UJI STABILITAS GALUR-GALUR PADI GOGO HASIL
KULTUR ANTERA SELAMA DUA MUSIM
ADIN AFIYATA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2013
Dengan ini saya menyatakan bahwa tesis Uji Stabilitas Galur-Galur Padi Gogo Hasil Kultur Antera Selama Dua Musim adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini
. Bogor, Januari 2013
Adin Afiyata
ABSTRACT
ADIN AFIYATA. Yield Stability Evaluation of Upland Rice Lines from Anther
Culture for Two Seasons. Under direction of BAMBANG SAPTA PURWOKO as chairman, ISWARI SARASWATI DEWI and MUHAMAD SYUKUR as members of the advisory committee.
The objective of this research was to study the adaptation and yield stability of upland rice lines obtained from anther culture. Ten lines of upland rice obtained from anther culture and two check varieties (Way Rarem and Batutegi) were evaluated for their potential yield in seven different locations in the rainy season 2010/2011 and 2011/2012. In each location, the design was Randomized Complete Block Design with four replications. The method of Francis & Kannenberg, Finlay & Wilkinson, Eberhart & Russel, and AMMI (Additive Main Effect Multiplicative Interaction) were used to analyze the adaptation and yield stability. WI-44 line achieved the highest yield average (4.88 ton ha-1), higher than Batutegi and not significantly different from Way Rarem and has the highest potential yield (10.28 ton ha-1) in Malang in the second season. IW-67 yielded an average of 4.53 ton ha-1, not significantly different from Batutegi and has potential yield (9.09 ton ha-1) in Malang in the second season. WI-44 line has a superior yield average in seven environment. WI-44, IW-67 and IW-56 lines have an ability to adapt in optimal environments. I5-10-1-1, GI-7 and O18-b-1 were adapted in the marginal environments with yield above the average of the genotype tested.
RINGKASAN
ADIN AFIYATA. Uji Stabilitas Galur-Galur Padi Gogo Hasil Kultur Antera Selama Dua Musim. Dibimbing oleh BAMBANG SAPTA PURWOKO sebagai ketua, ISWARI SARASWATI DEWI dan MUHAMAD SYUKUR sebagai anggota komisi pembimbing.
Ketahanan pangan menjadi penting sebagai salah satu faktor penjamin stabilitas nasional. Peningkatan produksi padi ditempuh melalui usaha pengembangan padi pada lahan kering. Program pemuliaan tanaman pengembangan padi gogo dipilih sebagai salah satu usaha potensial pemanfaatan lahan kering. Teknologi kultur antera dilaporkan mampu mempersingkat waktu untuk mendapatkan galur homozigot fertil.
Stabilitas adalah kemampuan tanaman untuk mempertahankan daya hasil terhadap perubahan kondisi lingkungan. Uji multilokasi sebagai uji adaptabilitas merupakan suatu tahapan penting sebelum varietas dilepas dan hasil uji multilokasi diharapkan dapat diperoleh genotipe-genotipe yang dapat beradaptasi baik di lingkungan tertentu dan berdaya hasil stabil pada beberapa lingkungan.
Penelitian ini bertujuan untuk mendapatkan galur padi gogo yang memiliki potensi hasil tinggi dan stabil pada lingkungan yang luas. Pengujian dilaksanakan di tujuh lokasi selama dua musim hujan (MH) 2010/2011 dan 2011/2012. Lokasi pengujiannya yaitu : Taman Bogo dan Natar – Lampung, Sukabumi dan Indramayu – Jawa Barat, Purworejo – Jawa Tengah, Wonosari – DI Yogyakarta, dan Malang – Jawa Timur.
Bahan pengujian ialah 12 genotipe, yang terdiri atas 10 galur harapan padi gogo hasil kultur antera dan dua varietas pembanding. Sepuluh galur harapan padi gogo tersebut ialah III3-4-6-1, I5-10-1-1, WI-44, GI-7, O18-b-1, IW-67, IG-19, IG-38, IW 56, B13-2e dan varietas pembandingnya adalah Batutegi dan Way Rarem. Pelaksanaan pengujian di tiap lokasi menggunakan Rancangan Acak Kelompok (RAK) yang diulang sebanyak empat kali. Satuan percobaan berupa petakan lahan berukuran 4 m x 5 m, sehingga tiap lokasi terdiri atas 48 satuan percobaan. Pengujian stabilitas menggunakan empat metode yaitu Francis & Kanennberg, Finlay & Wilkinson, Eberhart & Russel, dan AMMI.
Pengujian di tujuh lokasi selama dua musim menunjukkan, galur WI-44 memiliki rata-rata produktivitas paling tinggi (4.88 ton ha-1) lebih tinggi dibanding Batutegi dan tidak berbeda nyata dari Way Rarem dengan potensi produktivitas mencapai 10.28 ton ha-1 pada lingkungan Malang musim ke dua. Galur IW-67 memiliki rata-rata produktivitas (4.53 ton ha-1) tidak berbeda nyata dengan Batutegi dan memiliki potensi produktivitas (9.09 ton ha-1) pada lingkungan Malang musim ke dua. Produktivitas gabah kering giling rata-rata galur WI-44 unggul di tujuh lingkungan, I5-10-1-1 unggul di empat lingkungan dan IW-67 unggul di dua lingkungan. Galur WI-44, IW67 dan IW-56 merupakan galur-galur yang peka terhadap perubahan lingkungan dan beradaptasi khusus pada lingkungan optimal. Galur-galur I5-10-1-1, GI-7 dan O18-b-1 merupakan galur yang mampu beradaptasi pada lingkungan marginal dan memiliki rata-rata produktivitas yang lebih tinggi dibanding rata-rata lingkungannya.
© Hak Cipta Milik IPB, tahun 2013
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun tanpa izin IPB
UJI STABILITAS GALUR-GALUR PADI GOGO HASIL
KULTUR ANTERA SELAMA DUA MUSIM
ADIN AFIYATA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2013
Judul Tesis : Uji Stabilitas Galur-Galur Padi Gogo Hasil Kultur Antera Selama Dua Musim
Nama : Adin Afiyata
NRP : A253100111
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Bambang Sapta Purwoko, M.Sc. Ketua
Dr. Muhamad Syukur, S.P. M.Si. Dr. Ir. Iswari Saraswati Dewi
Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, M.Si.
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Alloh SWT atas segala rahmat dan karunia-Nya sehingga penulisan tesis ini dapat diselesaikan. Judul tesis ini adalah Analisis Stabilitas Hasil Galur-galur Padi Gogo Hasil Kultur Antera Selama Dua Musim. Tesis merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Sekolah Pasca Sarjana, Institut Pertanian Bogor.
Dengan terselesaikannya penulisan tesis ini, saya ucapkan terima kasih yang sebesar-besarnya kepada :
1. Prof. Dr. Ir. Bambang Sapta Purwoko, M.Sc., Dr. Ir. Iswari Saraswati Dewi dan Dr. Muhamad Syukur, S.P. M.Si. selaku komisi pembimbing atas bimbingan dan arahannya selama perencanaan, pelaksanaan, dan penulisan tesis ini.
2. Program IMHERE atas pendanaan dan bantuan fasilitas dalam pelaksanaan penelitian ini (Prof. Dr. Ir. Bambang Sapta Purwoko, M.Sc. sebagai ketua peneliti).
3. Dr. Desta Wirnas, S.P. M.Si. selaku dosen penguji ujian tesis ini.
4. Orang tua dan keluarga atas dukungan penyelesaian studi memperoleh gelar Magister Sains.
5. Teman-teman S2 PBT angkatan 2010 atas kebersamaan dan kekompakan selama ini.
Penulis berharap tesis ini bermanfaat bagi yang memerlukan.
Bogor, Desember 2012
RIWAYAT HIDUP
Penulis lahir di Purwokerto pada tanggal 31 Agustus 1988 sebagai anak pertama dari pasangan Subechi dan Sri Lestari Tutiningsih. Penulis menyelesaikan pendidikan dasar sampai Sekolah Menengah Atas di Purwokerto tahun 1994 sampai dengan 2006. Penulis menyelesaikan studi sarjana tahun 2010 di Program Studi Pemuliaan Tanaman, Jurusan Budidaya Pertanian, Fakultas Pertanian, Universitas Jenderal Soedirman.
Penulis tercatat sebagai mahasiswa pada Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pasca Sarjana, Institut Pertanian Bogor, Tahun akademik 2010/2011.
xvii
DAFTAR TABEL ... xix
DAFTAR GAMBAR ... xxi
DAFTAR LAMPIRAN ... xxiii
PENDAHULUAN ... 1 Latar Belakang... 1 Tujuan ... 3 Hipotesis ... 4 TINJAUAN PUSTAKA... 5 Botani Padi ... 5
Kultur Antera pada Program Pemuliaan Padi ... 7
Interaksi Genotipe x Lingkungan ... 10
Adaptabilitas dan Stabilitas Hasil ... 11
BAHAN DAN METODE ... 19
Waktu dan Tempat ... 19
Bahan dan Alat ... 19
Metode Penelitian ... 19
Pelaksanaan Penelitian ... 20
Pengamatan ... 21
Analisis Data ... 22
Tahapan Analisis Data ... 22
Uji Normalitas ... 22
Analisis Ragam Tiap Lokasi ... 22
Uji Kehomogenan Ragam ... 24
Analisis Ragam Gabungan ... 24
Analisis Stabilitas ... 25
HASIL DAN PEMBAHASAN ... 31
Kondisi Umum Penelitian ... 31
Analisis Stabilitas Hasil Selama Dua Musim Tanam ... 32
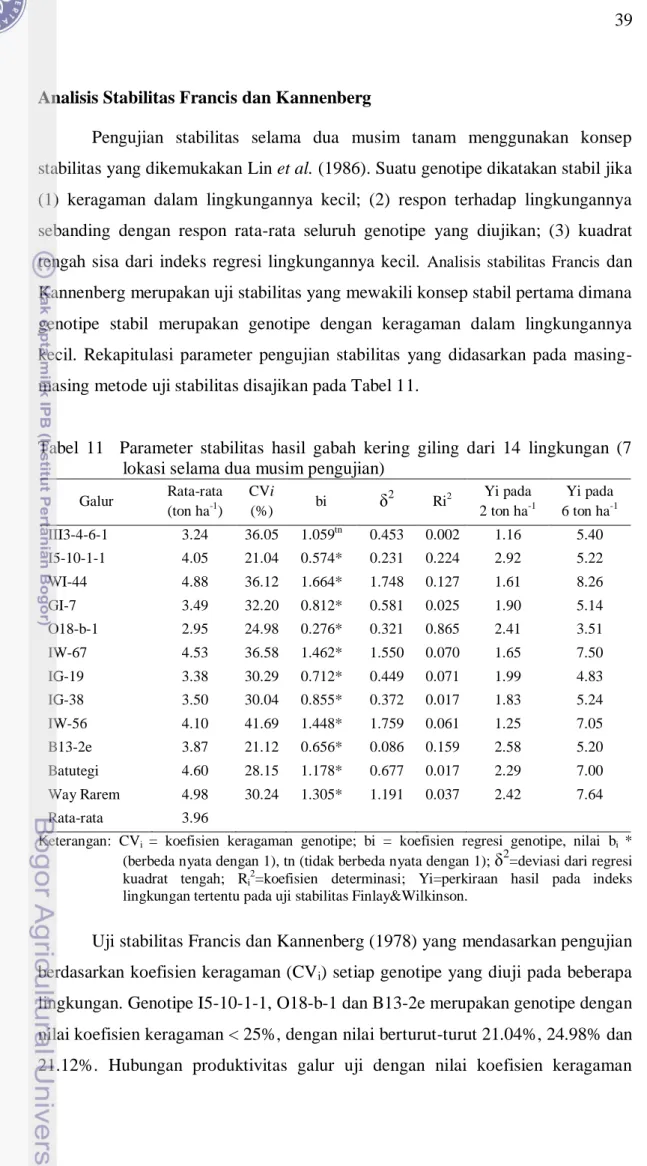
Analisis Stabilitas Francis dan Kannenberg ... 39
Analisis Stabilitas Finlay dan Wilkinson ... 41
xviii
Tinggi Tanaman ... 50
Jumlah Anakan Total dan Jumlah Anakan Produktif ... 53
Umur Berbunga dan Umur Panen ... 57
Panjang Malai ... 61
Gabah Isi, Persentase Gabah Isi dan Gabah Hampa ... 62
Bobot 1000 butir ... 68
SIMPULAN DAN SARAN ... 73
Simpulan ... 73
Saran ... 73
DAFTAR PUSTAKA ... 75
xix 1 Perbandingan waktu pemuliaan antara sistem pedigree dan kultur
antera ... 8
2 Masalah dan penelitian yang telah dilakukan untuk penggunaan kultur antera dalam program pemuliaan padi ... 9
3 Pengelompokan metode analisis stabilitas oleh Lin et al. (1986) ... 12
4 Galur-galur dan varietas pembanding padi gogo yang digunakan dalam penelitian ... 19
5 Sidik ragam karakter padi gogo pada masing-masing lokasi uji ... 24
6 Sidik ragam gabungan menggunakan model acak ... 25
7 Sidik ragam analisis stabilitas Eberhart dan Russel ... 28
8 Model analisis ragam AMMI ... 29
9 Sidik ragam gabungan produktivitas gabah kering giling dari 14 lingkungan ... 33
10 Rata-rata produktivitas di 7 lokasi selama dua musim ... 38
11 Parameter stabilitas hasil gabah kering giling dari 14 lingkungan ... 39
12 Sidik ragam AMMI 14 lingkungan ... 45
13 Rekapitulasi analisis stabilitas pada genotipe-genotipe yang diuji ... 48
14 Analisis ragam gabungan pengaruh genotipe (G), lokasi (E), dan interaksi G × E karakter agronomi padi gogo pada 7 lokasi selama dua musim ... 49
15 Variabel komponen hasil rata-rata dari 7 lokasi selama dua musim tanam ... 50
16 Rata-rata tinggi tanaman di 7 lokasi selama dua musim ... 52
17 Rata-rata jumlah anakan total di 7 lokasi selama dua musim ... 54
18 Rata-rata jumlah anakan produktif di 7 lokasi selama dua musim ... 56
19 Rata-rata umur berbunga di 7 lokasi selama dua musim ... 58
20 Rata-rata umur panen di 7 lokasi selama dua musim ... 60
21 Rata-rata periode pengisian biji dan efisiensi laju pembentukan hasil ... 61
xx
25 Rata-rata tingkat kerapatan malai ... 67 26 Rata-rata gabah hampa per malai di 7 lokasi selama dua musim ... 69 27 Rata-rata bobot 1000 butir di 7 lokasi selama dua musim ... 71
xxi 1 Skema jalur evolusi padi Asia dan Afrika ... 6 2 Bagan alir penelitian ... 23 3 Interpretasi umum nilai b dari pola populasi genotipe ketika koefisien
regresi genotipe diplot terhadap nilai rata-rata hasil genotipe... 26 4 Rata-rata kekurangan dan kelebihan produktivitas galur-galur yang diuji
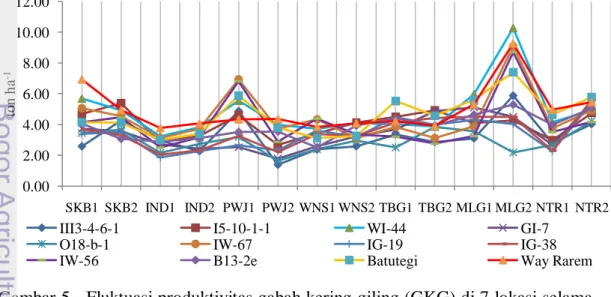
terhadap pembanding ... 34 5 Fluktuasi produktivitas gabah kering giling (GKG) di 7 lokasi selama
dua musim ... 35 6 Hubungan koefisien keragaman genotipe dengan rata-rata produktivitas ... 40 7 Pola linier produktivitas genotipe-genotipe padi gogo yang tergolong
stabil berdasarkan metode Finlay & Wilkinson... 41 8 Pola linier produktivitas genotipe-genotipe padi gogo yang tergolong
stabil di bawah rata-rata berdasarkan metode Finlay & Wilkinson ... 42 9 Pola linier produktivitas genotipe-genotipe padi gogo yang tergolong
stabil di atas rata-rata berdasarkan metode Finlay & Wilkinson ... 43 10 Interpretasi dari parameter bi dan δ2 pada pendekatan regresi untuk
menentukan stabilitas ... 44 11 Biplot pengaruh interaksi model AMMI2 untuk produktivitas GKG
xxiii
1 Asal persilangan galur-galur padi gogo yang digunakan ... 83
2 Silsilah dan deskripsi tetua galur-galur padi gogo yang digunakan ... 84
3 Deskripsi Varietas Padi Gogo... 85
4 Karakteristik umum lingkungan pengujian ... 90
Latar Belakang
Ketahanan pangan merupakan salah satu faktor penjamin stabilitas nasional. Beras merupakan makanan pokok di Indonesia. Konsumsi beras tahun 2011 139 kg/kapita (Puslitbang Tanaman Pangan 2012) dan perkiraan jumlah penduduk Indonesia pada tahun 2011 sebesar 241 juta jiwa (BPS 2012) memerlukan ketersediaan beras yang mencukupi. Dengan laju pertumbuhan penduduk 1.49% (BPS 2012), kebutuhan akan beras meningkat. Tingginya tingkat konsumsi beras dan terbatasnya produksi padi nasional dapat menurunkan ketahanan pangan nasional.
Data Kementerian Pertanian (2012) menunjukkan produksi padi tahun 2011 sebesar 66.76 juta ton dengan produktivitas 4.98 ton ha-1. Kontribusi padi gogo terhadap produksi padi nasional hanya sebesar 4.84% (3.23 juta ton). Produksi padi nasional masih belum mencukupi kebutuhan beras penduduk Indonesia.
Kendala dalam mempertahankan ketahanan pangan antara lain ialah adanya konversi lahan optimal bagi budidaya tanaman pangan terutama lahan sawah. Perluasan sawah makin sulit dilakukan. Hal ini menjadikan arah ekstensifikasi lahan tanaman pangan ke lahan suboptimal antara lain lahan kering. Abdurachman et al. (2008) menyebutkan bahwa Indonesia memiliki lahan kering sekitar 148 juta ha (78%) dan lahan basah seluas 40.20 juta ha (22%) dari 188.20 juta ha total luas daratan. Total luas lahan kering 148 juta ha, yang sesuai untuk budidaya pertanian hanya sekitar 76.22 juta ha (52%), sebagian besar terdapat di dataran rendah sebesar 70.71 juta ha atau (93%). Luas wilayah dataran rendah yang sesuai untuk pertanian tanaman pangan mencakup 23.26 juta ha dan pada dataran tinggi hanya sekitar 2.07 juta ha. Potensi lahan kering tidak terlepas dari masalah lahan sub optimal yaitu kesuburan tanah yang rendah, cekaman biotik (hama penyakit) dan cekaman abiotik.
Pengembangan padi gogo berpotensi meningkatkan produksi beras nasional. Program pengembangan tersebut perlu didukung pemuliaan tanaman sebagai upaya mengatasi permasalahan dalam pengembangan teknologi budidaya
padi gogo. Perakitan padi gogo yang toleran terhadap cekaman abiotik tertentu dan mampu beradaptasi luas pada lingkungan suboptimal menjadi topik program pemuliaan padi gogo. Program pemuliaan diharapkan mampu merakit beberapa kultivar padi gogo yang mempunyai potensi hasil tinggi. Program pemuliaan tanaman bertujuan untuk memperoleh varietas baru dengan produksi tinggi dan memiliki kestabilan pada berbagai lingkungan. Kegiatan pemuliaan yang dilakukan mencakup persilangan, introduksi, mutasi, introgresi gen, seleksi dan evaluasi.
Perakitan varietas baru perlu terus dilakukan karena varietas-varietas padi gogo yang sudah ada walaupun daya hasilnya sudah cukup tinggi, namun masih menghadapi cekaman abiotik dan biotik akibat lahan potensial pengembangan padi gogo merupakan lahan suboptimal. Perakitan padi gogo toleran lingkungan abiotik seperti toleran aluminium (Bakhtiar 2007) dan toleran naungan (Sasmita 2006) telah dilakukan untuk pengembangan padi gogo di daerah suboptimal. Perakitan dimulai dengan menyeleksi tetua yang memiliki ketahanan terhadap cekaman abiotik tersebut. Metode pemuliaan tanaman konvensional membutuhkan banyak waktu penggaluran, seleksi dan selanjutnya pengujian galur-galur yang mampu beradaptasi dengan cekaman abiotik tersebut. Proses mempercepat pembentukan galur murni padi gogo tipe baru dengan sifat-sifat yang diharapkan dari induknya dapat dilakukan dengan mempergunakan metode kultur antera (Abdullah et al. 2008, Dewi dan Purwoko 2001, 2011). Pembentukan galur murni dengan metode kultur antera hanya memerlukan waktu kurang lebih 12 bulan (Sasmita 2007). Proses ini lebih cepat bila dibandingkan dengan cara konvensional yang membutuhkan waktu yang lama (5-10 tahun) untuk mendapatkan kestabilan genetik galur-galur hasil persilangan (Safitri et al. 2010) atau memerlukan 10-12 generasi setelah persilangan (Sasmita 2007). Metode kultur antera akan menghasilkan tanaman dihaploid yang homozigos fertil (Safitri et al. 2010). Kultur antera mempercepat mendapatkan galur murni, tetapi waktu yang dibutuhkan untuk evaluasi uji daya hasil dan uji stabilitas tetap sama. Peran kultur antera dalam perakitan galur adalah memperoleh tanaman homogen homozigos dalam waktu relatif singkat, efisiensi proses seleksi meningkat, variabilitas genetik meningkat melalui produksi variasi gametoklonal,
mempercepat terekspresinya karakter yang dikendalikan gen resesif, efisiensi biaya, waktu dan tenaga kerja (Dewi dan Purwoko 2001, Abdullah et al. 2008, Safitri et al. 2010).
Galur-galur harapan padi gogo hasil program pemuliaan perlu diuji sebelum dilepas. Tahapan pelepasan varietas menurut peraturan Menteri Pertanian No 61 tahun 2011 mengenai metoda baku uji adaptasi dan uji observasi. Observasi daya hasil (ODH) merupakan uji lapangan untuk mengetahui karakter agronomi dan daya hasil pada tahap awal. Hal tersebut diikuti dengan uji daya hasil pendahuluan (UDHP) yang dilakukan pada petak uji minimum dan uji daya hasil lanjut (UDHL) yang dilakukan pada petak uji minimum serta dilakukan di beberapa lokasi pengujian berbeda atau musim berbeda. Uji stabilitas dilakukan setelah uji daya hasil lanjut dengan tujuan untuk memperoleh informasi tingkat stabilitas hasil galur-galur yang akan dilepas. Hasil pengujian menjadi dasar pertimbangan pelepasan suatu galur menjadi varietas.
Galur-galur padi gogo dihaploid dari beberapa persilangan beberapa varietas unggul pada penelitian sebelumnya melalui metode kultur antera telah diperoleh. Galur-galur IW-56, IW-67, IG-19, IG-38, GI-8 merupakan galur-galur hasil kultur antera yang toleran naungan (Sasmita 2007). Galur O18-b-1, B13-2e (Bambang Sapta Purwoko, komunikasi pribadi), III3-4-6-1 dan I5-10-1-1 (Herawati et al. 2009) adalah galur toleran aluminium hasil kultur antera. Galur-galur yang dihasilkan perlu diuji adaptasi dan stabilitasnya untuk melihat keragaannya di berbagai kondisi lingkungan sesuai aturan pelepasan varietas. Pengujian multi lokasi tahun I telah dilakukan pada tahun 2011. Oleh karena itu perlu dilakukan pengujian multi lokasi tahun II untuk melihat potensi hasil dan adaptabilitas padi gogo dihaploid di dua tahun uji, sesuai syarat pelepasan varietas.
Tujuan Tujuan penelitian ini ialah untuk:
1. mengevaluasi daya adaptasi dan stabilitas hasil galur-galur harapan padi gogo selama dua musim pengujian.
2. memperoleh galur-galur harapan padi gogo yang stabil statis untuk dilepas sebagai varietas
Hipotesis
Hipotesis yang diajukan pada penelitian ini ialah: 1. terdapat perbedaan hasil pada lokasi yang berbeda.
2. terdapat interaksi genotipe lingkungan (G x E) yang nyata. 3. terdapat galur yang stabil statis.
Botani Padi
Klasifikasi botani tanaman padi menurut Vaughan et al. (2003) tergolong kingdom Plantae, divisi Spermatophyta, digolongkan dalam sub divisi
Angiospermae, karena biji berkeping satu digolongkan ke dalam kelas Monocotyledonae, dengan ordo Glumiflorae termasuk dalam keluarga
rumput-rumputan atau disebut juga Graminae (Poaceae) dan dikenal dengan genus Oryza. Padi yang dikenal dan dibudidayakan sebagaian besar tergolong dalam spesies
Oryza sativa.
Genus Oryza terdistribusi di seluruh daerah tropis dan subtropis di dunia. Genus Oryza terdiri atas 23 spesies liar dan dua spesies yang dibudidayakan, yaitu, O. sativa di daerah Asia dan O. glaberrima di daerah Afrika. Oryza sativa didomestikasi di Asia, namun sekarang telah menyebar ke hampir seluruh daerah-daerah di dunia, sedangkan O. glaberrima, terbatas di barat Afrika. Jumlah kromosom dasar dari genus ini adalah n = 12. Spesies normal memiliki kromosom dalam keadaan diploid dengan 2n = 2x = 24 kromosom atau tetraploid dengan 2n = 2x = 48 kromosom. Berdasarkan analisis genom dan tingkat kompatibilitas persilangan, spesies Oryza dikelompokkan menjadi sembilan genom yang berbeda, yaitu, A, B, C, D, E, F, G, H dan J (Tripathi et al. 2011).
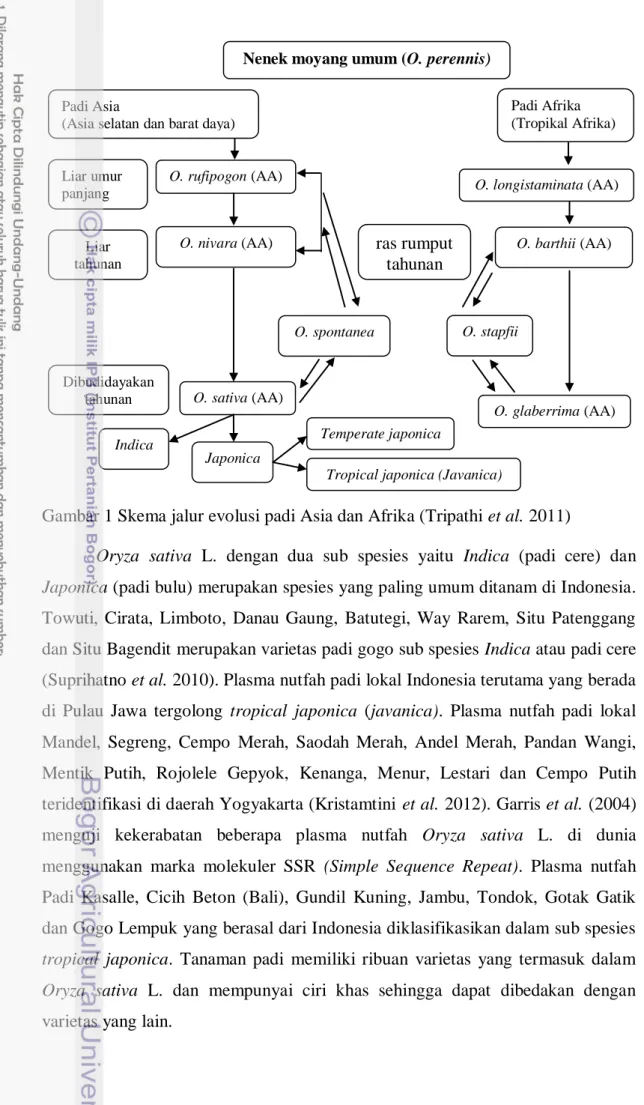
Pusat asal dan pusat keanekaragaman dua spesies yang telah dibudidayakan (O. sativa dan O. glaberrima) telah diidentifikasi menggunakan keanekaragaman genetik, bukti-bukti sejarah, arkeologi dan penyebaran geografis. Daerah Asia terutama daerah Cina dan India merupakan pusat asal O. sativa sementara daerah Niger di Afrika sebagai pusat utama O. glaberrima. Dua spesies mirip diyakini berevolusi secara alami dari dua nenek moyang yang berbeda, yaitu. O. nivara dan O. barthii. Gambar 1 merujuk pada skema evolusi padi di daerah Asia dan Afrika. Nenek moyang O. sativa disimbolkan menjadi genom AA sebagai spesies diploid Asia dan O. glaberrima menjadi genom AA spesies diploid Afrika (Gambar 1).
Gambar 1 Skema jalur evolusi padi Asia dan Afrika (Tripathi et al. 2011)
Oryza sativa L. dengan dua sub spesies yaitu Indica (padi cere) dan Japonica (padi bulu) merupakan spesies yang paling umum ditanam di Indonesia.
Towuti, Cirata, Limboto, Danau Gaung, Batutegi, Way Rarem, Situ Patenggang dan Situ Bagendit merupakan varietas padi gogo sub spesies Indica atau padi cere (Suprihatno et al. 2010). Plasma nutfah padi lokal Indonesia terutama yang berada di Pulau Jawa tergolong tropical japonica (javanica). Plasma nutfah padi lokal Mandel, Segreng, Cempo Merah, Saodah Merah, Andel Merah, Pandan Wangi, Mentik Putih, Rojolele Gepyok, Kenanga, Menur, Lestari dan Cempo Putih teridentifikasi di daerah Yogyakarta (Kristamtini et al. 2012). Garris et al. (2004) menguji kekerabatan beberapa plasma nutfah Oryza sativa L. di dunia menggunakan marka molekuler SSR (Simple Sequence Repeat). Plasma nutfah Padi Kasalle, Cicih Beton (Bali), Gundil Kuning, Jambu, Tondok, Gotak Gatik dan Gogo Lempuk yang berasal dari Indonesia diklasifikasikan dalam sub spesies
tropical japonica. Tanaman padi memiliki ribuan varietas yang termasuk dalam Oryza sativa L. dan mempunyai ciri khas sehingga dapat dibedakan dengan
varietas yang lain.
Nenek moyang umum (O. perennis)
O. rufipogon (AA)
O. longistaminata (AA)
O. nivara (AA) O. barthii (AA)
O. sativa (AA)
O. stapfii O. spontanea
O. glaberrima (AA) Temperate japonica
Tropical japonica (Javanica) Japonica
Indica
Padi Asia
(Asia selatan dan barat daya)
Padi Afrika (Tropikal Afrika) ras rumput tahunan Liar umur panjang Liar tahunan Dibudidayakan tahunan
Varietas unggul padi gogo yang mampu beradaptasi pada cekaman biotik maupun abiotik masih kurang sehingga memacu berkembangnya program-program pemuliaan padi gogo di Indonesia. Varietas unggul padi gogo yang telah dilepas dari tahun 1988 – 1993 antara lain: Batur, Danau Atas, Poso, Laut Tawar, C22 dan Danau Tempe memiliki hasil tinggi (potensi hasil 3.0-5.0 ton ha-1) namun memiliki kelemahan pada mutu beras yang rendah. Periode tahun 2001 – 2003 telah dilepas empat varietas padi gogo (potensi hasil 4.0-5.0 ton ha-1) yaitu Danau Gaung, Batutegi, Situ Patenggang dan Situ Bagendit. Inpago 4, Inpago 5, Inpago 6, Inpago 8, Inpago Unram 1 dan Inpago Unsoed 1 merupakan varietas padi gogo yang dilepas pada periode tahun 2010 – 2011 dengan potensi hasil 5.8-8.1 ton ha-1 (Kementerian Pertanian, 2012).
Kultur Antera pada Program Pemuliaan Padi
Perakitan varietas tanaman menyerbuk sendiri seperti padi ditujukan untuk memperoleh galur yang hampir mendekati 100% homozigotitas dengan sifat-sifat yang unggul. Metode konvensional dalam perakitan varietas memerlukan prosedur penelitian yang sistematik, mulai dari perumusan tujuan, pemilihan tetua, persilangan, seleksi galur, pengujian daya hasil hingga pelepasan varietas yang membutuhkan waktu 7 -10 tahun (Dewi dan Purwoko 2001). Galur-galur murni baru akan terbentuk dengan cara persilangan yang diikuti oleh serangkaian seleksi pada tiap generasi, seperti pada pedigree.
Kombinasi metode persilangan dengan metode kultur antera dalam program pemuliaan padi mampu meningkatkan efektifitas dan efisiensi seleksi. Dewi dan Purwoko (2001) menjelaskan bahwa galur murni dapat diseleksi dari populasi dihaploid yang homogen dan homozigot. Populasi seleksi menjadi lebih sedikit bergantung pada jumlah gen untuk seleksi. Jika perbedaan pada tetua persilangan sebanyak n (jumlah gen yang diinginkan) gen dan diasumsikan tidak terpaut maka dengan dihaploid hanya perlu menanam 2n tanaman agar semua genotipe homozigot terwakili. Pemuliaan konvensional membutuhkan 4n tanaman. Perbandingan waktu penggunaan metode kultur antera dan konvensional tertera pada Tabel 1.
Tabel 1 Perbandingan waktu pemuliaan antara sistem pedigree dan kultur antera (Dewi dan Purwoko 2001)
Waktu Sistem pedigree Kultur Antera
1 Hibridisasi Hibridisasi
2 F1 F1 dan Kulture Antera
3 F2 Perbanyakan Benih Skrining dan pengujian
4 Pedegree (F3-F9) (Skrining dan Pengujian-pengujian) Uji Adaptabilitas Regional
Uji daya hasil
5 Uji Adaptasi Regional Perbanyakan Benih
6 7 8 9 10 11 12 13 14 Perbanyakan Benih 15
Pembentukan tanaman dihaploid dari tanaman haploid diperoleh melalui dua tahap, yaitu tahap induksi butir tepung sari menjadi kalus atau embrioid dan selanjutnya tahap diferensiasi menjadi tanaman kecil atau planlet. Tanaman dihaploid asal kultur antera dapat diperoleh dengan induksi kolkisin atau secara spontan melalui fusi spontan inti sel dan dengan pemangkasan (ratooning) (Dewi dan Purwoko 2001). Tanaman haploid dapat diinduksi melalui proses
androgenesis dengan kultur antera dan kultur mikrospora atau proses gynogenesis
dengan kultur ovule, hibridisasi jenis tanaman yang berbeda (distant
hybridization), polinasi tertunda (delayed pollination), penggunaan polen yang
sudah diradiasi, perlakuan hormon, shock dengan temperatur tinggi (Poelman dan Sleper 1995). Karakter tanaman haploid dan dihaploid ditampilkan pada keadaan hemizygous yang dikendalikan oleh gen dominan maupun gen resesif, sehingga seleksi lebih mudah dilakukan pada tingkat monohaploid atau dihaploid untuk menghasilkan tanaman super jantan unggul pada asparagus contohnya, menghasilkan tanaman dihaploid dan tetraploid homozigot dan tanaman triploid pada kentang, dan sebagai tetua pembentuk hibrida F1 contohnya pada kentang atau tanaman lain yang dibudidayakan secara vegetatif (Dewi dan Purwoko 2011). Tanaman haploid dapat digunakan untuk menghasilkan galur murni secara cepat tanpa melalui banyak generasi silang, sebagaimana yang dilakukan pada metode pemuliaan konvensional (Syukur et al. 2012).
Kultur antera padi mulai diintroduksikan di Indonesia pada tahun 1991 oleh beberapa peneliti yang dikirim ke IRRI. Awal penelitian kultur antera masih berfokus pada media untuk induksi kalus dan regenerasi. Tahun 1997, IPB bekerja sama dengan Balitbio dalam pengembangan kultur antera pada padi gogo. Penelitian berupa perbaikan media kultur antera menggunakan poliamin dan usaha perakitan padi gogo toleran cekaman intensitas cahaya rendah dan aluminium (Dewi dan Purwoko 2001). Penelitian dengan kultur antera melalui pemberian spermin dapat meregenerasikan tanaman hijau pada tiga genotipe dari subspesies indica (Purwoko 2004), namun regenerasi pada kultur antera padi F1 yang diberi 10-3M putresin memberikan hasil yang lebih baik (Dewi et al. 2007). Beberapa penelitian perbaikan melalui kultur antera disajikan pada Tabel 2.
Tabel 2 Masalah dan penelitian yang telah dilakukan untuk penggunaan kultur antera dalam program pemuliaan padi (Dewi dan Purwoko 2001)
Masalah yang Dipecahkan Penelitian/Studi Telah dilakukan pada :
Metode untuk mendapatkan jumlah tanaman yang cukup (seleksi, evaluasi)
1. Media; komposisi media dasar (MS, N6, LS, He, L8, M8)
ZPT: 2,4-D, NAA, IAA, Kinetin, BA
Suplemen: CW ekstrak labu, ekstrak kentang CH, poliamin
2. Anther Culture Ability
Genotipe (high anther culturability)
Kondisi fisiologi tanaman donor
Tahap perkembangan antera (uninukleat)
Pra perlakuan (suhu rendah 5-10 0 C)
Populasi antera
Subkultur kalus (ukuran 1-2 mm, jenis embriogenik)
Aklimatisasi (air, lumpur, intensitas cahaya)
Konfirmasi stabilitas genetik dari progeni
Studi generasi DH1-DH5 pada kondisi yang sama dengan hasil, vigor tidak menurun, karakter morfologi dan agronomi tetap stabil. Seleksi dapat dilakukan pada generasi awal.
Metode pemuliaan yang efisien, karena ada batasan terhadap rekombinasi gen pada kultur antera F1
1. Eksplan dari F2 terseleksi
2. Eksplan F1 dari metode persilangan multiple crosses (pesilangan ganda) 3. Metode seleksi recurrent yang
Interaksi Genotipe x Lingkungan
Stabilitas dapat dianalisis dari adanya interaksi genotipe dan lingkungan. Interaksi genotipe lingkungan menjadi penting pada uji stabilitas jika suatu individu yang merupakan bagian dari populasi tidak menunjukkan penampilan yang berbeda nyata pada lokasi berbeda. Interaksi genotipe dan lingkungan terjadi bila penampilan nisbi (relative performance) atau peringkat variabel hasil beberapa genotipe akan berubah dengan perubahan lingkungan. Hal ini menunjukkan mengapa kultivar-kultivar berdaya hasil tinggi yang ditanam pada suatu lingkungan akan memberikan hasil yang berbeda pada lingkungan yang lain. Perbedaan lingkungan yang spesifik memiliki efek lebih besar terhadap variabel hasil untuk suatu genotipe dibandingkan genotipe yang lain (Falconer dan Mackay 1996).
Ada tidaknya pengaruh interaksi dapat dideteksi dari perilaku respon suatu faktor pada berbagai kondisi faktor lain. Jika respon suatu faktor berubah pola dari kondisi tertentu ke kondisi yang lain untuk faktor yang lain maka kedua faktor dikatakan berinteraksi. Jika pola respon dari suatu faktor tidak berubah pada berbagai kondisi faktor yang lain, maka dapat dikatakan kedua faktor tersebut tidak berinteraksi (Mattjik dan Sumertajaya 2006).
Ada atau tidaknya interaksi antar genotipe atau genotipe-genotipe tanaman dengan kisaran variasi lingkungan spasial yang luas, ataupun dengan variasi lingkungan pada suatu wilayah spesifik merupakan hal yang sangat penting bagi pemulia dalam menentukan genotipe tanaman yang akan dipilih untuk dilepas atau dirilis, ataupun untuk digunakan dalam mengukur komponen ragam suatu karakter tertentu (Baihaki dan Noladhi 2005).
Interpretasi dan pemanfaatan informasi interaksi G × E bervariasi antar peneliti. Eberhart dan Russel (1966) menyatakan bahwa interaksi G × E dapat mempengaruhi kemajuan seleksi dan sering mengganggu dalam seleksi genotipe-genotipe unggul. Nasrullah (1981) berpendapat bahwa interaksi G × E sering mempersulit pengambilan pilihan dari suatu percobaan varietas uji multilokasi yang kisaran lingkungannya luas. Informasi interaksi G × E sangat penting bagi negara-negara yang variabilitas biogeofisiknya luas seperti Indonesia. Pemulia dapat memanfaatkan potensi lingkungan spesifik dalam kebijakan penentuan
penerapan kebijakan wilayah sebaran suatu varietas unggul baru. Dalam hal ini ada dua alternatif pilihan, yaitu : (1) melepas varietas unggul baru dengan potensi hasil tinggi untuk kisaran spasial yang luas (wide adaptability), (2) melepas varietas unggul baru dengan potensi hasil tinggi pada wilayah tumbuh yang spesifik (spesifik lingkungan tumbuh-specific adaptability).
Adaptabilitas dan Stabilitas Hasil
Rendahnya jumlah varietas yang beradaptasi luas dan terabaikannya varietas yang beradaptasi sempit, serta rendahnya jumlah varietas unggul baru yang dilepas, dapat menggambarkan rendahnya tingkat efisiensi dan efektivitas proses pelepasan varietas unggul di Indonesia. Hal ini akan dapat diatasi apabila varietas unggul spesifik wilayah juga diperhitungkan dalam kebijakan pelepasan varietas, sehingga dapat menekan biaya dan waktu yang selama ini terbuang percuma. Baihaki dan Noladhi (2005) menyebutkan keuntungan yang akan diperoleh apabila varietas unggul spesifik wilayah dapat dilepas, antara lain : (1) efisiensi penggunaan dana dan waktu, (2) memperbanyak varietas unggul baru yang dilepas, (3) secara nasional produktivitas akan meningkat dan dengan sendirinya produksi akan meningkat pula, (4) akan menekan harga benih/ bibit, (5) akan terbentuk “regional buffering” yang sangat diperlukan untuk meredam meluasnya hama atau penyakit tanaman, (6) memberikan pilihan alternatif varietas yang cukup bagi petani, (7) memanfaatkan potensi kekayaan alam dengan baik, dan (8) mendorong terselenggaranya pembangunan pertanian yang sinambung. Beberapa pihak meragukan dilepasnya varietas unggul spesifik wilayah, dengan alasan bahwa varietas semacam ini tidak akan menarik industri perbenihan untuk memproduksinya, karena wilayah pemasarannya menjadi terbatas. Hal ini dapat diatasi dengan cara mengidentifikasi ciri-ciri wilayah yang serupa dengan wilayah pelepasan utama yang teridentifikasi dari uji multilokasi.
Konsep adaptabilitas dan stabilitas harus mampu diestimasi dan dibuktikan secara statistik. Metode analisis stabilitas dikelompokkan menjadi empat kelompok dan tiga konsep stabilitas berdasarkan deviasi pengaruh rata-rata genotipe, pengaruh interaksi genotipe dan lingkungan, serta pengaruh gabungan
keduanya (Lin et al. 1986). Pengelompokan metode analisis stabilitas tersebut tertera pada Tabel 3.
Kelompok A mendasarkan metode analisisnya pada deviasi pengaruh rata-rata genotipe. Stabilitas diukur berdasarkan pada terbentuknya variasi suatu genotipe dalam berbagai lingkungan. Kelompok B berdasar pada pengaruh interaksi genotipe dan lingkungan. Kelompok C dan D mendasarkan pada pengaruh gabungan deviasi rata-rata genotipe dan interaksi genotipe dan lingkungan.
Tabel 3 Pengelompokan metode analisis stabilitas oleh Lin et al. (1986)
Keempat kelompok tersebut mampu menjelaskan tiga konsep stabilitas yang dibuat Lin et al. (1986), dimana suatu genotipe dikatakan stabil jika : (1)
Kelompok Tipe Model Persamaan Penggagas Konsep
Stabilitas A 1 Si2 = (Xij− q j=1 Xi.)2/ q − 1 Francis & Kannenberg (1978) Statik 1 CVi= Si Xi. B 2 θi= p 2 p−1 (q−1) Xij− Xi.− X.j+ X.. q j=1 + SS GE 2 p−1 (q−1) Plaisted & Peterson (1959) Dinamik 2 θi= p−1 (p−2)(q−1)−p Xij− Xi.− X.j+ X.. q j=1 + SS GE p−2 (q−1) Plaisted (1960) Dinamik 2 Wi2= Xij− Xi.− X.j+ X.. q j=1 Wrickle (1962) Dinamik 2 ζi2= p p−2 (q−1) Xij− Xi.− X.j+ X.. q j=1 + SS GE (p−1) p−2 (q−1) Shukla (1972) Dinamik C 2 bi= Xij− Xi. q j=1 / X.j− X.. 2 q j=1 Finlay & Wilkinson (1963) Statik/ Dinamik 2 βi= Xij− Xi.− X.j+ X.. X.j− X.. / X.j− X.. 2 q j=1 q j=1 Perkins & Jinks (1968) Dinamik D 3 δi2= q−2 1 Xij− Xi. 2 q j=1 − βi2 X.j− X.. 2 q j=1 Eberhart & Russel (1966) Dinamik 3 δi2= q−2 1 Xij− Xi.− X.j+ X.. 2 q j=1 − βi2 X.j− X.. 2 q j=1 Perkins & Jinks (1968) Dinamik
memiliki koefisien keragaman yang kecil dalam lingkungannya (2) respon terhadap lingkungannya sebanding dengan rata-rata respon seluruh genotipe yang diuji, atau sebanding dengan indeks lingkungannya (3) memiliki kuadrat tengah sisa yang kecil dari garis regresi indeks lingkungannya. Konsep stabilitas tipe 1 dan 3 bersifat statis, dimana suatu genotipe hanya dapat dilihat stabil atau tidaknya saja. Adapun konsep stabilitas tipe 2 bersifat dinamis karena dapat menunjukan pola stabilitas dan adaptabilitas suatu genotipe. Kelompok A mampu menjelaskan konsep stabil tipe 1, kelompok B mampu menjelaskan konsep stabil tipe 2, kelompok D menjelaskan konsep stabil tipe 3, sedangkan kelompok C menjelaskan konsep stabil tipe 1 dan 2.
Analisis stabilitas Finlay dan Wilkinson (1963) didasarkan pada koefisien regresi (bi) antara hasil rata-rata suatu genotipe dengan rata-rata umum semua genotipe yang diuji dan semua lingkungan pengujian. Analisis ini dapat menjelaskan fenomena stabilitas dan adaptabilitas suatu genotipe. Genotipe-genotipe yang mempunyai slope regresi (bi) : > 1, = 1, dan < 1 berturut-turut mempunyai stabilitas di bawah rata-rata, setara rata-rata, dan di atas rata-rata.
Eberhart dan Russel (1966) mengembangkan metode pengujian stabilitas yang didasarkan pada deviasi dari regresi nilai rata-rata genotipe pada indeks lokasi (lingkungan). Suatu genotipe dikatakan stabil hanya bila kuadrat tengah sisa dari garis regresi adalah kecil. Nilai δ2 (parameter deviasi) yang besar atau Ri2
(koefisien determinasi) yang kecil menunjukkan bahwa model regresi yang diperoleh tidak menggambarkan data yang sebenarnya dan dengan sendirinya tidak dapat dipakai sebagai ukuran stabilitas.
Metode yang dapat digunakan dalam memvisualisasi dan menjelaskan respon genotipe terhadap lingkungan serta stabilitas daya hasilnya adalah metode
Additive Main Effect Multiplicative Interaction (AMMI). Analisis AMMI adalah
suatu teknik analisis data percobaan faktorial dengan pengaruh utama perlakuan bersifat aditif, sedangkan pengaruh interaksi dimodelkan dengan model bilinear. Pada dasarnya analisis AMMI menggabungkan analisis ragam aditif dengan pengaruh multiplikatif pada analisis komponen utama (Mattjik dan Sumertajaya, 2006).
Penggunaan analisis AMMI memiliki tiga tujuan, yaitu : (1) sebagai analisis pendahuluan untuk mencari model yang lebih tepat; (2) untuk menjelaskan interaksi galur x lingkungan (G × E); (3) meningkatkan keakuratan dugaan respon interaksi galur dan lingkungan. Tahapan analisis AMMI yang dilakukan adalah : (1) menyusun matriks pengaruh interaksi dalam bentuk matriks Ig × l (2) melakukan penguraian bilinear terhadap matriks Ig × l melalui SVD (singular value decomposition) (3) menentukan banyaknya Komponen Utama I (KUI) nyata melalui postdictive success (4) membuat biplot AMMI. Suatu galur dianggap stabil jika posisinya berada dekat dengan sumbu utama. Galur dianggap spesifik pada lokasi tertentu dapat dilihat melalui posisi masing-masing galur terhadap garis lokasi (Mattjik dan Sumertajaya 2006).
Untuk menentukan berapa banyak sumbu komponen utama yang dipakai sebagai penduga digunakan dua metode yaitu metode postdictive success dan
predictive succes. Metode postdictive success berhubungan dengan kemampuan
suatu model yang tereduksi untuk menduga data yang digunakan dalam membangun model tersebut. Salah satu caranya adalah berdasarkan banyaknya sumbu tersebut yang nyata pada uji F analisis ragam. Predictive success berhubungan dengan kemampuan suatu model dugaan untuk memprediksi data lain yang sejenis tetapi tidak digunakan dalam membangun model tersebut (data validasi). Penentuan jumlah sumbu komponen utama berdasarkan predictive
success dilakukan dengan validasi silang, yaitu membagi data menjadi dua
kelompok, satu kelompok untuk membangun model dan kelompok lain digunakan untuk validasi (menentukan jumlah kuadrat sisaan). Hal ini dilakukan berulang-ulang pada setiap berulang-ulangan dibangun model dengan berbagai sumbu komponen utama. Jumlah komponen utama yang terbaik adalah yang rataan akar kuadrat tengah sisa (Root Mean Square Predictive Different (RMSPD)) dari data validasi paling kecil (Mattjik dan Sumertajaya 2006).
Metode pengukuran stabilitas hasil seperti Stabilitas Francis dan Kannenberg, Finlay dan Wilkinson, Stabilitas Eberhart dan Russel, dan AMMI (Additive Main Effect Multiplicative) adalah sebagai berikut:
Konsep analisis stabilitas didasarkan pada terbentuknya variasi suatu genotipe dalam berbagai lingkungan. Terbentuknya variasi ini didekati kuadrat tengah genotipe serta koefisien variasi genotipe. Pendekatan tersebut menunjukkan bahwa dengan semakin kecilnya nilai pengukuran, maka semakin stabil genotipe tersebut. Jika nilai CV suatu genotipe kurang dari 20% dikategorikan stabil.
2. Stabilitas Finlay dan Wilkinson (1963)
Ukuran pengaruh lingkungan berasal dari rata-rata produksi dari masing-masing lingkungan dan musim. Regresi didasarkan pada produksi masing-masing-masing-masing varietas di plot terhadap rata-rata populasi. Rata-rata populasi mempunyai koefisien regresi = 1.0 sebagai genotipe yang stabil. Penambahan nilai koefisien terhadap 1.0 berarti meningkatkan kepekaan terhadap lingkungan, dan bila penurunan nilai koefisien terhadap 1.0 berarti meningkatkan ketahanan terhadap lingkungan. Regresi cukup efektif untuk mengetahui respon produksi varietas dalam kisaran lingkungan alami. Batas kisaran lingkungan berkurang akan mengurangi proporsi komponen keragaman bagi interaksi genotipe x lingkungan yang ditunjukkan oleh ragam pada koefisien regresi secara individu.
3. Stabilitas Eberhart dan Russel (1966)
Prinsip stabilitas Eberhart dan Russel yaitu menggabungkan jumlah kuadrat dari lingkungan (E) dan interaksi genotipe x lingkungan (GE) serta membaginya ke dalam pengaruh linier antar lingkungan (derajat bebas = 1) dan pengaruh linier dari genotipe x lingkungan (derajat bebas E = 2). Pengaruh residual kuadrat tengah dari model regresi antar lingkungan digunakan sebagai indeks stabilitas. Genotipe stabil bila memiliki nilai deviasi (simpangan) regresi kuadrat tengah δ2 = 0 dan memiliki nilai koefisien regresi (bi) = 1.
4. AMMI (Additive Main Effect Multiplicative).
Analisis AMMI adalah suatu teknik analisis data percobaan dua faktor perlakuan dengan pengaruh utama perlakuan bersifat aditif sedangkan pengaruh interaksi dimodelkan dengan model bilinier. Model AMMI dapat digunakan untuk menganalisis percobaan lokasi ganda. Pada dasarnya analisis AMMI menggabungkan analisis ragam aditif bagi pengaruh utama perlakuan dengan analisis komponen utama ganda dengan permodelan bilinier bagi pengaruh
interaksi (Mattjik dan Sumertajaya 2006). AMMI sangat efektif menjelaskan interaksi genotipe dengan lingkungan. Penguraian interaksi dilakukan dengan model bilinear, sehingga kesesuaian tempat tumbuh bagi genotipe akan dapat dipetakan. Selain itu biplot yang digunakan memperjelas pemetaan genotipe dan lingkungan secara simultan (Sumertajaya 2007).
Model AMMI sebagai berikut:
Yger = µ + g + βe + 𝜆𝑗 𝜑𝑔𝑗𝜌𝑒𝑗 + 𝛿𝑔𝑒 + εger
dimana:
Yger = nilai pengamatan pada genotipe ke -g, lingkungan ke-e dan kelompok ke-r
µ = rataan umum
g = pengaruh aditif dari pengaruh utama genotipe ke-g
βe = pengaruh aditif dari pengaruh utama lingkungan ke-e
𝜆𝑗 = nilai singular untuk komponen bilinier ke-n
𝜑𝑔𝑗 = pengaruh ganda genotipe ke-g melalui komponen bilinier ke-n 𝜌𝑒𝑗 = pengaruh ganda lokasi ke-e melalui komponen bilinier ke-n 𝛿𝑔𝑒 = simpangan dari pemodelan linier
εger = pengaruh acak pada genotipe ke-g, lokasi ke-e dan kelompok ke-r
Sumertajaya (1998) mengemukakan tiga tujuan penggunaan analisis AMMI yaitu:
1. AMMI sebagai analisis pendahuluan untuk mencari model yang lebih tepat. Jika tidak ada satupun komponen yang nyata maka pemodelan cukup dengan aditif saja. Sebaliknya jika hanya pengaruh ganda saja yang nyata maka pemodelan sepenuhnya ganda, berarti analisis yang tepat adalah komponen utama saja. Jika semua komponen interaksi nyata berarti pengaruh interaksi benar-benar sangat kompleks, tidak memungkinkan dilakukannya pereduksian tanpa kehilangan informasi penting.
2. Menjelaskan interaksi galur x lingkungan. Biplot AMMI meringkas pola hubungan antar galur, antar lingkungan dan antar interaksi galur dan lingkungan.
3. Meningkatkan keakuratan dugaaan respon interaksi galur x lingkungan. Hal ini terlaksana jika hanya sedikit komponen AMMI saja yang nyata dan tidak mencakup seluruh jumlah kuadrat interaksi. Sedikitnya komponen yang nyata menyatakan bahwa jumlah kuadrat sisanya hanya galat saja. Dengan menghilangkan galat ini berarti dugaan respon per galur x lingkungan menjadi lebih akurat.
Metode AMMI sudah dapat diterapkan untuk model tetap (AMMI) yaitu jika genotipe dan lingkungan ditentukan secara subyektif oleh peneliti dan kesimpulan yang diharapkan hanya terbatas pada genotipe dan lingkungan yang dicobakan saja. Pada model campuran (M-AMMI: Mixed AMMI) dimana salah satu sumber keragaman (genotipe atau lingkungan) bersifat acak maka kesimpulan untuk faktor acak hanya berlaku untuk populasi dari faktor acak. Model kategorik (GLM-AMMI/General Linear Model AMMI) yaitu jika respon yang diamati bersifat kategorik seperti tingkat serangan hama (ringan, sedang dan berat). Disamping itu, AMMI juga telah dikembangkan untuk menangani data hilang yaitu dengan EM-AMMI (Expectation Maximitation AMMI) (Sumertajaya 2007).
Waktu dan Tempat
Penelitian dilaksanakan pada musim hujan (MH) 2010/2011 dan 2011/2012 di 7 (tujuh) lokasi yaitu Taman Bogo dan Natar (Lampung); Sukabumi dan Indramayu (Jawa Barat); Purworejo (Jawa Tengah); Wonosari (DIY); Malang (Jawa Timur). Karakteristik masing masing lokasi terdapat pada Lampiran 1.
Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian antara lain: benih padi gogo yang terdiri atas 10 galur dan 2 varietas pembanding (Tabel 4) dengan sejarah persilangan masing-masing galur dan deskripsi varietas pada Lampiran 2 dan Lampiran 3, pupuk kandang/dasar (10 ton ha-1), Urea (200 kg ha-1), SP-36 (100 kg ha-1), KCl (100 kg ha-1) dan pestisida. Alat yang digunakan antara lain: meteran, timbangan, alat penghitung 1000 biji, hand counter, serta alat tulis dan
software analisis SAS 9.0.
Tabel 4 Galur-galur dan varietas pembanding padi gogo yang digunakan dalam penelitian
Kode Genotipe Keterangan Kode Genotipe Keterangan B1 III3-4-6-1 Toleran aluminium B7 IG-19 Toleran naungan B2 I5-10-1-1 Toleran aluminium B8 IG-38 Toleran naungan
B3 WI-44 Toleran naungan B9 IW-56 Toleran naungan
B4 GI-7 Toleran naungan B10 B13-2-e Toleran aluminium
B5 O18-b-1 Toleran aluminium B11 Batu Tegi Varietas cek B6 IW-67 Toleran naungan B12 Way Rarem Varietas cek
Metode Penelitian
Penelitiaan uji daya hasil di tiap lokasi menggunakan Rancangan Kelompok Lengkap Teracak (RKLT) dengan perlakuan genotipe padi gogo. Perlakuan terdiri atas 10 galur dan 2 varietas padi gogo, dengan ulangan sebanyak 4 (empat) dan ulangan tersarang dalam lokasi sehingga terdapat 48 satuan percobaan di tiap lokasi.
Pelaksanaan Penelitian
Tahap awal penelitian dimulai dengan persiapan lahan yang terdiri atas pengukuran luas lahan, pembersihan lahan, pengolahan tanah dan pembuatan petak percobaan. Petak percobaan penelitian dibuat berukuran 4 m x 5 m dengan jarak antar petak dalam ulangan 0.5 m dan antar ulangan 1 m. Total petak percobaan sebanyak 48 petak untuk 4 ulangan di tiap lokasi.
Penanaman menggunakan sistem tugal dengan kedalaman 3-5 cm. Jarak tanam 30 cm x 15 cm, sehingga terdapat 13 baris dan 33 lubang tanam tiap baris sehingga terdapat 429 lubang tanam untuk tiap petak. Setiap lubang berisi 3-5 benih padi gogo.
Aplikasi pemupukan yang dilakukan pada penelitian ini adalah pupuk kandang dan pupuk sumber NPK. Pupuk kandang sebanyak 10 ton ha-1 diberikan bersamaan dengan pengolahan lahan yaitu 1 minggu sebelum tanam. Pupuk kandang disebar dan dicampur dengan tanah. Sumber pupuk NPK yang digunakan adalah Urea (200 kg ha-1), SP-36 (100 kg ha-1) dan KCl (100 kg ha-1). Aplikasi pupuk sumber NPK dilakukan dengan cara memberikannya pada larikan berjarak 5 cm dari tanaman. Waktu aplikasi pupuk sumber NPK dibagi dalam 3 tahap, yaitu:
1. Pemupukan pertama dilakukan 1 minggu setelah penanaman yaitu dengan memberikan Urea (80 gram/petak), SP-36 (200 gram/petak) dan KCl (200 gram/petak),
2. Pemupukan kedua diberikan 4 MST (minggu setelah tanam) Urea (160 gram/petak),
3. Pemupukan ketiga diberikan setelah penyiangan pada 7 MST yaitu Urea (160 gram/petak).
Kegiatan pemeliharaan meliputi penyulaman dan penjarangan yang dilakukan bersamaan pada umur 2 MST. Penyulaman dilakukan dengan sistem sulam pindah. Pengendalian gulma dilakukan selama tanaman berumur 2-7 MST dengan cara penyiangan. Pengendalian hama dan penyakit dilakukan bila perlu hingga menjelang panen. Penyiraman disesuaikan dengan kondisi cuaca dan tanaman. Tahap akhir yaitu pemanenan tanaman yang dilakukan berdasar kriteria
masak fisiologis yang ditandai oleh malai yang berwarna kuning hingga mencapai 80% dalam satu plot.
Pengamatan
Unit pengamatan dilakukan terhadap 5 tanaman sampel dan populasi tiap petak percobaan untuk komponen-komponen tanaman padi gogo, antara lain:
1. Tinggi tanaman, diukur dari permukaan tanah sampai ujung malai tertinggi terhadap 5 rumpun tanaman sampel. Pengukuran tinggi tanaman dilakukan menjelang panen.
2. Jumlah anakan saat vegetatif, dihitung dari jumlah anakan pada saat tanaman berumur 50-60 hari setelah tanam.
3. Jumlah anakan produktif, dihitung berdasarkan jumlah anakan yang menghasilkan malai tiap rumpun. Penghitungan jumlah anakan produktif dilakukan menjelang panen.
4. Panjang malai, diukur dari pangkal malai (leher malai) hingga ujung malai.
5. Umur berbunga, diamati mulai tanam sampai tanaman berbunga 50% dalam tiap petak percobaan.
6. Umur panen, dihitung dari mulai tanam hingga gabah berwarna kuning (masak) telah mencapai 80% dalam tiap petak percobaan.
7. Jumlah gabah total per malai, dihitung dari jumlah gabah dalam tiap malai dari 5 malai utama. Jumlah gabah total per malai berasal dari total gabah isi maupun gabah hampa dalam tiap malai.
8. Jumlah gabah bernas per malai dan gabah hampa per malai, dihitung dari jumlah gabah bernas dan hampa dari 5 malai utama.
9. Persen gabah isi, dihitung menggunakan rumus:
Persen gabah isi = Jumlah gabah isi x 100% Jumlah gabah total
10. Hasil gabah kering per petak, dilakukan pada seluruh malai hasil panen dalam satu petak dikurangi 2 baris keliling (sebagai tanaman sampel dan border). Jumlah rumpun yang dipanen dihitung, kemudian ditimbang gabah kering panen (GKP). Kadar air dihitung berdasarkan nilai rata-rata dari 3 kali pengukuran kadar air gabah hasil panen (GKP). Setelah gabah
dijemur hingga mencapai kadar air + 14% dan dibersihkan, kemudian ditimbang gabah kering giling (GKG) tiap petak.
11. Bobot 1000 butir gabah (gram), ditimbang dari 1000 butir gabah bernas dari setiap petak dengan kadar air ± 14%.
Analisis Data Tahapan Analisis Data
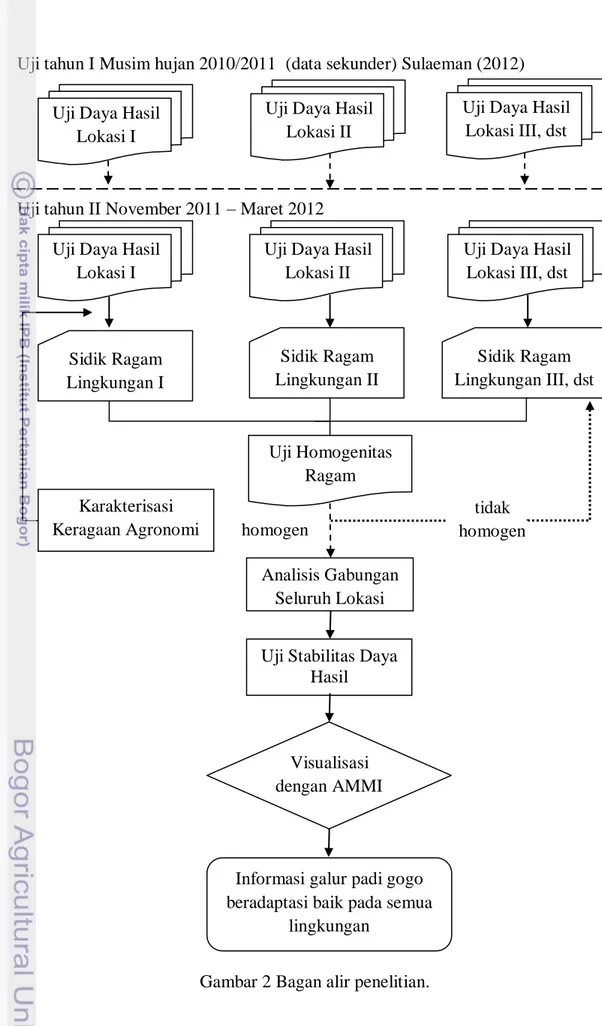
Data pengujian musim 2010/2011 merupakan data sekunder dari penelitian Sulaeman (2012) dan data pengujian musim 2011/2012 merupakan data primer. Bagan alir penelitian dan analisis data disajikan pada Gambar 2.
Uji Normalitas
Uji untuk mengetahui suatu data menyebar normal menurut Mattjik dan Sumertajaya (2006) yaitu uji Lilliefors. Hipotesis yang diuji adalah H0 : populasi
contoh menyebar normal. Statistik ujinya adalah:
D = maksimum 𝑆 𝑥 − 𝐹0(𝑥)
dimana : 𝑆 𝑥 = jumlah amatan contoh x
𝑛
𝐹0(𝑥) = 0.5 – P(0<Z<z); z =(𝑋− 𝜇 ) 𝜎
Analisis Ragam Tiap Lokasi
Analisis ragam dilakukan pada tiap lokasi untuk karakter-karakter padi gogo yang diamati. Analisis ragam berdasarkan metode yang digunakan oleh Singh dan Chaudhary (1979) disajikan pada Tabel 5. Model linier untuk RAK tiap lokasi, sebagai berikut:
Yik = µ + ρk + i + εik
i = 1,2,3, ….,12 dan k = 1,2,3,4 dimana:
Yik = Hasil pengamatan galur dan varietas pembanding ke-i dan ulangan ke-k
µ = Rataan umum
ρk = Pengaruh ulangan ke-k i = Pengaruh perlakuan ke-i
Uji tahun I Musim hujan 2010/2011 (data sekunder) Sulaeman (2012)
Uji tahun II November 2011 – Maret 2012
Gambar 2 Bagan alir penelitian. Sidik Ragam Lingkungan I Uji Homogenitas Ragam Karakterisasi Keragaan Agronomi Uji Daya Hasil
Lokasi I
Uji Daya Hasil Lokasi II
Uji Daya Hasil Lokasi III, dst
Uji Daya Hasil Lokasi I
Uji Daya Hasil Lokasi II
Uji Daya Hasil Lokasi III, dst
Sidik Ragam Lingkungan II
Sidik Ragam Lingkungan III, dst
Uji Stabilitas Daya Hasil Analisis Gabungan
Seluruh Lokasi
Visualisasi dengan AMMI
Informasi galur padi gogo beradaptasi baik pada semua
lingkungan homogen
tidak homogen
Tabel 5 Sidik ragam karakter padi gogo pada masing-masing lokasi uji.
Sumber Keragaman Derajat Bebas
Jumlah
kuadrat Kuadrat Tengah F-Hitung
Ulangan (r-1) JK 3 M3 = JK 3/(r-1) M3/ M1
Genotipe (G) g-1 JK 2 M2 = JK2/(g-1) M2/ M1
Galat (r-1) (g-1) JK 1 M1 = JK1/(r-1) (g-1) -
Total rg-1
Keterangan : r (jumlah ulangan), g (jumlah genotipe)
Uji Kehomogenan Ragam
Uji Bartlett menurut Mattjik dan Sumertajaya (2006) digunakan sebagai pengujian kehomogenan ragam galat. Hipotesis yang diuji adalah H0 : 𝜎12 = 𝜎22 = .... = 𝜎𝑘2. Prosedur pada uji Bartlett ini menggunakan pendekatan khi-kuadrat dengan (k-1) derajat bebas. Statistik ujinya adalah:
χ2= 2.3026 𝑟 𝑖− 1 𝑖 log S2 − 𝑟 𝑖− 1 𝑖 log Si2 dimana: 𝑆𝑖2= (𝑌𝑗 𝑖𝑗 − 𝑌 𝑖.)2 𝑟𝑖 − 1 ; 𝑆2= (𝑛𝑖− 1)𝑆𝑖2 𝑁 − 𝑡
Nilai χ2 dikoreksi sebelum dibandingkan dengan nilai χ 𝛼 ,𝑘−1
2 . Nilai χ2 terkoreksi adalah (1/FK) χ2, dengan FK adalah:
𝐹𝐾 = 1 + 1 3(𝑡 − 1) 1 𝑟𝑖 − 1− 1 𝑟𝑖− 1 𝑖
Analisis Ragam Gabungan
Analisis ragam gabungan (Tabel 6) dilakukan untuk menganalisis hasil pengamatan di semua lingkungan uji untuk karakter padi gogo yang diamati. Model linear untuk RAKL gabungan antara genotipe dan lingkungan seperti dikemukakan oleh Baihaki (2000) sebagai berikut:
Y
ijk= µ + α
i+ β
j/k+ τ
k+ (ατ)
ik+ ε
ijkdimana:
Y
ijk = nilai pengamatan pada perlakuan ke-i, ulangan ke-j, lingkungan ke-kµ
= nilai rata-rata umumα
i=
pengaruh perlakuan ke-iβ
j/k=
pengaruh ulangan ke-j dalam lingkungan ke-kτ
k = pengaruh lingkungan ke-kε
ijk = pengaruh acak pada perlakuan ke-i, ulangan ke-j, lokasi ke-kTabel 6 Sidik ragam gabungan menggunakan model acak
Sumber Keragaman db Kuadrat Tengah Nilai F
Lingkungan (E) L-1 M5 M5/ M4
Ulangan/Lingkungan L(r-1) M4 -
Genotipe (G) g-1 M3 M3/M1
G x E (L-1)(g-1) M2 M2/M1
Galat L(r-1)(g-1) M1 -
Keterangan : L (jumlah lokasi), r (jumlah ulangan), g (jumlah genotipe)
Analisis Stabilitas
Analisis stabilitas dilakukan untuk memperoleh galur-galur yang memiliki stabilitas hasil di semua lokasi uji. Pendugaan parameter kestabilan dilakukan dengan menggunakan empat pendekatan stabilitas hasil yaitu analisis stabilitas menurut Francis dan Kannenberg (1978), analisis stabilitas menurut Finlay dan Wilkinson (1963) analisis stabilitas menurut Eberhart dan Russel (1966) dan analisis AMMI.
a. Analisis stabilitas menurut Francis dan Kannenberg (1978)
Ragam lingkungan (𝑆𝑖2) dan koefisien ragam (CVi) digunakan untuk
menentukan kestabilan suatu genotipe.
CVi =
𝑆𝑖2
𝑌 𝑖𝑜 x 100%
dimana:
CVi = Koefisien variasi genotipe
𝑆𝑖2 = Kuadrat tengah dalam genotipe
Nilai koefisien variasi dari tiap genotipe digunakan untuk menentukkan stabil tidaknya suatu genotipe. Suatu genotipe dikatakan stabil jika nilai koefisien variasi genotipenya kurang dari 25%.
b. Analisis stabilitas Finlay dan Wilkinson (1963)
Analisis stabilitas menurut Finlay dan Wilkinsons menggunakan regresi antara genotipe dengan rataan genotipe di setiap lingkungan dalam skala log. Rata-rata hasil semua genotipe pada tiap lingkungan digunakan sebagai absis, dan hasil tiap genotipe pada tiap lingkungan digunakan sebagai ordinat. Sudut koefisien regresi menunjukkan wilayah adaptabilitas dan stabilitas genotipe.
1. suatu genotipe yang memiliki koefisien regresi b yang lebih besar dari satu dan signifikan menunjukkan bahwa genotipe tersebut beradaptasi baik pada lingkungan subur dengan kata lain adaptif terhadap perubahan lingkungan (Gambar 3),
2. genotipe dengan nilai b yang lebih kecil dari satu tidak sensitif terhadap perubahan lingkungan, karena itu beradaptasi pada lingkungan yang kurang subur (Baihaki 2000).
Spesifik beradaptasi pada lingkungan baik
1.0 kurang stabilitas beradaptasi baik
beradaptasi rata-rata pada semua lingkungan
Spesifik beradaptasi pada lingkungan kurang baik
Rata-rata hasil
Gambar 3 Interpretasi umum nilai b dari pola populasi genotipe ketika koefisien regresi genotipe diplot terhadap nilai rata-rata hasil genotipe (Finlay & Wilkinson 1963)
c. Analisis stabilitas Eberhart dan Russel (1966)
Analisis stabilitas untuk hasil dan komponen hasil menggunakan metode menurut Eberhart dan Russel (1966) dalam Singh dan Chaudhary (1985), dengan model regresi yang digunakan adalah :
Yij = m + βiIj + δij
dimana:
Yij = hasil/komponen hasil rataan dari genotipe ke-i di lingkungan ke-j
m = rataan umum untuk hasil/komponen hasil genotipe ke-i dari semua lingkungan
βi = koefisien regresi, respon genotipe ke-i pada lingkungan berbeda
Ij = indeks lingkungan yaitu rata-rata semua genotipe pada lingkungan ke-j
dikurangi rata-rata seluruh percobaan Ij =
𝑌𝑖 𝑖𝑗
𝑔 − 𝑌𝑖 𝑗 𝑖𝑗
𝑔𝑙
δij = simpangan regresi dari genotipe ke-i pada lingkungan ke-j
Parameter stabilitasnya: 1. Koefisien regresi (bi); bi =
𝑌𝑗 𝑖𝑗𝐼𝑗
𝐼𝑗2 𝑗
2. Simpangan dari regresi δ2; δ2 = δij 2 j l−2 − Se2 r dimana: 𝑆𝑒 2
𝑟 = dugaan galat gabungan
𝛿 𝑖𝑗2 𝑗 = 𝑌𝑖𝑗2 𝑗 −𝑌𝑖 2 𝑔 − ( 𝑌𝑗 𝑖𝑗𝐼𝑗) 2 𝐼𝑗2 𝑗
Genotipe stabil bila memiliki nilai koefisien regresi (bi) = 1 dan memiliki
nilai deviasi (simpangan) regresi kuadrat tengah δ2 = 0. Tabel sidik ragam analisis stabilitas menurut Eberhart dan Russel (1966) disajikan pada Tabel 7.
d. Analisis AMMI
Analisis AMMI merupakan teknik analisis data percobaan dua faktor perlakuan dengan pengaruh utama perlakuan bersifat aditif sedangkan pengaruh interaksi dimodelkan dengan model bilinier. Prinsipnya analisis AMMI menggabungkan analisis ragam aditif bagi pengaruh utama perlakuan dengan analisis komponen utama ganda dengan permodelan bilinier bagi pengaruh interaksi (Mattjik dan Sumertajaya 2006). AMMI sangat efektif menjelaskan
interaksi genotipe dengan lingkungan. Biplot digunakan untuk memperjelas pemetaan genotipe dan lingkungan secara simultan (Sumertajaya 2007).
Tabel 7 Sidik ragam analisis stabilitas Eberhart dan Russel
Sumber Keragaman Derajat Bebas Kuadrat Tengah
Galur (G) g – 1 𝑌𝑖 𝑖..2 𝑛 − 𝐹𝐾 Interaksi G x L g (L – 1) 𝑌𝑖𝑗2− 𝑌𝑖2 𝑖 𝑛 𝑗 𝑖 Lingkungan (linier) L 𝑌𝑗 .𝑗𝐼𝑗 2 𝑟𝑣 𝐼𝑗2 𝑗 Interaksi G x L (linier) g – 1 𝑌𝑖 𝑖𝑗𝐼𝑗 2 𝐼𝑗2 𝑗 𝑖 − 𝐽𝐾 𝑙𝑖𝑛𝑔𝑘. (𝑙𝑖𝑛𝑖𝑒𝑟) Simpangan gabungan g (L – 2) 𝛿𝑖𝑗2 𝑗 𝑖 Galur 1 L – 2 𝑌𝑖𝑗2− 𝑌𝑖2 𝑛 𝑗 − 𝑌𝑗 𝑖𝑗𝐼𝑗 2 𝐼𝑗2 𝑗 Galur 2 L – 2 ⋮ Galur 12 L – 2 𝑌𝑔𝑗2 − 𝑌𝑔2 𝑛 𝑗 − 𝑌𝑗 𝑔𝑗𝐼𝑗 2 𝐼𝑗2 𝑗 Galat gabungan L (g – 1) (r – 1) Total g L – 1 Yij2− FK j i
Keterangan: L (jumlah lokasi), r (jumlah ulangan), g (jumlah genotipe)
Pemodelan bilinier pengaruh interaksi genotipe dengan lingkungan sebagai berikut:
Menyusun pengaruh interaksi dalam bentuk matriks genotipe (baris)* lingkungan (kolom) sehingga matriks berukuran a x b:
𝛾 =
𝛾11 … 𝛾1𝑏
… … …
𝛾𝑎1 … 𝛾𝑎𝑏
Menguraikan bilinier terhadap matriks pengaruh interaksi 𝛾𝑔𝑒 = 𝜆𝑗
𝑛
𝑗 =1
𝜑𝑔𝑗𝜌𝑒𝑗 + 𝛿𝑔𝑒
Model AMMI secara lengkap dapat ditulis sebagai berikut dan model analisis ragam AMMI (Tabel 8):
Yger = µ + g + βe + 𝝀𝒋𝝋𝒈𝒋𝝆𝒆𝒋+ 𝜹𝒈𝒆 + εger
dimana:
Yger = nilai pengamatan pada genotipe ke -g, lingkungan ke-e dan kelompok ke-r
µ = rataan umum
g = pengaruh aditif dari pengaruh utama genotipe ke-g
βe = pengaruh aditif dari pengaruh utama lingkungan ke-e
𝜆𝑗 = nilai singular untuk komponen bilinier ke-n
𝜑𝑔𝑗 = pengaruh ganda genotipe ke-g melalui komponen bilinier ke-n 𝜌𝑒𝑗 = pengaruh ganda lokasi ke-e melalui komponen bilinier ke-n 𝛿𝑔𝑒 = simpangan dari pemodelan linier
εger = pengaruh acak pada genotipe ke-g, lokasi ke-e dan kelompok ke-r
Tabel 8 Model analisis ragam AMMI
Sumber Keragaman Derajat Bebas Kuadrat
Tengah Nilai F
Lingkungan (L) L-1 KTL KTL/ KTGalat
Ulangan /Lingkungan L(r-1) KTr/L KTr/L/KTGalat
Genotipe (G) G-1 KTG KTG/KTGalat
G x L (L-1)(G-1) KTG*L KTG*L/KTGalat
IAKU1 G+L-1-(2x1) KTIAKU1 KTIAKU1/ KTGalat
IAKU2 G+L-1-(2x2) KTIAKU KTIAKU2/ KTGalat
IAKUn g+L-1-(2xn) KTIAKUn KTIAKU1n/ KTGalat
Galat L(r-1)(G-1) KTGalat -
Total G L r-1
Keterangan: L (lingkungan), G (genotipe), r (ulangan), IAKU (Interaksi Analisis Komponen Utama)
Kondisi Umum Penelitian
Kondisi umum penelitian cukup baik. Karakteristik lingkungan pengujian berbeda-beda antar lokasi. Kondisi tanah di Taman Bogo pada musim uji pertama memiliki kejenuhan Al agak tinggi pada ulangan I sehingga tanaman kurang tumbuh secara optimal (Sulaeman 2012). Kondisi tanah di Taman Bogo pada musim kedua lebih baik dibanding musim pertama. Lingkungan tumbuh di Natar musim pertama dan kedua merupakan lahan kering tegalan. Karakteristik lahan lingkungan Indramayu pada musim pertama merupakan lahan di bawah tegakan pohon jati. Lingkungan penanaman Indramayu pada musim kedua merupakan lahan kering dengan kondisi tidak rata. Lokasi Sukabumi musim pertama dan kedua merupakan lahan sawah yang dikeringkan. Lokasi Purworejo musim pertama dan kedua merupakan lahan kering di dataran rendah dengan tekstur tanah sedikit berpasir. Lokasi pengujian Wonosari merupakan lahan kering berteras dataran rendah dan sedikit berbatu. Lokasi Malang musim pertama merupakan lahan kering dikelilingi pohon dan musim kedua merupakan lahan sawah dikeringkan dan terletak di dataran agak tinggi. Karakteristik umum lingkungan pengujian disajikan pada Lampiran 4.
Pengujian musim pertama dilaksanakan selama bulan Oktober 2010 hingga April 2011 (Sulaeman 2012). Pengujian musim kedua dilaksanakan bulan November 2011 hingga Maret 2012 kecuali lokasi Malang yang dilaksanakan bulan Maret hingga Juli 2011. Pertumbuhan awal tanaman secara umum baik di semua lokasi karena ketersediaan air yang cukup. Curah hujan yang cukup mendukung pertumbuhan vegetatif tanaman di semua lokasi penanaman. Data klimatologi lingkungan pengujian disajikan pada Lampiran 5.
Hama mentul (Phillophaga helleri) muncul di Wonosari pada musim pertama maupun kedua, tetapi penyebarannya dapat dicegah sehingga tidak sampai menyebabkan kerusakan tanaman pada fase pertumbuhannya. Serangan blas daun (Pyricularia grisea pv. oryzae) dan Hawar Daun Bakteri (Xanthomonas
oryzae pv. oryzae) terjadi di Sukabumi musim pertama dan kedua, Indramayu,
Sukabumi merupakan daerah endemik blas daun. Serangan ini masih dapat ditanggulangi dengan pengendalian penyakit secara intensif sehingga tidak menimbulkan kerusakan yang parah.
Serangan walang sangit (Leptocorisa oratorius) terjadi pada saat muncul malai sampai bulir padi matang susu. Bulir padi yang dihisap cairannya oleh walang sangit menyebabkan gabah menjadi hampa dan berubah warna. Serangan walang sangit terjadi di Natar, Indramayu dan Sukabumi dengan tingkat serangan rendah. Pengendalian intensif dengan aplikasi insektisida efektif mengendalikan serangan walang sangit.
Serangan burung terjadi pada fase generatif sampai menjelang panen. Lokasi Sukabumi, Purworejo, dan Malang musim pertama menunjukkan serangan burung sejak awal pengisian biji. Hal ini disebabkan karena ketidaksamaan waktu tanam dengan areal pertanaman sekitar dan umur genotipe yang diuji lebih genjah, sehingga serangan burung terkonsentrasi pada satu tempat tertentu. Rata-rata kehilangan hasil akibat serangan burung di lingkungan Malang musim pertama mencapai 30% untuk varietas berumur panjang (Sulaeman 2012). Serangan burung hampir tidak terjadi di semua lokasi pengujian musim kedua. Serangan burung yang cukup terlihat hanya pada lokasi Malang dengan tingkat serangan rendah.
Analisis Stabilitas Hasil Selama Dua Musim Tanam
Penelitian stabilitas musim tanam pertama sudah dilakukan oleh Sulaeman (2012). Data produktivitas GKG musim pertama dan musim kedua dikompilasi untuk kemudian dilakukan analisis stabilitas hasil. Sidik ragam gabungan karakter produktivitas GKG dari tujuh lokasi pengujian musim pertama dan kedua akan ditampilkan dengan asumsi perbedaan musim tanam dianggap sebagai lingkungan yang berbeda. Hal tersebut menjadikan lingkungan pengujian menjadi 14 lingkungan. Sidik ragam gabungan 14 lingkungan diperlihatkan pada Tabel 9.