Directory UMM :Data Elmu:jurnal:B:Brain Research:Vol884.Issue1-2.2000:
Teks penuh
Gambar
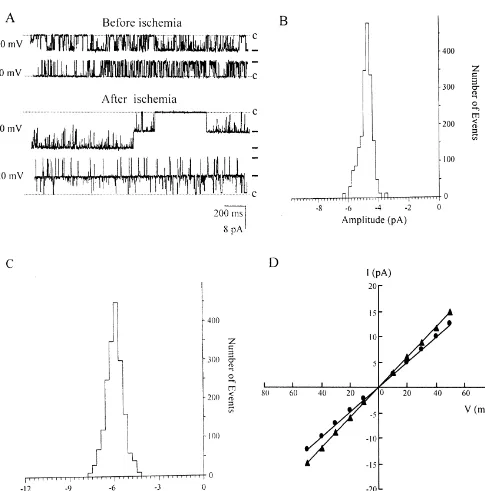
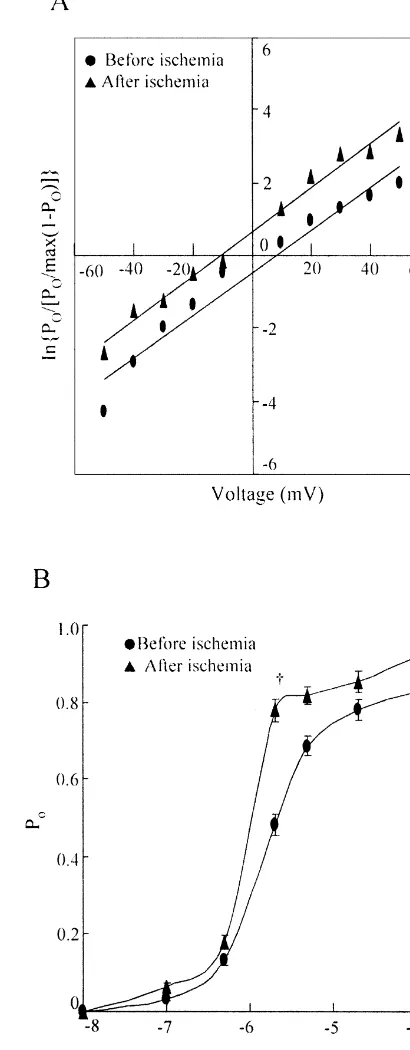
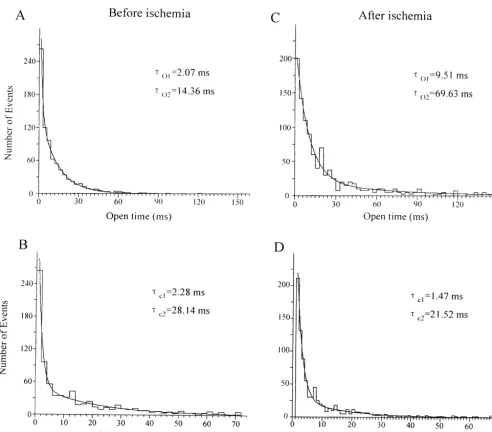
Dokumen terkait
The extended vase life of selected lines was related to low ethylene production at ¯ower senescence rather than to degree of ethylene sensitivity in young ¯owers.. Ethylene
Longer periods of chilling also resulted in increased uniformity in shoot emergence and decreased time to shoot emergence, suggesting that rhizomes may reach the physiological
ment, 83.3% (2.7) of the time intervals between The rotational grazing experiment showed that the cows entering the barn were shorter than 6 min, and sward height affects the
Following the short-lasting (1 h) hypoxia, the maximum level of preconditioning action was observed 7 days after hypoxia, whereas the longer-lasting hypoxia (8 h) produced the
Monitoring of fluorescence and conjugated dienes showed that peak values were reached in shorter times, and the peak values were lower for LDL aliquots oxidized more than 4 weeks
The abalone feeding experiment was conducted over an 8-week 56 days period from July to September, 1997, at HMSC. Dietary treatments consisted of dulse cultured under one of
In artificially inseminated herds, the proportion of cows returning for insemination after an interval longer than a normal oestrous cycle has been used as an estimate of
scopical observations. The ratio % of normal to atretic follicles decreased with time after the irradiation in primordial follicles and in primary follicles as well. At 6 h
