can. J. Bot. 42:603--631.
Berlyn, G.P. and J.P. Miksche. 1976. Botanical microtechnique and cytochemistry. The Iowa State University Press, Ames, Iowa, 326 p. Bonga, J.M. 1982. Vegetative propagation in relation to juvenility, maturity, and rejuvenation. In Cell and Tissue Culture in Forestry. Eds. J.M. Bonga and D.J. Durzan. Martinus Nijhoff Publishers, Hague, pp 387--412.
Borchert, R. 1976. The concept of juvenility in woody plants. Acta Hortic. 56:21--36.
Dhillon, S.S. 1987. DNA in tree species. In Cell and Tissue Culture in Forestry, Vol. 1. Eds. J.M. Bonga and D.J. Durzan. Martinus Nijhoff Publishers, Hague, pp 298--313.
Dhillon, S.S., G.P. Berlyn and J.P. Miksche. 1977. Requirement for an internal standard for microspectrophotometric measurements of DNA. Am. J. Bot. 64:117--121.
Dodd, R.S. 1988. Cyclophytic effects on wood structure in Pinus radiata D. Don. II. Tracheid morphology. Can. J. For. Res. 18:413--420.
Dodd, R.S. and N.K. Walker. 1988. Cyclophytic effects on wood structure in Pinus radiata D. Don. I. Densitometry and grain angle. Can. J. For. Res. 18:406--412.
Franclet, A. 1981. Rajeunissement et propagation végétative des lig-nuex. Annales de Recherches Sylvicals 1980, AFOCEL, Paris, France, pp 11--41.
Gosálvez, J., J. de la Torre, C. García de la Vega and C. López-Fernán-dez. 1986. The effect of double-strength standard saline citrate on silver staining. I. Nucleoli and micronucleoli in the somatic and germ line of the grasshopper Arcyptera fusca (Orthoptera). Can. J. Genet. Cytol. 28:219--226.
Greenwood, M.S. 1984. Phase change in loblolly pine: shoot develop-ment as a function of age. Physiol. Plant. 61:518--522.
Greenwood, M.S., C.A. Hopper and K.W. Hutchison. 1989. Matura-tion in larch. I. Effect of age on shoot growth, foliar characteristics, and DNA methylation. Plant Physiol. 90:406--412.
Greenwood, M.S. and K.W. Hutchison. 1993. Maturation as a devel-opmental process. In Clonal Forestry I: Genetics and Biotechnol-ogy. Eds. M.R. Ahuja and W.J. Libby. Springer-Verlag, New York, pp 14--33.
Hackett, W.P. 1985. Juvenility, maturation, and rejuvenation in woody plants. Hortic. Rev. 7:109--155.
Hackett, W.S. and J.R. Murray. 1993. Maturation and rejuvenation in woody plants. In Forestry Science Series, Vol. 41. Micropropaga-tion of Woody Plants. Ed. M.R. Ahuja. Kluwer Academic Publica-tions, Dordrecht, pp 93--105.
Howell, W.M. and D.A. Black. 1980. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 36:1014--1015.
Jensen, W.A. 1962. Botanical histochemistry. Principles and practice. W.H. Freeman and Company, San Francisco, 408 p.
Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill Book Company, New York, 523 p.
Kessler, B. and S. Reches. 1977. Structural and functional changes of chromosomal DNA during aging and phase change in plants. Chro-mosomes Today 6:237--246.
König, C., I. Ebert and J. Greilhuber. 1987. A DNA cytophotometric and chromosome banding study in Hedera helix (Araliaceae), with reference to differential DNA replication associated with juvenile-adult phase change. Genome 29:498--503.
Lillie, R.D. 1969. H.J. Conn’s biological stains. Williams and Wilkins, Baltimore, 498 p.
Lloyd, A., E.J. Mellerowicz, C. Chow, R.T. Riding and C.H.A. Little. 1994. Fluctuations in ribosomal RNA gene amount and nucleolar activity in the cambial region of Abies balsamea (Pinaceae) shoots during reactivation. Am. J. Bot. 81:1384--1389.
Materi, D.M. and B.G. Cumming. 1991. Effects of carbohydrate deprivation on rejuvenation, apospory, and regeneration in the os-trich fern (Matteuccia struthiopteris) sporophytes. Can. J. Bot. 69:1241--1245.
Mellerowicz, E.J., R.T. Riding and C.H.A. Little. 1989. Genomic variability in the vascular cambium of balsam fir. Can J. Bot. 67:990--996.
Mellerowicz, E.J., R.T. Riding and C.H.A. Little. 1990. Nuclear size and shape changes in fusiform cambial cells of Abies balsamea
during the annual cycle of activity and dormancy. Can. J. Bot. 68:1857--1863.
Mellerowicz, E.J., R.T. Riding and C.H.A. Little. 1992. Periodicity of cambial activity in Abies balsamea. II. Effects of temperature and photoperiod on the size of the nuclear genome in fusiform cambial cells. Physiol. Plant. 85:526--530.
Mellerowicz, E.J., R.T. Riding and C.H.A. Little. 1993. Nucleolar activity in the fusiform cambial cells of Abies balsamea (Pinaceae): effect of season and age. Am. J. Bot. 80:1168--1174.
Olesen, P.O. 1982. The effect of cyclophysis on tracheid width and basic density in Norway spruce. For. Tree Improv. Arbor. Hørsholm 15:1--80.
Poething, R.S. 1990. Phase change and the regulation of shoot mor-phogenesis in plants. Science 250:923--930.
Polito, V.S. and V. Alliata. 1981. Growth of calluses derived from shoot apical meristems of adult and juvenile English ivy (Hedera helix
L.). Plant Sci. Lett. 22:387--393.
Polito, V.S. and Y.-C. Chang. 1984. Quantitative nuclear cytology of English ivy (Hedera helix L.). Plant Sci. Lett. 34:369--377. Riding, R.T. and C.H.A. Little. 1986. Histochemistry of the dormant
vascular cambium of Abies balsamea: changes associated with tree age and crown position. Can. J. Bot. 64:2082--2087.
Schäffner, K.-H. and W. Nagl. 1979. Differential DNA replication involved in transition from juvenile to adult phase in Hedera helix (Araliaceae). Plant Syst. Evol. 2(suppl.):105--110.
Sweet, G.B. and J. Maddern Harris. 1976. Wood properties of Pinus radiata: seed-grown trees compared with grafts from different-aged ortets. N.Z. J. For. Sci. 6:114--121.
Takemoto, Y. and M.S. Greenwood. 1993. Maturation in larch: age-re-lated changes in xylem development in the long-shoot foliage and the main stem. Tree Physiol. 13:253--262.
Trippi, V.S. 1990. Aging of meristems and morphogenetic potentiali-ties. In Plant Aging: Basic and Applied Approaches. Ed. R. Ro-dríguez Plenum Press, New York, pp 3--10.
Wareing, P.F. and V.M. Frydman. 1976. General aspects of phase change, with special reference to Hedera helix L. Acta Hortic. 56:57--69.
Weissbach, A., C. Ward and A. Bolden. 1989. Eukaryotic DNA methy-lation and gene expression. Curr. Top. Cell. Regul. 30:1--21. Zhong, Y., E.J. Mellerowicz, A.D. Lloyd, V. Leinhos, R.T. Riding and