KONSEP TERJADINYA SPESIASI ALOPATRIK KONSEP TERJADINYA SPESIASI ALOPATRIK
Oleh : Oleh : Nama
Nama : : Widya Widya Esti Esti PurwaningtyPurwaningtyasas NIM
NIM : : B1J014108B1J014108 Rombongan
Rombongan : : VIIIVIII Kelompok
Kelompok : : 11 Asisten
Asisten : : Andreanne Andreanne Poppy Poppy EstaniaEstania
LAPORAN PRAKTIKUM SISTEMATIKA HEWAN I LAPORAN PRAKTIKUM SISTEMATIKA HEWAN I
KEMENTERIAN RISET, TEKNOLOGI, DAN PENDIDIKAN TINGGI KEMENTERIAN RISET, TEKNOLOGI, DAN PENDIDIKAN TINGGI
UNIVERSITAS JENDERAL SOEDIRMAN UNIVERSITAS JENDERAL SOEDIRMAN
FAKULTAS BIOLOGI FAKULTAS BIOLOGI PURWOKERTO PURWOKERTO 2016 2016
BAB I.
BAB I. PENDAHULUANPENDAHULUAN
A. Latar Belakang A. Latar Belakang
Kekerabatan dalam sistematik organisme dapat diartikan sebagai pola Kekerabatan dalam sistematik organisme dapat diartikan sebagai pola hubungan atau total kesamaan antara kelompok organisme berdasarkan sifat atau ciri hubungan atau total kesamaan antara kelompok organisme berdasarkan sifat atau ciri tertentu dari masing-masing kelompok organisme tersebut. Berdasarkan jenis data tertentu dari masing-masing kelompok organisme tersebut. Berdasarkan jenis data yang digunakan untuk menentukan jauh dekatnya kekerabatan antara dua kelompok yang digunakan untuk menentukan jauh dekatnya kekerabatan antara dua kelompok organisme, maka kekerabatan dapat dibedakan atas kekerabatan fenetik dan organisme, maka kekerabatan dapat dibedakan atas kekerabatan fenetik dan kekerabatan filogenetik (filetik). Kekerabatan fenetik didasarkan pada persamaan kekerabatan filogenetik (filetik). Kekerabatan fenetik didasarkan pada persamaan sifat-sifat yang dimiliki masing-masing kelompok tumbuhan tanpa memperhatikan sifat-sifat yang dimiliki masing-masing kelompok tumbuhan tanpa memperhatikan sejarah keturunannya, sedangkan kekerabatan filogenetik didasarkan pada sejarah keturunannya, sedangkan kekerabatan filogenetik didasarkan pada asumsi-asumsi evolusi sebagai acuan utama (Clifford & Stephenson, 1975). Pengelompokan asumsi evolusi sebagai acuan utama (Clifford & Stephenson, 1975). Pengelompokan taksa dalam satu klaster menggambarkan kedekatan kekerabatannya. Semakin tinggi taksa dalam satu klaster menggambarkan kedekatan kekerabatannya. Semakin tinggi kemiripan urutan nukleotidanya, semakin tinggi similiritasnya menyebabkan kemiripan urutan nukleotidanya, semakin tinggi similiritasnya menyebabkan posisinya ada dalam percabangan poh
posisinya ada dalam percabangan pohon filogeni menjadi berdekatan (Sahara, 2015).on filogeni menjadi berdekatan (Sahara, 2015). Spesiasi juga membentuk jembatan konseptual antara mikroevolusi, Spesiasi juga membentuk jembatan konseptual antara mikroevolusi, perubahan
perubahan dari dari waktu waktu ke ke waktu waktu dalam dalam frekuensi frekuensi alel alel dalam dalam suatu suatu populasi, populasi, dandan makroevolusi, pola luas evolusi di atas tingkat spesies. Contoh perubahan makroevolusi, pola luas evolusi di atas tingkat spesies. Contoh perubahan makroevolusi adalah asal grup baru organisme, seperti mamalia atau tanaman makroevolusi adalah asal grup baru organisme, seperti mamalia atau tanaman berbunga, melalui serangkaian peristiwa spesiasi (Reece et al., 2015).
berbunga, melalui serangkaian peristiwa spesiasi (Reece et al., 2015).
Evolusi isolasi reproduksi antara populasi sejenis yang berbeda merupakan Evolusi isolasi reproduksi antara populasi sejenis yang berbeda merupakan persyaratan
persyaratan utama utama untuk untuk proses proses spesiasi. spesiasi. Populasi Populasi allopatrik allopatrik pada pada tahap tahap awalawal perbedaan,
perbedaan, oleh oleh karena karena itu, itu, dapat dapat memberikan memberikan kesempatan kesempatan berharga berharga untukuntuk mempelajari timbulnya hambatan reproduksi dan alasan subur untuk pengujian mempelajari timbulnya hambatan reproduksi dan alasan subur untuk pengujian hipotesis mengenai peran adaptasi lingkungan, seleksi seksual dan perbedaan genetik hipotesis mengenai peran adaptasi lingkungan, seleksi seksual dan perbedaan genetik acak dalam menghasilkan hambatan reproduksi antara populasi. Bahkan hambatan acak dalam menghasilkan hambatan reproduksi antara populasi. Bahkan hambatan reproduksi lengkap yang berevolusi selama allopatri mungkin memainkan peran reproduksi lengkap yang berevolusi selama allopatri mungkin memainkan peran penting
penting dalam dalam kasus kasus kontak kontak sekunder sekunder dengan dengan mencegah mencegah campuran campuran populasi populasi dandan semakin memperkuat isolasi seksual sampai spesiasi selesai (Jennings et al., 2011). semakin memperkuat isolasi seksual sampai spesiasi selesai (Jennings et al., 2011).
Meskipun spesies adalah istilah yang berguna dan umum dalam biologi, Meskipun spesies adalah istilah yang berguna dan umum dalam biologi, penggunaannya
penggunaannya bervariasi bervariasi antara antara ahli ahli biologi biologi yang yang tertarik tertarik dalam dalam berbagai berbagai aspekaspek spesiasi perbedaan dari garis keturunan biologis dan munculnya isolasi reproduktif spesiasi perbedaan dari garis keturunan biologis dan munculnya isolasi reproduktif antara garis keturunan. Ahli biologi yang berbeda berpikir tentang spesies berbeda antara garis keturunan. Ahli biologi yang berbeda berpikir tentang spesies berbeda
karena mereka mengajukan pertanyaan yang berbeda, bagaimana kita dapat mengenali dan mengidentifikasi spesies, Bagaimana spesies baru muncul, bagaimana spesies yang berbeda tetap terpisah, mengapa tingkat spesiasi berbeda antara kelompok organisme. Dalam menjawab pertanyaan ini, ahli biologi fokus pada atribut yang berbeda dari spesies, yang mengarah ke beberapa cara berpikir yang berbeda tentang apa spesies dan bagaimana mereka terbentuk (Hillis et al., 2012).
Praktikum kali ini menggunakan satu jenis ikan yaitu ikan baceman yang berasal dari sungai yang berbeda, yang nantinya akan di analisis kekerabatannya dari satu sama lain. Praktikum kali ini akan membandingkan antara ciri yang mirip dengan semua ciri yang digunakan berupa nilai rata-rata kemiripan ciri, sekaligus menunjukan tingkat hubungan kekerabatan antara taksa yang dibandingkan. Nilai rata-rata kemiripan ciri, selanjutnya dapat digunakan untuk menggambar fenogram mengguanakan perangkat lunak ( software) dengan analisis data atau biasa disebut MEGA 5.05.
B. Tujuan
Tujuan dari praktikum konsep terjadinya spesiasi alopatrik adalah: 1. Memahami konsep spesiasi.
2. Memahami konsep spesiasi pada ikan.
3. Menggunakan software aplikasi komputer yang mendukung penelitian tentang konsep terjadinya spesiasi.
BAB II. TINJAUAN PUSTAKA
Menurut Mallet (2001), spesies dalam konsepnya didefinisikan sebagai spesies adalah kelompok perkembangbiakan alami suatu populasi yang terisolasi secara reproduksinya dari kelompok lainnya. Spesies merupakan perkembangbiakan komunitas sejenis yang setiap individu mempunyai total variasi genetik yang tidak cocok dengan komunitas lain ketika bereproduksi. Konsep spesies berdasarkan konsep perkembangbiakan yakni yang diungkapkan oleh Mayr mampu bertahan hingga beberapa kurun waktu walaupun terjadi perbedaan dengan realitas spesies yang ada di zaman ini maupun seterusnya. Konsep lainnya mengenai spesies berdasarkan konsep isolasi mendefinisikan bahwa spesies merupakan kebanyakan populasi yang termasuk individu dengan dua induk organisme tersebut dan
melakukan sistem perkawinan (reproduksi).
Spesiasi merupakan proses pembentukan spesies baru yang berbeda dari spesies sebelumnya melalui proses perkembangbiakan secara natural dalam kerangka evolusi. Spesiasi menjadi fokus biologi evolusioner. Spesiasi sangat terkait dengan evolusi, keduanya merupakan proses perubahan yang berangsur-angsur, sedikit demi sedikit, secara gradual, perlahan tetapi pasti terjadi. Spesiasi lebih ditekankan pada perubahan yang terjadi pada populasi jenis tertentu. Kecepatan spesiasi maupun
kepunahan sebagian tergantung pada ukuran kisaran geografis dari suatu daerah. Daerah yang luas cenderung meningkatkan kecepatan spesiasi dan menurunkan kecepatan kepunahan. Jenis yang terdapat di daerah yang luas akan mengalami spesiasi lebih cepat, sedangkan menurunnya luas area akan meningkatkan kepunahan suatu jenis, jadi menurunkan jumlah jenis yang akan mengalami spesiasi (Widodo, 2007).
Spesiasi atau terbentuknya spesies baru dapat diakibatkan oleh adanya isolasi geografi, isolasi reproduksi, dan perubahan genetika (Campbell et al., 2008). Adapun proses spesiasi ini dapat berlangsung secara cepat atau lama hingga berjuta-juta
tahun. Hal terebut didukung pula oleh Sahara (2015), yang menyatakan bahwa isolasi geografi merupakan salah satu penyebab awal suatu spesia si, yang merupakan contoh dari keragaman genetik. Spesiasi pada organisme disebabkan karena faktor ekologi dan biografi diikuti oleh isolasi reproduksi. Perbedaan genetik mungkin terdapat antar individu dalam populasi, antar populasi di dalam daerah yang sama atau daerah geografi yang berbeda.
Faktor-faktor yang mendorong terjadinya spesiasi menurut Campbell et al., (2008) yaitu :
a. Isolasi geografis
Isolasi geografis merupakan bentuk pembatasan alam yang berupa pemisahan populasi oleh kondisi alam. Hal ini dapat terjadi jika populasi makhluk hidup yang sama bermigrasi dari lingkungan lama menuju lingkungan baru yang terpisah dengan lingkungan awal dan menetap membentuk populasi tersendiri. Jika sistem populasi yang mula-mula kontinyu dipisahkan oleh kondisi geografis sehingga terbentuk hambatan bagi penyebaran spesiesnya, maka sistem populasi yang demikian tidak akan lagi bertukar susunan gen, dan evolusinya berlangsung secara sendiri-sendiri. Seiring dengan berjalannya waktu, kedua populasi tadi akan semakin berbeda sebab masing-masing menjalani evolusi dengan caranya sendiri.
b. Isolasi reproduksi
Isolasi reproduksi merupakan salah satu penghambat untuk terjadinya perkawinan silang. Jika individu-individu dalam suatu populasi berkumpul dalam satu tempat, maka mungkin terjadi kompetisi untuk mendapatkan makanan, tempat maupun pasangan. Kompetisi ini memungkinkan individu yang kalah akan beradaptasi dengan mengembangkan hanya sebagai faktor geografis (isolasi dengan pemisahan fisis) yang sebenarnya populasi itu masih memilki potensi untuk melakukan interbreeding dan mereka sebenarnya masih dapat dikatakan dalam satu spesies. Selanjutnya kedua populasi tersebut begitu berbeda secara genetis sehingga gene flow yang efektif tidak akan berlangsung lagi seandainya bercampur lagi. Jika titik pemisahan itu telah tercapai, maka kedua populasi itu telah menjadi dua spesies yang terpisah.
Menurut Dharmayanti (2011), spesiasi pada tingkat populasi terdiri dari beberapa model yaitu sebagai berikut:
a. Spesiasi Alopatrik
Spesiasi alopatrik merupakan pembentukan spesies baru yang terjadi melalui pemisahan populasi-populasi yang diturunkan dari nenek moyang bersama dalam geografis yang berbeda. Spesiasi alopatrik banyak dibuktikan melalui studi variasi geografi. Spesies yang beranekaragam secara geografis dari seluruh karakter dapat menghalangi pertukaran gen antara spesies simpatrik. Populasi yang terpisah secara geografis dapat terisolasi oleh kemandulan atau perbedaan perilaku dibandingkan dengan populasi yang berdekatan. Populasi yang terisolasi mungkin tidak dapat
melakukan interbreeding jika mereka bertemu, karena bentuknya sangat menyimpang dan kemudian masuk ke dalam simpatrik tetapi tidak terjadi interbreeding . Spesiasi alopatrik merupakan mekanisme isolasi yang terjadi secara gradual.
b. Spesiasi parapatrik (semi geografik)
Spesiasi parapatrik merupakan spesiasi yang terjadi karena adanya variasi frekuensi kawin dalam suatu populasi yang menempati wilayah yang sama. Spesies induk tinggal di habitat yang kontinyu tanpa ada isolasi geografi. Spesies baru terbentuk dari populasi yang berdekatan. Suatu populasi yang berada di dalam
wilayah tertentu harus berusaha untuk beradaptasi dengan baik untuk menjamin kelangsungan hidupnya, dan usaha itu dimulai dengan memperluas daerah ke daerah lain yang masih berdekatan dengan daerah asalnya. Apabila di area yang baru ini terjadi seleksi, maka perubahan gen akan terakumulasi dan dua populasi akan berubah menjadi teradaptasikan dengan lingkungan barunya. Jika kemudian mereka berubah menjadi spesies lain (spesies yang berbeda), maka perbatasan ini akan
diakui sebagai zona hibrid. Dua populasi tersebut akan terpisah, namun secara geografis letaknya berdekatan sepanjang gradien lingkungan. Spesiasi parapatrik tidak memiliki barrier ekstrinsik yang spesifik untuk gene flow. Populasi berlanjut, tetapi populasi tidak kawin secara acak. Individu lebih mudah kawin dengan tetangganya secara geografis daripada individu dalam cakupan populasi yang berbeda. Individu lebih mungkin untuk kawin dengan tetangganya daripada dengan individu yang ada dalam cakupan. Penyimpangan bisa terjadi karena arus gen dikurangi di dalam populasi dan bermacam-macam tekanan pemilihan ke seberang cakupan populasi.
c. Spesiasi Simpatrik
Spesiasi simpatrik adalah pembentukan spesies baru yang terjadi karena tinggal pada daerah yang sama. Menurut Campbell et al., (2004), dalam spesiasi simpatrik, spesies baru muncul di dalam lingkungan hidup populasi tetua. Model spesiasi simpatrik meliputi spesiasi gradual dan spontan. Sebagian besar model spesiasi simpatrik masih dalam kontroversi, kecuali pada model spesiasi spontan dan spesiasi poliploidi yang terjadi pada tumbuhan.
MEGA 5.05 ( Molecular Evolutionary Genetics Analysis) merupakan software yang digunakan untuk menganalisi komparatif statistik dengan berbasis data sekuens molekul berdasarkan prinsip evolusi. MEGA digunakan oleh para ahli biologi di
sejumlah besar laboratorium untuk merekonstruksi sejarah evolusi spesies dan menyimpulkan tingkat dan sifat selektif yang membentuk evolusi gen dan spesies. MEGA digunakan dalam banyak ruang kelas sebagai alat untuk mengajarkan metode yang digunakan dalam bioinformatika evolusi, salah satu fitur kunci MEGA adalah antar muka pengguna grafis yang memfasilitasi visualisasi rinci dan eksplorasi interaktif urutan data, pohon filogenetik dan hasil analisis. Waktu kebutuhan pengguna MEGA telah berkembang karena meningkatnya ketersedian multi-gen dan
genom-skala data yang mengharuskan berulang (Kumar et al., 2012).
Terdapat lima jenis tes yang ada pada menu phylogeny di software MEGA. Jenis test itu diantaranya Construct / Test Maximum Likelihood Tree, Construct / Test Neighbor-Joining Tree, Construct / Test Maximum-Evolution Tree, Construct / Test UPGMA Tree, Construct / Test Maximum Parsimony Tree . Menurut Tamura et al., (2011), masing-masing tes memiliki perbedaan penganalisisan data. Hal tersebut dapat diperinci sebagai berikut:
1. Unweighted Pair-Group Method with Arithmethic mean ( UPGMA )
Mengasumsikan kesamaan laju evolusi antar sequence/ OTU sehingga metode ini mengestimasi nilai rerata (menyamaratakan) nilai distance untuk menggabungkan pasangan OTU dan perhitungan branch length. Construct/ test UPGMA tree biasanya untuk melihat jauh dekatnya kekerabatan namun tidak menggunakan pembobotan semua dianggap sama.
2. Neighbor Joining (NJ)
Menggunakan logika bahwa pohon terbaik adalah pohon yang meminimalisir nilai tree length yang merupakan penjumlahan dari total branch length. Kondisi ini dicapai dengan konstruksi kombinasi pasangan OTU yang memberikan nilai branch length terkecil (neighbor ) diantara seluruh kombinasi yang ada. Construct/ test neighbor-joining tree biasanya melihat jauh dekatnya hubungan kekerabatan dan apabila terdapat karakter yang berbeda walaupun sedikit tetap diberi pembobotan. Neighbor-joining tree dengan UPGMA tree hampir sama perbedannya hanya pada
ada tidaknya pembobotan 3. Minimum Evolution (ME)
Memiliki logika yang sama dengan metode NJ. Perbedaannya terletak pada metode ME menggunakan optimality search criterion sehingga proses penentuan tree length terkecil dilakukan dengan mengevaluasi seluruh kandidat pohon yang mungkin dibuat untuk sejumlah OTU. Dengan demikian, maka metode NJ dapat
dipandang sebagai pendekatan heuristik (cepat namun tidak menjamin) untuk mendapatkan pohon ME. construct/ test minimum-Evolution tree biasanya untuk melihat hasil evolusi namun tidak secara keseluruhan.
4. Maximum Parsimony (MP)
Metode ini memiliki kesamaan konsep dengan metode ME. Namun demikian penentuan pohon dengan tree length terkecil tidak dilakukan berdasarkan matriks distance seperti pada ME. Perhitungan branch length dan tree length pada metode MP didapatkan dari jumlah substitusi minimum antar character state setiap situs pada sequence alignment . Contruct/ test Maximum parsimony biasanya untuk
melihat data mikroevolusi dan makroevolusi hingga terbentuk spesies baru. 5. Maximum Likelihood (ML)
Metode ini menggunakan perhitungan probabilitas kombinasi seluruh character state ancestral yang dapat memberikan bentukan character state seperti pada sequence alignment yang dimiliki saat ini untuk seluruh situs pada setiap OTU. Metode MP dalam hal ini dapat dipandang sebagai pendekatan heuristik terhadap metode ML karena hanya menggunakan kombinasi character state yang memberikan jumlah substitusi minimum untuk menghasilkan character state sequence yang ada pada saat ini. Construct/ test maximum likelihood tree biasanya untuk melihat masing-masing pasangan spesies dilihat dari basa-basa yang sekarang. Maximum likelihood tree hampir sama dengan maximum parsimony tree yaitu untuk melihat evolusi.
BAB III. MATERI DAN METODE
A. Materi
Alat-alat yang digunakan dalam praktikum konsep terjadinya spesiasi allopatrik meliputi bak preparat, pinset, jangka sorong, sterofoam, kertas milimeter blok, jarum pentul dan kamera.
Bahan yang digunakan dalam praktikum konsep terjadinya spesiasi allopatrik adalah beberapa spesimen hewan ikan baceman ( Hemibagrus nemurus) dari Sungai Serayu, Sungai Klawing dan Sungai Banjaran.
B. Metode
Metode yang dilakukan dalam praktikum antara lain:
1. Preparat diletakkan di atas kertas millimeter blok yang telah dialasi dengan sterofoam.
2. Preparat diukur morfologi tubuhnya berdasarkan titik-t itik patokan dan hasilnya dicatat.
3. Preparat dihitung jumlah perbedaan pada morfologi dibandingkan dengan preparat lain.
4. Dibuat pohon filogenetik dari jumlah perbedaan morfologi prepar at yang sudah dihitung sebelumnya.
5. Hubungan kekerabatan antar preparat dianalisis lebih lanjut menggunakan software MEGA 5.05.
BAB IV. HASIL DAN PEMBAHASAN A. Hasil
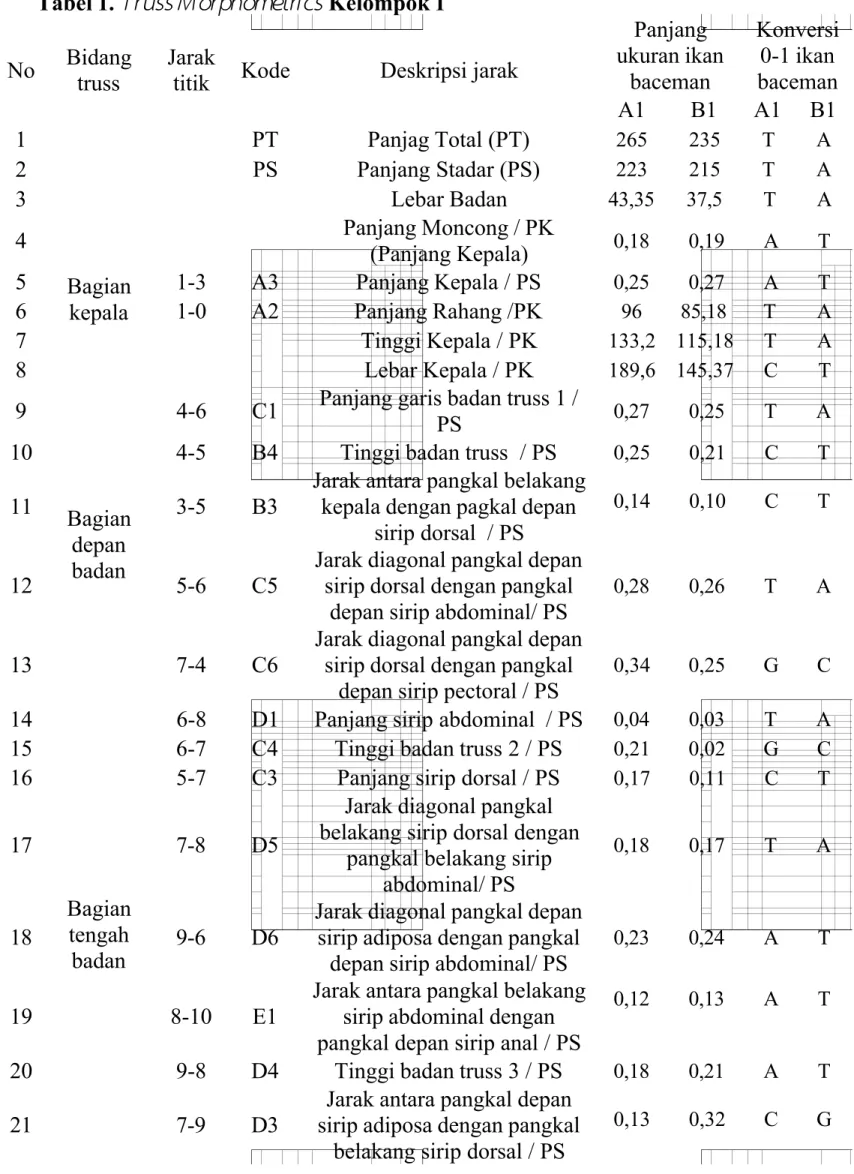
Tabel 1.
Truss M orphometrics
Kelompok 1No Bidang truss
Jarak
titik Kode Deskripsi jarak
Panjang ukuran ikan baceman Konversi 0-1 ikan baceman A1 B1 A1 B1 1 PT Panjag Total (PT) 265 235 T A 2 PS Panjang Stadar (PS) 223 215 T A 3 Lebar Badan 43,35 37,5 T A 4 Bagian kepala Panjang Moncong / PK (Panjang Kepala) 0,18 0,19 A T 5 1-3 A3 Panjang Kepala / PS 0,25 0,27 A T 6 1-0 A2 Panjang Rahang /PK 96 85,18 T A 7 Tinggi Kepala / PK 133,2 115,18 T A 8 Lebar Kepala / PK 189,6 145,37 C T 9 Bagian depan badan
4-6 C1 Panjang garis badan truss 1 /
PS 0,27 0,25 T A
10 4-5 B4 Tinggi badan truss / PS 0,25 0,21 C T
11 3-5 B3
Jarak antara pangkal belakang kepala dengan pagkal depan
sirip dorsal / PS
0,14 0,10 C T
12 5-6 C5
Jarak diagonal pangkal depan sirip dorsal dengan pangkal
depan sirip abdominal/ PS
0,28 0,26 T A
13 7-4 C6
Jarak diagonal pangkal depan sirip dorsal dengan pangkal
depan sirip pectoral / PS
0,34 0,25 G C
14
Bagian tengah badan
6-8 D1 Panjang sirip abdominal / PS 0,04 0,03 T A
15 6-7 C4 Tinggi badan truss 2 / PS 0,21 0,02 G C
16 5-7 C3 Panjang sirip dorsal / PS 0,17 0,11 C T
17 7-8 D5
Jarak diagonal pangkal belakang sirip dorsal dengan
pangkal belakang sirip abdominal/ PS
0,18 0,17 T A
18 9-6 D6
Jarak diagonal pangkal depan sirip adiposa dengan pangkal
depan sirip abdominal/ PS
0,23 0,24 A T
19 8-10 E1
Jarak antara pangkal belakang sirip abdominal dengan pangkal depan sirip anal / PS
0,12 0,13 A T
20 9-8 D4 Tinggi badan truss 3 / PS 0,18 0,21 A T
21 7-9 D3
Jarak antara pangkal depan sirip adiposa dengan pangkal
belakang sirip dorsal / PS
22
Bagian tengah badan
9-10 E5
Jarak diagonal pangkal depan sirip adiposa dengan pangkal
depan sirip anal / PS
0,16 0,13 T A
23 11-8 E6
Jarak diagonal pangkal belakang sirip adiposa dengan
pangkal depan sirip abdominal / PS 0,18 0,22 T C 24 Bagian belakang badan
12-10 F1 Pangkal sirip anal / PS 0,09 0,09 A A
25 11-10 E4 Tinggi badan truss 4 / PS 0,18 0,13 T C
26 11-9 E3 Panjang sirip adiposa / PS 0,15 0,02 G C
27 11-12 F5
Jarak diagonal pangkal belakang sirip adiposa dengan
pangkal belakang sirip anal / PS
0,12 0,08 C T
28 13-10 F6
Jarak diagonal pelipatan ekor bagian ventral dengan pangkal sirip anal / PS
0,21 0,17 C T 29 Bagian pangkal ekor 12-14 G1
Jarak antara pangkal belakang sirip anal dengan pelipatan
ekor bagian ventral / PS
0,17 0,06 G C
30 13-14 G3
Jarak antara pelipatan ekor bagian ventral dengan pelipatan ekor bagian dorsal /
PS
0,14 0,07 G C
31 11-13 F3
Jarak antara pelipata ekor bagian dorsal dengan pangkal
belakang sirip adiposa / PS
0,12 0,07 C T
32 11-14 G5
Jarak diagonal pangkal belakang sirip adiposa dengan pelipatan ekor bagian ventral /
PSJ
0,18 0,11 G C
32 13-12 F4
Jarak diagonal pelipatan ekor bagian dorsal dengan pangkal
belakang sirip anal / PS
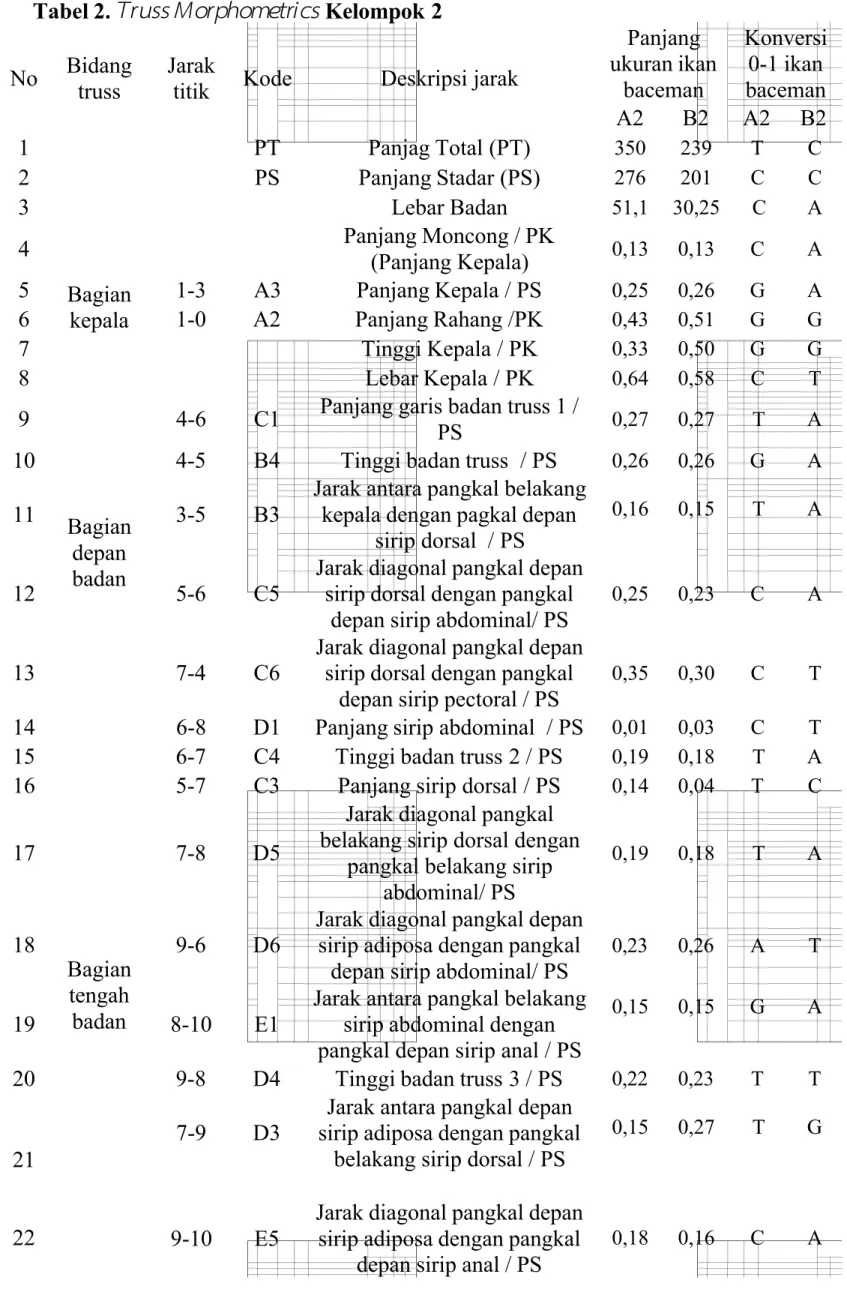
Tabel 2.
Truss Morphometrics
Kelompok 2No Bidang truss
Jarak
titik Kode Deskripsi jarak
Panjang ukuran ikan baceman Konversi 0-1 ikan baceman A2 B2 A2 B2 1 PT Panjag Total (PT) 350 239 T C 2 PS Panjang Stadar (PS) 276 201 C C 3 Lebar Badan 51,1 30,25 C A 4 Bagian kepala Panjang Moncong / PK (Panjang Kepala) 0,13 0,13 C A 5 1-3 A3 Panjang Kepala / PS 0,25 0,26 G A 6 1-0 A2 Panjang Rahang /PK 0,43 0,51 G G 7 Tinggi Kepala / PK 0,33 0,50 G G 8 Lebar Kepala / PK 0,64 0,58 C T 9 Bagian depan badan
4-6 C1 Panjang garis badan truss 1 /
PS 0,27 0,27 T A
10 4-5 B4 Tinggi badan truss / PS 0,26 0,26 G A
11 3-5 B3
Jarak antara pangkal belakang kepala dengan pagkal depan
sirip dorsal / PS
0,16 0,15 T A
12 5-6 C5
Jarak diagonal pangkal depan sirip dorsal dengan pangkal
depan sirip abdominal/ PS
0,25 0,23 C A
13 7-4 C6
Jarak diagonal pangkal depan sirip dorsal dengan pangkal
depan sirip pectoral / PS
0,35 0,30 C T
14
Bagian tengah badan
6-8 D1 Panjang sirip abdominal / PS 0,01 0,03 C T
15 6-7 C4 Tinggi badan truss 2 / PS 0,19 0,18 T A
16 5-7 C3 Panjang sirip dorsal / PS 0,14 0,04 T C
17 7-8 D5
Jarak diagonal pangkal belakang sirip dorsal dengan
pangkal belakang sirip abdominal/ PS
0,19 0,18 T A
18 9-6 D6
Jarak diagonal pangkal depan sirip adiposa dengan pangkal
depan sirip abdominal/ PS
0,23 0,26 A T
19 8-10 E1
Jarak antara pangkal belakang sirip abdominal dengan pangkal depan sirip anal / PS
0,15 0,15 G A
20 9-8 D4 Tinggi badan truss 3 / PS 0,22 0,23 T T
21
22
7-9 D3
Jarak antara pangkal depan sirip adiposa dengan pangkal
belakang sirip dorsal / PS
0,15 0,27 T G
9-10 E5
Jarak diagonal pangkal depan sirip adiposa dengan pangkal
depan sirip anal / PS
23
Bagian tengah
badan 11-8 E6
Jarak diagonal pangkal belakang sirip adiposa dengan
pangkal depan sirip abdominal / PS 0,28 0,24 G T 24 Bagian belakang badan
12-10 F1 Pangkal sirip anal / PS 0,11 0,07 A T
25 11-10 E4 Tinggi badan truss 4 / PS 0,16 0,14 T A
26 11-9 E3 Panjang sirip adiposa / PS 0,11 0,03 T C
27 11-12 F5
Jarak diagonal pangkal belakang sirip adiposa dengan
pangkal belakang sirip anal / PS
0,10 0,09 A A
28 13-10 F6
Jarak diagonal pelipatan ekor bagian ventral dengan pangkal sirip anal / PS
0,28 0,25 G A 29 Bagian pangkal ekor 12-14 G1
Jarak antara pangkal belakang sirip anal dengan pelipatan
ekor bagian ventral / PS
0,14 0,14 T A
30 13-14 G3
Jarak antara pelipatan ekor bagian ventral dengan pelipatan ekor bagian dorsal /
PS
0,13 0,11 T A
31 11-13 F3
Jarak antara pelipata ekor bagian dorsal dengan pangkal
belakang sirip adiposa / PS
0,15 0,16 C T
32 11-14 G5
Jarak diagonal pangkal belakang sirip adiposa dengan
pelipatan ekor bagian ventral / PSJ
0,19 0,19 C A
32 13-12 F4
Jarak diagonal pelipatan ekor bagian dorsal dengan pangkal
belakang sirip anal / PS
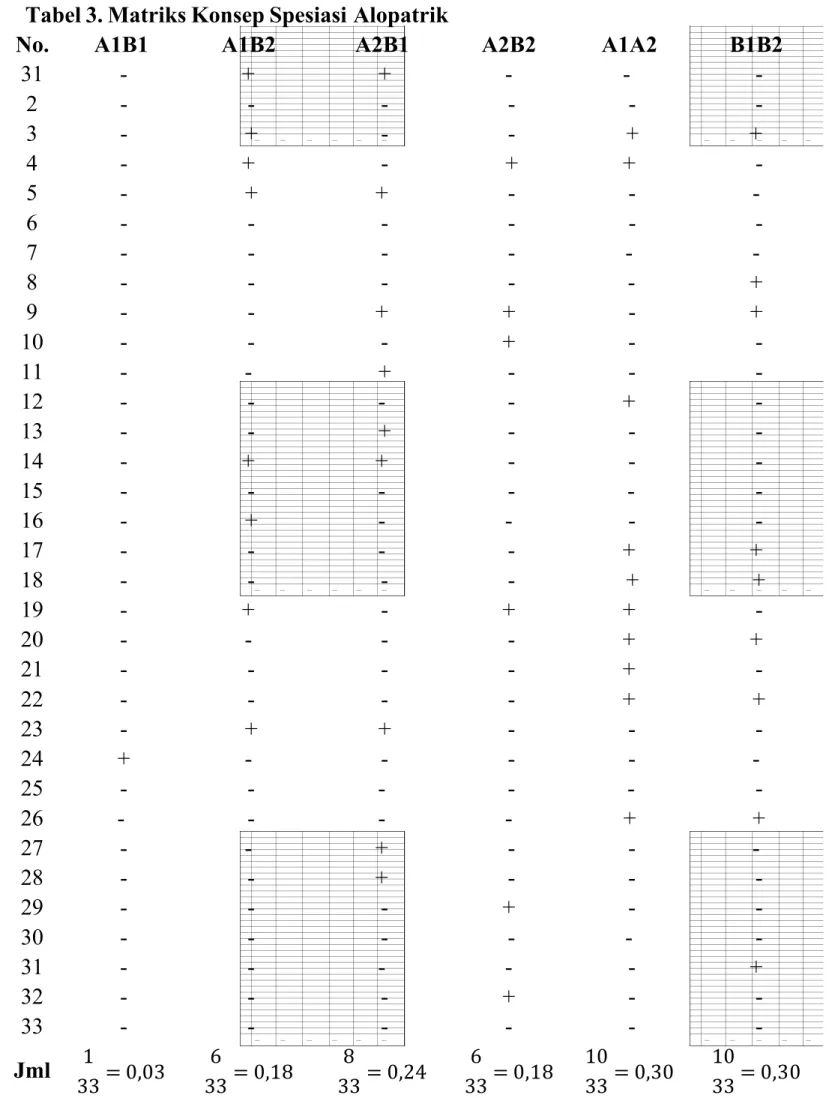
Tabel 3. Matriks Konsep Spesiasi Alopatrik
No. A1B1 A1B2 A2B1 A2B2 A1A2 B1B2
31 - + + - - -2 - - - -3 - + - - + + 4 - + - + + -5 - + + - - -6 - - - -7 - - - -8 - - - + 9 - - + + - + 10 - - - + - -11 - - + - - -12 - - - - + -13 - - + - - -14 - + + - - -15 - - - -16 - + - - - -17 - - - - + + 18 - - - - + + 19 - + - + + -20 - - - - + + 21 - - - - + -22 - - - - + + 23 - + + - - -24 + - - - - -25 - - - -26 - - - - + + 27 - - + - - -28 - - + - - -29 - - - + - -30 - - - -31 - - - + 32 - - - + - -33 - - - -Jml 1 33 = 0,03 6 33 = 0,18 8 33 = 0,24 6 33 = 0,18 10 33 = 0,30 10 33 = 0,30
0,3 0,3 A1 A2 B2 B1 Tabel 4. Matriks Konsep Spesiasi Alopatrik
A1 A2 B1 B2
A1 ………. - -
-A2 0,30 ……….. -
-B1 0,03 0,24 ……..
-B2 0,18 0,18 0,30 ……
Gambar 1. Fenogram Spesiasi Alopatrik Secara Manual
Gambar 2. Fenogram Spesiasi Alopatrik Hasil MEGA 5.05 (UPGMA)
Gambar 3. Spesiasi Alopatrik Hasil MEGA 5.05 (
Maximum Parsimony
)Keterangan :
A = Ikan Baceman Asal Sungai Serayu B = Ikan Baceman Asal Sungai Banjaran
A1 A2 B1 B2 A1 A2 B1 B2
B. Pembahasan
Praktikum kali ini digunakan spesimen Ikan Baceman ( Hemibagrus nemurus) yang berasal dari Sungai Serayu (A), Sungai Banjaran (B), dan Sungai Klawing (C). Kelompok 1 dan 2 mendapatkan ikan yang berasal dari Sungai Serayu (A) dan Sungai Banjaran (B). Ikan Baceman tersebut diukur menggunakan metode truss morphometrics. Ikan Baceman ditempatkan diatas millimeter blok, kemudian ditentukan titik-titik morfometrinya. Terdapat 14 titik, diantaranya (0) titik rahang paling posterior, (1) titik anterior moncong, (2) titik operculum bagian ventral, (3) titik operculum bagian dorsal, (4) titik permulaan sirip pectoral, (5) titik permulaan sirip dorsal, (6) titik pangkal depan sirip abdominal, (7) titik akhir sirip dorsal, (8) titik akhir sirip abdominal, (9) titik permulaan sirip adiposa, (10) titik permulaan sirip anal, (11) titik akhir sirip adiposa, (12) titik akhir sirip anal, (13) titik akhir sirip caudal dorsal, (14) titik akhir sirip caudal ventral. Titik-titik tersebut saling dihubungkan dan akan menghasilkan 33 panjang ukuran. Dikarenakan keterbatasan waktu maka pengukuran dilakukan oleh asisten dan telah disediakan tabel pengukuran morfometri.
Tabel pengukuran morfometri kemudian di analisa perbandingan Ikan Beceman dari Sungai Serayu dan Sungai Banjaran. Hasil pengukuran morfometri, selanjunya dikonversikan ke dalam bentuk basa-basa nitrogen (A, T, G, C). Apabila perbandingan bernilai bilangan bulat 1-30 maka dikonversikan ke dalam basa A
(untuk nilai yang lebih kecil) dan T (untuk nilai yang lebih besar), perbandingan bernilai 31-60, maka dikonversikan kedalam basa T (untuk nilai yang lebih kecil)
dan C (untuk nilai yang lebih besar). Begitu juga apabila hasil >60, maka akan dikonversikan ke dalam basa C (untuk nilai yang lebih kecil) dan G (untuk nilai yang lebih besar). Pengukuran dengan melibatkan nilai desimal, maka yang bernilai 0 dikonversikan kedalam basa A. Apabila selisih nilai desimalnya 1-3, maka dikonversikan kedalam basa A (untuk nilai yang lebih kecil) dan T (untuk nilai yang lebih besar), selisih nilai desimalnya 4-6, maka dikonversikan kedalam basa T (untuk nilai yang lebih kecil) dan C (untuk nilai yang lebih besar). Sedangkan untuk selisih nilai desimal mencapai >7, maka dikonversikan kedalam basa C (untuk nilai yang lebih kecil) dan G (untuk nilai yang lebih besar).
Nilai yang telah dikonversikan menjadi rangkaian basa nitrogen kemudian saling dibandingkan A1, A2, B1 dan B2 yang merupakan kode Ikan Baceman dari
Sungai Surayu dan Ikan Baceman dari Sungai Banjaran. Hasil perbandingan yang antara A1B1, A1B2, A2B1, A2B2, A1A2, dan B1B2 yang memiliki basa nitrogen yang sama diberi tanda positif (+) pada tabel matriks konsep spesiasi alopatrik, sedangkan basa nitrogen yang saling berbeda diberi tanda negatif (-). Jumlah tanda (+) yang melambangkan persamaan kemudian dihitung berkolom tabel dan dibagi dengan jumlah titik yang ada. Kemudian hasil dari perhitungan tersebut dimasukkan pada matriks konsep spesiasi alopatrik untuk selanjutnya dibuat fenogram.
Pembuatan fenogran berpedoman pada nilai matriks konsep spesiasi alopatrik. Nilai yang lebih besar menandakan hubungan kekerabatan yang lebih dekat. Berdasarkan hasil perhitungan kelompok 1 dan 2 rombongan VIII, nilai terbesar adalah nilai antara A1 dengan A2 dan B1 dengan B2 yaitu sebesar 0,3. Sehingga kekerabatan antara A1 dan A2 lebih dekat daripada antara A1 dengan B1 ataupun sebaliknya. Maka percabangan dalam fenogram pun dibuat lebih dekat. Kemudian dilihat nilai yang terbesar kedua pada matriks yaitu antara A2 dengan B1 sebesar 0,24, nilai ini digunakan untuk menentukan percabangan berikutnya. Dengan begitu maka dapat menentukan fenogramnya.
Tahapan-tahapan penggunaan software MEGA 5.05 pada praktikum ini yaitu sebagai berikut.
1. Urutan basa hasil pengamatan disalin di Microsoft Word dengan format yang sudah ditentukan.
2. Buka software MEGA 5.05 dan pilih Align, kemudian Edit built alignment dan Create a new alignment .
3. Pada jendela baru akan terdapat beberapa pilihan, karena data yang dimiliki adalah rangkaian basa nitrogen maka pilih DNA, kemudian pilih edit dan Insert blank sequences.
4. Maka akan muncul kotak berwarna biru dan urutan basa nitrogen yang telah disalin pada Microsoft word kemudian di salin kembali pada kotak tersebut, kemudian ganti nama sequence.
5. Simpan data dengan cara pilih menu data, export alignment , dan pilih export kemudian MEGA format, save.
6. Buka software MEGA 5.05, klik data lalu pilih open a file, pilih file yang tadi disimpan, kemudian pilih open.
7. Untuk melihat pohon filogeni berdasarkan data yang dimiliki maka pilih menu Phylogeny. Terdapat 5 tipe pohon filogeni yang dapat digunakan sesuai dengan
kebutuhan.
8. itu test of phylogeny diganti dengan Bootstrap method. Dan No of Bootstrap replication diganti dengan 1000. Lalu pilih compute.
9. Selanjutnya akan muncul fenogram baru dan klik image dan pilih copy to clipboard lalu kembali ke Ms. Word lalu klik paste.
Berdasarkan hasil menggunakan software MEGA 5.05, A1 dengan A2 dan B1 dengan B2 memiliki kekerabtan yang lebih dekat. Hal ini juga sesuai dengan dengan hasil pada pembuatan fenogram secara manual. A1 lebih dekat hubungan
BAB V. KESIMPULAN DAN SARAN
A. Kesimpulan
Berdasarkan hasil dan pembahasan, dapat disimpulkan bahwa:
1. Konsep spesiasi adalah pemisahan suatu organisme karena faktor eksternal (geografi) ataupun internal (genetik dan reproduksi) sehingga terbentuk spesies baru.
2. Konsep spesiasi pada ikan adalah dengan menggunakan Truss Morfometrics untuk bahan pembanding antar ikan.
3. Software MEGA 5.05 dapat digunakan untuk menentukan fenogram hubungan kekerabatan dengan data hasil Truss Morfometrics yang dikonversikan.
B. Saran
Saran untuk praktikum kali ini adalah membutuhkan keahlian teknik truss morfometric yang benar karena data yang kita gunakan dapat mempengaruhi hasil pada saat memasukkan data ke MEGA.
DAFTAR REFERENSI
Campbell, A. N. 2004. Biologi Edisi Kelima Jilid Kedua. Jakarta: Erlangga.
Campbell, N. A., Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V. dan Jackson, R. B. 2008. Biologi Jilid 2. 8th Edition. San Fransisco: Pearson Education Inc.
Clifford dan Stephenson. 1975. An Introduction to Numerical Classification. New York: Academic Press.
Dharmayanti, I. 2011. Filogenetika Molekuler: Metode Taksonomi Organisme Berdasarkan Sejarah Evolusi. WARTAZOA. 22(1): 25-31.
Hillis D.M., Heller H.C., Sadava D., dan Price M.V. 2012. Principles od Life. Sunderland: Sinauer Associates.
Jennings J.H., Mazzi D., Ritchie M.G., dan Hoikkala A. 2011. Sexual And Postmating Reproductive Isolation Between Alopatrik Drosophila Montana Populations Suggest Speciation Potential. BMC Evol. Biol., 11(68): 1-10. Kumar, S., Stecher, G., Peterson, D.,Tamura, K. 2012. MEGA-CC: Computing Core
of Molecular Evolutionary Genetics Analysis Program for Automated and Iterative Data Analysis. Bioinformatics Application Note. 28 (20) : 2685 –
2686.
Mallet, J. 2001. Species, Concept of . Encyclopedia of Biodiversity. London: Academic Press.
Reece J.B., Urry L.A., Cain M.L., Wasserman S.A., Minorsky P.V., dan Jackson R.B. 2015. Campbell Biology : Tenth Edition. San Francisco: Pearson Education.
Sahara, A., Prastowo, J., Wirdayanti, R., Kurniasih, Nurcahyo, W. 2015. Kekerabatan Genetik Caplak Rhiphicephalus ( Boophilus) microplus Asal IndonesiaBerdasarkan Sekuen Internal Transcribed Spacer-2. Jurnal Veteriner 16(3): 310-319.
Tamura., K, Peterson., D, Peterson., N, Stecher., G, Nei., M dan Kumar., S. 2011. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance and Maximum Parsimony Methods. Journal of Molecular Biology and Evolution, 28(10): 2731 – 2739.
Widodo Pudji. 2007. Spesiasi pada Jambu-Jambuan (Myrtaceae): Model Cepat dan Lambat. Biodiversitas. 8(1): 79-82.