KARAKTERISTIK ANATOMI SKELET APENDIKULAR
BUAYA SENYULONG (Tomistoma schlegelii)
ALIF RAHMAN RAHIM PUARADA
FAKULTAS KEDOKTERAN HEWAN
INSTITUT PERTANIAN BOGOR
BOGOR
2017
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini Saya menyatakan bahwa skripsi berjudul Karakteristik Anatomi Skelet Apendikular Buaya Senyulong (Tomistoma schlegelii) adalah benar karya Saya dengan arahan dari Komisi Pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari Penulis telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.Dengan ini Saya melimpahkan hak cipta dari karya tulis Saya kepada Institut Pertanian Bogor.
Bogor, Januari 2017 Alif Rahman Rahim Puarada NIM B04120220
ABSTRAK
ALIF RAHMAN RAHIM PUARADA. Karakteristik Anatomi Skelet Apendikular Buaya Senyulong (Tomistoma schlegelii). Dibimbing oleh NURHIDAYAT dan SUPRATIKNO.
Penelitian ini bertujuan untuk mempelajari karakteristik anatomi skelet apendikular buaya senyulong (Tomistoma schlegelii) sebagai sistem lokomosi yang dikaitkan dengan fungsi dan perilakunya. Anatomi skelet kaki buaya senyulong dipelajari dengan melakukan pengamatan dan pengukuran pada tulang-tulang penyusun ekstremitas dan bidang persendian. Penelitian ini menunjukkan bahwa pada kaki depan, os scapula dan os coracoidea bertaut dan membentuk cavitas glenoidalis, os humerus lebih melengkung ke medial, olecranon pada os ulna tidak berkembang, dan di ekstremitas distal os radius terdapat facies articularis radiale yang luas untuk bersendi dengan os carpi radiale. Pada kaki belakang, os coxae bertaut dengan os sacrum, os pubis berhubungan dengan ossa gastralia, di distal os femoris tidak ditemukan os patella, di daerah tarsi, os astragalus dan os calcaneus membentuk sendi gerak. Skeleton manus dan pedis memiliki struktur yang mirip, tetapi digit V pada kaki belakang rudimenter. Struktur skelet apendikular tersebut memungkinkan buaya untuk melakukan gerakan berjalan, berenang, serta melindungi organ dan membantu respirasi. Kata kunci: ekstremitas, Tomistoma schlegelii, skelet kaki
ABSTRACT
ALIF RAHMAN RAHIM PUARADA. Morphological Characteristic of Appendicular Skeleton of False Gharial (Tomistoma schlegelii). Supervised by NURHIDAYAT and SUPRATIKNO.
This study was aimed to observe the morphological characteristic of appendicular skeleton of false gharial (Tomistoma schlegelii) as a locomotion system, associated with the function and behavior. The anatomy of appendicular skeleton of false gharial was observed and the length of bones was measured. This study shows that on the forelimb, os scapula and os coracoidea fit together formed cavitas glenoidalis, medial of os humerus more curved, olecranon of os ulna was undeveloped, and on the distal extremities of os radius there are a wide facies articularis radiale which jointed with os carpi radiale. On the hind limb, os coxae has an amphiarthrosis joint with os sacrum, os pubis jointed with ossa gastralia, on distal of os femoris the patella was absent, on tarsi region, os astragalus and os calcaneus form a motion joint. Skeleton manus and pedis has a similar structure, but the digit V of hind limb were rudiment. The appendicular skeleton structure allows false gharial to perform a walking motion, swimming also to protect organ and respiration.
KARAKTERISTIK ANATOMI SKELET APENDIKULAR
BUAYA SENYULONG (Tomistoma schlegelii)
ALIF RAHMAN RAHIM PUARADA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kedokteran Hewan
pada
Fakultas Kedokteran Hewan
FAKULTAS KEDOKTERAN HEWAN
INSTITUT PERTANIAN BOGOR
BOGOR
2017
PRAKATA
Puji dan syukur Penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga skripsi ini berhasil diselesaikan. Judul yang dipilih dalam penelitian yang akan dilakukan adalah “Karakteristik Anatomi Skelet Apendikular Buaya Senyulong (Tomistoma schlegelii)”. Adapun penulisan proposal penelitian ini dilakukan sebagai salah satu syarat melakukan penelitian dan pertimbangan yang selanjutnya akan dilanjutkan ke dalam bentuk skripsi.
Terimakasih Penulis ucapkan kepada Dr Drh Nurhidayat, MS, PAVet dan Drh Supratikno, MSi, PAVet sebagai dosen pembimbing yang telah memberikan arahan dan bimbingan untuk persiapan penyusunan dan pelaksanaan penelitian. Tak lupa ucapan terimakasih juga disampaikan kepada keluarga tercinta atas doa dan kasih sayangnya, seluruh Dosen dan Staf Bagian Anatomi atas bantuannya selama penelitian. Terimakasih juga disampaikan kepada teman-teman sepenelitian, penghuni rumah Cempaka 3, serta teman-teman dari kelas BB Astrocyte atas bantuan moral dan semangatnya. Semoga penelitian ini dapat bermanfaat bagi Penulis, Pembaca dan semua pihak yang berkepentingan.
Penulis mengetahui bahwa karya ini belum sempurna, sehingga perlu bimbingan dan arahan yang membangun demi hasil penelitian yang lebih baik.
Bogor, Januari 2017 Alif Rahman Rahim Puarada
DAFTAR ISI
DAFTAR TABEL vi DAFTAR GAMBAR vi PENDAHULUAN 1 Latar Belakang 1 Tujuan Penelitian 1 Manfaat Penelitian 1 TINJAUAN PUSTAKA 2 Ordo Crocodylia 2Ciri Morfologi dan Struktur Tubuh 2
Habitat dan Perilaku Alamiah 4
Lokomosi 5
Skelet Apendikular Reptil 7
BAHAN DAN METODE 8
Waktu dan Tempat Penelitian 8
Bahan dan Alat 8
Metode Penelitian 9
HASIL DAN PEMBAHASAN 9
Hasil 9
Tulang Gelang Bahu (Cingulum membri thoracici) 9
Tulang Lengan Atas (Skeleton brachii) 9
Tulang Lengan Bawah (Skeleton antebrachii) 10 Tulang Telapak Kaki Depan (Skeleton manus) 10 Tulang Gelang Panggul (Cingulum membri pelvini) 12
Tulang Paha (Skeleton femoris) 13
Tulang Kaki Belakang Bawah (Skeleton cruris) 13 Tulang Telapak Kaki Belakang (Skeleton pedis) 13
Pembahasan 16
SIMPULAN DAN SARAN 19
Simpulan 19
Saran 19
DAFTAR TABEL
1 Perbandingan skelet kaki hewan 7
DAFTAR GAMBAR
1 Morfologi eksternal tubuh buaya senyulong 2
2 Skelet kepala buaya senyulong: (A) Tampak lateral (B) Tampak dorsal 3
3 Distribusi buaya senyulong 4
4 Lokomosi darat buaya (a) belly crawl, (b) belly run 5 5 Gerakan high walk (a) A. mississippiensis tampak lateral (b) struktur
sendi pergelangan kaki buaya dan kadal
5 6 Tahap-tahap gerakan gallop (a) ilustrasi, (b) C. johnstoni 6 7 Gambar 7 Lokomosi buaya di perairan (a) berenang (b) mengapung 6 8 Struktur os scapula, os coracoidea dan os humerus buaya senyulong
bagian kiri
11 9 Struktur ossa radius et ulna, ossa carpi, ossa metacarpalia, dan
ossa phalanges manus buaya senyulong bagian kiri
12 10 Struktur os coxae buaya senyulong bagian kiri 14 11 Struktur os femoris, ossa tibia et fibula, ossa tarsi, ossa metatarsalia,
dan ossa phalanges pedis buaya senyulong bagian kiri
1
PENDAHULUAN
Latar Belakang
Indonesia adalah negara dengan keanekaragaman hayati tertinggi kedua di dunia setelah Brasil dan tertinggi kedua keberadaan hewan endemik setelah Australia (Brown 1997). Flora dan fauna Indonesia adalah campuran dari spesies di Benua Asia dan Australia. Pulau Sumatera, Jawa, Kalimantan dan Bali adalah pecahan dari Benua Asia dan memiliki banyak spesies Asia. Pulau Sulawesi, Kepulauan Nusa Tenggara, dan Maluku memiliki flora dan fauna yang unik karena telah lama terpisah dari Benua Asia dan Australia (Monk et al. 2013; Whitten et al. 2013).
Buaya senyulong atau sepit (Tomistoma schlegelii) adalah salah satu hewan endemik di Indonesia yang dapat ditemukan di Pulau Jawa, Kalimantan, dan Sumatera (Bezuijen et al. 2014). Berdasarkan morfologinya buaya ini memiliki kemiripan dengan famili buaya sejati (Crocodylidae) tetapi dengan ukuran tubuh yang lebih kecil (panjang rata-rata 3.5 m) serta bentuk moncong yang lancip dan runcing (Piras et al. 2010). Namun, secara molekuler buaya senyulong lebih memiliki kemiripan dengan famili Gavialidae dengan adanya gen mecistops dan osteolaemus (Willis et al. 2007).
Buaya adalah hewan semi akuatik yang menghabiskan sebagian siklus hidupnya di air. Lokomosi di air dilakukan dengan menggunakan ekor untuk berenang sementara kaki didekatkan ke tubuh untuk mengurangi gesekan dengan air (Fish 1984). Buaya mempunyai tiga cara untuk bergerak di darat, yaitu belly crawl, high walk, dan gallop. Belly crawl adalah gerakan seperti kadal yaitu gerakan dengan merentangkan kaki ke samping dan dengan pusat gravitasi rendah. Gerakan high walk berbeda dengan gerakan reptil lain yaitu gerakan dengan mengangkat tubuhnya untuk berjalan seperti mamalia. Terakhir gallop adalah gerakan seperti „melompat‟ pada kelinci yang biasa dilakukan oleh buaya untuk melarikan diri (Reilly dan Elias 1998).
Kemampuan gerakan-gerakan yang dilakukan buaya sangat berkaitan dengan anatomi skelet apendikular buaya. Sejauh ini belum dilakukan penelitian mengenai anatomi skelet apendikular buaya senyulong. Oleh karena itu, studi komparatif tentang anatomi skelet apendikular buaya senyulong penting dilakukan.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari karakteristik anatomi skelet apendikular buaya senyulong sebagai sistem lokomosi yang dikaitkan dengan fungsi dan perilaku gerak buaya senyulong.
Manfaat Penelitian
Penelitian ini bermanfaat sebagai sumber informasi anatomi skelet apendikular buaya senyulong untuk mempelajari fisiologi, adaptasi, dan perilaku buaya senyulong.
2
TINJAUAN PUSTAKA
Ordo CrocodyliaOrdo Crocodilia terdiri atas tiga famili yaitu Crocodylidae, Alligatoridae, dan Gavialidae (Halliday dan Adler 2002). Famili Gavialidae memiliki status kepunahan tertinggi di antara famili lain dari ordo Crocodilia. Gavialidae secara umum hanya memiliki satu spesies yang belum punah yaitu buaya gharial (Gavialis gangenticus), buaya lokal di India dan Nepal. Buaya senyulong biasanya dimasukkan dalam famili Crocodylidae berdasarkan morfologi skeletnya, tapi berdasarkan morfologi dan habituasi dimasukkan ke dalam famili Gavialidae. Namun, banyak studi molekuler secara konsisten memasukkan kedua spesies dalam famili Gavialidae (Willis et al. 2007).
Ciri Morfologi dan Struktur Tubuh
Ordo Crocodilia secara morfologis memiliki ciri-ciri yang sama, dengan beberapa perbedaan pada besar tubuh, bentuk moncong, dan warna kulit. Bentuk moncong pada famili Crocodylidae dapat berbentuk lebar maupun ramping dan jika dilihat dari dorsal memiliki bentuk seperti huruf „V‟, sedangkan pada famili Alligatoridae bentuk moncong lebih luas dan pendek seperti huruf „U‟. Buaya senyulong dan buaya gharial yang termaksud dalam famili Gavialidae memiliki moncong yang panjang dan ramping.
Moncong buaya gharial yang panjang dan sempit merupakan adaptasi hewan terhadap diet memakan ikan. Permukaan dorsal buaya gharial berwarna hijau gelap, hampir hitam pada saat dewasa dan pada bagian ventral berwarna putih kekuningan (Saikia 2013). Buaya jantan memiliki tonjolan hidung berangga di ujung moncong (Grigg dan Kirshner 2015). Tonjolan ini didiuga sebagai sex indikator secara visual dan sebagai resonator suara serta perilaku sexual lainnya (Martin dan Bellairs 1977).
Buaya senyulong memiliki moncong yang mirip dengan buaya gharial namun tidak ditemukan tonjolan pada ujung moncong. Moncong buaya senyulong sangat panjang dan ramping dengan perbandingan panjang moncong tiga kali lebar moncong itu sendiri (Grigg dan Kirshner 2015). Selain itu buaya senyulong juga memiliki kulit dengan warna cokelat kemerahan pada bagian dorsal beserta spot-spot hitam pada ekor. Bagian ventral berwarna putih keabuan dengan belang hitam. Buaya muda mempunyai belang pada rahang, badan, dan ekor (Gambar 1).
Gambar 1 Morfologi eksternal tubuh buaya senyulong (sumber: http://animaldiversity.org)
3 Buaya senyulong memiliki moncong yang paling ramping di antara buaya-buaya yang lain, dalam segi ukuran dapat dibandingkan dengan buaya-buaya moncong-kecil (Mecistops cataphractus) dan buaya air tawar (Crocodylus johnsoni
)
(Piras et al. 2010). Buaya senyulong adalah buaya besar, dengan ukuran tubuh yang sedikit lebih kecil dari buaya gharial. Mathew et al. (2011) menyatakan bahwa panjang dari tiga buaya jantan yang dibiakkan di Zoo Negara (Malaysia) mencapai 3,6–3,9 m dengan bobot badan 190–210 kg sedangkan betina mempunyai panjang 3,27 m dengan bobot badan 93 kg. Hewan ini juga dianggap memiliki skelet kepala terbesar dari buaya lain, karena adanya moncong yang ramping dan panjang. Skelet tengkorak kepala atas tercatat dapat mencapai 84 cm dan os mandibular mencapai 104 cm (Gambar 2) (Whitaker dan Whitaker 2008).Gambar 2 Skelet kepala buaya senyulong: (A) Tampak lateral (B) Tampak dorsal (sumber: http://digimorph.org)
Buaya memiliki salah satu bite force terkuat di antara hewan lain. Penelitian Erickson et al. (2012) menyatakan bahwa seekor buaya air laut memiliki bite force sebesar 16 000 N. Penelitian ini juga melihat hubungan antara bite force dengan bentuk moncong. Oleh karena itu, buaya gharial dengan moncong yang ramping dan panjang memiliki bite force relatif lebih lemah dari jenis buaya lain.
Buaya memiliki beragam bentuk gigi yang tumpul sampai yang berbentuk seperti jarum (Erickson et al. 2012). Spesies dengan moncong lebar memiliki ukuran gigi yang beragam, sedangkan moncong yang lebih ramping seperti buaya gharial memiliki bentuk yang seragam. Gigi buaya sejati dan buaya gharial lebih tampak daripada aligator dan kaiman saat rahang tertutup (Grigg dan Gans 1993). Bangsa buaya adalah polyphyodonts yang memiliki 80 gigi dan dalam satu masa hidupnya dapat mengganti gigi sebanyak 50 kali.
Buaya adalah hewan tetrapoda dengan empat kaki. Menurut Kelly (2006) kaki depan buaya memiliki lima digit tanpa selaput dan kaki belakang mempunyai empat digit berselaput dengan digit kelima rudimenter. Sistem skelet buaya mirip dengan tetrapoda lain, namun memiliki perbedaan pada tengkorak, pelvis dan rusuk. Tulang rawan pada rusuk dapat membuat thorax kolaps saat menyelam. Hal ini menyebabkan udara lebih banyak masuk ke dalam paru-paru. Struktur pelvis juga dapat mengakomodasi sebagian besar makanan (Huchzermeyer 2003).
4
Habitat dan Perilaku Alamiah
Buaya adalah reptil semi akuatik yang menghabiskan sebagian siklus hidupnya di air. Kebanyakan buaya hidup pada dataran rendah dan sebagian dapat ditemukan pada ketinggian 1 000 m, dengan suhu 5°C lebih rendah dari suhu pesisir. Buaya air laut memiliki distribusi wilayah terluas di antara jenis buaya lain, mulai dari India bagian timur sampai Papua Nugini dan Australia bagian utara (Alcala dan Dy-Liacco 1989).
Buaya senyulong adalah spesies buaya air tawar dan jenis buaya yang membangun sarang. Buaya ini sering berada pada daerah rawa-rawa, danau, dan sungai dataran rendah. Hewan ini dapat ditemukan di timur Sumatera, Kalimantan, dan Jawa Barat (Indonesia), serta Serawak dan Pesisir Malaysia (Malaysia). Belum lama ini buaya senyulong juga dikabarkan terlihat di Brunei Darussalam dan Thailand. Namun, populasi tertinggi berada di Kalimantan dan Sumatera (Gambar 3) (Stuebing et al. 2006).
Gambar 3 Distribusi buaya senyulong (Bezuijen et. al. 2010)
Buaya dapat hidup dalam berbagai tipe habitat perairan tergantung kepada perilaku makannya. Beberapa spesies seperti kaiman bersifat terestrial dan lebih memilih rawa-rawa, telaga, dan danau sedangkan spesies lain seperti aligator cina dapat menghabiskan lebih banyak waktu di dalam air dan mendiami sungai, hutan bakau, dan muara. Bangsa gharial memiliki habitat di sekitar kolam dan sungai dengan arus yang cepat (Alcala dan Dy-Liacco 1989). Buaya senyulong juga memiliki jenis makanan yang sama dengan buaya gharial, yaitu ikan dan vertebrata kecil. Namun beberapa observasi menunjukkan bahwa buaya senyulong dewasa juga memakan vertebrata besar termasuk bekantan, monyet ekor panjang, rusa, unggas air, dan reptil (Rachmawan dan Brend 2009).
Walaupun buaya lebih sering hidup di perairan, hewan ini juga dapat hidup di daratan untuk mengatur suhu tubuhnya karena buaya adalah hewan poikiloterm. Hewan poikiloterm atau berdarah dingin adalah hewan yang suhu tubuhnya terpengaruh oleh suhu lingkungan. Beberapa cara untuk mengatur suhu tubuh adalah dengan berjemur, membuka mulut lebar-lebar, dan mandi lumpur. Hal ini dilakukan agar tubuh dapat melakukan metabolisme secara optimal (Davenport 2012).
5 Lokomosi
Lokomosi adalah suatu perilaku hewan memindahkan dirinya dari satu tempat ke tempat lain untuk memenuhi suatu kebutuhan seperti termoregulasi, menemukan makanan, interaksi sosial, bersarang, dan berlari dari bahaya. Lokomosi di darat dan air memiliki cara yang berbeda tergantung fungsinya. Lokomosi melibatkan kaki depan dan belakang (apendikular) serta sumbu tubuh dan ekor untuk berenang (aksial) (Biewener 2003).
Menurut Britton (2002), buaya memiliki gerakan yang berbeda di darat dan di perairan. Gerakan belly crawl atau merayap merupakan gerakan yang umum terlihat di darat. Umumnya dada, perut, dan ekor menyentuh permukaan dan kaki diangkat sedikit dari permukaan untuk memberi ruang yang cukup pada saat mendorong tubuh ke depan. Buaya dapat memodifikasi gerakan ini saat menemukan permukaan kasar dengan mengangkat tubuh secara vertikal kemudian mendorongnya ke depan. Gerakan ini juga dapat dimodifikasi agar lebih cepat dengan menggerakkan tubuh seperti huruf „S‟ secara horizontal seperti kadal untuk menambahkan kekuatan dorongan pada kaki. Gerakan ini sering disebut gerakan belly run (Gambar 4).
Gambar 4 Lokomosi darat buaya (a) belly crawl, (b) belly run (Britton 2002) High walk adalah gerakan saat buaya mengangkat seluruh tubuhnya dari permukaan tanah dan kakinya diposisikan di ventral tubuh. Gerakan ini digunakan saat keluar dari air dan melewati hambatan. Buaya dapat melakukan gerakan high walk karena sendi pergelangan kaki buaya berada di antara os astragalus dan os calcaneus, sehingga buaya dapat memutar pergelangan kaki dan menopang tubuh (Parrish 1987). Namun, buaya gharial dewasa tidak dapat melakukan high walk karena kaki buaya gharial tidak cukup kuat untuk menopang tubuhnya. Hal ini menyebabkan hewan ini lebih banyak menghabiskan siklus hidup di dalam air (Gambar 5) (Whitaker dan Basu 1982).
Gambar 5 Gerakan high walk (a) A. mississippiensis tampak lateral (b) struktur sendi pergelangan kaki buaya dan kadal (Parrish 1987; Britton 2002).
Gallop adalah gerakan tercepat buaya di darat yang digunakan pada saat terancam, walaupun hanya sebagian kecil spesies buaya yang dapat melakukannya. Gallop pada buaya lebih menyerupai lompatan pada kelinci dibandingkan gallop pada kuda. Kaki belakang mendorong tubuh ke atas dan ke depan saat itu pula kaki depan diangkat dari tanah dan buaya akhirnya „melayang‟ (Tahap 1 dan 2).
a
b
a b
6
Saat mendarat, kaki depan menopang berat tubuh dan kaki belakang diayunkan ke depan sampai melewati kaki depan (Tahap 3). Saat kaki depan diangkat kembali gaya dorongan dari kaki belakang akan lebih maksimum mendorong tubuh (Tahap 4) (Gambar 6) (Webb dan Gans 1982)
Gambar 6 Tahap-tahap gerakan gallop (a) ilustrasi, (b) C. johnstoni (Britton 2002)
Berenang adalah gerakan yang dilakukan buaya saat berada di perairan. Saat dalam keadaan istirahat seluruh permukaan tubuh hewan dapat terlihat dengan ekor tidak bergerak (air yang tenang) atau dengan gerakan konstan (air mengalir). Gerakan berenang didominasi dengan pergerakan ekor dan sedikit gerakan kaki untuk mengatur pergerakan. Namun pada saat berburu, tubuh berada dalam air secara vertikal sedangkan kepala berada di atas permukaan air. Dalam keadaan ini kaki berfungsi untuk menstabilkan tubuh. Posisi vertikal ini memungkinkan hewan untuk segera menyelam saat berburu (Gambar 7) (Fish 1984).
Gambar 7 Lokomosi buaya di perairan (a) berenang (b) mengapung
Buaya memiliki dua cara bergerak di bawah permukaan air. Gerakan pertama dilakukan pada air dengan arus lambat dengan meggerakan kaki dan gerakan ekor menjadi kurang penting. Secara sederhana buaya berjalan di dasar sungai seperti gerakan di darat. Namun pada beberapa situasi kaki depan didekatkan ke samping tubuh dan kaki belakang sebagai pendorong. Gerakan yang lain adalah ekor sebagai penggerak utama dengan kaki didekatkan pada sisi tubuh untuk mengurangi gesekan dengan arus sungai. Gerakan ini dilakukan saat berada pada air dengan arus yang deras (Fish 1984).
a b a b 1 2 3 4
7 Skelet Apendikular Reptil
Struktur skelet dan otot kaki hewan beradaptasi dengan perilaku, fungsi, dan kebiasaan hewan tersebut. Hewan yang berlari dengan kecepatan tinggi akan memiliki kaki yang panjang serta cara menumpu dengan permukaan dan langkah yang panjang (Hildebrand 1960). Reptil memiliki struktur kaki yang cukup unik dengan konformasi skelet yang kompak. Struktur ini terdiri dari banyak tulang kecil-kecil di daerah carpus, tarsus, metacarpus dan metatarsus. Susunan tulang ini mendukung aktivitas reptil dalam kehidupan dan perilaku alamiahnya.
Tabel 1 Perbandingan skelet kaki hewan
Hewan Lengan atas Lengan bawah
dan digit
Fungsi
Katak 1 15 Lokomosi darat
Paus 1 31 Berenang
Anjing 1 23 Lokomosi darat
Penguin 1 9 Berenang
Manusia 1 28 Menggenggam
Kelelawar 1 26 Terbang
Unggas 1 9 Terbang
Aligator 1 28 Lokomosi darat
Sumber: (Holt et al. 2000)
Reptil memiliki os scapula dan os coracoidea yang terpisah. Menurut Getty (1975), os scapula memiliki bentuk yang telah beradaptasi sesuai dengan permukaan dinding dada. Selain itu menurut Purwanto (2014), biawak air memiliki os scapula berbentuk konveks dengan struktur yang unik dan berfungsi mendukung pergerakan serta menopang tubuh bagian depan. Os scapula dan os coracoidea akan membentuk cavitas glenoidalis yang akan berhubungan dengan facies articularis humeri. Cavitas glenoidalis pada komodo memiliki bidang sendi 130º(Surahya 1989).
Facies articularis humeri berbentuk elips dengan sudut bidang sendi transversal 220º dan horizontal 110ºdan menyebabkan os humerus dapat bergerak ke kranial dan kaudal (Surahya 1989). Reptil primitif memiliki corpus humerus, os radius dan os ulna berbentuk silindris. Biawak air hanya memiliki satu crista pectoralis pada bagian proksimal os humerus (Purwanto 2014), sedangkan pada komodo terdapat dua crista pectoralis dan satu crista deltoidea (Surahya 1989).
Os coxae pada biawak air terdiri atas os ilium, os pubis, dan os ischii. Facies articularis ossis ilii komodo memanjang sampai ke kaudal dari os ilium dan berbentuk lekukan yang relatif dalam. Hal ini menyebabkan os sacrum sama sekali tidak dapat bergerak. Struktur acetabulum biawak air yang khas, berbentuk seperti mangkuk memungkinkan terjadinya gerakan rotasi meskipun terbatas (Purwanto 2014).
Reptil primitif memiliki os femoris dan os tibia yang membesar pada bagian proksimal dan distal (Colbert 1955). Schaeffer (1941) menjelaskan bahwa os tibia memiliki facies articularis proximalis tibiae di bagian proksimal dan facies articularis distal tibiae pada ujung distal. Os fibula memiliki facies articularis distalis fibulae pada ujung distal yang bersendi dengan os astragalocalcaneus.
8
Persendian yang dibentuk antar tulang tersebut memungkinkan terjadinya gerakan ekstensor dan fleksor yang luas pada biawak air (Purwanto 2014).
Menurut Colville dan Bassert (2002), meskipun terdapat variasi jumlah ossa carpi antar spesies, pada umumnya hewan tetap memiliki os carpi radiale, os carpi ulnare, dan beberapa spesies selalu memiliki os carpi intermedium. Watson (1917) juga mengatakan ossa carpi berjumlah sepuluh buah tulang yang terdiri dari satu os carpi radiale, os carpi intermedium, os carpi ulnare, os carpi accesorium, dua ossa carpi centrale, dan empat ossa carpale distal. Colville dan Bassert (2002) menyatakan bahwa ossa tarsi yang memiliki ukuran terbesar adalah os talus (os tarsi tibiale) dan os calcaneus (os tarsi fibulare). Buaya memiliki persendian pergelangan kaki di antara os astragalus dan os calcaneus yang menyebabkan kaki dapat diputar 90º(Parrish 1987).
Buaya memiliki lima buah digit pada kaki depan dan empat buah digit pada kaki belakang (Kelly 2006). Digit I memiliki dua os phalanx sedangkan yang lain memiliki tiga os phalanx (Palastanga et al. 2013). Perbedaan struktur digit kaki depan dan kaki belakang adalah ukurannya. Kaki belakang memiliki digit yang lebih panjang dibandingkan kaki depan. Secara keseluruhan struktur digit kaki depan dan belakang reptil ini memiliki struktur yang khusus dan tersusun secara kompak serta memiliki bidang persendian yang lebih luas. Hal ini mendukung aktivitas reptil ini dalam berlari, berdiri, berenang, dan bertahan hidup di lingkungannya (Purwanto 2014). Os phalanx distalis bagian distal terdapat cakar yang relatif panjang, kokoh, dan tumpul. Cakar ini digunakan untuk membantu mendorong tubuh untuk bergerak (Britton 2002).
BAHAN DAN METODE
Waktu dan Tempat PenelitianPenelitian ini dilaksanakan pada bulan Juli sampai dengan Oktober 2016, bertempat di Laboratorium Anatomi, Divisi Anatomi Histologi dan Embriologi, Departemen Anatomi Fisiologi dan Farmakologi, Fakultas Kedokteran Hewan Institut Pertanian Bogor.
Bahan dan Alat
Bahan yang digunakan pada penelitian ini adalah satu set preparat skelet apendikular buaya senyulong dengan jenis kelamin jantan. Buaya senyulong yang digunakan dalam penelitian ini diperoleh dari Taman Margasatwa Ragunan Jakarta. Pihak Taman Margasatwa Ragunan Jakarta menyerahkan satu set tulang buaya senyulong ke Laboratorium Anatomi Fakultas Kedokteran Hewan IPB.
Alat-alat yang digunakan dalam penelitian ini adalah penggaris, alat tulis, dan kamera Canon EOS® 700D.
9 Metode Penelitian
Penelitian dilakukan secara eksploratif dengan melakukan pengamatan, pencatatan, dan pengambilan gambar dari skelet apendikular buaya senyulong. Pengamatan preparat skelet dilakukan dengan mengamati bentuk dan bagian skelet yang khas, pengukuran panjang dan lebar tulang, serta membandingkan dengan skelet apendikular buaya lain. Adapun pengolahan data dilakukan secara deskriptif dengan menjelaskan bagian-bagian tulang dan perbedaannya dengan skelet buaya apendikular lain. Selanjutnya dilakukan pengambilan gambar dengan menggunakan kamera Canon EOS® 700D. Gambar yang diperoleh kemudian diolah dengan software Adobe Photoshop CS4. Kemudian dilakukan penamaan pada setiap bagian tulang berdasarkan Nomina Anatomica Veterinaria (ICVGAN 2012) dan referensi lain tentang buaya. Skelet apendikular buaya senyulong selanjutnya dirangkai menurut posisi alamiahnya.
HASIL DAN PEMBAHASAN
HasilBuaya senyulong memiliki struktur skelet kaki depan dan belakang yang kokoh dan kompak. Skelet kaki depan terdiri atas os scapula, os coracoidea, os interclavicula, os humerus, os radius, os ulna, ossa carpi, ossa metacarpalia dan ossa phalanges manus, sedangkan skelet kaki belakang terdiri atas os coxae
(os ilium, os pubis, os ischii), os femoris, os tibia, os fibula, ossa tarsi, ossa metatarsalia, dan ossa phalanges pedis.
Tulang Gelang Bahu (Cingulum membri thoracici)
Tulang gelang bahu pada buaya senyulong ini terdiri dari os scapula dan os coracoidea. Os scapula berbentuk konveks dan terletak kraniolateral thorax. Bagian dorsal tulang ini ditemukan cartilago suprascapularis yang membentuk seperempat bagian dorsal os scapula. Os coracoidea memiliki bentuk yang mirip dengan os scapula dan terletak di ventral thorax. Di proksimal tulang ini ditemukan foramen coracoidea. Pada bagian medial ditemukan cartilago
coracoidea yang bersendi dengan os interclavicula. Tulang ini berbentuk segitiga
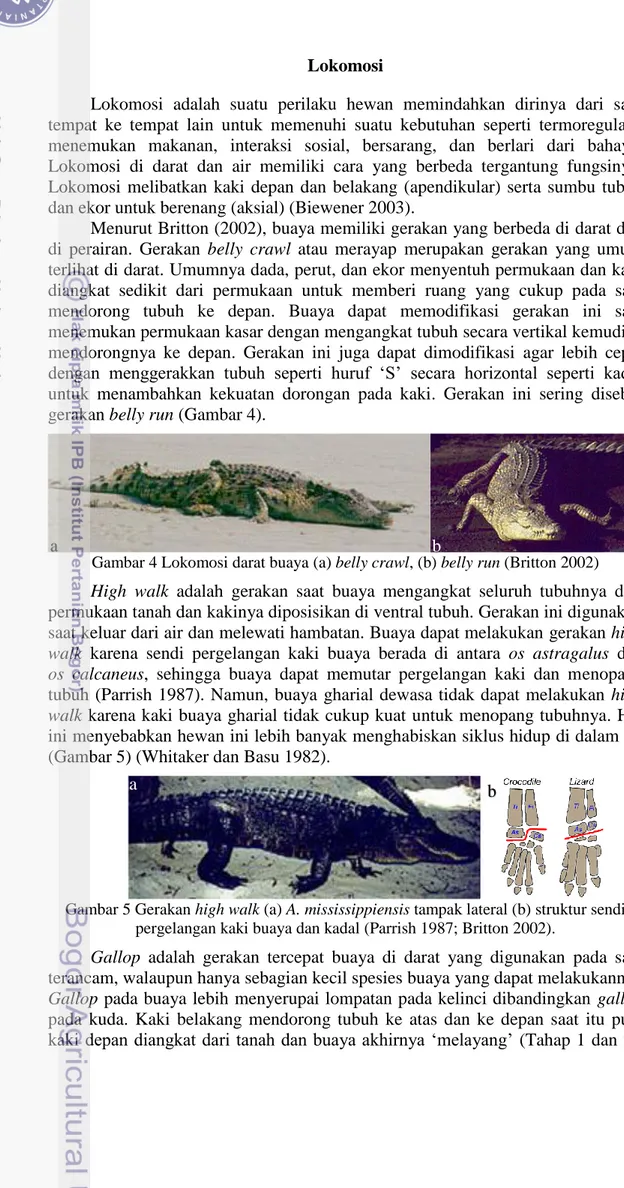
dan bersendi dengan os sternum serta menghubungkan os coracoidea kanan dan kiri. Os scapula bagian distal dan os coracoidea bagian proksimal bertaut membentuk cavitas glenoidalis yang memiliki lengkungan relatif dalam untuk bidang sendi dengan caput humeri (Gambar 8).
Tulang Lengan Atas (Skeleton brachii)
Os humerus merupakan tulang terbesar dari kaki depan, berbentuk silindris, dan pada bagian distal melengkung ke medial. Tulang ini memiliki panjang ± 20 cm dan di bagian proksimal terdapat caput humeri yang terletak di antara tuberculum majus dan tuberculum minus. Pada lateral tuberculum majus terdapat
crista humeri yang memanjang ke ventral dan bertemu dengan tuberositas deltoidea. Bidang persendian dengan tulang gelang bahu adalah caput humeri
10
yang berbentuk konveks dengan permukaan licin. Os humerus pada bagian distal terdapat dua bungkul yaitu condylus ulnaris dan condylus radialis yang memiliki permukaan halus. Condylus ulnaris terletak di lateral dan bersendi dengan os ulna sedangkan condylus radialis terletak di medial dan bersendi dengan os radius (Gambar 8).
Tulang Lengan Bawah (Skeleton antebrachii)
Ossa radius et ulna buaya senyulong terpisah oleh spatium interosseum antebrachii sepanjang kedua tulang tersebut. Os radius berbentuk silindris dengan
panjang ± 11 cm sedangkan os ulna berbentuk relatif pipih dengan panjang ± 14 cm. Os ulna pada bagian proksimal lebih besar dengan adanya olecranon yang relatif tidak berkembang sedangkan bagian distal lebih pipih dan melengkung ke medial. Bagian proksimal terdapat caput radii dan caput ulnaris yang bersendi dengan condyli radialis et ulnaris dari os humerus. Permukaan ini berbentuk cekung dengan permukaan yang licin. Bagian distal os ulna terdapat facies articularis ulnare yang berbentuk tidak beraturan dengan permukaan licin dan bersendi dengan os carpi ulnare dan os carpi accessorium, sedangkan os radius terdapat facies articularis radiale dengan bidang yang menjulur membentuk processus styloideus dan bersendi dengan os carpi radiale (Gambar 9).
Tulang Telapak Kaki Depan (Skeleton manus)
Buaya senyulong memiliki empat buah ossa carpi dengan bentuk yang tidak beraturan. Tulang-tulang ini tersusun dalam dua baris; baris proksimal terdiri atas os carpi radiale, os carpi ulnare, dan os carpi accessorium, sedangkan pada baris distal terdapat ossa carpale IV et V. Os carpi radiale memiliki ukuran terbesar
dan bersendi dengan os radius pada bagian proximocaudal. Os carpi accessorium berupa tulang kecil di kaudal os carpi ulnare, kedua tulang ini bersendi dengan
os ulna (Gambar 9).
Ossa metacarpalia berjumlah lima buah dan berbentuk silindris yang terdiri
dari ossa metacarpale I, II, III, IV et V. Bagian kranial ossa metacarpalia bersendi dengan ossa phalanges manus berbentuk konveks hampir setengah lingkaran dengan permukaan yang licin (Gambar 9).
Ossa digitorum manus terdiri dari lima digit dan setiap digit memiliki os phalanx proximalis, os phalanx media, dan os phalanx distalis. Ossa phalanges manus berbentuk silindris pendek dengan persendian antar tulang yang memungkinkan terjadinya gerakan yang luas. Digit I-V kaki depan buaya senyulong memiliki jumlah os phalanx media yang berbeda, masing-masing 0, 1, 2, 1, dan 0. Os phalanx distalis pada bagian distal terdapat cakar yang relatif panjang dan kuat pada digit I, kemudian semakin mengecil pada digit yang lebih lateral (Gambar 9).
11
Gambar 8 Struktur os scapula, os coracoidea dan os humerus buaya senyulong bagian kiri.
A. Os scapula dan os caracoidea tampak dorsal B. Os humerus tampak dorsal, lateral dan distal
1. Os scapula, 2. Os coracoidea, 3. Cartilago suprascapularis, 4. Foramen coracoidea, 5. Cavitas glenoidalis, 6. Os humerus, 7. Crista humeri,
8. Caput humeri, 9. Tuberculum majus, 10. Tuberculum minus,
11. Tuberositas deltoidea, 12. Condylus ulnaris, 13. Condylus radialis B
12
Gambar 9 Struktur ossa radius et ulna, ossa carpi, ossa metacarpalia, dan ossa phalanges manus buaya senyulong bagian kiri
A. Ossa radius et ulna, ossa carpi, ossa metacarpalia dan ossa phalanges manus tampak dorsal
B. Ossa carpi dan ossa metacarpalia tampak dorsal C. Ossa carpi tampak ventral
1. Os radius, 2. Os ulna, 3. Olecranon, 4. Spatium interosseum antebrachii, 5. Caput ulnare, 6. Caput radii, 7. Facies articularis radiale, 8. Facies
articularis ulnare, 9. Processus styloideus, 10. Os carpi radiale, 11. Os carpi ulnare, 12. Os carpi accessorium, 13. Ossa carpale IV et V,
14. Os metacarpale I, 15. Os metacarpale II, 16. Os metacarpale III,
17. Os metacarpale IV, 18. Os metacarpale V, 19. Os phalanx proximalis, 20. Os phalanx media, 21. Os phalanx distalis
Tulang Gelang Panggul (Cingulum membri pelvini)
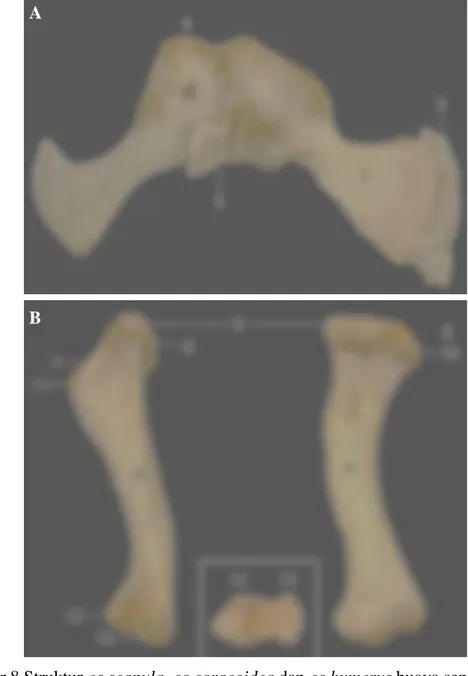
Os coxae pada senyulong dibentuk oleh tiga tulang yaitu os ilium, os ischii dan os pubis. Os coxae kiri dan kanan bertemu pada symphysis pubis dan symphysis ischiadica. Pada os coxae ini ditemukan foramen obturatum dan foramen puboischiadicus. Foramen obturatum pada buaya senyulong berukuran relatif kecil dan terletak di kranial acetabulum. Lubang ini terbentuk dari pertautan antara os ilium dan os ischii, sedangkan foramen puboischiadicus merupakan lubang yang terletak di antara os ischii dan os pubis (Gambar 10).
Os ischii merupakan tulang yang terletak di bagian kaudal dari os coxae. Tulang ini menghadap ke ruang panggul, berbentuk pipih dengan aspek permukaan licin. Os pubis berbentuk pipih seperti kipas dan menghadap ke
kranial. Tulang ini di kraniodistal berhubungan dengan ossa gastralia skelet tubuh buaya. Os ilium memiliki bentuk yang tidak beraturan dan menjulur ke kaudal. Tulang ini memiliki dua permukaan yaitu facies glutea yang menghadap ke
dorsolateral dan bagian kaudalnya membentuk ala ossis ilii, serta facies sacropelvina yang menghadap ke medial dan bertaut dengan os sacrum pada
C
13
facies articularis ossis ilii. Facies ini memiliki bentuk bergelombang dengan permukaan yang kasar dan menutupi 2/3 bagian medial os ilium. Pertemuan dari os ilium dan os ischii membentuk acetabulum, yang berbentuk cekung dengan permukaan licin dan bersendi dengan caput ossis femoris (Gambar 10).
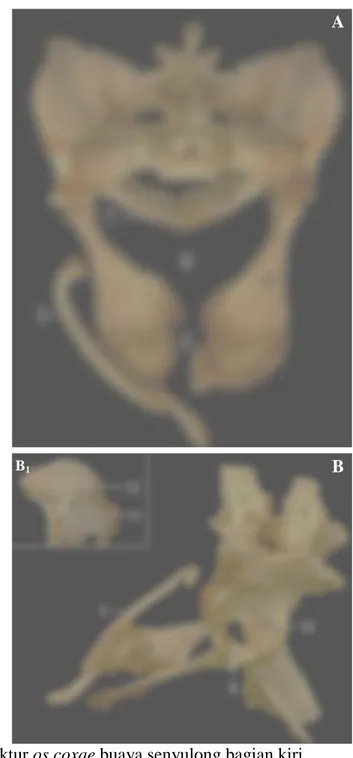
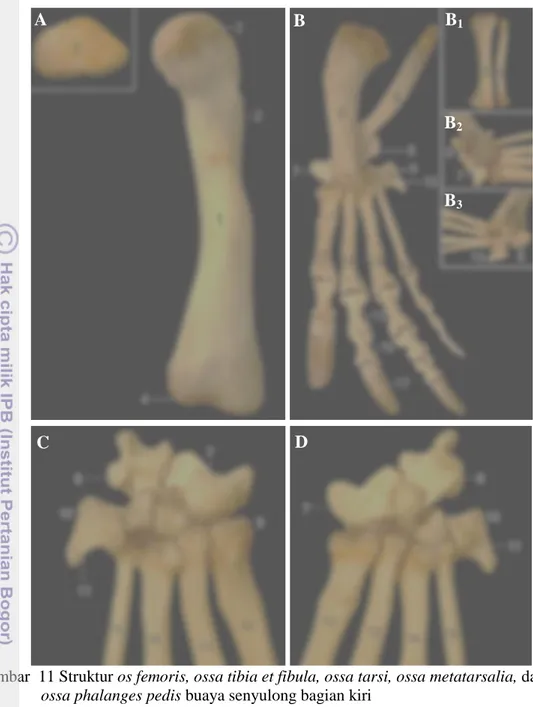
Tulang Paha (Skeleton femoris)
Os femoris merupakan tulang terpanjang dari buaya senyulong dengan panjang ± 24 cm. Bagian proksimal tulang ini terdapat caput femoris yang berhubungan langsung dengan corpus femoris tanpa adanya collum ossis femoris. Caput femoris berupa bungkul yang relatif besar, berbentuk konveks dan jajaran genjang dengan bagian kaudal mengecil dan permukaan licin. Caput ini terletak di sisi proksimomedial dan bersendi dengan acetabulum. Os femoris pada buaya
senyulong tidak ditemukan adanya trochanter mayor maupun trochanter minor tetapi memiliki trochanter tertius. Os femoris bagian distal memiliki condyli
lateralis et medialis yang berbentuk seperti angka delapan (8) dengan permukaan
licin. Trochlea ossis femoris dan os patella tidak ditemukan pada buaya senyulong (Gambar 11).
Tulang Kaki Belakang Bawah (Skeleton cruris)
Tulang kaki belakang bawah terdiri dari ossa tibia et fibula, berbentuk silindris dengan panjang masing-masing ± 14 cm dan ± 15 cm. Os tibia berukuran lebih besar dibandingkan dengan os fibula yang relatif lebih pipih. Kedua tulang ini membentuk spatium interosseum cruris di sepanjang tulang tersebut. Os tibia bagian proksimal terdapat facies articularis proximalis tibiae berbentuk datar dengan permukaan licin. Pada bagian distal terdapat facies articularis distalis tibiae yang berbentuk lekukan tidak terlalu dalam dengan permukaan licin dan bersendi dengan os astragalus secara kompak. Os fibula lebih panjang dari os tibia dan di proksimal, tulang ini bersendi dengan condylus lateralis dari os femoris, sedangkan di distal terdapat facies articularis distalis fibulae berbentuk konveks dengan permukaan licin, dan bersendi dengan os calcaneus (Gambar 11).
Tulang Telapak Kaki Belakang (Skeleton pedis)
Skeleton pedis buaya senyulong mempunyai lima buah ossa tarsi dan empat digit. Ossa tarsi tersusun menjadi baris proksimal dan distal. Baris proksimal dibentuk oleh os astragalus yang bersendi dengan os tibia dan os calcaneus yang bersendi dengan os fibula. Kedua tulang ini berbentuk persegi panjang tidak
beraturan. Baris distal terdiri atas os tarsi centrale, os tarsale III, serta
ossa tarsale IV et V yang bersatu dengan os metatarsale V. Ossa metatarsalia
berjumlah empat buah berbentuk silindris, yaitu ossa metatarsale I, II, III, et IV. Bagian distal ossa metatarsalia bersendi dengan ossa phalanges pedis (Gambar 11).
Ossa digitorum pedis terdiri dari empat digit, yaitu digit I, digit II, digit III, dan digit IV. Digit tersusun atas os phalanx proximalis, os phalanx media, dan os phalanx distalis. Ossa phalanges pedis secara umum memiliki struktur yang mirip pada kaki depan, dengan os phalanx media sama pada kaki depan, masing-masing berjumlah 0, 1, 2 dan 1 tanpa digit V. Os phalanx distalis pada bagian distal terdapat cakar yang relatif panjang dan tebal pada setiap digit (Gambar 11)
14
Gambar 10 Struktur os coxae buaya senyulong bagian kiri
A. Os coxae kiri dan kanan dan persendian sacroiliaca tampak kraniodorsal B. Os coxae tampak lateral
B1. Inset B (os ischi) tampak medial
1. Os sacrum, 2. Os ilium, 3. Os ischii, 4. Os pubis, 5. Ossa gastralia 6. Symphysis pubis, 7. Symphysis ischiadica, 8. Foramen obturatum,
9. Foramen puboischiadicus, 10. Acetabulum, 11. Facies articularis ossis
ilii, 12. Ala ossis ilii,
B1
A
15
Gambar 11 Struktur os femoris, ossa tibia et fibula, ossa tarsi, ossa metatarsalia, dan ossa phalanges pedis buaya senyulong bagian kiri
A. Os femoris tampak dorsal
B. Os tibia, os fibula, ossa tarsi, ossa metatarsalia, dan ossa phalanges pedis tampak dorsal
B1. Os tibia dan os fibula tampak dorsal
B2. Inset B tampak medial
B3. Inset B tampak lateral
C. Ossa tarsi dan ossa metatarsalia tampak dorsal D. Ossa tarsi tampak ventral
1. Caput femoris, 2. Corpus femoris, 3. Trochanter tertius, 4. Condyli
lateralis et medialis, 5. Os tibia, 6. Os fibula, 7. Os astragalus, 8. Os calcaneus, 9. Os tarsi centrale, 10. Os tarsale III, 11. Ossa tarsale IV
et V, 12. Os metatarsale I, 13. Os metatarsale II, 14. Os metatarsale III,
15. Os metatarsale IV, 16. Os phalanx proximalis, 17. Os phalanx media,
18. Os phalanx distalis
A B B1
B2
B3
16
Pembahasan
Buaya senyulong merupakan hewan tetrapoda yang memiliki empat buah kaki yang sangat kuat. Serupa dengan buaya jenis lainnya, kaki depan buaya memiliki lima digit tanpa selaput sedangkan kaki belakang mempunyai empat digit berselaput dengan digit ke lima rudimenter. Kaki depan dan belakang memiliki tulang-tulang kecil pada daerah carpus, tarsus, metacarpus, dan metatarsus. Susunan tulang ini mendukung aktivitas reptil dalam kehidupan dan perilaku alamiahnya (Grzimek 1975).
Buaya memiliki struktur tulang yang telah beradaptasi dengan fungsi dan perilakunya seperti pertulangan gelang bahu (pectoral girdle) dan pertulangan gelang panggul (pelvic girdle). Hal ini sesuai dengan Hildebrand (1960) yang menyatakan struktur skelet dan otot kaki hewan akan mengalami adaptasi sesuai dengan fungsi, perilaku, dan kebiasaan hewan tersebut. Struktur tulang gelang bahu tersusun dari os scapula, os coracoidea, dan os interclavicula. Reptil yang
merupakan hewan melata memiliki os coracoidea yang bersendi dengan
os clavicula dan os interclavicula, tetapi pada buaya senyulong hanya ditemukan os interclavicula, sedangkan os scapula tidak bersendi langsung dengan skelet
tubuh (Hyman 1992). Struktur ini diduga berfungsi untuk melindungi organ-organ di daerah rongga thorax beserta os sternum yang hanya berupa kartilago. Selain itu, bentuk dan struktur persendian gelang bahu juga diduga mendukung pergerakan hewan dalam membantu meningkatkan panjang langkah hewan tanpa memerlukan pergerakan berlebihan pada persendian bahu dan siku (Jenkins dan Goslow 1983). Struktur seperti ini juga dapat membantu efisiensi penggunaan energi metabolisme buaya yang tergolong hewan berdarah dingin. Hewan poikiloterm hanya memiliki 5-10% total energi metabolisme jika dibandingkan pada hewan homoiterm (Campbell dan Reece 2002).
Lokomosi darat buaya senyulong, memiliki beberapa gerakan seperti belly crawl, high walk, dan gallop. Umumnya dada, perut, dan ekor menyentuh permukaan tanah kemudian kaki diangkat sedikit dari permukaan cukup untuk mendorong tubuh ke depan. Gerakan ini didukung oleh struktur persendian-persendian dari buaya, antara lain persendian-persendian bahu yang merupakan pertemuan
antara cavitas glenoidalis tulang gelang bahu dengan caput humeri dari os humerus. Caput humeri terletak di antara tuberculum majus et minus sehingga
dapat melakukan gerakan ekstensor dan fleksor yang leluasa. Menurut Skerritt dan Lelland (1984) persendian ini memungkinkan gerakan ekstensor dan fleksor namun sedikit gerakan abduksi, adduksi, dan rotasi.
Pada os ulna buaya senyulong, olecranon relatif kurang berkembang jika dibandingkan dengan mamalia, seperti pada biawak air (Purwanto 2014). Kondisi ini diduga untuk mendukung fleksibilitas buaya dalam berjalan dan berenang. Pada kaki belakang buaya senyulong tidak ditemukan trochlear ossis femoris, juga tidak terdapat os patella pada persendian lutut. Pada tetrapoda primitif seperti amfibi dan reptil (kecuali Lepidosaur), tendo dari kaki belakang bagian atas langsung berhubungan dengan os tibia dan os patella tidak ditemukan (Romer dan Parson 1977). Hal ini diduga dapat membantu buaya bergerak lebih cepat terutama saat melakukan high walk. Saat bergerak cepat gerakan bagian bawah baik depan maupun belakang akan membentuk sudut sendi lutut yang lebih luas pada saat ekstensor dan fleksor (Reilly dan Elias 1998).
17 Bagian distal os humerus pada buaya senyulong lebih mengarah ke medial tubuh. Hal ini diduga untuk memudahkan kaki depan aduksi ke tubuh sehingga hambatan antara tubuh dan air menjadi lebih kecil ketika berenang. Selain itu pada persendian sacroiliaca, facies articularis ossis ilii pada os coxae langsung bertaut dengan os sacrum dan tidak memungkinkan terjadi gerakan antara kedua tulang. Hal ini mendukung fleksibilitas daerah ekor buaya dan lokomosi buaya saat berjalan dan berenang. Struktur ini dapat membantu gerakan buaya senyulong dalam lokomosi air. Menurut Whitaker dan Basu (1982) bangsa gharial menghabiskan sebagian besar siklus hidupnya di dalam air dan memiliki kemampuan berenang yang baik.
Pada pertemuan dari os ilium dan os ischii membentuk acetabulum. Struktur acetabulum buaya senyulong yang cekung serta bentuk caput femoris seperti jajaran genjang menyebabkan gerakan ke kranial dan ke kaudal lebih mudah dilakukan dibandingkan dengan gerakan ekstensor, fleksor dan rotasi. Gerakan rotasi ini pada komodo juga tidak dapat dilakukan karena bentuk acetabulum bidang vertikal lebih dalam daripada bidang horizontal, sehingga gerakan abduksi dan adduksi kaki belakang lebih luas dibandingkan gerakan ekstensor dan fleksor (Surahya 1989).
Persendian antara os astragalus (homolog os talus) dan os calcaneus yang memiliki persendian gerak menyebabkan buaya dapat memutar sumbu kaki sejajar dengan sumbu tubuh dan melakukan high walk (Parrish 1987). High walk adalah gerakan khusus buaya dengan mengangkat seluruh tubuhnya dari permukaan tanah dan kakinya diposisikan di ventral tubuh seperti pada mamalia. Gerakan ini tidak dapat dilakukan oleh biawak air karena kedua tulang ini tumbuh menyatu membentuk os astragalocalcaneus, tetapi masih terlihat batas antara kedua tulang tersebut (Purwanto 2014).
Selain persendian antara os astragalus dan os calcaneus, celah antara tulang kaki bawah (antebrachii dan cruris) juga penting dalam gerakan buaya. Spatium interosseum antebrachii antara ossa radius et ulna dan spatium interosseum cruris
antara ossa tibia et fibula terdapat di sepanjang kedua tulang tersebut. Hal ini memungkinkan terjadinya gerakan pronasio dan supinasio yang leluasa. Semakin luas celah antara kedua tulang tersebut semakin leluasa pula gerakan pronasio dan supinasio yang dapat dilakukan (Dyce et al. 2010). Gerakan ini memudahkan buaya untuk mengontrol gerakan saat berjalan, berenang, dan gerakan lain yang memerlukan fleksibilitas tinggi seperti saat menangkap mangsa. Pada karnivora celah ini juga lebih luas dibandingkan pada babi dan kuda, yang merupakan hewan herbivora (Dyce et al. 2010).
Saat bergerak, baik di darat maupun di air (berenang), skeleton pedis dan skeleton manus menumpu pada permukaan untuk mendorong tubuh buaya. Skeleton pedis buaya senyulong terdiri dari empat buah ossa carpi seperti pada aligator (Kent dan Carr 2001), tetapi berbeda pada komodo yang mempunyai sebelas buah ossa carpi (Surahya 1989). Buaya senyulong memiliki empat buah ossa carpi dengan baris proksimal terdiri dari os carpi radiale, os carpi ulnare, os carpi accessorium dan pada baris distal hanya memiliki ossa carpales IV et V. Selain itu terdapat bantalan berupa jaringan ikat di antara sendi-sendi tulang kaki. Struktur tersebut menyebabkan gerakan pergelangan kaki buaya lebih fleksibel sehingga buaya dapat memutar sumbu kaki sejajar dengan sumbu tubuh dan melakukan gerakan high walk. Tetapi menurut Whitaker dan Basu (1982) bangsa
18
Gharial tidak dapat melakukan gerakan high walk, belly run, dan gallop karena memiliki otot yang lemah. Hal ini menyebabkan ektremitas distal os radius memiliki facies articularis radiale yang luas dan menjadi bidang tumpuan pada kaki depan. Os radius yang menjadi tumpuan menyebabkan os ulna lebih aktif dalam menggerakkan skeleton pedis (Borgeskov et al. 1966).
Pada buaya senyulong, ossa metacarpalia dan ossa metatarsalia memiliki berbentuk silindris. Pada proksimal tulang-tulang ini saling menumpu karena jumlah ossa carpi dan ossa tarsi yang sedikit sedangkan pada bagian distal
bersendi dengan ossa phalanges manus dan pedis. Permukaan sendi
ossa metacarpalia dan ossa metatarsalia berbentuk konveks dengan permukaan
licin, yang memungkinkan gerakan ektensor dan fleksor yang relatif luas pada daerah digit (Purwanto 2014). Menurut Kent dan Carr (2001) hewan yang memiliki lima digit biasanya merupakan hewan plantigradi yang berjalan dengan digit dan metacarpale/metatarsale yang menyentuh bidang tumpuan. Buaya senyulong memiliki lima digit pada kaki depan dan empat digit pada kaki belakang, dengan setiap digit terdiri atas os phalanx proximalis, os phalanx media, dan os phalanx distalis. Digit I memiliki dua ossa phalanges sedangkan digit yang lain memiliki tiga ossa phalanges (Palastanga et al. 2013) namun pada digit III buaya senyulong memiliki dua buah ossa phalanges media.
Struktur digit kaki depan dan kaki belakang berbeda dalam ukuran dan jumlah digit. Kaki belakang memiliki digit yang lebih panjang dibandingkan kaki depan serta satu digit rudimenter karena memerlukan permukaan yang luas sebagai tumpuan untuk mendorong tubuh. Secara keseluruhan struktur digit kaki depan dan belakang reptil ini memiliki struktur yang khusus dan persendian antar tulang yang renggang. Kondisi ini diduga terkait dengan perilaku berenang buaya senyulong yang memerlukan struktur jari yang relatif panjang dan fleksibel sehingga dapat bergerak dengan lebih leluasa. Pada os phalanx distalis buaya terdapat cakar yang relatif panjang, kokoh dan melengkung. Cakar ini dapat digunakan untuk mendorong tubuh dan menggali pasir saat akan bertelur.
Struktur skelet apendikular buaya selain dalam bergerak, juga disesuaikan dengan perilaku buaya lainnya seperti melindungi organ, membantu respirasi dan reproduksi. Os coracoidea buaya senyulong diduga berfungsi dalam melindungi
organ-organ dalam rongga thorax sama halnya dengan os coxae. Os coxae terdiri dari os pubis, os ischi, dan os ilium diduga berfungsi untuk melindungi organ genitalia buaya. Os pubis pada kraniodistal bersendi dengan ossa gastralia yang diduga untuk menahan organ viscera buaya agar tetap berada di abdominal, selain itu menurut Claessens (2004) dengan adanya persambungan ini os pubis dapat digerakkan untuk membantu dalam respirasi buaya. Saat melakuan inspirasi os pubis akan berotasi ke arah vental dan menyebabkan permukaan rongga dada meluas serta tekanan yang menurun, sedangkan pada saat ekspirasi os pubis berotasi ke arah dorsal dan menyebabkan permukaan rongga dada meyempit serta tekanan yang meningkat.
Pada os coxae juga ditemukan foramen obturatum dan foramen puboischiadicus. Foramen obturatum pada buaya senyulong berukuran relatif kecil dan terletak di antara pertemuan os ischi dan os ilium. Foramen puboischiadicus bersama symphysis pubis dan symphysis ischiadica, yang mempertemukan os coxae kiri dan kanan, akan melebar saat hewan bertelur (Romer dan Parsons 1977).
19
SIMPULAN DAN SARAN
SimpulanBuaya senyulong memiliki struktur skelet kaki depan dan belakang yang kokoh. Pada kaki depan, os scapula dan os coracoidea bertaut dan membentuk cavitas glenoidalis, os humerus lebih melengkung ke medial, olecranon pada os ulna tidak berkembang, dan di ekstremitas distal os radius terdapat facies articularis radiale yang luas untuk bersendi dengan os carpi radiale. Pada kaki belakang, os coxae bertaut dengan os sacrum, os pubis berhubungan dengan ossa gastralia, di distal os femoris tidak ditemukan os patella, di daerah tarsi, os astragalus dan os calcaneus membentuk sendi gerak. Skeleton manus dan pedis memiliki struktur yang mirip, tetapi digit V pada kaki belakang rudimenter. Struktur skelet apendikular tersebut memungkinkan buaya untuk melakukan gerakan berjalan, berenang, serta melindungi organ dan membantu respirasi.
Saran
Penelitian lanjutan mengenai struktur anatomi fungsional pada buaya senyulong masih perlu dilakukan terutama pada bagian struktur otot. Hal ini bertujuan untuk mendapatkan data yang lebih lengkap mengenai anatomi fungsional buaya senyulong.
DAFTAR PUSTAKA
Alcala AC, Dy-Liacco MTS. 1989. Habitats. Di dalam: Crocodiles and Alligators: Facts on File. Editor: Ross CA, Garnett S. New York (US): New York, Inc. hlm. 136–153
Brown LR. 1997. State of the World 1997: A Worldwatch Institute Report on Progress Toward a Sustainable Society 14th Ed. New York (US): Norton & Co. hlm. 7
Bezuijen MR, Shwedick B, Simpson BK, Staniewicz A, Stuebing R. 2014. Tomistoma schlegelii. The IUCN Red List of Threatened Species 2014. [internet]. [diunduh 2016 April 16] tersedia pada: http://dx.doi.org/10.2305/IUCN.UK.2014-1.RLTS.T21981A2780499.en
Biewener AA. 2003. Animal locomotion. London (UK) Oxford University Press. hlm. 1–14
Britton A. 2002. Locomotion. Di dalam: Crocodile General Biology. [internet]. [diunduh 2016 Juli 21] tersedia pada: http://crocodilian.com/cnhc/cbd-gb3.htm Borgeskov S, Christiansen B, Kjaer A, Balslev I. 1966. Fractures of the carpal
20
Campbell NA, Reece JB. 2002. Biology. 6th Ed. International Edition. San Fransisco, Boston, New York (US): Pearson Education, Inc.
Claessens LP. 2004. Archosaurian respiration and the pelvic girdle aspiration breathing of crocodyliforms. Proceedings of the Royal Society of London B: Biological Sciences, 271(1547):1461-1465.
Colbert EH. 1955. Evolution of the Vertebrates. New York (US): John Wiley and Sons, Inc. 223.
Colville T, Bassert JM. 2002. Clinical Anatomy & Physiology for Veterinary Technicians. Philadelphia (US): Mosby. 95-118
Davenport J. 2012. Animal Life at Low Temperature. London (UK): Chapman & Hall. Hlm 51–87
Dyce KM, Sack WO, Wensing CJG. 2010. Textbook of Veterinary Anatomy. 4th Ed. Philadelphia (US): WB Saunders.
Erickson GM, Gignac PM, Steppan SJ, Lappin AK, Vliet KA, Brueggen JD, Webb GJW. 2012. Insights into the ecology and evolutionary success of crocodilians revealed through bite-force and tooth-pressure experimentation. PLoS ONE, 7(3).
Fish FE. 1984. Kinematics of undulatory swimming in the American alligator. Copeia. 84(4):839–843.
Getty R. 1975. Osteology. Di dalam: Sisson and Grossman: The Anatomy of the Domestic Animal. 5th Ed. Editor: Getty, R. Philadelphia (US): WB Saunders. Hlm 19–34
Grigg G, Gans C. 1993. Morphology and physiology of the crocodylia. Di dalam: Fauna of Australia. Volume 2A, Amphibia and Reptilia. Editor: Glasby CJ, Ross GJB, Beesley PL. Melbourne (AU): Australian Government Publishing Service. 326–336
Grigg G, Kirshner D. 2015. Biology and evolution of crocodylians. Ithaca and London (UK): CSIRO PUBLISHING. Hlm 81–132
Grzimek B. 1975. Grzimek’s Animal Life Encyclopedia. London (UK): Van Nostrand Reinhold Company. 321–337.
Hildebrand M. 1960. How Animal Run. Dalam: Hildebrand M, Goslow GEJ. 2001. Analysis of Vetebrate Structure. 5th Ed. New York (US): John Willey and Sons. 31–37
Huchzermeyer FW. 2003. Crocodiles: Biology, Husbandry and Disease. South Africa (ZA): CABI Publishing. Hlm 7–10
Halliday T, Adler K. 2002. The Firefly Encyclopedia of Reptiles and Amphibians. Toronto (CA): Firefly Books. Hlm 212–221
Hyman LH. 1992. Hyman's comparative vertebrate anatomy. Chicago (US): University of Chicago Press. Hlm 238–264
[ICVGAN] International Committee on Veterinary Gross Anatomical Nomenclature. 2012. Nomina Anatomica Veterinaria. 5th Ed. Hannover (DE): Editorial Commitee of WAVA.
Jenkins FA, Goslow GE. 1983. The functional anatomy of the shoulder of the savannah monitor lizard (Varanus exanthematicus). Journal of Morphology. 175(2):195-216.
Kelly L. 2006. Crocodile: Evolution's Greatest Survivor. Crows Nest (AU): Allen & Unwin. 67–92
21 Kent GC, Carr RK. 2001. Comparative Anatomy of the Vertebrates. 9th Ed. New
York (US): McGraw-Hill Book, Co. 505 hlm.
Martin BGH; Bellairs A. 1977. The narial excrescence and pterygoid bulla of the gharial, Gavialis gangeticus (Crocodilia). Journal of Zoology. 182(4):541–558. Mathew A, Ganesan M, Majid RA, Beastall C. 2011. Breeding of False Gharial (Tomistoma schlegelii) at Zoo Negara, Malaysia. Zoo Negara. [internet] .
[diunduh 2016 April 16] tersedia pada:
http://zoonegaramalaysia.my/RPFalseGharial.pdf.
Monk K, De Fretes Y, Reksodiharjo-Lilley G. 2013. Ecology of Nusa Tenggara and Maluku. Jakarta (ID): Tuttle Publishing. Hlm 9–57
Palastanga N, Field D, Soames R. 2013. Anatomy and Human Movement. 2nd Ed. Melbourne (AU): Butterworth Heinemann. Hlm 320–325
Parrish JM. 1987. The origin of crocodilian locomotion. Paleobiology, 13(4):396-414.
Piras P, Colangelo P, Adams DC, Buscalioni A, Cubo J, Kotsakis T, Raia P. 2010. The Gavialis–Tomistoma debate: the contribution of skull ontogenetic allometry and growth trajectories to the study of crocodylian relationships. Evolution & Development. 12(6):568-579.
Purwanto E. 2014. Karakteristik struktur skelet kaki biawak air (Varanus salvator). [skripsi] Bogor (ID): Institut Pertanian Bogor.
Rachmawan D, Brend S. 2009. Human-Tomistoma interactions in Central Kalimantan, Indonesian Borneo. Crocodile Specialist Group Newsletter. 28:9-11.
Reilly SM, Elias JA. 1998. Locomotion in Alligator mississippiensis: kinematic effects of speed and posture and their relevance to the sprawling-to-erect paradigm. Journal of Experimental Biology. 201(18):2559-2574.
Romer AS, Parsons TS. 1977. The Vertebrate Body. Philadelphia (US): Holt-Saunders International. 590.
Saikia PK. 2013. Indian Gharial (Gavialis gangeticus): Status, Ecology and Conservation. Rare Animals of India. 1:76.
Skerritt GC, Lelland JM. 1984. An Introduction to the Functional Anatomy of the Limbs of the Domestic Animals. England (UK): John Wright & Sons.
Surahya S. 1989. Komodo: Studi Anatomi dan Kedudukannya dalam Sistematika Hewan. Yogyakarta (ID). Gadjah Mada University Press.
Schaeffer B. 1941. The morphological and functional evolution of the tarsus in amphibians and reptiles.
American Museum of Natural History. 78:395–472.
Stuebing RB. 2006. The current and historic distribution of Tomistoma schlegelii(the False Gharial) (Müller 1838) (Crocodylia, Reptilia). The Raffles Bulletin of Zoology 54(1):181-197.
Whitaker R, Whitaker N. 2008. Who‟s got the biggest. Crocodile Specialist Group Newsletter, 27(4):26-30.
Whitaker R, Basu D. 1982. The gharial (Gavialis gangeticus): A review. Journal of the Bombay Natural History Society. 79(3):531-548.
Whitten T, Henderson G, Mustafa M. 2013. Ecology of Sulawesi. Jakarta (ID): Tuttle Publishing. Hlm 34–53.
Willis RE, McAliley LR, Neeley ED, Densmore LD. 2007. Evidence for placing the false gharial (Tomistoma schlegelii) into the famili Gavialidae: inferences
22
from nuclear gene sequences. Molecular Phylogenetics and Evolution. 43(3):787-794.
Watson LDMS. 1917. The evolution of the tetrapod shoulder girdle and forelimb. Journal of Anatomy. 52:1-63.
Webb GJ, Gans C. 1982. Galloping in Crocodylus johnstoni: A reflection of terrestrial activity. Records of the Australian Museum, 34:607-618.
23
RIWAYAT HIDUP
Penulis memiliki nama lengkap Alif Rahman Rahim Puarada, lahir di Jayapura, 09 November 1994 dari ayah Wahidin Puarda dan ibu Siti Hadra Kadir. Penulis merupakan anak ketiga dari enam bersaudara. Penulis menyelesaikan pendidikan dasar di SD Yapis Fak-fak, Papua Barat. Kemudian Penulis melanjutkan pendidikan di SMP N1 Fak-Fak, Papua Barat dan SMA Lazuardi GIS Depok, Jawa Barat. Penulis melanjutkan pendidikan S1 di Institut Pertanian Bogor pada tahun 2012 melalui jalur Ujian Seleksi Masuk IPB. Selama masa perkuliahan, Penulis pernah tergabung ke dalam HIMPRO (Himpunan Minat dan Profesi) Satwa Liar. Penulis juga aktif dalam Organisasi Mahasiswa IMAKAHI (Ikatan Mahasiswa Kedokteran Hewan Indonesia). Penulis pernah memimpin acara pengabdian masyarakat „Masyarakat Abdi Nusantara IX‟ yang dilaksanakan di Kabupaten Banten.