Ternak sapi membutuhkan protein untuk membangun dan menjaga protein jaringan dan organ tubuh yang normal, serta untuk meningkatkan produksi yang optimal. Bahan suplemen protein untuk ransum sapi relatif mahal, oleh karena itu diperlukan seleksi yang tepat dan penggunaan suplemen protein yang efisien agar dapat mengurangi biaya produksi. Beberapa kriteria yang harus dipertimbangkan dalam penggunaan suplemen protein antara lain: palatibilitas, degradasi protein rumen, kualitas protein, absorbsi asam amino di usus halus, biaya per unit protein, ketersediaan dan konsistensi produk, dan dampaknya terhadap penampilan produksi ternak (Stern et al. 2006). Suatu model nutrisi baru untuk sistem pemberian protein pakan ternak sapi perah di Amerika telah berkembang dari sistem yang berbasis protein kasar (NRC 1978), ke sistem yang lebih kompleks berbasis degradabilitas protein rumen (Rumen Degradable Protein= RDP), protein tidak terdegradasi di rumen (Rumen Undegradable Protein=RUP) dan RUP tercerna di usus halus (NRC 2001). Jadi, model nutrisi baru ini lebih unggul dari pada sistem sebelumnya, karena dirancang guna memenuhi kebutuhan protein bagi mikrobial rumen dan juga pasokan protein yang lolos degradasi rumen, tetapi tercerna dalam usus halus.
Isyarat tentang adanya defisiensi protein pada ternak ruminansia di negara-negara tropis termasuk Indonesia sebetulnya telah lama dikemukakan oleh Leng (1991). Menurut Rossi & Silcox (2007), defisiensi protein dapat mengakibatkan penurunan bobot badan, laju pertumbuhan yang rendah, rendahnya reproduksi, mengurangi produksi susu, dan menurunkan nafsu makan. Bila defisiensi protein terjadi sebelum sapi beranak, maka akan menurunkan produksi kolustrum, dan kondisi pedet lemah pada saat lahir. Guna meningkatkan efisiensi konversi pakan dan produksi ternak pada ruminansia yang diberi pakan berkualitas rendah termasuk pakan berbasis jerami padi perlu suplementasi nutrien-nutrien, seperti mineral, urea, dan protein yang lolos fermentasi dalam rumen (bypass protein) tetapi tercerna dalam usus halus (Leng 1991; Galina et al. 2000; Ortiz et al. 2001; Loest et al. 2001). Menurut Leng (1991) defisiensi nutrien-nutrien yang diperlukan oleh mikroorganisme rumen menurunkan pertumbuhan mikrobial,
yang pada akhirnya dapat menurunkan kecernaan dan konsumsi pakan, utamanya pakan berserat misalnya jerami padi.
Jerami padi telah dikenal sebagai salah satu makanan pokok untuk ruminansia di Indonesia, dan sudah biasa diberikan pada ternak sapi, terutama didaerah persawahan padi. Kebiasaan ini mengingat penyediaan hijauan makanan ternak semakin berkurang seiring dengan menyempitnya kepemilikan lahan sebagai akibat pertambahan jumlah penduduk. Produksi jerami padi sangat berlimpah seiring dengan meningkatnya produksi padi di Indonesia. Data produksi jerami padi di Indonesia menunjukkan potensi cukup besar yaitu 60 135 501 ton bahan kering (BPS 2004). Disisi lain, sebaran populasi ternak sapi pedaging di Indonesia tidak merata dan populasinya masih rendah yaitu 10 726 347ekor pada tahun 2004 (DITJENAK 2004). Pada Tabel 1 disajikan data tentang populasi sapi pedaging dan produksi jerami padi di Indonesia. Berdasarkan data tersebut, menunjukkan bahwa jerami padi di Indonesia sebagai produk samping hasil pertanian di daerah persawahan padi, tersedia dalam jumlah yang cukup banyak. Disisi lain, sebaran jumlah sapi di berbagai propinsi di Indonesia masih rendah, sehingga terdapat kesenjangan antara ketersediaan jerami padi dengan populasi sapi pedaging. Dengan demikian, jerami padi di daerah persawahan padi banyak yang tidak termanfaatkan sebagai pakan sapi. Akibatnya, masih banyak terlihat adanya pembakaran jerami padi di daerah persawahan padi.
Penggunaan jerami padi sebagai pakan mempunyai keterbatasan karena nilai protein dan nilai cernanya rendah, selain itu juga kurang palatabel. Kandungan dinding sel jerami padi kurang lebih 86% dari BK, sedangkan komponen utama dari dinding sel adalah selulosa, hemiselulosa dan lignin dengan kandungannya secara berturut-turut adalah 30-51%, 6-28% dan 4-10% dari BK (Doyle et al. 1986). Sedangkan kandungan protein kasarnya juga rendah yaitu rata-rata 4.21%, TDN (43.2%), serat kasar (32.5%), lemak kasar (1.47%), abu (16.9%), bahan ekstrak tanpa nitrogen (45%), Ca (0.41%), dan P (0.29%) dari bahan kering (Sutardi 1981). Lebih lanjut disebutkan bahwa peningkatan fermentabilitas jerami sebagai pakan sapi dapat diupayakan dengan memberikan beberapa perlakuan pendahuluan (pretreatment), misalnya dengan perlakuan kimia (perlakuan alkali,
Tabel 1. Populasi sapi pedaging dan produksi jerami padi di Indonesia
No. Propinsi Jerami padi
(ton bh. kering)a Sapi pedaging (ekor)b 1 NAD 1 860 233 702 689 2 Sumut 4 157 211 248 971 3 Sumbar 2 134 039 623 520 4 Riau 725 917 116 035 5 Jambi 790 487 150 220 6 Sumatera selatan 3 155 720 506 203 7 Bengkulu 560 191 79 122 8 Lampung 2 502 371 388 977
9 Kep Bangka Belitung 37 380 20 831
10 DKI Jakarta 14 852 0 11 Jawa Barat 9 494 717 227 504 12 Jawa Tengah 8 261 406 1 346 955 13 DI Yogyakarta 670 963 226 489 14 Jawa Timur 8 570 537 2 517 227 15 Banten 1 841 841 10 432
16 Nusa Tenggara Barat 1 646 219 427 960
17 Nusa Tenggara Timur 908 046 523 036
18 Kalimantan Barat 1 843 664 160 500 19 Kalimantan Tengah 1 156 465 45 530 20 Kalimantan Selatan 2 239 715 175 790 21 Kalimantan Timur 722 165 57 268 22 Sulawesi Utara 466 817 126 026 23 Sulawesi Tengah 888 946 196 040 24 Sulawesi Selatan 3 906 119 751 283 25 Sulawesi Tenggara 428 684 214 470 26 Gorontalo 190 592 168 267 27 Maluku 56 358 64 047 28 Papua 76 841 71 346 29 Maluku Utara 98 091 35 132 INDONESIA 60 135 501 10 726 347
amoniasi), perlakuan biologi (fermentasi dengan berbagai jenis mikroorganisme erob maupun anerob), dan perlakuan fisik (penggilingan, pembuatan pellet, dan pemanasan pada tekanan tinggi). Metode-metode tersebut sudah banyak dikaji dan telah memperlihatkan hasil cukup baik. Tetapi, penerapan metode-metode tersebut masih menghadapi banyak kendala apabila dalam skala yang besar, sehingga sulit diadopsi oleh peternak di Indonesia.
Berbagai jenis tanaman leguminosa yang memiliki kandungan protein tinggi juga telah banyak dicobakan ke ternak ruminansia sebagai suplemen. Hasil penelitian Manurung (1989) menunjukkan bahwa suplementasi hijauan lamtoro (Leucaena leucocephala) pada ransum berbasis jerami padi mendapatkan pertambahan bobot badan sapi jantan peranakan ongol sebesar 394 g/ekor/hari. Penerapan penggunaan suplementasi hijauan lamtoro juga sulit, karena produksi hijauan leguminosa ini terbatas dan ketersediaannya tidak kontinyu akibat semakin menyempitnya lahan penanaman hijauan pakan.
Suplemen urea dapat dikombinasikan dengan sumber energi yang siap tersedia (Readily available energy sources) misalnya molases, dengan cara disemprot pada pakan berserat berkualitas rendah, seperti jerami padi (Preston & Leng 1987). Namun demikian menurut Sahoo et al. (2004) berbagai upaya untuk mempopulerkan campuran urea molasses di tingkat petani di India tidaklah berhasil karena sering terjadi insiden kelebihan urea (overdosage) atau campurannya yang tidak homogen dengan pakan berserat, sehingga menimbulkan keracunan bagi ternak ruminansia. Hasil penelitian Sahoo et al. (2004) tentang suplementasi urea-molases pada ransum kerbau jantan berbasis jerami gandum menunjukkan pertambahan bobot badan yang nyata dibanding ransum kontrol atau tanpa suplemen (kontrol=213 g/ekor/hari dan dengan suplemen= 356 g/ekor/hari). Lebih lanjut juga disebutkan bahwa difisit protein pada pakan serat berkualitas rendah dapat dilakukan dengan memberikan tambahan protein pakan yang tidak tercerna dalam rumen (RUP) dan campuran urea-molases serta mineral dalam ransum ternak ruminansia.
Menurut Sellier (2003), ada tiga pendekatan untuk memasok protein bagi ternak ruminansia: (1) pasokan sumber protein dalam ransum yang tidak mudah didegradasi oleh enzim mikrobial dalam rumen (RUP) dan dapat langsung ke usus
halus, sehingga akan meningkatkan jumlah asam amino pakan yang mudah diserap; (2) optimalisasi fermentasi rumen untuk meningkatkan jumlah protein mikrobial, sehingga akan meningkatkan asam amino mikrobial yang mudah diserap; (3) enkapsulasi spesifik asam amino (atau asam amino analog) menggunakan agen proteksi degradasi mikrobial rumen. Stern et al. (2006) menegaskan bahwa setelah memaksimalkan sintesis protein mikrobial, jumlah substansi protein tidak terdegradasi di rumen (RUP) dari suplemen protein harus diinkoporasi kedalam pakan ruminansia yang berproduksi tinggi. Protein mikrobial rumen merupakan protein berkualitas tinggi untuk ruminansia, yaitu memiliki kecernaan yang tinggi didalam usus halus. Mengingat laju pertumbuhan mikrobial rumen mempengaruhi pasokan asam amino ternak ruminansia, maka penting untuk memaksimalkan sintesis protein mikrobial dalam rumen. Sintesis protein mikrobial dalam rumen memberikan sejumlah besar protein yang disuplai ke usus halus ruminansia, yaitu sampai 50-80% dari total protein yang dapat diserap. Jumlah protein mikrobial yang masuk ke usus halus tergantung pada ketersediaan nutrien dan efisiensi penggunaan nutrien-nutrien oleh bakteri rumen. Menurut Brunetti (2004) sekitar 60% protein murni didegradasi menjadi ammonia dalam rumen yang digunakan sebagai makanan mikrobial, selanjutnya akan menjadi protein mikrobial bagi ternak ruminansia, sedangkan yang 40% lagi lolos degradasi ke organ pasca rumen untuk diserap dalam usus halus. Disisi lain 100% Nitrogen bukan protein (NBP) dalam makanan didegradasi menjadi ammonia rumen, dan jika ammonia tersebut tidak dimetabolismekan oleh mikrobial, akan diserap oleh dinding rumen, yang pada akhirnya akan kembali menjadi nitrogen darah yang berasal dari urea. Oleh karena itu, berbagai alasan ini oleh para ahli gizi ternak ruminansia pada jaman sekarang ingin mengetahui seberapa banyak protein terlarut (protein soluble) dan bypass dalam ransum.
Penggunaan Nitrogen Bukan Protein untuk Optimalisasi Biosintesis Protein Mikrobial Rumen
Ruminansia memiliki empat bagian perut, yaitu rumen, retikulum, omasum dan abomasum. Bagian yang terbesar (rumen) mengandung banyak mikroorganisme untuk mencerna pakan. Lebih separuh dari protein yang
dikonsumsi sapi didegradasi dalam rumen menjadi peptide, asam amino, dan ammonia oleh aksi enzim-enzim yang diproduksi oleh mikroorganisme-mikroorganisme tersebut. Selain itu, senyawa NBP, misalnya urea (NH2)2C=O
juga didegradasi menjadi karbon dioksida (CO2) dan ammonia (NH3) didalam
rumen. Mikroorganisme-mikroorganisme rumen akan menggunakan ammonia yang dilepas dari degradasi protein dan NBP untuk dikonversi menjadi protein mikrobial (Sewell 1993; Rossi & Silcox 2007). Urea mudah larut dalam rumen dan mikroorganisme rumen akan cepat mendekomposisi menjadi NH3 dan CO2.
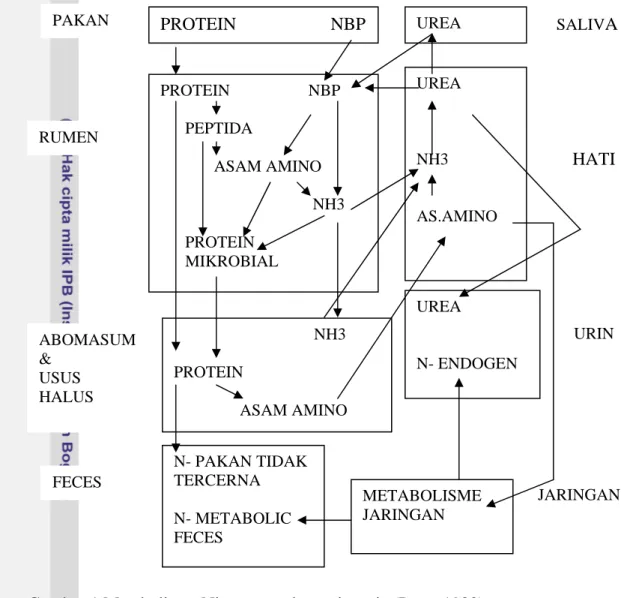
Menurut Perry (1980), nasib NH3 yang dilepas dari urea kemudian melewati
dua jalur (Gambar 1), yaitu 1) digunakan untuk sintesis protein mikrobial, dan 2) diabsorbsi melalui dinding rumen masuk pembuluh darah yang membawanya sampai ke hati. Hati akan mendetoxifikasi ammonia, dengan mengkonversi ammonia menjadi urea, untuk selanjutnya diekskresi ke urin. Tetapi, sebagian urea ini juga masuk kembali ke rumen melalui saliva atau langsung menembus dinding rumen (dengan melalui saluran darah) masuk kedalam cairan rumen. Namun demikian, bila NH3 yang terlepas dari rumen terlalu cepat, kapasitas NH3
di hati akan berlebihan, dan akan meluap ke sistem pembuluh darah. Level NH3
yang tinggi dalam sirkulasi darah dapat menyebabkan keracunan dan bahkan kematian. Fenomena ini menunjukkan bahwa, urea akan efektif menjadi sumber protein sapi, bila kondisi didalam rumen menguntungkan bagi mikroorganisme untuk menggunakan ammonia dari urea sebelum NH3 lepas dari rumen.
Penggunaan urea oleh mikroorganisme rumen tergantung pada pertumbuhan populasi mikrobial dan jumlah nutrien esensial yang diperlukan untuk pertumbuhan yang cepat, seperti vitamin, mineral, dan sumber energi yang siap tersedia (readily available source of energy). Jadi, pemenuhan kebutuhan nutrisi bagi mikrobial rumen sangat mempengaruhi metabolisme Nitrogen dalam rumen (Bach et al. 2005; Stern et al. 2006). Ammonia yang berlebih dapat digunakan lebih efisien pada ransum yang memiliki energi tinggi (seperti pati dan biji-bijian), bila dibandingkan dengan ransum yang berenergi rendah (Sewell 1993). Karbohidrat yang mudah terfermentasi seperti pati dan gula lebih efektif dari pada sumber karbohidrat lain, misalnya selulosa dalam mendukung pertumbuhan mikrobial rumen (Stern & Hoover 1979). Pendapat ini juga sejalan dengan
PAKAN PROTEIN NBP UREA SALIVA UREA NH3 AS.AMINO PROTEIN NBP PEPTIDA ASAM AMINO NH3 PROTEIN MIKROBIAL
Gambar 1 Metabolisme Nitrogen pada ruminansia (Perry 1980).
Hoover & Stokes (1991) bahwa bila jumlah bahan organik yang terfermentasi dalam rumen meningkat maka sintesis protein mikrobial juga meningkat. Beberapa percobaan in vitro (Stern et al. 1978) dan in vivo (Casper & Schingoethe 1989; Cameron et al. 1991) telah dilakukan bahwa infusi peningkatan sejumlah karbohidrat yang mudah terfermentasi dapat menurunkan konsentrasi N-ammonia, karena meningkatnya konsumsi N oleh mikrobial rumen. Ditemukan pula bahwa bila laju degradasi protein melebihi laju fermentasi karbohidrat, maka sejumlah besar N dapat hilang sebagai ammonia, dan sebaliknya bila laju fermentasi karbohidrat melebihi laju degradasi protein, maka sintesis protein mikrobial dapat
RUMEN HATI UREA N- ENDOGEN URIN NH3 PROTEIN ASAM AMINO ABOMASUM & USUS HALUS N- PAKAN TIDAK TERCERNA N- METABOLIC FECES JARINGAN FECES METABOLISME JARINGAN
menurun (Nocek & Russel 1988). Ditegaskan oleh Jouany (1991) mikrobial rumen selain membutuhkan N (NH3, oligopeptida untuk bakteri, peptida untuk
protozoa), juga rantai karbon (VFA rantai cabang) dan ATP sebagai sumber energi untuk sintesis protein mikrobial.
Berbagai metoda digunakan untuk memprediksi sintesis protein mikrobial rumen antara lain marker mikrobial ribonucleic acid (RNA) dan di-amino-pimilic acid (DAPA), serta isotop (35S, 15N, 32P), namun demikian proses penggunaan marker-marker atau isotop tersebut sangat komplex dan sulit diterapkan di kondisi lapang (Singh et al. 2007). Metode lain yang dapat digunakan untuk memprediksi sintesis protein mikrobial adalah melalui derivate purin yang diekskresikan melalui urin. Purin (hasil degradasi asam nukleat dalam rumen) yang diabsorbsi dikonversikan menjadi derivate purin yaitu berupa asam urat dan allantoin. Purin yang diekskresikan per hari mempunyai korelasi linier dengan jumlah purin mikrobial yang diabsorbsi. Dengan asumsi rasio protein dan purin konstan dalam populasi mikrobial, maka ekskresi derivate purin digunakan sebagai index untuk menentukan protein mikrobial (Chen et al. 1990; Singh et al. 2007).
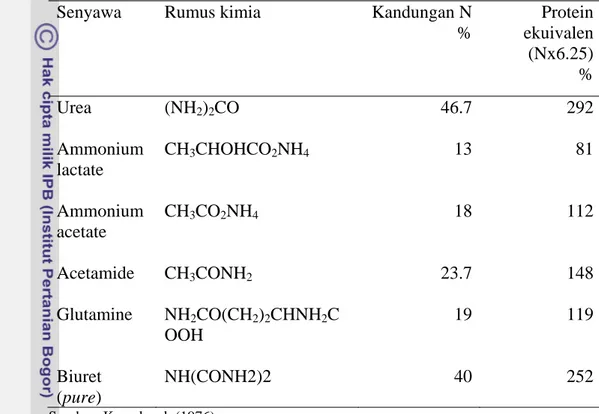
Prioritas utama dalam pemberian pakan ruminansia adalah menjamin tidak ada defisiensi nutrien dalam pakan yang mempengaruhi pertumbuhan mikrobial. Lebih lanjut Galo et al. (2003) menegaskan bahwa NBP dalam pakan sapi pedaging dan sapi perah, dapat digunakan secara efektif sebagai sumber nitrogen untuk mikrobial rumen. Urea sebagai suplemen ransum ruminansia sering dipakai di beberapa negara, terutama yang memiliki keterbatasan tersedianya protein pakan dan secara ekonomi penggunaannya menguntungkan. Menurut Kowalczyk (1976) senyawa-senyawa NBP yang dapat digunakan sebagai pengganti protein untuk ruminansia harus tidak mahal, digunakan mikrobial rumen dengan baik, dan tidak berpengaruh negatip terhadap kesehatan ternak. Dibandingkan dengan senyawa NBP yang lain (Tabel 2), harga urea tidak mahal untuk setiap unit protein ekuivalen (N x 6.25), sehingga memungkinkan untuk menyusun ransum dengan harga yang relatif murah (Rossi & Silcox 2007). Disamping itu urea sudah dikenal dan secara luas digunakan petani di Indonesia.
Acetamide, glycine dan glutamine dihidrolisis dengan laju yang lebih lambat dari pada urea, dan Nitrogen yang terkandung dapat digunakan efisien oleh ternak
ruminansia, tetapi penggunaannya dalam praktek pakan adalah terbatas karena harganya mahal. Biuret, relatif tahan hidrolisis dalam rumen, tetapi perlu adaptasi yang lama untuk penggunaannya oleh mikrobial rumen secara efisien. Kondisi
Tabel 2. Senyawa-senyawa Nitrogen bukan protein Senyawa Rumus kimia Kandungan N
% Protein ekuivalen (Nx6.25) % Urea (NH2)2CO 46.7 292 Ammonium lactate CH3CHOHCO2NH4 13 81 Ammonium acetate CH3CO2NH4 18 112 Acetamide CH3CONH2 23.7 148 Glutamine NH2CO(CH2)2CHNH2C OOH 19 119 Biuret (pure) NH(CONH2)2 40 252 Sumber: Kowalczyk (1976)
penggunaan ini dan tingkat penggunaan biuret untuk ternak ruminansia masih kontroversi (Fonnesbeck et al. 1975).
Penggunaan urea memiliki keterbatasan yaitu terlalu cepatnya urea melepaskan ammonia begitu kontak dengan enzim urease di dalam cairan rumen (Coppock et al. 1976; Visek 1984). Bloomfield et al. (1960) mendapatkan bahwa kecepatan pelepasan ammonia dari hidrolisis urea dapat mencapai 4 kali lebih cepat daripada kecepatan penggunaannya oleh mikrobial rumen.
Upaya untuk meningkatkan efisiensi penggunaan urea adalah melalui sinkronisasi laju fermentasi karbohidrat dengan laju pelepasan ammonia dari fermentasi urea. Helmer & Bartley (1971) menyebutkan bahwa penggunaan pati berpengaruh lebih baik sebagai sumber energi karena pati dalam menyediakan kerangka karbon dan energi lebih sejalan dengan waktu pelepasan ammonia dari
fermentasi urea dalam rumen. Lebih lanjut, Kowalczyk (1976) menegaskan bahwa pakan yang mengandung gula (molasses, gula tebu) dan pati (biji-bijian, umbi-umbian) sangat baik dan cocok untuk suplementasi dengan urea.
Hasil penelitian Prasetiyono (1992) menunjukkan bahwa sinkronisasi kecepatan pelepasan amonia asal urea dengan sumber energi asal ubi jalar yang dikukus telah mampu memacu pertumbuhan mikrobial rumen. Namun demikian kenyataannya sulit diterapkan karena membutuhkan bahan sumber energi yang terlalu banyak dan energi bahan bakar untuk pengukusan yang relatif kurang praktis. Hasil penelitian Chanjula et al. (2004) menunjukkan bahwa sinkronisasi penggunaan urea dengan pati yang berasal dari ubi kayu dan jagung dalam ransum sapi perah memberikan respon yang tidak berbeda terhadap penampilan produksi sapi perah, namun menghasilkan income over feed cost yang lebih tinggi pada pati asal ubi kayu (54.0 US$/bulan vs 51.40 US$/bulan). Fenomena ini menunjukkan bahwa ubi kayu dapat dijadikan bahan sumber energi yang potensial sebagai pakan ruminansia.
Nampaknya berbagai cara telah dilakukan dalam upaya untuk menekan biaya penggunaan protein alami dengan menggunakan pengganti nitrogen bukan protein, seperti urea. Teknik perlambatan pelepasan amonia di rumen (slow release of ammonia=SRA) dari hidrolisis urea telah banyak diminati pada akhir-akhir ini, karena dipandang lebih efisien dan praktis dalam penerapannya. Galo et al. (2003) menemukan bahwa SRA terbuat dari urea yang dilapisi (coated urea=CU) terbukti mampu meningkatkan efisiensi penggunaan Nitrogen oleh mikrobial rumen. Selanjutnya, Antonelli et al. (2004) menemukan bahwa 20% urea yang diekstrusi dengan 80% pati jagung dapat mencegah keracunan ammonia dari hidrolisis urea dalam rumen. Menurut Stanton & Whittier (2006) keracunan urea ditandai dengan tremor, salivasi yang berlebihan, bernapas terengah-engah, kembung, dan tetani. Disebutkan pula bahwa ternak sapi biasanya mati bila level ammonia darahnya mencapai 5 mg%. Lebih lanjut disebutkan bahwa penggunaan bahan sumber energi akan memperbaiki penggunaan urea dalam ransum.
Peneliti lain, Coppock et al. (1976) menemukan bahwa produk pati-urea tergelatinisasi (GSUP= Gelatinized Starch Urea Product) melalui pemasakan ekstrusi campuran sorgum dan urea, dengan kandungan protein kasar GSUP
sebesar 70% dapat menurunkan konsentrasi amonia rumen dibandingkan kontrol yang menggunakan urea dalam formula konsentrat sapi perah laktasi.
Penelitian Helmer et al. (1970) mengungkapkan bahwa produk pemasakan ekstrusi campuran pati jagung dan urea, dapat meningkatkan efisiensi penggunaan ammonia untuk dapat dikonversi menjadi protein mikrobial rumen, dibandingkan tanpa ekstrusi. Hal ini karena adanya gelatinisasi pati pada proses ekstrusi dapat menyediakan energi yang lebih besar, sehingga dapat meningkatkan penggunaan amonia oleh mikrobial rumen. Produk ekstrusi ini dengan kadar protein kasar 34% menghasilkan efisiensi yang tinggi dalam penggunan nitrogen oleh mikrobial rumen. Disebutkan pula bahwa kompleks pati-urea tersebut juga merupakan sumber energi yang baik serta memungkinkan sebagai substitusi protein untuk ruminansia.
Proses ekstrusi sering digunakan dalam proses pengolahan bahan pangan dan bahan yang dipakai kebanyakan merupakan sumber karbohidrat. Menurut Muchtadi et al. (1988) proses ekstrusi bahan pangan adalah suatu proses dimana bahan tersebut dipaksa mengalir dibawah satu atau lebih kondisi operasi seperti pencampuran, pemanasan dan pemotongan melalui suatu cetakan yang dirancang untuk membentuk hasil ekstrusi yang bergelombang kering (puff-dry). Alat untuk melakukan ekstrusi adalah ekstruder. Lebih lanjut disebutkan bahwa, ada dua jenis ekstruder berdasarkan jumlah ulir yang dipergunakan dalam proses ekstrusi yaitu ekstruder berulir tunggal dan ekstruder berulir ganda. Mesin ekstruder banyak digunakan dalam pengembangan produk-produk baru seperti makanan ringan, makanan ternak atau produk modifikasi pati lainnya. Pada proses pemasakan ekstrusi digunakan aplikasi suhu tinggi dengan waktu olah yang singkat. Temperatur optimum proses ekstrusi kurang lebih 180oC dan kecepatan ulir sekitar 300 rpm dalam waktu kurang lebih 10 detik. Disebutkan pula, bahwa melalui aplikasi suhu tinggi bersaat olah singkat, maka kerusakan termal senyawa-senyawa gizi dapat diusahakan seminimal mungkin, terutama untuk protein dan vitamin, sekaligus berkemampuan merusak senyawa-senyawa anti-nutrisi dan senyawa-senyawa toksik.
Scott et al. (1976) menyatakan bahwa proses pemasakan dapat memecah dinding granula pati, sedangkan Cullison (1968) menyebutkan bahwa pemasakan
pati dapat mengakibatkan ikatan pati menjadi lebih longgar (Gambar 2) sehingga terjadi pembebasan amilosa yang akan menyebabkan daya kelarutan meningkat. Pendapat ini didukung oleh hasil penelitian Sofyan (1983) bahwa ubi kayu yang dimasak dengan pengukusan dapat mempengaruhi kelarutan pati. Sedangkan Szylit et al. (1978) menyebutkan bahwa proses pemasakan pati diantaranya bertujuan untuk mempercepat pemecahan pati oleh mikrobial rumen sehingga pelepasan energinya akan sejalan dengan pelepasan ammonia dari senyawa nitrogen bukan protein yang mudah dihidrolisis seperti urea dan akibatnya ammonia dapat digunakan lebih efisien dalam rumen.
Gambar 2 Struktur molekul amilosa amilopektin pada proses pengembangan granula pati
Pati terdiri dari suatu rantai unit-unit D-glucopyranosil dan memiliki rumus umum (C6H10O5)n dengan n = 250 sampai diatas 1000. Ada 2 komponen utama
pada molekul pati yaitu amylopektin (75-80% ) yang bentuknya merupakan suatu rantai bercabang dan sisanya adalah amylosa yang berbentuk linear (Austin 1986).
Ubi kayu merupakan sumber pati yang sangat potensial, dan merupakan tanaman lokal yang mudah dibudidayakan di Indonesia serta dapat tumbuh sepanjang tahun. Produksi ubi kayu di Indonesia cukup tinggi yang ditunjukkan data statistik selama lima tahun terakhir mengalami peningkatan yaitu dari 18 523 810 ton pada tahun 2003 menjadi 18 950 274 ton pada tahun 2007 (BPS 2007).
Menurut Gerpacio et al. (1979) ubi kayu merupakan sumber karbohidrat yang relatif murah dibanding jagung dan memiliki kandungan pati yang lebih tinggi (48.49%) dari pada jagung (45.35%). Disamping itu, ubi kayu mengandung karbohidrat non-struktural lebih tinggi dari pada jagung (Sommart et al. 2000; Chanjula et al. 2003). Namun demikian, ubi kayu memiliki kandungan protein yang rendah (Kiyothong & Wanapat 2004; Wanapat & Khampa 2007). Rendahnya protein ini dapat di perbaiki dengan bahan aditif protein lainnya (Balagopalan 2002). Selain memanfaatkan umbinya, daun ubi kayu dapat digunakan sebagai sumber asam amino. Suryahadi & Amrullah (1989) menemukan bahwa daun tanaman ubi kayu dapat digunakan sebagai sumber asam amino leusin, phenilalanin dan valin, berturut-turut: 10.5, 3.6 dan 6.8% dari protein total daun ubi kayu. Penambahan asam-asam amino ini sangat tepat dalam ransum yang mengandung nitrogen bukan protein dan dapat meningkatkan produksi ternak.
Menurut Huber & Kung (1981) pada ransum yang mengandung NBP, kerangka karbon bercabang sangat potensial untuk menjadi faktor pembatas pertumbuhan mikrobial rumen. Asam-asam amino rantai cabang (branced-chain amino acids = BCAA), seperti valin, leusin, dan isoleusin di dalam rumen akan mengalami proses dekarboksilasi dan deaminasi menjadi asam-asam lemak rantai cabang. Asam isobutirat, asam 2-metilbutirat, dan asam isovalerat berturut-turut berasal dari asam amino valin, isoleusin, dan leusin. Asam-asam lemak rantai cabang ini juga mendukung dalam sintesis mikrobial rumen. Proses deaminasi dan dekarboksilasi asam-asam amino rantai cabang dapat disederhanakan pada reaksi seperti berikut (Andries et al. 1987):
R-CH(NH2)COOH + H2O Æ RCOCOOH + NH3 + 2H+ RCOCOOH + H2O Æ RCOOH + CO2 + 2H+ R = (CH3)2CH-(valin) = (CH3)2CHCH2-(leusin) = CH3CH2CH(CH3)-(isoleusin)
Proteksi Protein Kedelai dari Degradasi dalam Rumen
Perbaikan nutrisi protein ternak ruminansia dapat ditempuh melalui peningkatan pasokan protein asal mikrobial (protein mikrobial) dan protein asal pakan yang lolos degradasi. Suatu sumber protein tidak tahan terhadap degradasi mungkin hanya akan memberikan masukan protein berupa protein mikrobial saja kepada hewan induk semang. Sebaliknya pemberian protein yang tahan degradasi, disamping protein mikrobial, hewan induk semang juga akan mendapat protein asal pakan yang lolos degradasi, sehingga persediaan asam amino bagi penyerapan usus menjadi lebih banyak (Stern et al. 2006).
Menurut Stern et al. (2006), degradasi protein dalam rumen yang berasal dari protein pakan yang terkonsumsi merupakan salah satu faktor penting yang mempengaruhi suplai asam amino dalam usus halus ruminansia. Proteolisis menentukan ketersediaan N-amonia, asam amino, peptide, VFA rantai cabang, yang mempengaruhi laju pertumbuhan mikrobial dalam rumen. Laju dan tingkat proteolisis dalam rumen tidak hanya mempengaruhi sintesis protein mikrobial, tetapi juga jumlah dan kualitas protein pakan yang tidak terdegradasi yang masuk dalam duodenum. Meskipun protein mikrobial sendiri mungkin mampu mencukupi kebutuhan ruminansia yang berproduksi rendah, tetapi tidak mampu mencukupi kebutuhan ruminansia yang berproduksi tinggi, terutama untuk pertumbuhan, produksi wool, dan produksi susu. Oleh karena itu, untuk ruminansia berproduksi tinggi, disamping pemberian suplemen protein pakan yang terdegradasi dalam rumen untuk memaksimalkan protein mikrobial rumen, perlu penambahan suplemen protein pakan yang lolos degradasi rumen (Rumen Undegradable Protein= RUP) karena akan memperbaiki suplai asam amino ke ternak ruminansia (Stern et al. 2006).
Beberapa faktor penting yang mempengaruhi degradasi protein dalam rumen adalah tipe protein, solubilitas dalam rumen, pH rumen, tipe substrat, serta tingkat laju lolos degradasi rumen (Bach et al. 2005; Stern et al. 2006). Berikut ini disajikan dalam Tabel 3, contoh RUP, kecernaan protein kasar (PK) dalam usus halus (Intestinal CP Digestion= ID), dan daya serap protein pakan dalam usus halus (Intestinally Absorbable Dietary Protein= IADP) dari berbagai suplemen protein.
Solubilitas protein merupakan faktor kunci dalam menentukan kerentanan terhadap enzim protease mikrobial dan degradabilitasnya, misalnya: prolamin dan glutelin tidak larut dan lambat terdegradasi, sedangkan globulin larut dan mudah terdegradasi dalam rumen (Stern et al. 2006). Selain itu, struktur protein juga penting, misalnya: beberapa albumin dapat larut, tetapi mengandung ikatan disulfide sehingga menyebabkan lambat terdegradasi dalam rumen.
Tabel 3 Contoh RUP, ID, dan IADP dari berbagai suplemen protein.
No. Sumber protein RUP
(% PK)
ID (% RUP)
IADP (%PK)
1. Bungkil biji kapuk 55 80 43
2. Tepung kedelai 25 90 22
3. Tepung kedelai terekstrusi 66 88 58
4. Ampas bir 57 77 44
5. Tepung darah 88 63 55
6. Tepung bulu 76 67 51
7. Tepung ikan 65 80 52
8. Tepung tulang &daging (MBM) 59 55 33 Sumber: Stern et al. (2006).
Menurut Kopecny & Wallace (1982) pH optimal untuk enzim proteolitik berkisar antara 5.5 – 7.0; tetapi degradasi protein berkurang pada pH rumen yang rendah. Selain degradasi protein terjadi karena aktivitas enzim proteolitik, juga dapat disebabkan oleh aktivitas enzim lain.
Jumlah total protein yang tersedia untuk absorbsi dari usus halus tergantung pada pasokan mikrobial dan protein pakan lolos degradasi kedalam duodenum dan kecernaannya dalam usus halus (Stern et al. 2006). Disebutkan pula bahwa protein pakan yang terabsorbsi di usus halus (IADP) didefinisikan sebagai jumlah protein dari suatu specifik pakan yang tersedia untuk absorbsi dalam usus halus. Nilai IADP merupakan suatu index kualitas suplemen protein sebagai sumber RUP untuk ruminansia.
Kedelai merupakan suplemen protein untuk ternak ruminansia yang sudah banyak digunakan terutama di luar negeri seperti Amerika digunakan sebagai suplemen protein untuk ternak sapi pedaging dan sapi perah. Protein asal kedelai memiliki keunggulan tersendiri dalam kandungan asam aminonya, terutama kaya akan lysine, tetapi memiliki asam-asam amino pembatas pertama, kedua, dan ketiga masing-masing adalah methionine, valine dan isoleucine (Schingoethe 1996). Oleh karena itu, bila kedelai dikombinasikan dengan jagung (yang memiliki asam amino pembatas lysine) akan menghasilkan imbangan protein yang baik. Namun demikian, kedelai memiliki efisiensi protein yang relatif rendah, karena masih terdegradasi dalam rumen. Jumlah protein kedelai yang lolos dari fermentasi rumen adalah 25% (Stern et al. 2006). Sehingga dalam aplikasinya sebagai suplemen protein ruminansia masih perlu diproteksi agar dapat meningkatkan kemampuan lolos dari degradasi rumen yang sangat penting memberikan kontribusi bagi industri peternakan sapi pedaging dan sapi perah, serta perkembangan industri kedelai.
Upaya untuk proteksi sumber protein pakan dari degradasi dalam rumen sudah banyak dilakukan, yaitu dengan cara pemanasan bahan makanan, pemberian air minum yang banyak, dan perlakuan kimiawi serta pembungkusan protein dengan kapsul (Chalupa 1974). Orskov (2002) mencoba melindungi protein dengan asam tannin. Hasil penelitiannya memperlihatkan bahwa asam tannin dapat meningkatkan penyerapan N dalam saluran pencernaan pasca rumen dibandingkan penggunaan kapsul dan tanpa perlindungan. Peneliti lain ada yang menggunakan formaldehid sebagai protektor (Kanjanapruthipong et al. 2002; Sahoo et al. 2004; Abdullah & Awawdeh 2004). Namun demikian, penggunaan protektor tersebut kurang ekonomis dan sulit penerapannya, karena formaldehid sebagai bahan protektor memiliki potensi karsinogenik (carcinogenic effect), sehingga kurang aman bagi ternak (Leng 1991).
Menurut Leng (1991), perlakuan xylosa memberikan harapan sebagai pilihan metoda untuk proteksi protein dari degradasi rumen. Hal ini karena, selain tidak memiliki efek buruk terhadap ternak, xylosa juga sangat mudah diproduksi dan relatif tidak mahal. Xylosa dapat diperoleh dari black liquor (BL), yaitu hasil samping (by product) dari proses hidrolisis basa pada pabrik kertas.
Dijelaskan pula bahwa xylosa lebih cepat bereaksi dengan asam amino melalui reaksi pencoklatan (browning reaction) dibandingkan gula pereduksi lainnya. Temuan ini juga didukung oleh Can & Yilmaz (2002), bahwa xylosa dapat digunakan sebagai protektor protein kedelai dari degradasi rumen, melalui reaksi Mailard (non-enzimatic browning reaction) dan aman dikonsumsi ternak.
Menurut Harper et al. (1979) xylosa termasuk karbohidrat dari grup monosakarida yang memiliki lima atom karbon (pentose: C5H10O5). Hasil
penelitian Lewis et al., (1988) yang disitasi oleh Leng (1991) tentang pengaruh suplementasi pakan sapi berbahan dasar hijauan dan protein tepung kedelai yang disemprot dengan xylosa (berbahan asal cairan sulfite) dan dipanaskan pada suhu 200 oF selama 2 jam menghasilkan bobot badan harian sapi seperti yang tersaji pada Tabel 4.
Tabel 4 Pengaruh suplementasi hijauan pakan terhadap pertambahan bobot badan Sapi
Perlakuan Pertambahan bobot hidup (g/hari)
Tanpa suplemen + 7 % tepung kedelai
+ 9 % tepung kedelai + 10 % sulfite liquor + 8 % tepung kedelai + 5 % sulfite liquor
591 673 823 841
Sumber: Lewis et al. (1988) yang disitasi oleh Leng (1991).
Pemberian Pakan pada Sapi Pedaging
Pakan ternak sapi pedaging terdiri dari pakan kasar dan konsentrat (Perry 1980). Pakan kasar atau hijauan memiliki kandungan serat kasar 18% atau lebih, sedangkan konsentrat memiliki kandungan serat kasar yang rendah atau kurang dari 18% (Ensminger 1990). Bahan konsentrat yang diberikan ke sapi sebagai sumber energi antara lain: bekatul, gandum, dedak gandum, sorghum, tepung ubi kayu, sedangkan yang merupakan sumber protein antara lain: tepung kedelai, bungkil kapuk, bungkil wijen, bungkil kelapa (Perry 1980).
Prinsip dasar dalam pengembangan sistem pemberian pakan pada usaha sapi pedaging, adalah: 1) memaksimalkan fungsi rumen sapi, melalui pemberian
nutrisi yang sesuai dengan kebutuhan mikroflora rumen dan memperbaiki ekosistem rumen, 2) mengoptimalkan kebutuhan nutrisi untuk metabolisme melalui suplementasi bypass protein, karbohidrat dan lemak, 3) meningkatkan palatibilitas dan kecernaan, 4) mengoptimalkan kebutuhan mineral (Gurnadi 1993).
Menurut Frisch (1978) teknologi pakan yang digunakan untuk usaha penggemukan sapi sistem feedlot sebaiknya dengan “Grainfed” , dengan komposisi konsentrat 80 s/d 85% dan pakan kasar atau hijauan 15%. Disebutkan pula bahwa pemberian konsentrat diatas 85% dapat mengakibatkan acidosis atau asam lambung. Pakan kasar yang ditambahkan tersebut dapat menstimulasi proses ruminansi dan mencegah kemungkinan adanya gangguan pencernaan. Menurut Jesse et al. (1976) penggunaan konsentrat tinggi (lebih dari 70%) pada usaha penggemukan sapi, akan meningkatkan konsumsi pakan, laju pertumbuhan, efisiensi pakan, persentase karkas, dan persentase lemak, serta dapat menurunkan alokasi biaya pakan untuk setiap unit pertambahan berat badan. Disebutkan pula bahwa sapi pedaging mampu mengkonsumsi bahan kering pakan sebanyak 2.5 – 3% dari berat badan untuk setiap hari. Pemeliharaan sapi yang selalu dikandangkan secara feedlot dengan pakan yang dominan adalah biji-bijian, sering mengalami kekurangan vitamin A dan D, sehingga kedalam ransumnya perlu adanya penambahan vitamin A dan D (Preston & Willis 1974). Beberapa faktor yang mempengaruhi konsumsi bahan kering dan bahan organik antara lain: sifat fisik atau kimia pakan, permintaan fisiologis ternak untuk hidup pokok dan produksi sesuai dengan kapasitas saluran pencernaan, bobot hidup yang berhubungan dengan perkembangan saluran pencernaan, karena pada umumnya kapasitas saluran pencernaan meningkat seiring dengan bobot hidup sehingga mampu menampung bahan kering dalam jumlah yang banyak (Parakkasi 1999).
Komposisi Tubuh
Komponen penyusun utama dari tubuh seekor ternak adalah air, lemak, protein, dan abu. Komposisi tubuh ternak dipengaruhi oleh jenis ternak, bobot badan, umur, jenis kelamin, dan status nutrisi (Soeparno 1992). Disebutkan pula bahwa sejalan dengan kenaikan bobot badan, maka akan terjadi perubahan
komposisi tubuh, yaitu persentase air tubuh mengalami penurunan, persentase lemak meningkat, proporsi tulang menurun, sedangkan persentase protein tubuh relatif konstan. Sebaliknya, penurunan bobot badan menyebabkan peningkatan proporsi tulang, sedangkan proporsi otot sedikit menurun dan proporsi lemak tidak mengalami perubahan yang berarti (Soeparno 1992; Parakkasi 1999). Menurut Berg & Butterfield (1976) komposisi tubuh dari bobot kosong pada ternak sapi adalah air 39.8-77.6%, protein 12.4-20.6%, lemak 1.8-44.6%, dan abu 3.0-6.1%.
Beberapa metoda pendugaan komposisi tubuh telah banyak dilakukan, yaitu metode langsung dan tidak langsung. Pendugaan komposisi tubuh dengan metode langsung dapat dilakukan dengan pemotongan pada ternak. Pengukuran komposisi tubuh dengan pemotongan ternak memiliki hasil yang cukup akurat, tetapi biayanya cukup mahal. Sapi yang telah dipotong kemudian dilakukan pemisahan daging, lemak dan tulang, kemudian diambil sampel untuk analisis proksimat, sehingga dapat diketahui kadar air, lemak, protein, karbohidrat, dan mineral ternak tersebut (Astuti dan Sastradipradja 1999).
Pendugaan komposisi tubuh dengan metode tidak langsung dapat dilakukan dengan cara menginjeksikan perunut (tracer) atau dilution techniques ( McDonald et al. 1988). Beberapa syarat yang harus digunakan pada metode dilution techniques, antara lain: 1) mudah larut dan terbawa keseluruh tubuh, 2) tidak bersifat racun dan tidak berpengaruh secara fisiologis, 3) tidak disekresi dan tidak ikut dalam metabolisme tubuh, 4) konsentrasi dalam darah dapat diukur dengan mudah. Beberapa perunut yang dapat digunakan untuk menduga komposisi tubuh antara lain tritium (T2O), deuterium (D2O), potassium (40K), dan urea (Berg &
Butterfield 1976; Andrew et al. 1995).
Penggunaan metode dilution techniques dengan tracer urea banyak direkomendasikan oleh para peneliti sebelumnya, karena bahan tracer ini mudah didapat, murah harganya, dan dalam pengukurannya tidak terlalu sulit. Berdasarkan penelitian yang dilakukan Astuti dan Sastradipradja (1999) yang telah membandingkan metode langsung dengan urea space technique didapatkan hasil tidak ada perbedaan yang nyata dan disimpulkan bahwa urea space technique berkorelasi yang tinggi dengan metode langsung.
Penggunaan larutan urea sebagai perunut didasarkan atas kenyataan bahwa molekul urea dapat bercampur merata dengan cepat bersama cairan tubuh dalam waktu 12 – 15 menit pada sapi. Disamping itu, urea bukan merupakan senyawa yang asing bagi tubuh sapi, serta mudah dalam pelaksanaan dan lebih akurat hasilnya (Rule et al. 1986 ; Astuti & Sastradipradja (1999).