BAB II
KAJIAN PUSTAKA
2.1 Epidemiologi Kasus HIV di Dunia dan Indonesia
Infeksi HIV/AIDS merupakan masalah di seluruh dunia termasuk Indonesia. Menurut laporan UNAIDS (2010) walaupun terdapat penurunan angka insiden, prevalensi penderita yang hidup dengan HIV/AIDS tetap meningkat. Hal ini berhubungan dengan menurunnya jumlah kematian akibat AIDS sebagai dampak pengobatan antiretroviral (ARV) pada beberapa tahun terakhir.
Menurut UNAIDS, pada tahun 2009, diperkirakan 2,6 juta orang mendapatkan infeksi baru (newly infected) oleh HIV. Angka ini 19% lebih kecil dibandingkan tahun 1999 (3,1 juta orang) dan 21% lebih kecil dibandingkan tahun 1997 (3,2 juta orang) tahun dimana infeksi baru HIV mencapai puncaknya. UNAIDS juga memperkirakan terdapat 33,3 juta orang hidup dengan HIV/AIDS pada akhir tahun 2009, meningkat 27% dibandingkan tahun 1999 (26,2 juta orang).
Berdasarkan laporan Kemenkes RI (2011) terdapat total 26483 kasus AIDS di seluruh Indonesia secara kumulatif. Prevalensi kasus AIDS di Indonesia secara nasional adalah 11,09 per 100.000 penduduk. Sedangkan jumlah kasus baru HIV/AIDS secara nasional pada tahun 2010 adalah 4.158 kasus, lebih tinggi daripada tahun sebelumnya (3.863 kasus). Angka ini didominansi golongan usia produktif (20-40 tahun), serta heteroseksual sebagai faktor risiko.
2.2 Anemia Pada Penderita Terinfeksi HIV 2.2.1 Epidemiologi
Anemia adalah komplikasi yang sering ditemukan pada penderita terinfeksi HIV dan disebabkan oleh penyebab yang multifaktorial. Pada studi retrospektif terhadap 32.867 rekam medis penderita terinfeksi HIV di 9 kota di Amerika didapatkan distribusi dari kadar hemoglobin sangat bervariasi tergantung stadium dari penyakit. Pada penderita terinfeksi HIV tanpa gejala klinis AIDS, 72% pada pria dan 69% pada wanita memiliki kadar hemoglobin dalam batas normal (masing-masing lebih dari 14 atau 12 g/dL). Akan tetapi pada penderita dengan gejala klinis AIDS hanya 13% pada pria dan 23% pada wanita yang memiliki kadar hemoglobin dalam batas normal. Insiden anemia berhubungan dengan stadium penyakit dimana insiden dalam satu tahun adalah 3,2 % penderita terinfeksi HIV tanpa gejala klinis AIDS dan 36,9% pada penderita dengan gejala klinis AIDS. Kejadian anemia pada infeksi HIV lebih banyak terjadi pada ras kulit hitam serta jenis kelamin perempuan (Sullivan, 2008).
Penelitian Levine, dkk (2001) pada 2056 perempuan dengan HIV positif dan 569 perempuan dengan HIV negatif menunjukkan bahwa 37% dari perempuan dengan HIV positif menderita anemia sedangkan hanya 17% prevalensi anemia pada HIV negatif. Perempuan dengan HIV positif juga lebih cenderung mengalami anemia berat dibandingkan dengan HIV negatif. Anemia lebih sering terjadi pada ras kulit hitam daripada ras lainnya, pada perempuan terinfeksi HIV, 44,9% terjadi pada ras kulit hitam, sedangkan 25,7% pada ras kulit putih dan 24,8% pada ras latina.
Penelitian di Ghana pada 442 penderita dengan HIV/AIDS mendapatkan insiden anemia sebesar 63% pada penderita pra ARV, lebih besar secara signifikan dibandingkan penderita dengan terapi ARV yaitu 46% (Owiredu, 2011).
2.2.2 Etiologi
Penyebab anemia pada penderita HIV adalah multifaktorial, namun penurunan produksi eritrosit akibat eritropoesis yang inefektif merupakan faktor utama dibandingkan penyebab lain. HIV secara langsung mempengaruhi sel progenitor hematopoetik sumsum tulang atau secara tidak langsung melalui melalui efek sitokin yang menyebabkan penurunan produksi eritrosit. Interferon-γ (IFN-γ), tumour necrosis factor-α (TNF-α) dan beberapa sitokin lain dapat menghambat hematopoesis. Level sitokin tersebut ditemukan meningkat pada penderita terinfeksi HIV. Pengobatan terhadap infeksi HIV dan penurunan viral load menggunakan ARV dapat memperbaiki hematopoesis (Claster S, 2002).
Infeksi opportunistik juga dapat menyebabkan anemia pada penderita HIV. Infeksi tuberkulosis merupakan infeksi opportunistik tersering pada infeksi HIV dan diketahui menyebabkan anemia on chronic disease (ACD). Infeksi parvovirus B19 yang menyebabkan penurunan yang signifikan terhadap jumlah eritrosit. Penelitian Naides, dkk (1993) menunjukkan empat dari sembilan pasien dengan viremia B19 yang persisten setelah diperiksa secara serial menderita anemia berat (hemoglobin < 8,5 g/dL) bahkan setelah zidovudin dihentikan. Sedangkan 5 penderita dengan anemia berat yang mengalami perbaikan setelah zidovudin dihentikan tidak menunjukkan viremia B19. Penelitian pada tujuh penderita HIV dengan pure red cell aplasia menunjukkan terdapat giant pronormoblast pada sumsum tulang akibat infeksi akut dari parvovirus B19 (Frichofen, 1990). Micobacterium avium complex dapat mengenai beberapa organ. Organ yang paling sering terkena adalah darah,
sumsum tulang, hati, limpa, dan limfonodi. Anemia berat adalah salah satu tanda infeksi mikroorganisme ini (Desforges, 1991).
Anemia juga dapat disebabkan oleh efek samping dari pengobatan, walaupun banyak obat yang digunakan untuk terapi HIV/AIDS adalah bersifat myelosupresif, anemia berat lebih sering terjadi akibat penggunaan zidovudin. Cotrimoxazole menghambat metabolisme asam folat dan beberapa penelitian telah melaporkan adanya kelainan akibat penggunaannya. Kelainan ini potensial diperburuk jika penggunaannya dikombinasi dengan myelosupresif dan obat anti asam folat seperti zidovudin. Penelitian pada 982 penderita HIV dengan pemberian cotrimoxazole profilaksis menunjukkan peningkatan persentase penderita dengan kelainan hematologi antara lain: leukopenia (2,1%), neutropenia (2,0%), trombositopenia (2,3%), dan anemia (2,3%). Data ini telah distratifikasi dengan mean jumlah CD4+ (Watera, 2007). Penelitian oleh Keizu tahun 1990, dalam periode 10 tahun (1976-1985) terdapat total 154 kasus dengan diskrasia darah dilaporkan di Swedia dimana setelah dievaluasi berhubungan penggunaan cotrimoxazole. Kelainan tersebut berupa 61 kasus leukopenia, 28 kasus trombositopenia, dan 2 kasus anemia non hemolitik. Ganciclovir adalah analog nukleosid dengan dengan aktivitas antivirus secara invitro terhadap grup herpes virus dan beberapa virus DNA yang lain. Efek terhadap sistem hematologi sering terjadi pada pemberiannya, tetapi efek ini bersifat reversibel (Faulds dan Heel, 1990)
Perubahan pada hormon eritropoesis seperti penurunan hematopoetic growth factor dan eritropoetin dapat menyebabkan anemia. Produksi eritropoetin oleh ginjal dibutuhkan untuk menstimulasi prekursor eritroid sumsum tulang untuk
berproliferasi dan meningkatkan produksi eritrosit, jadi insufisiensi renal yang berat memberikan kontribusi terhadap terjadinya anemia pada penderita terinfeksi HIV. Produksi eritrosit normal juga membutuhkan fungsi sumsum tulang yang normal, jadi harus bebas dari infeksi dan tumor.
Walaupun relatif jarang, defesiensi vitamin B 12 merupakan penyebab anemia pada penderita HIV. Studi prospektif terhadap 60 penderita terinfeksi HIV yang dirawat di rumah sakit. Kadar serum vitamin B12 yang rendah didapatkan pada 10 penderita (16,7%). Pemberian terapi parenteral memberikan respon yang berbeda walaupun serum vitamin B12 telah terkoreksi (Remacha, 1991).
Walaupun penyebab utama terjadinya anemia pada penderita terinfeksi HIV disebabkan oleh penurunan produksi eritrosit, adalah penting untuk mempertimbangkan penyebab lain seperti hemolisis serta perdarahan gastrointestinal yang dapat terjadi pada penderita ini. Hasil yang positif pada direct Coombs tes dilaporkan terdapat pada 20% sampai 40% penderita HIV. Walaupun demikian autoimmune hemolytic anemia (AIHA) jarang terjadi pada penderita ini. Walaupun belum jelas, mekanisme patofisiologi penyakit ini berhubungan dengan produksi antibodi antieritrosit, hipergammaglobulinemia, atau terbentuknya kompleks imun. Hal ini mungkin disebabkan oleh defek secara umum pada regulasi produksi antibodi akibat infeksi HIV (Saif, 2001).
Anemia pada penderita terinfeksi HIV sering dihubungkan dengan apa yang dinamakan “anemia on chronic disease” karena sering penurunan produksi eritrosit terjadi sekunder akibat Infeksi HIV yang kronik. Hal ini disertai dengan menurunnya respon terhadap eritropoetin.
2.3 Dampak Anemia Pada Penderita Terinfeksi HIV 2.3.1 Dampak anemia terhadap kualitas hidup
Fatigue adalah adalah gejala utama tersering pada penderita HIV, hal ini menyebabkan gangguan fungsi fisik dan penurunan kualitas hidup. Perkiraan prevalensi fatigue pada penderita HIV yang asimtomatik berkisar antara 10% sampai 30%, sedangkan pada penderita dengan stadium lanjut mencapai 50% (Breibhart, dkk., 1998; Darko, dkk., 1992).
Saat anemia yang berhubungan dengan infeksi HIV dapat diatasi, secara umum terdapat peningkatan kualitas hidup penderita. Bukti ini ditunjukkan oleh penelitian Revicki, dkk (1994) pada studi multisenter untuk mengevaluasi efek terhadap pemberian recombinant human erythropoietin (epoetin alfa) pada kualitas hidup penderita HIV dengan anemia. Penderita dengan anemia yang terkoreksi dengan epoetin alfa menunjukkan perbaikan yang signifikan dalam energi/fatigue, home management, kesehatan yang memuaskan (health satisfaction), kesehatan global dibandingkan dengan penderita dengan anemia yang tidak terkoreksi.
Studi selanjutnya meneliti hubungan pemberian epoetin alfa terhadap kualitas hidup penderita HIV. Pada penderita ini tetap diberikan zidovudin dengan kombinasi dengan antiviral yang lain termasuk protease inhibitor. Analisis kualitas hidup pasien distratifikasi berdasarkan jumlah limfosit CD4+. Hasilnya, terapi epoetin alfa secara signifikan memperbaiki anemia dan menurunkan kebutuhan transfusi, independent terhadap jumlah CD4+. Lebih lanjut, peningkatan kadar hemoglobin secara signifikan memperbaiki kualitas hidup penderita terutama kesehatan fisik (Abrams, 2000).
2.3.2 Anemia dan survival
Beberapa penelitian menunjukkan bukti yang konsisten mengenai hubungan antara anemia dengan mortalitas pada penderita terinfeksi HIV. Hasil penelitian multisenter menunjukkan bahwa penderita terinfeksi HIV dengan anemia memiliki resiko yang lebih tinggi untuk mortalitas dibandingkan dengan penderita tanpa anemia, walaupun setelah dikontrol dengan beberapa faktor yang diketahui mempengaruhi survival seperti viral load atau jumlah limfosit T CD4+. The Center for Disease Control and Prevention (CDC) melakukan penelitian pada kohort retrospektif yang besar (large retrospective observational cohort study) dimana informasi dikumpulkan dari rekam medis dari 32.867 penderita HIV positif yang diobati di rumah sakit, klinik dan praktek swasta di seluruh Eropa. Hasilnya menunjukkan bahwa pada penderita anemia memiliki median survival yang lebih pendek secara signifikan dibandingkan dengan penderita tanpa anemia (Sullivan, 1998).
The EuroSIDA study group melakukan dan menganalisis data kohort retrospektif yang dikumpulkan di Eropa pada 6725 penderita dengan kadar hemoglobin normal, anemia ringan, dan berat. Kurva Kaplan Meier menunjukkan proporsi penderita yang meninggal dalam beberapa bulan sejak observasi (gambar 2.1). Pada penderita, 3,1% tanpa anemia, 15,9% dengan anemia ringan dan 40,8% dengan anemia berat, meninggal setelah 12 bulan. Perbedaan antara 3 kelompok berbeda secara signifikan (p<0,001) (Mockrof dkk., 1999).
Gambar 2.1 Kurva Kaplan-Meier waktu kematian pada penderita terinfeksi HIV dengan hemoglobin normal (Hb), anemia ringan, atau anemia berat dalam penelitian EuroSIDA
Sumber: Mockroft A, Kirk O, Barton SE, Dietrich M, Proenca R, Colebunders R, Pradier C, Monforte A, Ledergerber B, Lundgren JD, 1999. Anemia is an independent predictive marker for clinical prognosis in HIV-infected patients from across Europe. J Acquir Immune Defic Sindr 13: 943-50.
Penelitian yang lain menanalisis hubungan antara kadar hemoglobin dan survival pada pada 2343 penderita terinfeksi HIV. Anemia didefinisikan sebagai kadar hemoglobin <9,5 g/dL. Hasilnya menunjukkan penderita dengan anemia memiliki risiko kematian yang lebih besar secara signifikan dibandingkan dengan penderita tanpa anemia. Hubungan ini independen terhadap jumlah limfosit CD4+, adanya infeksi oportunistik, umur, serta penggunaan antiretroviral maupun terapi infeksi opportunistik (Moore, dkk., 1998).
Perbaikan anemia juga menunjukkan hubungan terhadap survival pada penderita HIV. Pada studi kohort yang dilakukan CDC terhadap 3203 penderita HIV dengan anemia menunjukkan perbaikan terhadap anemia secara signifikan menurunkan risiko terhadap kematian walaupun telah dikontrol dengan jumlah limfosit T CD4+. (Sulivan, dkk., 1998). Penelitian Moore dkk. (1998) juga menunjukkan koreksi anemia dengan pemberian epoetin alfa menyebabkan
penurunan yang signifikan terhadap risiko kematian. Hubungan ini tetap setelah dikontrol dengan faktor yang diketahui mempengaruhi prognosis.
2.4 Eritropoesis dan Hemoglobin
Sistem eritroid terdiri dari sel darah merah (red cell) atau eritrosit dan prekursor eritroid. Unit fungsional dari sistem eritroid ini dikenal sebagai eritron (erythron) yang mempunyai fungsi penting sebagai pembawa oksigen (oxygen carrier).
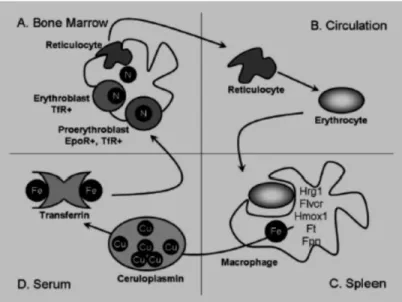
Prekursor eritroid dalam sumsum tulang berasal dari sel induk hematopoetik, melalui jalur sel induk myeloid, kemudian menjadi sel induk eritroid, yaitu BFU-E dan selanjutnya CFU-E. Prekursor eritroid yang dapat dikenal secara morfologik konvensional dalam sumsum tulang dikenal dengan pronormoblast, kemudian berkembang menjadi basophilic (early normoblast), selanjutnya polychromatophilic normoblast, dan acidophilic (late) normoblast. Sel ini kehilangan intinya, masih tertinggal sisa-sisa RNA, yang jika di cat dengan pengecatan khusus akan tampak seperti jala sehingga disebut retikulosit. Retikulosit akan dilepas ke darah tepi, kehilangan sisa RNA sehingga menjadi eritrosit dewasa. Proses ini dikenal sebagai eritropoesis. Proses pembentukan eritrosit memerlukan: sel induk (BFU-E, CFU-E, normoblast); bahan pembentuk eritrosit (besi, vitamin B12, asam folat, protein); mekanisme regulasi (faktor pertumbuhan hematopoetik dan hormon eritropoetin) (Greer, dkk., 2004).
Prekursor eritroid pada beberapa stadium perkembangan berinteraksi dengan makrofag sumsum tulang. Lingkungan mikro ini sering disebut dengan erythroid island (gambar 2.2). Proses perkembangan eritrosit tergantung dari adanya
eritropoetin (Epo-dependent) dimana secara langsung akan mempengaruhi proliferasi dan survival dari progenitor eritroid. Prekursor eritroid yang masih memiliki nukleus disebut eritroblas mengekspresikan transferin reseptor (TfR) dalam kadar yang tinggi, reseptor ini diperlukan untuk uptake besi dari transferin serum. Jika produksi hemoglobin telah mencukupi, TfR disimpan dan membentuk soluble-transferin reseptor (sTfr) (Roy, 2010).
Gambar 2.2. Gambaran proses eritropoesis normal
A. Prekursor eritroid matur dalam sumsum tulang dalam kontak dengan sentral makrofag; B. Setelah proses hemoglobinisasi lengkap, prekursur eritroid mengeluarkan nukleusnya dan masuk ke dalam sirkulasi; C. Senescent erythrocytes difagosit oleh makrofag jaringan, dimana besi mengalami recycled; D. Besi dikembalikan ke eritron via transferin.
Sumber: Roy CN, 2010. Anemia of inflammation. Blood 83: 276-80.
Eritrosit hidup dan beredar dalam darah tepi (life span) rata-rata selama 120 hari. Setelah 120 hari eritrosit mengalami penuaan (senescence) kemudian dikeluarkan dari sirkulasi oleh sistem RES.
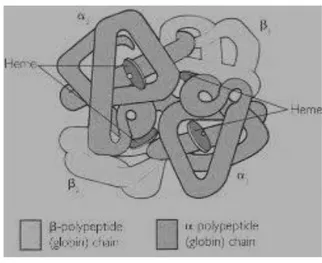
Eritrosit matang merupakan suatu cakram bikonkaf dengan diameter sekitar tujuh mikron. Eritrosit merupakan sel dengan struktur yang tidak lengkap. Sel ini hanya terdiri atas membran dan sitoplasma tanpa inti sel. Komponen eritrosit terdiri dari: membran eritrosit, sistem enzim, dan hemoglobin. Hemoglobin berfungsi sebagai alat angkut oksigen. Komponennya terdiri atas: heme, yang merupakan gabungan protoforfirin dengan besi; dan globin, bagian protein yang terdiri dari dua rantai alfa dan dua rantai beta (gambar 2.3).
Gambar 2.3 Struktur Hemoglobin.
Anemia merupakan kelainan hematologi yang paling sering dijumpai baik di klinik maupun di lapangan. Untuk mendapatkan pengertian anemia maka kita perlu menetapkan definisi anemia. Anemia adalah keadaan dimana massa eritrosit dan/atau massa hemoglobin yang beredar tidak dapat memenuhi fungsinya untuk menyediakan oksigen bagi jaringan tubuh. Secara laboratorik dijabarkan sebagai penurunan di bawah normal kadar hemoglobin, hitung eritrosit dan hematokrit (Beutler, dkk., 2001).
Untuk menjabarkan definisi anemia di atas maka perlu ditetapkan batas hemoglobin yang bisa kita anggap sudah tejadi anemia. Cutoff point yang umum dipakai adalah kriteria WHO (1989). Dinyatakan anemia bila: laki-laki dewasa hemoglobin < 13 gr/dl; perempuan dewasa tidak hamil emoglobin < 12 gr/dl.
2.5 Aktivasi Imun Pada Infeksi HIV
Untuk mengetahui patogenesis anemia pada penderita terinfeksi HIV, terlebih dahulu harus dipahami pengaruh infeksi HIV terhadap aktivasi sel limfosit dan monosit yang akan menyebabkan adanya disregulasi sitokin, kemudian bagaimana sitokin pro inflamasi mempengaruhi eritropoesis normal.
Penelitian menunjukkan bahwa limfosit T, monosit, dan makrofag pada infeksi HIV berada dalam keadaan teraktivasi yang ditandai peningkatan marker akivasi pada permukaan selnya serta peningkatan jumlah memory/primed/activated T cell dan penurunan naïve/resting T cell. Aktivasi sel limfosit T terjadi karena presentasi antigen pada T cell receptor (TCR) oleh antigen precenting cells (APC) serta protein HIV seperti gp120 pada reseptor CD4 sel T. Parameter dari aktivasi imun ini dapat memberikan nilai prognostik yang kuat dalam memprediksi progresifitas penyakit.
Hazenberg, dkk (2003) melakukan penelitian pada 149 pria homosexual penderita terinfeksi HIV yang dilakukan pada peripheral blood mononuclear cells (PBMC). Sampel diambil sebelum terjadinya serokonversi dan 1 serta 5 tahun setelah serokonversi. Level dari marker aktivasi sel dari sel limfosit T CD4+ dan CD8+ seperti CD38, HLA-DR, CD70, dan Ki67 diukur menggunakan flow cytomery
activated cell shorter scan (FACS scan). CD38 dan HLA-DR (MHC class II antigen) adalah dua antigen yang diketahui meningkat ekspresinya pada sel T yang telah teraktivasi. CD70 merupakan famili dari reseptor tumor necrosis factor (TNF) diekspresikan pada sel T yang teraktivasi dan mengontrol besar dan durasi dari respon sel T. Penderita terinfeksi HIV menunjukkan peningkatan ekspresi CD70 dimana mencerminkan menetapnya peningkatan level dari aktivasi imun. Ki67 adalah antigen inti sel dimana diekspresikan secara khusus oleh sel yang dalam keadaan membelah, karena itu sering digunakan sebagai marker untuk proliferasi sel T. Hasil penelitian menunjukkan bahwa setelah serokonversi terjadi peningkatan secara signifikan ekspresi Ki67, HLA-DR, CD38 dan CD70 pada sel T CD4+ dan CD8+ dibandingkan nilai pada preserokonversi. Pada penderita yang progresifitas penyakitnya lambat (slow progresor) ekspresi Ki67 stabil stiap waktu, akan tetapi pada yang progresifitas penyakitnya cepat (fast progresor) terdapat peningkatan secara signifikan ekspresi antigen ini pada sel T CD4+ dan CD8+. Korelasi negatif kuat didapatkan pada jumlah total sel CD4+ dengan dengan level ekspresi dari Ki67, HLA-DR, CD 38 (r: -0,506 - 0,599; p<0,005). Hasil penelitian menunjukkan bahwa terdapat peningkatan level aktivasi sistem imun yang persisten setelah fase akut dari infeksi HIV.
Seperti pada infeksi HIV-1, jumlah sel T CD4+ mengalami penurunan secara progresif pada infeksi HIV-2, akan tetapi penurunannya lebih lambat dan level viremia lebih rendah pada setiap stadium penyakit. Sousa, dkk (2002) melakukan penelitian potong lintang pada 27 penderita terinfeksi HIV-2, dan 26 penderita terinfeksi HIV-1, serta 25 individu sehat sebagai kontrol. Hasilnya menunjukkaan
bahwa untuk jumlah sel T CD4+ yang sama, penderita terinfeksi HIV-1 maupun HIV-2 menunjukkan level yang sama pada marker aktivasi dan proliferasi sel meskipun terdapat perbedaan yang besar pada plasma viral load. Hal ini ditandai dengan: 1. Imbalans rasio populasi sel T naïve : memory; 2. Peningkatan ekspresi marker aktivasi sel T CD4+ dan CD8+ (HLA-DR, CD38, CD69, Fas/CD95). Hasil penelitian ini menunjukkan hubungan yang kuat antara aktivasi imun dan penurunan jumlah sel T CD4+ pada infeksi HIV dan hubungan yang tidak langsung dengan viral load.
Penelitian lain bertujuan mengetahui hubungan antara penurunan jumlah sel T CD4+ dan aktivasi imun pada anak yang terinfeksi HIV-1 dengan terapi ARV. Penelitian potong lintang dilakukan pada 143 anak dengan terapi ARV lebih dari 24 minggu, sebagai kontrol juga diperiksa 23 anak sehat. Hasilnya, anak yang terinfeksi HIV dengan persentase CD4+ yang rendah memiliki sel T CD4+ memori (CD45RO+) yang tinggi dan sel CD4 naïve+ (CD45RA+) yang rendah. Lebih lanjut, anak dengan CD4 > 25% memiliki jumlah sel T CD4+ memori dan naive yang sama dengan grup kontrol. Pada anak dengan persentase CD4+ rendah menunjukkan peningkatan yang signifikan ekspresi HLA-DR, dan CD38 pada sel T CD4+ dan CD8+ independen terhadap level viral load. Penelitian ini mencerminkan bahwa pada penderita terinfeksi HIV dengan terapi ARV memiliki ekspresi marker aktivasi sel T yang tinggi pada jumlah sel T CD4+ yang rendah. Anak dengan infeksi HIV dengan persentase CD4+ yang terendah memiliki jumlah sel T CD4+ memori dan teraktivasi tertinggi dan juga jumlah naive sel T CD4+ dan CD8+ yang terendah. Jadi, penurunan
jumlah sel T CD4+ pada penderita berhubungan dengan level yang tinggi dari aktivasi sel T yang diinduksi oleh replikasi HIV yang kontiniu (Resino, 2006).
2.6 Aktivasi Imun dan Disregulasi Sitokin
Inflamasi telah diketahui sebagai penyebab disfungsi imun pada penderita terinfeksi HIV, mengindikasikan bahwa aktivasi imun adalah faktor penyebab lemahnya imunitas dan progresifitas penyakit pada penderita terinfeksi HIV. Penelitian menunjukkan replikasi virus sendiri merupakan penyebab utama keadaan chronic inflamatory state pada penderita HIV, dimana pada penderita menunjukkan peningkatan marker dari inflamasi dan aktivasi imun. Aktivasi imun dapat mempengaruhi proses eritropoesis melalui peningkatan produksi sitokin pro inflamasi seperti tumor necrosis factor-α (TNF-α) dan interferon-γ (IFN-γ), serta interleukin-10 (IL-10) oleh sel T yang telah teraktivasi.
Infeksi HIV berhubungan dengan perubahan pada imunitas seluler (cell mediated immunity) yang dapat diidentifikasi melalui kultur sel dari penderita terinfeksi secara invitro. Peripheral blood mononuclear cells (PBMC) yang diisolasi dari penderita terinfeksi HIV menunjukkan ketidakseimbangan produksi sitokin yang berhubungan dengan disfungsi imunitas seluler.
Penelitian Graziosi, dkk (1996) mempelajari kinetik ekspresi dari beberapa sitokin seperti IL-2, IL-4, IL-6, IL-10, IFN-γ, dan TNF-α pada PBMC sembilan penderita terinfeksi HIV menggunakan teknik semiquantitative reverse transcriptase-polymerise chain reaction( RT-PCR). Hasilnya menunjukkan ekspresi dari IL-2 dan IL-4 tidak atau minimal sekali terdeteksi pada PBMC. Akan tetapi kadar IL-2 yang
cukup tinggi ditemukan pada sel mononuklear yang diisolasi dari limfonodi (lymph node mononuclear cell). Ekspresi dari IL-6 terdeteksi hanya pada tiga dari sembilan pasien, dan ditemukan pada saat transisi dari fase akut ke fase kronik. Eksprei dari IL-10 dan TNF-α konsisten terdeteksi pada semua pasien, dan kadar semua sitokin ini bersifat stabil dan meningkat menurut waktu. Sama dengan IL-10 dan TNF-α, peningkatan ekspresi dari IFN-γ terdeteksi pada semua penderita. Lima dari sembilan penderita menunjukkan puncak level ekspresi IFN-γ yang sangat awal dari infeksi HIV. Ekspresi puncak dari IFN-γ bersamaan dengan ekspansi oligoklonal dari sel T CD8+ dan sel T CD8+ paling banyak mengekspresikan sitokin ini. Sebagai tambahan, ekspresi dari IL-6, IL-10, IFN-γ, dan TNF-α oleh sel mononuklear pada limfonodi dibandingkan dengan darah perifer lebih tinggi secara signifikan (tiga kali lipat untuk tiap sitokin). Dari hasil ini dapat disimpulkan bahwa sitokin tertentu seperti IL-2 dan IL-4 pada darah perifer mungkin tidak akurat merefleksikan situasi pada organ yang lain seperti limfonodi dan sumsum tulang. Hal ini mengindikasikan bahwa beberapa sitokin mungkin diproduksi dan bekerja lokal pada jaringan tanpa masuk kedalam sirkulasi. Level dari sitokin dalam serum adalah relatif rendah dan lebih mudah dipengaruhi pleh fluktuasi diurnal dari aktivitas imun. Lebih jauh lagi, sitokin berbeda dengan hormon, dimana sitokin merupakan soluble factor yang menunjukkan efek lokal pada lingkungan seluler tertentu. Jadi, secara in vitro stimulasi terhadap sel T memberikan gambaran yang lebih baik bagaimana sel T memberikan reaksi dalam bentuk produksi sitokin. T-cell mediated response sangat berhubungan dengan adanya puncak yang awal dari ekspresi IFN-γ, dan penelitian menunjukkan bahwa respon sitokin pada saat awal memainkan peranan utama dalam menekan replikasi
virus. Akhirnya, respon yang bifasik dari ekspresi sitokin (puncak yang awal dari IFN-γ dan puncak yang lambat dari IL-6, IL-10, dan TNF-α) secara jelas menggambarkan peranan sel T dalam antiviral immune respon.
Hasil penelitian yang sama juga ditunjukkan oleh Caruso, dkk (1995). Dengan teknik immunoflorescence dan flow cytometry didapatkan bahwa pada penderita HIV mulai dari stadium awal infeksi terdapat peningkatan persentase limfosit yang memproduksi IFN-γ dibandingkan dengan kontrol sehat. Peningkatan ekspresi IFN-γ oleh limfosit meningkat dengan meningkatnya stadium penyakit. Peningkatan ekspresi sitokin ini terjadi pada sel T CD4+ maupun CD8+.
Westby, dkk. (1998) menggunakan three colour- flowcytometry activated cell shorter (FACS) menganalisis produksi sitokin intraselular pada 20 penderita terinfeksi HIV dan 20 orang sehat sebagai kontrol. Hasilnya, sel limfosit T pada penderita HIV baik yang simtomatik maupun asimtomatik menunjukkan penurunan kapasitas untuk memproduksi IL-2 khususnya pada sel T CD4+ dan peningkatan produksi IFN-γ khususnya pada sel T CD8+ dibandingkan dengan kontrol. Produksi IL-2 oleh sel T CD4+ juga juga lebih rendah secara signifikan pada penderita HIV yang simtomatik dibandingkan dengan asimtomatik. Produksi IL-2 oleh sel T CD8+ juga lebih rendah pada penderita HIV simtomatik dibandingkan dengan asimtomatik walaupun secara statistik tidak signifikan. Di sisi lain tidak terdapat perbedaan produksi IFN-γ antara penderita HIV simtomatik dan asimtomatik. Jadi, aktivasi sel T yang ditandai dengan tingginya produksi IFN-γ tampak pada awal terjadinya infeksi HIV dan menetap seiring perjalanan penyakit. Sedangkan penurunan produksi IL-2 oleh sel T bersifat progresif . Perbedaan produksi IL-2 dan IFN-γ antara
kelompok sampel dan kontrol tidak secara sederhana dijelaskan oleh terbaliknya rasio jumlah limfosit CD4:CD8, karena telah distratifikasi dengan jumlah sel T CD4+ maupun CD8+ pada masing-masing kelompok. Akan tetapi hasil ini disebabkan oleh penurunan kemampuan sel T CD4+ untuk memproduksi IL-2 dan peningkatan kemampuan sel T CD8+ untuk memproduksi IFN-γ akibat aktivasi sel. Terdapat korelasi negatif antara produksi IL-2 oleh sel T dengan viral load (r = -0,832), korelasi ini terdapat pada sel T CD4+ dan CD8+ (masing r = -0,435 dan -0,622).
Penurunan ekspresi IL-2 dan peningkatan ekspresi IFN-γ oleh sel T juga didapatkan Fan, dkk (1993). Dengan teknik quantitative polymerise chain reaction, didapatkan ekspresi mRNA untuk IL-2 yang lebih rendah dan ekspresi mRNA IFN-γ yang lebih tinggi pada PBMC penderita HIV dibandingkan kontrol sehat. Penelitian dengan teknik immunofluorescence dan flow cytometric analysis juga menunjukkan peningkatan produksi IFN-γ pada penderita HIV asimtomatik dibandingkan kontrol sehat. (Caruso, dkk., 1996).
Penelitian Haissman, dkk. (2009) pada 229 penderita HIV, dan 54 kontrol bertujuan mengevaluasi peranan aktivasi imun dalam peningkatan produksi sitokin dalam patogenesis infeksi HIV serta efek pemberian ARV pada parameter ini. Konsentrasi IL-6, IL-8, IL-10, TNF-α, Interleukin-1 reseptor antagonis (IL-1ra), dan monosite chemotactic protein-1 (MCP-1) dari plasma penderita terinfeksi HIV dan kontrol distratifikasi berdasarkan jumlah sel T CD4+ dan diikuti selama 2 dan 4 bulan setelah pemberian ARV. Hasilnya, pada penderita terinfeksi HIV menunjukkan peningkatan kadar sitokin TNF-α, IL-6, IL-8, IL-1ra, dan MCP-1 dibandingkan kontrol. Progresifitas HIV juga memiliki efek terhadap peningkatan sitokin, dimana
terdapat kadar yang lebih tinggi secara signifikan dari TNF-α, IL-6, IL-8, IL-1ra, dan MCP-1 pada kelompok penderita dengan jumlah sel T CD4+ kurang dari 200 sel/mm3. Kadar semua sitokin menurun setelah 2 bulan pemberian ARV dan berlanjut setelah 4 bulan pengobatan. Pada analisis univariat, semua sitokin memiliki korelasi positif dengan viral load dan berkorelasi negatif dengan jumlah sel T CD4+. Inflamasi merupakan penyebab utama dari HIV-related immunodeficiency dimana replikasi virus sendiri merupakan faktor utama yang menyebabkan aktivasi imun dan peningkatan kadar sitokin walaupun terdapat bermacam koinfeksi. Penelitian ini juga menggambarkan bahwa ARV sangat efisien untuk menurunkan aktivasi imun dan kadar sitokin.
Tuberkulosis (TB) adalah infeksi opportunistik tersering pada penderita AIDS. Manifestasi klinis dari tuberkulosis pada penderita HIV biasanya lebih berat dengan kerusakan paru yang difus dan sering dengan manifestasi ekstrapulmoner yang berat. Oleh karena itu parameter imunologik mungkin berbeda pada penderita AIDS dengan TB dibandingkan dengan penyakit itu secara sendiri-sendiri. Dengan stimulasi mitogen pada PBMC, produksi IFN-γ dan TNF-α oleh sel T dianalisis dengan metode enzim-linked immunoabsorbent assay (ELISA). Sampel didapat melalui 33 penderita dan dibagi menjadi empat kelompok: sebelas penderita AIDS dengan tuberkulosis, enam penderita HIV asimtomatik, delapan pasien dengan tuberkulosis, dan delapan kontrol sehat. Hasilnya menunjukkan proporsi sel T CD4+ yang mengekspresikan TNF-α lebih tinggi pada grup penderita TB dibandingkan dengan penderita HIV asimtomatik, sedangkan kelompok HIV-TB menunjukkan nilai intermediet yaitu diantara kelompok penderita HIV asimtomatik dan kelompok TB.
Pada sel T CD8+ ekspresi IFN-γ dan TNF-α berbeda pada keempat kelompok. Persentase sel T CD8+ yang mengekspresikan IFN-γ lebih tinggi pada kelompok penderita TB dibandingkan dengan kelompok penderita HIV asimtomatik, sedangkan kelompok HIV-TB menunjukkan nilai intermediet namun lebih mendekati nilai kelompok HIV asimtomatik. Yang menarik, ekspresi TNF-α oleh sel T CD8+ pada kelompok HIV-TB menunjukkan nilai yang sama dengan kelompok HIV asimtomatik dan lebih rendah dengan kelompok TB. Respon sel T terhadap Micobacterium tuberculosis membutuhkan produksi IFN-γ. Mekanisme proteksi ini berhubungan dengan terbentuknya granuloma untuk melokalisir infeksi yang tergantung oleh terdapatnya IFN-γ. Pada penelitian ini tidak terdapat perbedaan yang signifikan ekspresi sitokin ini oleh sel T CD4+ pada keempat kelompok penderita sedangkan pada sel T CD8+ ekspresi IFN-γ lebih tinggi pada kelompok TB daripada kelompok TB-HIV. Hal ini menunjukkan bahwa pada penderita HIV terdapat hiporesponsif sel T dalam mengenali antigen Mycobacterium tuberculosis yang mengakibatkan gangguan produksi IFN-γ pada kelompok TB-HIV. Hiporesponsif sel T ini terjadi akibat aktivasi imun secara umum (Cunha, dkk., 2005).
2.7 Abnormalitas Eritropoesis Pada Infeksi HIV
Beberapa penelitian menunjukkan terdapat hambatan pertumbuhan dan diferensiasi sel progenitor hematopoetik secara in vitro. Infeksi langsung HIV pada sel progenitor CD34+ pernah dilaporkan (Steinberg, dkk., 1991; Stanly, dkk., 1992), akan tetapi Thomas, dkk menemukan hanya 2 dari 10 sel CD34+ penderita yang positif, dan jumlah kopi dari proviral DNA pada sampel ini adalah 2 sampai 5 per 250.000 sel.
Dari hasil ini dapat disimpulkan bahwa sel progenitor hematopoetik CD34+ tidak rentan terhadap infeksi HIV dan bukan merupakan reservoir virus pada sebagian besar penderita HIV asimtomatik (Thomas, dkk., 1995). Hal ini mengindikasikan bahwa HIV mempengaruhi mekanisme hematopoesis secara indirek.
HIV menghambat beberapa jalur hematopoesis tanpa infeksi langsung pada sel progenitor eritroid CD34+ tetapi dengan merubah lingkungan mikro (microenvironment) yang mendukung hematopoesis. Hematopoesis abnormal terjadi pada infeksi HIV sebagai akibat dari disregulasi sitokin yang akan mempengaruhi ekspresi beberapa gen yang berhubungan dengan hematopoesis (Koka dan Reddy, 2004).
2.7.1 Pengaruh inflamasi/aktivasi imun terhadap eritropoesis
Beberapa penelitian menyatakan bahwa eritropoesis (dan myelopoesis) menurun pada penderita HIV naïve dibandingkan dengan kontrol akibat disregulasi produksi sitokin dan growth factor dan peningkatan apoptosis sel progenitor sumsum tulang. Isgro, dkk (2005) meneliti efek terapi ARV terhadap produksi sitokin oleh sel limfosit dan sel stromal sumsum tulang pada penderita terinfeksi HIV sebelum dan sesudah terapi ARV. Dibandingkan dengan penderita sehat, terdapat penurunan produksi IL-2 dan peningkatan TNF-α oleh sel sumsum tulang, bersamaan dengan terdapatnya penurunan aktivitas clonogenic. Terapi ARV menunjukkan perbaikan aktivitas stem cell, perbaikan gambaran fungsi sel stromal, peningkatan produksi IL-2 pada sumsum tulang, penurunan ekspresi Fas antigen bersamaan dengan penurunan produksi
TNF-α. Fas antigen bertanggung jawab terhadap terjadinya apoptosis pada sel eritroid progenitor maupun sel CD4+ yang secara detail akan dijelaskan kemudian.
Penelitian Isgro, dkk (2000) menunjukkan bahwa perubahan morfologi dan tidak efektifnya fungsi sumsum tulang bertanggung jawab terhadap terjadinya sitopenia pada infeksi HIV. Penelitian mengenai efek terapi ARV pada jumlah colony forming units erythroid (CFU-E) menggunakan bone marrow cell cultures bertujuan untuk mengevaluasi mekanisme pengaruh imun rekonstitusi terhadap sumsum tulang. Pada semua penderita setelah terapi ARV, peningkatan pertumbuhan koloni adalah homogen pada semua tipe sel progenitor hematopoetik. Hasil ini berhubungan dengan data in vivo yang menunjukkan peningkatan jumlah bone marrow mononuclear cells (BMMCs) disertai dengan peningkatan jumlah sel T CD4+ darah perifer serta penurunan plasma HIV-RNA. Hasil ini mengindikasikan bahwa penurunan jumlah sel progenitor hematopoetik dan atau tidak efektifnya rangsangan terhadap pertumbuhan sel progenitor merupakan penyebab abnormalitas hematologi pada penderita HIV. Pengendalian replikasi virus oleh ARV menyebabkan perbaikan aktivitas sel progenitor yang disebabkan supresi terhadap beberapa sitokin yang menginhibisi hematopoesis normal.
Constantini dkk. (2009) meneliti kapasitas eritropoetik dari sel progenitor CD34+ yang telah dimurnikan (purified CD34 progenitors) melalui analisis longitudinal terhadap pertumbuhan burst forming unit erythroid (BFU-E) sebelum dan sesudah terapi ARV pada 12 penderita terinfeksi HIV dan 31 kontrol sehat. Hasilnya menunjukkan bahwa secara in vitro terdapat akumulasi dari jumlah BFU-E pada penderita HIV naïve dibandingkan dengan kontrol dan supresi terhadap replikasi
virus oleh ARV diikuti dengan penurunan akumulasi BFU-E. Peningkatan jumlah BFU-E pada penderita HIV dengan replikasi virus aktif disebabkan oleh: (1) peningkatan produksi pro-erythropoietic factor yang bekerja pada sumsum tulang untuk mempertahankan eritrosit normal, (2) peningkatan eritropoesis yang tidak efektif dengan akumulasi prekursor eritroid, dan (3) penyakit inflamasi kronik. Hasil tersebut di atas mengindikasikan bahwa pada penderita HIV naïve, imflamasi kronik atau aktivasi imun berhubungan dengan tidak efektifnya eritropoesis dan akumulasi dari pekursor eritroid BFU-E. Interferon-γ dan TNF-α menghambat pembentukan perkembangan sel eritroid khususnya pada pertumbuhan diferensiasi BFU-E menjadi CFU-E sehingga terjadi akumulasi dari BFU-E. ARV menyebabkan supresi terhadap replikasi HIV, menurunkan aktivasi imun dan menormalkan jumlah BFU-E.
Penelitian Constantini mendapatkan bahwa baseline soluble transferin receptor (sTfR) pada infeksi HIV lebih rendah dibandingkan dengan kontrol dan meningkat dengan pemberian ARV. Sitokin pro inflamasi khususnya IFN-γ menurunkan kadar serum sTfR melalui mekanisme transkripsional dan postranskripsional (Weiss, dkk., 2002). sTfR diproduksi ketika prekursor eritroid telah cukup memproduksi hemoglobin dan kemudian menyimpan reseptornya. Saat persediaan besi untuk eritron menurun, atau kapasitas eritroid meningkat, sTfR dalam plasma meningkat. Walaupun terjadi restriksi besi pada anemia akibat inflamasi, sTfR tidak meningkat tetapi tetap dalam rentang normal akibat down regulation oleh sitokin pro inflamasi (Roy, 2010). Hal ini memperkuat indikasi bahwa inflamasi kronik atau aktivasi imun memang berperan terhadap penurunan eritropoesis.
Eritropoetin secara langsung akan mempengaruhi proliferasi dan survival dari eritroid progenitor. Penelitian Constantini juga mendapatkan bahwa kadar eritropoetin serum lebih mendekati kadar normal rendah dan meningkat setelah pemberian ARV. Hal ini menunjukkan gangguan terhadap produksi eritropoetin pada penderita HIV naïve walaupun tidak terdapat bukti adanya gagal ginjal berdasarkan kadar serum kreatinin. Hasil ini sesuai dengan penelitian sesuai dengan penelitian Wang, dkk (1993) dimana HIV dapat menginduksi penurunan produksi eritropoetin secara in vitro. Kadar eritropoetin serum relatif lebih rendah pada penderita HIV dan ini mungkin memberikan kontribusi terhadap terjadinya diseritropoesis dimana kadar eritropoetin yang normal atau normal rendah tidak cukup untuk melawan efek inhibisi dari sitokin inflamasi sehingga diperlukan kadar eritropoetin yang tinggi untuk mempertahankan eritropoesis normal (Levine, 2003).
Hasil yang berbeda didapatkan Salome dkk (2002) mengenai kadar serum eritropoetin pada 111 penderita HIV dimana didapatkan pada kebanyakan penderita menunjukkan kadar eritropoetin serum yang normal atau sedikit mengalami peningkatan serta korelasi terbalik yang signifikan antara kadar eritropoetin dan hemoglobin (r = - 0,6353). Penelitian Rarick dkk (1991) pada 110 penderita HIV simtomatik juga menunjukkan hubungan terbalik yang signifikan antara kadar eritropoetin serum dan kadar hemoglobin (r = - 0,42). Data ini menunjukkan bahwa terdapat penurunan kemampuan eritroid progenitor untuk berespon terhadap eritropoetin sehingga kompensasi sumsum tulang terhadap terjadinya anemia menjadi tidak adekuat. Menurunnya respon eritroid progenitor terhadap eritropoetin terjadi pada keadaan chronic inflammatory states dimana terjadi efek inhibisi oleh sitokin
pro inflamasi terhadap proses eritropoesis, kadar eritropoetin yang lebih tinggi diperlukan untuk melawan efek tersebut.
Adanya inflamasi kronik pada infeksi HIV juga menyebabkan gangguan hemoostasis besi. Di Indonesia, Wisaksana, dkk (2011) melakukan penelitian potong lintang terhadap beberapa parameter metebolisme besi pada penderita HIV di RS Hasan Sadikin Bandung, Jawa Barat dari bulan September 2007 sampai agustus 2009. Hasilnya menunjukkan bahwa anemia pada penderita HIV naïve sebagian normokromik-normositik dengan retikulosit indek yang normal atau rendah, kadar sTfR yang rendah, serta peningkatan kadar high sensitive C-reactive protein (hsCRP). Mengindikasikan bahwa anemia on chronic disease (ACD) merupakan penyebab utama mekanisme anemia pada infeksi HIV. Kadar ferritin yang lebih tinggi secara signifikan ditemukan khususnya pada penderita dengan jumlah limfosit CD4+ < 200 sel/mm3. Kadar feritin serum juga lebih tinggi juga ditemukan pada penderita dengan anemia sedang-berat dibandingkan dengan penderita dengan anemia ringan (p = 0,01) atau tanpa anemia (p = 0,07). Terdapat korelasi negatif yang signifikan antara kadar feritin dengan jumlah limfosit CD4+ (r = -0,48). Kadar ferritin yang tinggi dan sTfR yang rendah mengindikasikan bahwa redistribusi besi (iron redistribution) dan aktifitas eritropoetik yang rendah lebih berkontribusi dalam menimbulkan anemia dibandingkan defesiensi besi pada penderita terinfeksi HIV.
Akibat dari inflamasi, besi dialihkan dari sirkulasi ke system retikuloendotelial atau tempat penyimpanan lain dalam bentuk feritin. Hepcidin (HepC) memainkan peranan dalam proses ini (Nemeth, 2009). HepC adalah hormon peptida yang diproduksi oleh hepatosit. Ferroportin (Fpn) adalah suatu transporter
untuk memfasilitasi transport besi dari makrofag (gambar 2.2). Fpn berada pada enterosit duodenal dan makrofag jaringan. HepC adalah regulator yang poten dengan menginduksi internalisasi dan degradasi dari Fpn. Jadi, HepC berikatan dengan Fpn pada permukaan sel dan mengakibatkan restriksi absopsi besi melalui enterosit serta restriksi pelepasan besi oleh makrofag (Roy, 2010; Weiss, dkk., 2005). IFN-γ juga berperan dalam menyebabkan retensi besi oleh makrofag dengan menurunkan ekspresi Fpn yang akhirnya menghambat pelepasan besi oleh makrofag (Ludwiczek, dkk., 2003).
2.7.2 Apoptosis sel progenitor eritroid, peranan Interferon-γ
Beberapa penelitian seperti yang dijelaskan sebelumnya menunjukkan peningkatan IFN-γ pada infeksi HIV. Interferon-γ adalah sitokin multifungsional yang disekresi oleh sel limfosit T yang teraktivasi dan natural killer cell. Sitokin ini sangat poten dalam menginduksi aktivasi dan diferensiasi sel fagosit dan limfosit, juga sebagai antiviral, antiproliferatif, dan imunomodulator pada beberapa jenis sel. IFN-γ juga menunjukkan efek inhibisi terhadap perkembangan ganulocyte-macrophage colony forming unit (CFU-GM), burst forming units-erythroid (BFU-E) dan colony forming units-erythroid (CFU-E) secara in vitro. IFN-γ menghambat pembentukan koloni sel eritroid progenitor, hal ini menunjukkan bahwa IFN-γ memainkan peranan penting dalam menimbulkan anemia pada penyakit kronik.
Interferon-γ merupakan inhibitor eritropoesis yang paling poten, peningkatan konsentrasi IFN-γ mengindikasikan aktivasi dari cell-mediated immunity yang persisten. Terdapat korelasi negatif yang signifikan antara penurunan kadar
hemoglobin dengan peningkatan ekspresi IFN-γ sebagai marker aktivasi imun seluler. Hal ini menunjukkan bahwa inhibisi eritropoesis oleh IFN-γ memiliki peran penting sebagai penyebab anemia pada penderita terinfeksi HIV (Fuchs, dkk., 1993; Denz, dkk., 1990).
Interferon-γ diketahui menghambat proliferasi dan diferensiasi sel eritroid progenitor dan menyebabkan apoptosis sel eritroid, sebaliknya eritropoetin berperan meningkatkan produksi sel eritroid dan dan mencegah apoptosis. Efek yang berlawanan dari eritropoetin dan IFN-γ pada keadaan inflamasi kronis dibuktikan oleh beberapa penelitian. Means dan Krantz (1991) meneliti pengaruh recombinant human Interferon-γ (rhγIFN) terhadap pembentukan koloni secara invitro pada higly purified erythroid colony forming units (CFU-E) yang didapat melalui sumsum tulang. Hasil penelitian menunjukkan pertumbuhan koloni CFU-E secara signifikan dihambat oleh rhγIFN pada semua konsentrasi yang di tes. Inhibisi oleh rhγIFN dapat diatasi oleh peningkatan konsentrasi recombinant human erythropoietin (rhEpo). Penelitian ini menunjukkan bahwa inhibisi pertumbuhan CFU-E oleh rhγIFN dan beberapa sitokin lain dapat dikoreksi dengan paparan (exposure) rhEpo dosis tinggi. Peningkatan dosis rhEpo bukan bukan secara langsung menstimulasi pertumbuhan koloni akan tetapi rhEpo mengatasi efek inhibisi yang diinduksi oleh rhγIFN. Sebab pemberian rhEpo pada dosis yang lebih tinggi lagi tidak secara signifikan menyebabkan peningkatan pertumbuhan koloni. Hal tersebut dapat disebabkan karena rhγIFN menurunkan ekspresi reseptor Epo pada CFU-E dan membutuhkan konsentrasi reEpo yang tinggi untuk pertumbuhan koloni yang optimal.
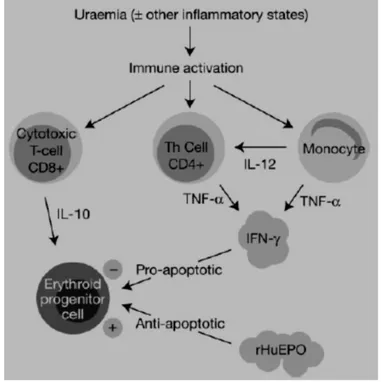
Gambar 2.4 Peningkatan aktivitas imun pada kondisi inflamasi mengakibatkan apoptosis sel eritroid progenitor
Keterangan: IFN, interferon; TNF, tumor necrosis factor; IL, interleukin; Th, sel T-helper
Sumber: Macdougall IC, Cooper AC, 2002. Erythropoietin resistance: the role of inflammation and pro-inflammatory cytokines. Nephrol Dial Transplant 17: 39-43
Interferon-γ adalah mediator utama supresi terhadap sel progenitor eritroid karena sitokin ini merupakan antagonis efek anti-apoptotik eritropoetin pada CFU-E. Karena IFN-γ hanya diproduksi oleh sel T yang teraktivasi, maka aktivasi sel T adalah sangat penting dalam memediasi supresi terhadap proses eritropoesis. Beberapa penelitian menunjukkan bahwa suatu keadaan chronic inflammatory state menunjukkan resistensi terhadap pemberian rhEpo yang anti-apoptotik oleh karena pengaruh sitokin pro inflamasi yang pro-apoptotik. Macdougall, dkk (2002) serta Cooper, dkk (2002) meneliti peningkatin level aktivasi imun melalui studi terhadap sel T pada penderita yang menjalani hemodialisis yang dibagi menjadi kelompok
berespon baik (good responders) dan berespon buruk (poor responders) terhadap eritropoetin. Hasilnya menunjukkan bahwa secara signifikan ekspresi TNF-α, IFN-γ, dan IL-10 pada sel T CD4+ dan CD8+ lebih besar secara signifikan pada penderita yang berespon buruk dibandingkan yang berespon baik terhadap rhEpo. Hasil ini mendukung konsep bahwa penderita yang berespon buruk terhadap terapi rhEpo disebabkan oleh peningkatan kapasitas sel T CD4+ dan CD8+ untuk memproduksi sitokin pro inflamasi terutama IFN-γ (gambar 2.4).
Penderita dengan kondisi inflamasi kronik seperti infeksi HIV dapat meningkatkan aktivasi imun yang melibatkan sel T dan monosit (gambar 2.3). Sel T dan monosit mensekresikan beberapa sitokin pro-inflamasi dimana salah satunya dapat meningkatkan produksi IFN-γ. Monosit saat teraktivasi memproduksi IL-12 dalam jumlah yang tinggi, dimana diketahui dapat menstimulasi sel T-helper untuk memproduksi IFN-γ (Gately, dkk., 1998) . Terdapat juga peningkatan produksi TNF-α oleh monosit dimana dapat memacu aksi dari IFN-γ sebagai agen pro-apoptotik baik melalui mekanisme addiktif maupun sinergistik (Sitter dan Bergner, 2000). IFN-γ memicu kematian sel eritroid progenitor pada sumsum tulang dengan cara mengantagonis aksi anti-apoptotik dari eritropoetin.
2.7.3 Mekanisme IFN-γ menyebabkan apoptosis sel progenitor eritroid
Beberapa penelitian telah menunjukkan efek inhibis IFN-γ terhadap sel progenitor eritroid. IFN-γ menghambat pertumbuhan dan diferensiasi CFU-E pada sumsum tulang dan menginduksi apoptosis. Proses apoptosis melibatkan banyak perubahan metabolik yang akan menimbulkan degradasi genomik DNA menjadi fragmen
nukleosomal. Mekanisme bagaimana IFN-γ menimbulkan efek inhibisi terhadap pertumbuhan sel progenitor eritroid dan menginduksi apoptosis adalah multifaktorial. Beberapa mekanisme yang diketahui antara lain:
1. Interferon-γ menginduksi ekspresi Fas reseptor pada sel progenitor eritroid sumsum tulang
Fas antigen adalah suatu molekul reseptor permukaan sel, dimana aktivasi oleh ligand-nya dapat menimbulkan tranduksi sinyal apoptosis. Beberapa penelitian menunjukkan bahwa sistem Fas/Fas ligand (FasL) memainkan peranan penting dalam menimbulkan apoptosis pada beberapa sel yang berbeda akibat interaksi antara fas dengan FasL. Fas (APO-1/CD95) adalah membran glikoprotein berukuran 45 kD dan merupakan famili tumour necrosis factor-receptor. Fas mengandung 70 asam amino yang dibutuhkan untuk menimbulkan sinyal transduksi apoptosis. Fas mRNA diekspresikan oleh sel limfosit, monosit, dan juga sel leukemia. Sedangkan FasL adalah suatu protein famili dari TNF berukuran 40 kD. FasL diproduksi oleh sel T yang teraktivasi terutama sel T sitotoksik (Takahashi, dkk., 1994). Antigen presenting cells adalah sumber dari FasL, dan infeksi HIV menyebabkan peningkatan ekspresi FasL oleh makrofag. Hal ini mengindikasikan bahwa makrofag juga berperan dalam apoptosis sel limfosit dan eritroid progenitor pada infeksi HIV (Baddley, dkk., 1996).
Dai, dkk (1998) meneliti ekspresi Fas pada CFU-E sumsum tulang manusia setelah diinkubasi dengan IFN-γ dan interaksi Fas dan FasL dalam menimbulkan kematian sel akibat apoptosis yang diinduksi oleh IFN-γ (IFN-γ induced apoptotic cell death). Untuk menentukan bahwa ekspresi Fas/FasL berhubungan dengan IFN-γ
dalam menginduksi apoptosis, ditentukan ada atau tidaknya molekul ini pada permukaan CFU-E dalam keadaan terdapat atau tidaknya IFN-γ. Analisis menggunakan northern and flow cytometric menunjukkan persentase yang kecil dari CFU-E manusia normal yang mengekspresikan Fas pada permukaannya dan juga levelnya sangat rendah. Ekspresi Fas pada permukaan mengalami peningkatan setelah enam jam inkubasi CFU-E dengan IFN-γ pada konsentrasi kurang dari 50 U/mL dan meningkat secara gradual dan mencapai puncak dalam 72 jam setelah inkubasi dengan IFN-γ dimana efek inhibisi IFN-γ terhadap pertumbuhan CFU-E jelas terlihat. Jadi, efek inhibisi IFN-γ terhadap CFU-E bersifat dose and time dependent. Eksperimen selanjutnya menunjukkan aktivasi Fas oleh anti-Fas monoclonal antibody (anti Fas MoAb) yang secara fungsional mirip dengan FasL menyebabkan penurunan viabilitas CFU-E. Apoptosis yang diinduksi IFN-γ sangat menurun setelah pemberian antihuman FasL antibody yang menghambat interaksi Fas dengan FasL. Penelitian ini menunjukkan bahwa IFN-γ meningkatkan ekspresi Fas pada permukaan sel progenitor eritroid serta menginisiasi interaksi antara Fas dengan FasL yang akan menimbulkan efek inhibisi dan apoptosis sehingga memberikan kontribusi yang besar dalam menghambat eritropoesis normal.
Penelitian lain juga membuktikan efek inhibisi dari TNF-α dan IFN-γ pada hematopoesis yang dimediasi atau dipotensiasi oleh sistem Fas/FasL. Terdapat peningkatan ekspresi Fas pada permukaan sel CD34+ setelah diinkubasi dengan TNF-α atau IFN-γ atau keduanya secara bersama-sama. Ekspresi Fas paling kuat diinduksi oleh adanya IFN-γ dan TNF-α secara bersamaan. Pemberian anti-Fas MoAb sangat memperkuat efek inhibisi dari TNF-α dan IFN-γ pada pertumbuhan koloni sel CD34+
bahkan pada konsentrasi TNF-α dan IFN-γ yang sangat rendah. Untuk menentukan apakah anti-Fas Mo-Ab memediasi terjadinya apoptosis dilakukan pemeriksaan terhadap DNA, dimana terdapat gambaran degradasi nukleosom yang merupakan tipikal terjadinya apoptosis. Hasil penelitian ini menunjukkan bahwa efek dari FasL adalah sinergistik dengan TNF-γ dan TNF-α dalam mempengaruhi proliferasi dan viabilitas sel. Jadi ekspresi Fas reseptor yang diinduksi oleh TNF-α dan IFN-γ akan menyebkan sel rentan terhadap FasL dimana interaksi Fas-FasL akan mentransduksi sinyal apoptosis (Maciejewski, dkk., 1995).
Kerjasama antara beberapa sitokin inhibisi seperti TNF-α, IFN-γ, dan FasL akan menimbulkan efek inhibisi yang poten pada sel sumsum tulang pada setting yang dikenal dengan immune-system attack. Ekspresi Fas juga terdapat pada sel limfosit penderita terinfeksi HIV yang akan dijelaskan pada sub bab selanjutnya. 2. IFN-γ menurunkan reseptor eritropoetin dan stem cell factor pada sel eritroid
progenitor
Eritropoetin dan stem cell factor (SCF) menunjukkan kerja yang bersifat sinergis untuk merangsang proliferasi dan/atau diferensiasi sel eritroid progenitor dan mencegah apoptosis in vitro (Muta dan Krantz, 1993; Muta, dkk., 1994). SCF dan eritropoetin juga dapat mengatasi efek inhibisi dari IFN-γ pada sel eritroid progenitor in vitro (Dai, dkk., 1995; Means, dkk., 1991) . Jadi, IFN-γ dan growth factor ini memiliki efek yang berlawanan terhadap sel progenitor eritroid.
Penelitian Taniguchi, dkk. (1997) menunjukkan bahwa IFN-γ menurunkan ekspresi reseptor eritropoetin dan SCF pada permukaan sel CFU-E dengan menurunkan ekspresi mRNA untuk kedua reseptor ini. IFN-γ menekan ekspresi
mRNA pada tingkat transkripsi. Hasil ini mengindikasikan bahwa IFN-γ menghambat proliferasi dan diferensiasi sel progenitor eritroid dan menurunkan survival dengan menurunkan jumlah target reseptor untuk eritropoetin dan SCF. Hasil penelitian ini dapat menjelaskan bagaimana dosis yang tinggi dari eritropoetin ini dapat mengatasi efek inhibisi dari IFN-γ, karena dengan sedikit reseptor yang terdapat pada permukaan sel, membutuhkan konsentrasi yang tinggi dari eritropoetin untuk mengembalikan probabilitas interaksi antara reseptor dengan ligand-nya.
3. IFN-γ menginduksi peningkatan ekspresi dan aktivasi caspase 1, 3 dan 8
Terdapat 10 enzym caspase yang telah teridentifikasi. Caspase merupakan protease yang memainkan peranan penting dalam perubahan biokimia yang menyebabkan kematian sel. Konversi caspase dari bentuk proenzym yang bersifat dorman ke bentuk enzym aktif adalah langkah penting dalam menimbulkan kaskade apoptosis yaitu penyusutan sel (cell shrinkage), kondensasi kromatin, pecahnya DNA, pembengkakan membran dan pembentukan apoptotic bodies (Cohen, 1997).
Dai dan Krantz (1999) meneliti efek IFN-γ terhadap ekspresi dan aktivasi beberapa caspase pada sel progenitor eritroid (CFU-E) menggunakan RNase protection assay dan western blot analysis. Hasilnya menunjukkan terdapat peningkatan ekspresi caspase 1,3 dan 8. Caspase ini tidak hanya meningkat ekspresinya, tetapi juga teraktivasi ketika sel mengalami apoptosis. Caspase inhibitor sangat efisien memblokade aktivasi kaspase dan mengurangi inhibisi pertumbuhan sel oleh IFN-γ.
Penelitian Cohen, dkk. (1997) menunjukkan bahwa Fas menginduksi apoptosis dengan mengaktivasi caspase 8 yang mengandung Fas-associating protein with death domain (FAAD) yang berfungsi sebagai death effector. Jadi terdapat hubungan langsung antara reseptor untuk kematian sel (cell death teceptors) dan caspase.
2.8 Mekanisme Patogenesis Penurunan Jumlah Sel Limfosit T CD4+
Keadaan imunodefisiensi pada penderita HIV ditandai oleh penurunan baik jumlah maupun fungsi dari sel limfosit T CD4+. Penyebab penurunan jumlah sel T CD4+ pada infeksi HIV antara lain adalah mekanisme direk dan indirek. Efek langsung HIV terhadap penurunan jumlah sel T CD4+ (direct HIV-mediated cytophatic effects) pernah dilaporkan pada studi in vitro namun tidak secara in vivo (Pantaleo, dkk., 1993). Namun penelitian menunjukkan bahwa derajat penurunan sel limfosit CD4+ tidak berkorelasi dengan derajat sitopatisitas HIV secara in vitro. Infeksi oleh strain HIV yang noncytophatic menyebabkan deplesi sel limfosit CD4+ yang ekstensif, sedangkan strain virus yang highly cythopatic menyebabkan deplesi sel limfosit CD4+ yang minimal walaupun dengan beban virus (viral burden) yang sama (Mosier, 1993). Penelitian dengan “melabel” sel limfosit pada limfonodi menunjukkan bahwa sel limfosit CD4+ yang mati akibat apoptosis bukanlah sel yang terinfeksi HIV melainkan bystander cells atau sel disekitarnya yang tidak terinfeksi (Finkel, dkk., 1995). Fakta lain menyebutkan bahwa sangat sedikit PBMC yang aktif terinfeksi HIV sedangkan terjadi penurunan yang cepat dan agresif dari sel limfosit CD4+ di sirkulasi (Wood, dkk., 1993). Jumlah sel yang terinfeksi HIV secara aktif adalah
sangat rendah, dengan rasio sel terinfeksi dengan tidak terinfeksi adalah 1:105 dengan kata lain sekitar 107 sel yang terinfeksi dari 1012 sel limfosit yang terdapat dalam tubuh (Chun, dkk., 1997). Bukti tersebut di atas secara jelas mengindikasikan bahwa mekanisme indirek dari infeksi HIV lebih berperan dalam penurunan jumlah sel limfosit CD4+ daripada efek langsung HIV terhadap sel.
Beberapa penelitian in vitro telah dapat mengidentifikasi mekanisme indirek dari infeksi HIV, dan HIV-induced apoptosis adalah mekanisme yang paling bisa menjelaskan penurunan jumlah sel limfosit CD4+. HIV-induced apoptosis telah dipublikasikan oleh beberapa penelitian. PBMC dari penderita HIV mengalami apoptosis secara in vitro (Meyaard, dkk., 1992; Oyaizu, dkk., 1993) dan kematian sel akibat apoptosis juga terjadi pada limfonodi. Derajat apoptosis sel pada limfonodi pada penderita terinfeksi HIV 3-4 kali lebih besar daripada yang diamati pada individu sehat. Apoptosis ini terjadi secara ekstensif pada semua kompartemen fungsional dari limfonodi seperti kortex, parakortex, dan sinus, berbeda pada individu dengan HIV negatif yang hanya terjadi pada germinal centers (Muro-Cacho, dkk., 1995).
2.8.1 Aktivasi imun dan apoptosis sel limfosit T CD4+ (activation-induced cell death/apoptosis)
Beberapa penelitian seperti yang dijelaskan pada bab sebelumnya menunjukkan bahwa pada infeksi HIV, adanya replikasi virus menyebabkan aktivasi imun yang ditandai dengan peningkatan marker aktivasi seperti HLA-DR, CD38, CD69, CD70, dan juga Fas/CD95 pada permukaan sel T CD4+. Peningkatan marker aktivasi ini
berhubungan dengan penurunan jumlah limfosit T CD4+ independent terhadap viral load. Penelitian tersebut di atas menunjukkan bahwa hubungan langsung (direct causal relationship) antara aktivasi imun dan penurunan jumlah limfosit T CD4+ dan hubungan yang tidak langsung antara penurunan jumlah sel limfosit T CD4+ dengan virus replication rate karena status imunologik berhubungan langsung dengan aktivasi imun dan bukan viral load (Resino, dkk., 2006; Hazenberg, dkk., 2003; Sousa, dkk., 2002). Aktivasi imun juga terjadi pada awal infeksi, sebelum terjadi penurunan yang signifikan dari jumlah sel T CD4+ (Sousa, dkk., 2002).
Penelitian Muro-Cacho, dkk. (1995) pada limfonodi penderita terinfeksi HIV juga menunjukkan bahwa terdapat korelasi yang signifikan antara intensitas terjadinya apoptosis dengan derajat aktivasi imun, sedangkan tidak terdapat korelasi antara intensitas apoptosis dengan viral burden pada limfonodi. Hasil ini mengindikasikan bahwa peningkatan intensitas apoptosis pada infeksi HIV disebabkan oleh akivasi imun secara umum (general state of immune activation), dan independent terhadap viral load.
Beberapa penelitian membuktikan bahwa aktivasi limfosit T CD4+ oleh antigen tertentu dapat menimbulkan apoptosis. Banda, dkk (1992) membuktikan bahwa ikatan silang (cross linked) envelove glycoprotein dari HIV, gp120 dengan anti-gp120 pada pada reseptor CD4 limfosit diikuti oleh ikatan antigen yang terikat MHC class II (MHC class II restricted antigen) pada T Cell Receptor (TCR) menyebabkan aktivasi sel T CD4+ yang menyebabkan terjadinya apoptosis atau programmed cell death. Data ini menunjukkan bahwa hanya dengan konsentrasi pikomolar dari gp120 menyebabkan activation-induced cell death. Kematian sel ini
memberikan gambaran yang karakteristik untuk apoptosis, seperti perubahan histologik dari inti sel dan kondensasi sitoplasma serta fragmentasi DNA. Fakta ini didukung oleh penelitian Groux, dkk (1992) yang menunjukkan bahwa rangsangan pada TCR oleh MHC class II dependent antigen menyebabkan kematian sel T CD4+ yang aktif dengan gambaran biokimia dan ultrastruktural dari apoptosis. Activation-induced cell death terjadi terutama pada sel limfosit T CD4+ dan kematian sel ini tidak tampak pada kontrol yang seronegatif HIV walaupun dengan infeksi akut maupun kronis. Terjadinya Activation induced cell death dapat dicegah dengan pemberian cyclosporin A.
Organ limfoid adalah reservoir utama HIV, dan monosit serta makrofag adalah target utama infeksi HIV. Sel limfosit T CD4+ baik yang terinfeksi maupun tidak terinfeksi HIV akan bermigrasi menuju limfonodi saat timbulnya respon imun. Pada keadaan ini sel limfosit CD4+ yang tidak terinfeksi akan kontak dengan APC yang terinfeksi HIV menyebabkan aktivasi dari sel T CD4+. Setelah teraktivasi, sel T CD4+ mengalami resirkulasi ke perifer dan mengalami apoptosis. Jadi APC seperti monosit dan makrofag memainkan peranan penting dalam imunosupresi akibat infeksi HIV (Cotrez, dkk., 1997).
Peranan aktivasi monosit akibat infeksi HIV dalam menyebabkan apoptosis limfosit CD4+ dibuktikan oleh penelitian Gascon, dkk (2002). Pada penderita dengan peningkatan ekspresi HLA-DR pada monosit memiliki jumlah limfosit T CD4+ yang lebih rendah secara signifikan dibandingkan dengan penderita tanpa marker aktivasi ini. Pada penderita yang memiliki marker aktivasi HLA-DR pada monosit yang persisten setelah 24 minggu pemberian ARV menunjukkan jumlah limfosit T CD4+
yang tetap rendah sampai 2 tahun pengamatan dibandingkan dengan penderita tanpa marker aktivasi yang persisten. Dari hasil penelitian ini dapat disimpulkan bahwa aktivasi monosit yang didefinisikan dengan peningkatan ekspresi HLA-DR berhubungan dengan penurunan jumlah limfosit T CD4+ pada infeksi HIV dan menyebabkan respon yang buruk setelah pemberian ARV.
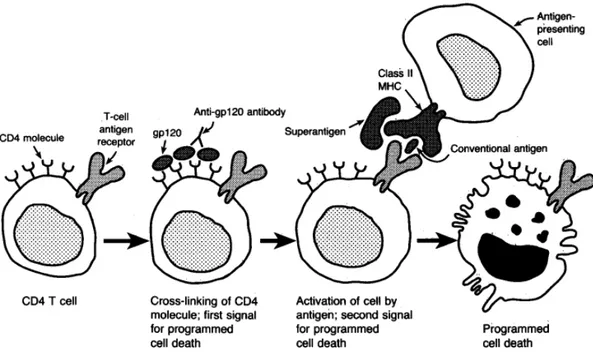
Gambar 2.5 Apoptosis (programmed cell death) pada infeksi HIV
Sinyal pertama oleh cross-linking molekul CD4 oleh gp120 dan anti-gp120 serta aktivasi T cell receptor (TCR) oleh antigen maupun superantigen sebagai sinyal kedua menyebabkan apoptosis limfosit T CD4+.
Sumber: Pantaleo G, Graziosi C, Fauci AS, 1993. The immunopathogenesis of Human Immunodeficiency Virus infection. N Engl J Med 328: 327-35.
Peranan monosit dalam menimbulkan apoptosis diperkuat oleh penelitian Cotrez, dkk (1997) yang menunjukkan bahwa apoptosis limfosit T CD4+ membutuhkan 2 sinyal dari APC, pertama stimulasi antigen pada T cell receptor (TCR) oleh APC melalui MHC clas II dan kedua adalah protein HIV gp 120. Kedua
sinyal ini akan membuat limfosit T CD4+ yang tidak terinfeksi virus menjadi teraktivasi dan kemudian mengalami apoptosis (gambar 2.5). Apoptosis terjadi tanpa adanya replikasi virus dalam sel.
Lebih lanjut pada limfosit T CD4+ yang teraktivasi terjadi peningkatan ekspresi L-selectin (CD62L) sebuah homing receptor atau reseptor pada limfosit yang menyebabkan limfosit mengalami migrasi ke limfonodi dari sirkulasi perifer (homing). Gambaran lalulintas sel (cell trafficking) pada infeksi HIV mengalami perubahan). Pada lalu lintas sel yang normal sel limfosit T CD4+ naïve bermigrasi ke dalam limfonodi dan jaringan limfoid lainnya. Jika tidak terdapat antigen akan kembali ke dalam sirkulasi dalam 1-2 hari. Akan tetapi pada infeksi HIV sel limfosit T CD4+ akan kontak dengan APC menyebabkan peningkatan ekspresi homing receptor CD62L. Rangsangan reseptor ini oleh ligannya akan menyebabkan peningkatan jumlah sel yang bermigrasi ke limfonodi, di dalam limfonodi sinyal lainnya dari APC akan menyebabkan apoptosis sel T CD4+. Hal ini mungkin dapat menjelaskan mengapa limfadenopati sering terjadi pada waktu yang bersamaan dengan terjadinya penurunan jumlah limfosit T CD4+ dalam darah (Wang, dkk., 1999; Kirschner, dkk., 2000).
2.8.2 Apoptosis sel limfosit T CD4+, peranan Fas/APO-1/CD95
Penelitian lebih lanjut mengenai mekanisme biologis apoptosis menunjukkan suatu reseptor memediasi apoptosis pada limfosit T CD4+. Data terbaru menunjukkan, interaksi antara Fas dan FasL, bersamaan dengan aktivasi dari TCR oleh antigen menyebabkan apoptosis pada limfosit T CD4+. Jadi, aktivasi dari limfosit oleh
antigen dibutuhkan untuk terjadinya apoptosis pada infeksi HIV yang dimediasi oleh interaksi Fas-FasL (Wang, dkk., 1994).
Seperti yang dijelaskan sebelumnya, pada infeksi HIV limfosit T dalam sirkulasi memiliki derajat aktivasi yang tinggi dibandingkan dengan kontrol yang ditunjukkan oleh peningkatan marker aktivasi. Sel T yang teraktivasi menyebabkan peningkatan ekspresi Fas reseptor pada permukaannya. Penelitian Debatin, dkk (1994) menunjukkan bahwa terdapat peningkatan yang signifikan ekspresi Fas pada sel T CD4+ dan CD8+ dibandingkan dengan kontrol. Mekanisme peningkatan ini dapat disebabkan langsung oleh infeksi HIV atau stimulasi oleh produk HIV atau aktivasi secara umum pada sel T.
Penelitian lain juga menunjukkan bahwa ekspresi Fas pada sel T CD4+ naïve mengalami peningkatan sesuai dengan progresifitas penyakit dan mengalami penurunan sebagai akibat terapi ARV. Bohler, dkk (2000) meneliti 37 penderita HIV dan 12 kontrol sehat. Ekspresi Fas pada resting/naïve dan primed/memory sel T CD4+ serta CD8+ diperiksa secara kuantitatif menggunakan four-colour dan three-colour flow cytometry. Hasilnya menunjukkan bahwa terdapat peningkatan sel yang mengekspresikan Fas secara kuantitatif pada resting/naïve T cells dan peningkatan persentase CD95high primed/memory T cells sesuai dengan progresifitas penyakit. Ekspresi Fas pada resting/naïve CD4+ cells mengalami penurunan sampai mendekati nilai normal pada penderita dengan jumlah total limfosit T CD4+ darah perifer yang stabil setelah 6 bulan pemberian ARV, sedangkan ekspresi Fas pada sel limfosit T CD8+ tetap tinggi dibandingkan kontrol.
Makrofag yang terinfeksi HIV merupakan sumber dari FasL. Penelitian menunjukkan bahwa infeksi HIV pada sel monositik tidak hanya menyebabkan peningkatan ekspresi Fas pada permukaannya tetapi juga peningkatan produksi FasL. Makofag dari penderita sehat juga menunjukkan peningkatan produksi FasL setelah diinfeksi dengan HIV monositotropic strain. Fakta tersebut mengindikasikan bahwa sel makrofag yang terinfeksi HIV dan APC yang lain seperti sel dendritik tidak hanya sebagai reservoir virus tetapi juga sumber dari FasL (Badley, dkk., 1996).
Penelitian Debatin, dkk (1994) menunjukkan bahwa terdapat peningkatan ekspresi Fas secara signifikan pada sel limfosit T CD4+ dan CD8+ anak penderita HIV dibandingkan kontrol. Yang menarik, sensitasisi terhadap Fas terutama mempengaruhi limfosit T CD4+. Hal ini mengindikasikan bahwa kostimulasi reseptor CD4 pada sel T oleh ikatan silang gp120-anti gp120 membuat sel T CD4+ menjadi lebih sensitif terhadap apoptosis yang diinduksi oleh Fas dibandingkan dengan sel limfosit T CD8+. Produk lain dari HIV, Trans-Activator Transcription (Tat), yang diproduksi oleh sel yang terinfeksi HIV dapat melakukan penetrasi ke dalam sel yang tidak terinfeksi dan dan bekerja seperti “toxin” menyebabkan sel menjadi hipersensitif terhadap apoptosis yang dimediasi oleh Fas (Accornero, dkk., 1997) (gambar2.6).
Gambar 2.6 Apoptosis limfosit CD4+ yang dimediasi oleh Fas/APO-1. Model bagaimana produk gen HIV seperti HIV Tat dan gp120 yang diproduksi oleh sel T CD4+ dan makrofag yang terinfeksi HIV dapat mengaktivasi sel T CD4+ yang tidak terinfeksi menjadi sensitif terhadap apoptosis yang dimediasi oleh Fas/APO-1. Sumber: Krammer PH, Dhein J, Walczak H, Behrmann I, Mariani S, Matiba B, Fath M, Daniel PT, Knipping E, dkk., 1994. The role of APO-1-mediated apoptosis in the immune system. Immunological Reviews 142: 175-91
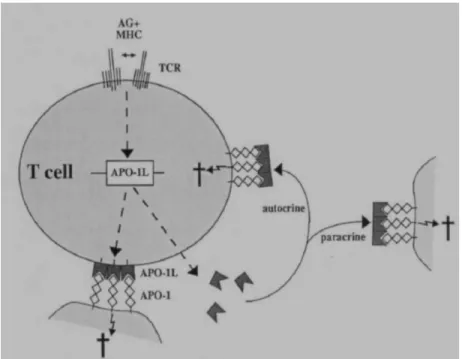
Bagaimana mekanisme kerja sistem Fas-FasL dalam menimbulkan apoptosis sel limfosit T? interaksi antara Fas dengan FasL adalah spesifik dalam menimbulkan apoptosis sel. Sel T yang teraktivasi akan menghasilkan FasL/APO-1L yang akan berinteraksi dengan Fas reseptor sel yang berada disekitarnya (bystander/neighboring T-cell), akibat interaksi ini sel disekitarnya akan mengalami apoptosis, mekanisme ini dikenal dengan fratricide atau bystander effect. FasL yang diproduksi sel limfosit T yang teraktivasi dapat berinteraksi dengan Fas reseptor dan menyebabkan apoptosis pada dirinya sendiri yang dikenal dengan autocrine suicide dan juga sel yang jauh yang dikenal dengan paracrine death (gambar 2.7). Mekanisme paracrine death ini dapat menjelaskan mekanisme sel limfosit T yang teraktivasi dalam menyebakan apoptosis sel lain yang mengekspresikan Fas reseptor seperti sel eritroid progenitor yang terpapar dengan IFN-γ (Krammer, dkk., 1994).