Makalah Mata Kuliah Seminar Killer Yeast di Daerah Tropis
Disusun oleh Marta Diana Kuncoro
31101236
Fakultas Bioteknologi
Halaman Pengesahan
Naskah Seminar dengan judul : Killer Yeast di Daerah Tropis
merupakan hasil olahan jurnal-jurnal berikut :
1. Tan, H.W. & Tay, S.T. (2010). Anti-Candida activity and biofilm inhibitory effects of secreted products of tropical environmental yeasts. Tropical Biomedicine 28(1): 175 180.
2. Abranches, J., Morais, P.B., Rosa, C.A., Mendonca-Hagler, L.C., & Hagler, A.N. (1997). The incidence of killer activity and extracellular proteases in tropical yeast
communities. Can.J. Microbiol. 43: 328-336.
3. Lim, S.L. & Tay, S.T. (2011). Diversity and killer activity of yeasts in Malaysian fermented food samples. Tropical Biomedicine 28(2): 438-443.
disusun oleh Marta Diana Kuncoro
31101236
disajikan dalam matakuliah Seminar
Disetujui, Pembimbing I,
ABSTRAK
Killer yeast adalah strain yeast tertentu yang dapat memproduksi protein toksin bersifat
lethal bagi strain yeast sensitif. Killer yeast berperan dalam pembentukan struktur komunitas yeast. Pada habitat tropis, killer yeast ditemukan di buah, bunga, daun, kaktus, bromelia,
Drosophila, serta berbagai macam makanan fermentasi. Ditemukan beberapa genus yang bersifat
lethal terhadap Candida, antara lain Aureobasidium, Cryptococcus, Issatchenkia, Kloeckera apis,
Kluyveromices, Pichia, Pseudozyma, Trichosporon, dan Ustilago. Beberapa diantara genus
tersebut memiliki kemampuan menghambat aktivitas metabolisme biofilm Candida albicans. Aktivitas proteolitik pada komunitas yeast yang ditemukan pada berbagai habitat menggambarkan kekayaan nutrien pada habitat tersebut dan mungkin penting bagi keberadaan atau kompetisi yeast terkait.
ABSTRACT
Killer yeasts are yeasts that secrete proteinaceous killer toxins lethal to sensitive yeast strains. Killer yeast plays a role in the formation of community structure. In tropical habitats, killer yeasts could be found on fruits, flowers, leaves, cacti, bromeliad, Drosophila and various fermented foods. Among these interesting yeast, there are several genera that are lethal to
Candida, including Aerobasidium, Cryptococcus, Issatchenkia, Kloeckera, Kluyveromices,
Pichia, Pseudozyma, Trichosporon, and Ustilago. Some of theme has the ability to inhibit
Candida albicans biofilm. Proteolytic activities were also found for yeast communities reflecting
BAB 1 PENDAHULUAN
Yeast atau khamir adalah mikroorganisme eukariotik bersel tunggal yang termasuk dalam
kerajaan Fungi. Yeast telah dimanfaatkan oleh manusia sejak zaman dahulu, bahkan sebelum bahasa tulisan berkembang. Oleh karena itu, yeast dikenal sebagai “Man’s oldest industrial company” dalam bidang pangan, khususnya berperan sebagai mikroorganisme yang memfermentasi jus anggur menjadi wine dan berperan pula dalam proses pembuatan roti dimana karbondioksida yang dihasilkan membuat roti dapat mengembang.
Seiring dengan perkembangan pengetahuan, saat ini telah ditemukan kelompok yeast yang disebut dengan killer yeast. Killer yeast adalah strain yeast tertentu yang dapat memproduksi protein toksin bersifat lethal bagi strain yeast sensitif. Banyak penelitian membuktikan bahwa protein toksin memiliki aktivitas anti-mycotic untuk menghambat pertumbuhan strain yeast sensitif.
Sama seperti yeast lainnya, killer yeast secara alami dapat ditemukan pada berbagai habitat. Pada habitat alami, killer yeast dapat ditemukan di tanah, tumbuhan dan serangga(1,2). Selain itu, dapat juga ditemukan di dalam makanan, antara lain tape, tempe, wine, cuka, miso, fermentasi buah dan sayuran(3). Dalam habitat tersebut, yeast, khususnya killer yeast memiliki peran yang unik, baik itu dapat merugikan atau membawa manfaat bagi manusia. Aktivitas proteolitik penting bagi yeast untuk beradaptasi di dalam habitatnya. Aktivitas proteolitik menjadi sarana yeast untuk mengambil nutrien dalam substrat. Sehingga tidak dapat dipungkiri bahwa kemampuan killer dan kemampuan proteolitik yeast berperan dalam kompetisi di dalam suatu habitat.
Killer yeast dan toksin yang diproduksinya, secara potensial dapat diaplikasikan baik
dalam bidang lingkungan, kesehatan maupun industri. Dalam bidang industri fermentasi atau pengawetan makanan, Sulo et al. (1992) menyatakan kehadiran killer yeast dapat mengeliminasi mikroorganisme yang tidak diinginkan. Dalam bidang kesehatan, killer yeast dimungkinkan dapat menghambat atau membunuh mikroorganisme patogen bagi manusia, contohnya
BAB 2
TINJAUAN PUSTAKA
2.1 Ekologi Yeast
Yeast adalah organisme eukariotik bersel tunggal yang termasuk dalam kerajaan
Fungi. Yeast biasanya tumbuh di lingkungan lembab yang memiliki nutrisi sederhana,
seperti gula dan asam amino. Hal tersebut juga yang menjadi alasan mengapa yeast secara umum dapat ditemui pada tumbuhan (daun, bunga, buah akar), serangga, dan juga berbagai jenis makanan(5). Sebuah studi menyatakan bahwa buah yang rusak atau membusuk merupakan habitat yang baik bagi pertumbuhan yeast karena kondisi keasamannya, faktor pendukung suksesi dan intensitas yang tinggi akan hadirnya vektor serangga(2).
Kemampuan dari banyak yeast untuk mengatur komunitasnya sangat berpengaruh pada pertumbuhan dan kelangsungan hidupnya. Selama beberapa tahun terakhir, studi mengenai Saccharomyces cerevisiae mengungkapkan beberapa karakter dari komunitas
yeast. Sementara itu, matriks ekstraseluler di sekeliling komunitas dapat melindungi
mereka dari stress lingkungan dan berperan dalam pengeluaran sinyal oleh sel (cell
signaling) yang berkontribusi terhadap organisasi dalam komunitas, sehingga lingkup
komunitas yang berbeda akan mengekspresikan gen dan sifat organisme yang berbeda(11). Peran yeast di dalam kehidupan serangga telah banyak dipelajari, salah satunya adalah hubungan yeast dengan lalat buah (Drosophila). Pada studi tersebut menyatakan bahwa yeast mampu mensintesis accessory substance (nutrisi tambahan) yang dibutuhkan oleh Drosophila (pada buah)(6). Dari hasil studi ini, akhirnya yeast diaplikasikan untuk menjebak Drosophila suzukii yang menjadi hama buah cherry karena adanya kecenderungan jenis lalat buah tersebut bertelur pada buah yang terdapat yeast di dalamnya (berhubungan dengan accessory substance yang dibutuhkan untuk nutrisi larva)(12).
Yeast tertentu mengambil peran utama dalam proses fermentasi makanan,
suatu jenis yeast menjadi indikator adanya yeast lain dalam lingkungan yang sama karena
yeast selalu ditemukan dalam komunitas. Yeast-yeast tersebut dapat mendukung maupun
menghambat proses fermentasi makanan(3).
2.2 Killer Yeast
Killer yeast adalah strain yeast yang memiliki kemampuan untuk mensekresi
protein atau glikoprotein toksin (killer toxin) yang bersifat lethal bagi strain yeast tertentu (sensitive yeast). Penemuan killer toxin tersebut pertama kali dilaporkan oleh Bevan dan Makover (1963) pada strain yeast Saccharomyces cerevisiae. Berdasarkan penemuan itu, maka dalam suatu komunitas, yeast dikelompokkan menjadi killer, neutral dan sensitive. Strain yeast dikatakan netral apabila tidak memproduksi killer toxin tetapi juga tidak sensitif terhadap strain yeast lain. Sedangkan, dinyatakan sebagai killer apabila toksin yang dihasilkan (killer toxin) bersifat lethal terhadap strain yeast tertentu yang kemudian disebut sensitive yeast. Setelah itu, pada tahun 1970-an, ditemukan bahwa kemampuan
Saccharomyces cerevisiae untuk memproduksi killer toxin berhubungan dengan
keberadaan suatu virus double stranded RNA (dsRNA)(4). Seiring dengan perkembangan penelitian yang dilakukan, ditemukan spesies yeast lain (selain S. cerevisiae) yang memiliki killer toxin, contohnya beberapa spesies dari genus Pichia, Kluyveromyces,
Debaryomyces, Ustilago, Pseudozyma, dan Candida(1,2,3).
Kini, diketahui bahwa killer toxin yang diproduksi oleh spesies yeast yang berbeda, ada yang dikode oleh plasmid DNA maupun RNA. Dengan perbedaan pengkode pada masing-masing killer toxin tersebut, maka targetnya pun berbeda, baik jenis strain
yeast sensitifnya maupun reseptor yang diserang(7).
2.3 Killer Toxin
Jenis killer toxin dari dsRNA-killer yeast yang sering dipelajari yaitu toksin pada
Saccharomyces cerevisiae. Killer yeast ini memiliki killer toxin K1, K2 dan K28 yang
dikode oleh jenis virus ScV-M1, ScV-M2, ScV-M28 yang merupakan virus dsRNA sejenis
mycovirus yang terus berkembang pada yeast dan fungi. Pada Saccharomyces cerevisiae ini,
mengkode enkapsidasi, pembentukan struktur partikel virus dan sebagai RNA polymerase. Sedangkan, M virus merupakan pengkode toksin sekaligus memberikan imunitas kepada
Saccharomyces cerevisiae terhadap toksin tersebut(4).
Pada beberapa strain killer lainnya, killer toxin yang dimiliki dikode oleh dsDNA plasmid. Salah satu contohnya adalah strain killer Kluyveromyces lactis IFO1267 yang memiliki plasmid pGKL1 dan pGKL2(8). Hasil studi berbagai macam killer toxin menunjukkan bahwa karakteristik umum killer toxin hampir semuanya merupakan protein yang sensitif terhadap protease, labil pada temperatur tinggi dan aktif hanya pada pH rendah (asam)(9).
2.5 Killer Yeast dan Habitatnya
Sama seperti yeast pada umumnya, killer yeast dapat hidup di alam (tumbuhan, tanah, serangga)(1,2), maupun pada makanan fermentasi (tape, tempe, wine, cuka, miso, fermentasi buah dan sayuran)(3). Sejauh ini belum diketahui pasti fungsi killer toxin di komunitas alami yeast dan spekulasi mengenai peran killer toxin(10).
Buah merupakan habitat yang optimal bagi aktivitas killer toxin, yaitu memiliki pH rendah (asam) dan kandungan gula tinggi sehingga tidak mengherankan apabila komunitas killer yeast pada buah memiliki proporsi yang tinggi(10). Pada buah yang membusuk, killer yeast dapat lebih unggul karena pada lingkungannya yang sesuai tersebut, keuntungan yang akan didapat lewat sekresi killer toxin lebih besar daripada energi yang diperlukan untuk proses sekresi toksin. Sementara itu, misalnya pada kaktus busuk dan batang pohon lapuk yang bukan merupakan habitat ideal bagi aktivitas killer
toxin, energi yang dikeluarkan untuk mensekresi killer toxin lebih besar daripada
keuntungan yang didapatkan, sehingga fenotip killer di habitat semacam ini mengalami kerugian. Dalam keadaan ini, kompetisi yang bersifat eksploitatif lebih kerap terjadi(10).
Killer yeast dapat ditemukan lebih banyak pada habitat alami daripada dalam koleksi
BAB 3 ISI
3.1 Killer Yeast pada Sampel Makanan
Killer yeast dapat diisolasi dari berbagai sampel makanan. Berikut adalah contoh
killer yeast yang berhasil diisolasi dan diidentifikasi dari sampel makanan.
Tabel 1. Killer Yeast pada Sampel Makanan yang Ditemukan di Malaysia
Di Malaysia (pasar Klang valley) diambil sampel makanan fermentasi antara lain tape, tempe, kecap, rice wine, yogurt, cuka, fermentasi kacang dan miso, serta fermentasi buah dan sayuran. Dari total 19 spesies yeast yang diisolasi dari makanan tersebut, ditemukan 4 spesies killer yeast yang bersifat lethal terhadap beberapa strain Candida (C.
albicans ATCC90028, C. parapsilosis ATCC22019, Candida krusei ATCC6258, dan
isolate klinis C. dubliniensis, C. tropicalis, C. glabrata, C. guilliermondii, C. rugosa), yaitu Trichosporon asahii, Pichia anomala, Pichia norvegensis dan Issatchenkia
orientalis.
Berdasarkan hasil uji aktivitas anti-Candida, Trichosporon asahii yang diisolasi dari tempe memiliki aktivitas killer yang paling luas terhadap berbagai jenis Candida, yaitu C. tropicalis, C. guilliermondii, C. rugosa, C. glabrata, C. parapsilosis
Trichosporon pullulans, Trichosporon asteroids dan Trichosporon porosum (Lim, S.L.
dan Tay, S.T., 2011)
Studi ini menunjukkan adanya aktivitas killer pada C. dubliniensis, C. tropicalis,
C.guilliermondii dan C. rugosa. Pichia norvegensis sebelumnya dilaporkan memiliki
spektrum killer yang luas terhadap berbagai jenis fungi patogen oportunistik, seperti C. albicans. Sementara itu, pada studi ini Pichia norvegensis menunjukkan killer activity pada C. glabrata, C. guilliermondii, C. rugosa. Sedangkan Issatchenkia orientalis menunjukkan killer activity hanya pada C. glabrata, dan C. krusei ATCC6258.
3.2 Killer Yeast pada Sampel Bunga, Daun dan Tanah (Malaysia)
Penelitian dilakukan dengan mengisolasi yeast dari bunga, daun dan tanah kawasan kampus University of Malaya, Kuala Lumpur. Dari total 100 sampel bunga, daun dan tanah, didapat 4 genus yeast yang dinyatakan sebagai killer yeast, yaitu Aureobasidium pullulans,
Ustilago (U. sparsa, U. trichophora, U. tragana), Candida (C. parapsilosis, C. tropicalis)
dan Pseudozyma (P. hubeiensis, P. spp.). Dalam studi ini, Aureobasidium pullulans dinyatakan sebagai isolat yang paling banyak ditemukan (50% dari total isolat teridentifikasi) dan merupakan killer yeast yang paling unggul. Studi lain menyatakan bahwa yeast ini mudah ditemukan di permukaan (phyllosphere) tumbuhan dan bermacam-macam buah tropis (Lotrakul et al., 2009). Organisme ini diketahui memproduksi antifungal aureobasidin A yang memiliki aktivitas fungisidal yang kuat.
Pada studi lain, genus Ustilago yang teridentifikasi sebagai killer yeast hanya spesies
Ustilago maydis. Namun, pada studi ini untuk pertama kalinya dilaporkan U. sparsa, U.
trichophora, U. tragana memiliki aktivitas killer.
Sensitive yeast yang digunakan dalam uji aktivitas killer ini, antara lain C. albicans
ATCC90028, C. parapsilosis ATCC22019, C. parapsilosis ATCC90018, C. krusei ATCC6258 dan isolat klinis C. dubliniensis, C. tropicalis, C. glabrata, C. guilliermondii dan
C. rugosa. Dari semua sensitive yeast yang digunakan untuk uji aktivitas killer, C. rugosa
Killer toxin yang disekresi oleh killer yeast, diuji aktivitas penghambatannya terhadap
biofilm Candida albicans ATCC90028. Pengujian dilakukan dengan cara menambahkan filtrat kultur (killer yeast) ke dalam medium berisi biofilm C. albicans. Efek penghambatan biofilm ditentukan dari persentase reduksi aktivitas metabolik dari biofilm tersebut.
Uji tersebut menunjukkan bahwa Aureobasidium pullulans, Candida tropicalis,
Ustilago sparsa, Ustilago tragana dan Pseudozyma spp. dapat menghambat pertumbuhan
biofilm Candida albicans dengan tingkat penurunan aktivitas lebih dari 50%. Isolat
Pseudozyma menunjukkan presentase penurunan aktivitas metabolism biofilm Candida
tertinggi yaitu 71,9% (gambar 1). Ada banyak faktor yang memengaruhi formasi biofilm C.
albicans. Diperkirakan bahwa kandungan produk alkohol seperti ethanol, isoamyl alcohol,
2-phenylethanol, 1-dodecanol, E-nerolidol, glycolipid biosurfactant dan signaling molecules
yang ditemukan pada kultur filtrat killer yeast, dapat memengaruhi pertumbuhan biofilm C.
albicans.
3.2 Killer Yeast pada Bunga, Buah, Bromelia, Kaktus dan Serangga
Pada studi ini, yeast diisolasi dari buah Parahancornia amapa dan lalat Drosophila terkait di hutan hujan Brazilian Amazon, lalat Drosophila di hutan hujan Atlantis, kaktus
Pilosocereus arabidae, kaktus Opuntia vulgaris dan Drosophila serido terkait, air dari
kantung bromelia Neoregelia cruenta, serta bunga Ipomea litoralis dan Ipomoea pes-caprae dari ekosistem pasir pantai Rio de Janeiro dan Espirito Santo. Semua isolat diuji sifat
nya terhadap Candida glabrata IMUFRT-50083 (NCYC 388).Yeast yang dinyatakan sebagai killer kemudian diuji aktivitas killer-nya terhadap yeast lain dalam satu komunitas.
Killer yeast ditemukan pada semua habitat kecuali pada bunga Ipomea dan air dari
kantung bromelia N. cruenta. Semua killer yeast diuji aktivitas killer-nya terhadap yeast dalam satu komunitas. Killer yeast yang ditemukan, antara lain Candida apis dan
Kluyveromyces marxianus pada lalat Drosophila di hutan hujan Atlantis; Candida krusei,
Candida sorbosa, Hanseniaspora uvarum, Issatchenkia occidentalis, dan Pichia
membranaefaciens dari buah amapa dan Drosophila terkait; yeast yang menyerupai Pichia
ohmeri dan Sporobolomyces roseus dari kaktus; Candida bombicola, Candida fructus, dan
Kloeckera apis dari komunitas amapa dan hutan hujan Atlantis; Pichia kluyveri var. kluyveri
dari ketiga komunitas, yaitu buah amapa, kaktus dan lalat hutan. Komunitas yeast dari buah amapa memiliki diversitas killer yeast paling tinggi.
Studi pada buah Amapa menunjukkan Pichia kluyveri var. kluyveri termasuk killer
yeast yang memiliki aktivitas killer paling tinggi. Pichia kluyveri var. kluyveri
IMUFRJ-51498 memiliki spektrum aktivitas killer paling luas dan membunuh 38% dari isolat yang mewakili 46 dari spesies yang diuji dari buah amapa.
Candida sorboxyla menjadi yeast yang paling sensitif (ditemukan pada buah amapa)
dari semua yeast. Sementara itu, hampir semua Candida citrea, Candida fructus, Candida
krusei, Candida norvegensis, Candida sorbosa, lssatchenkia occidentalis, Kloeckera
apiculata, Kloeckera apis, Pichia acaciae, Pichia kluyveri, Pichia membranaefaciens, dan
Pichia pijperi bersifat sensitif terhadap toksin Pichia kluyveri var. kluyveri.
Dalam komunitas yang ditemukan pada Drosophila dari hutan hujan Atlantis, hampir semua yeast yang ditemukan bersifat resisten terhadap semua isolat killer Pichia kluyveri var.
kluyveri, Kloeckera apis, Candida fructus, Candida bombicola, Candida apis, dan
Kluyveromyces marxianus. Sementara itu, komunitas yeast pada Pilosocereus arabidae,
Opuntia vulgaris dan Drosophila serido memiliki diversitas paling rendah. Hampir semua
spesies pada komunitas ini memiliki isolat sensitif. Di dalam komunitas ini, Candida
guilliermondii hanya dapat dibunuh (sensitif) oleh Pichia kluyveri var. kluyveri.
mungkin berhubungan dengan kenyataan bahwa yeast tersebut diisolasi dari kaktus O. vulgaris dan diuji aktivitas killer-nya terhadap sensitive yeast dari komunitas lain (Pilocereus arabidae). Selain itu, bukti lainnya adalah fenomena M. mucilagina yang dibunuh oleh Pichia
ohmeri yang berasal dari kaktus tetapi tidak dapat dibunuh oleh Pichia kluyveri var. kluyveri
dari kaktus yang sama.
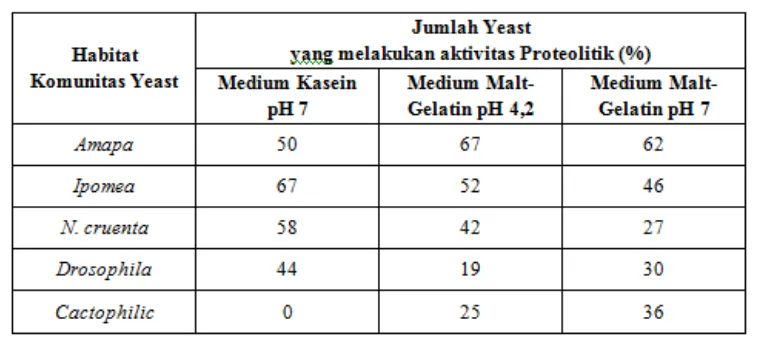
3.3 Aktivitas Proteolitik
Dalam studi yang dibahas dalam sub bab 3.3, semua yeast yang diisolasi dan diidentifikasi, diuji aktivitas proteolitiknya di dalam dua jenis medium, yaitu medium yang kaya protein (kasein) dan kaya karbohidrat (gelatin). Uji aktivitas proteolitik iini dilakukan dengan maksud untuk mengetahui apakah ada hubugan antara resistensi sensitive yeast terhadap killer toxin. Aktivitas proteolitik (produksi protease) tersebar di dalam komunitas
yeast. Berdasarkan uji protease pada komunitas yeast Amapa, 67% spesies positif di dalam
malt-gelatin pada pH 4,2; sebanyak 62% spesies positif di dalam malt-gelatin pada pH 7; dan
50% spesies mampu menghidrolisais kasein pada pH 7. Isolat killer lebih banyak dapat menghidrolisis kasein daripada gelatin dan hanya Pichia kluyveri var. kluyveri yang dinyatakan proteolitik dari 3 uji tersebut.
Kelompok yeast yang diisolasi dari bunga Ipomea dan kantung N. cruenta menunjukkan frekuensi strain proteolitik yang tinggi (64% spesies positif terhadap sedikitnya terhadap satu jenis uji protease). Dari hasil uji yang positif dari komunitas yeast pada Ipomea, kebanyakan spesies dapat menghidrolisis kasein (67% spesies positif dalam medium kasein; 52% spesies positif dalam medium malt-gelatin pH 4,2; 46% spesies positif dalam medium malt-gelatin pH 7). Sementara itu, pada kantung air bromelia N. cruenta, 58% positif dalam medium kasein, 42% spesies positif dalam medium malt-gelatin pH 4,2; dan 27% spesies positif dalam medium malt-gelatin pH 7.
positif dalam medium malt-gelatin pH 4,2; 36% spesies positif dalam medium malt-gelatin pH 7).
Tabel 2. Jumlah Yeast yang melakukan Aktivitas Proteolitik di Berbagai Habitat
Dalam studi ini, tidak semua uji mendeteksi semua proteolityc yeast sehingga diperlukan adanya uji lanjutan. Studi sebelumnya menyatakan bahwa produksi protease ekstraseluler merupakan salah satu faktor dimana killer yeast dapat hidup berdampingan dengan sensitive
yeast (coexist). Protease ekstraseluler diklaim mampu melindungi isolat sensitif dengan cara
memotong atau memecah toksin. Walaupun begitu, data statistik tidak menunjukkan adanya korelasi yang signifikan antara resistensi terhadap killer toxin dengan produksi protease ekstraseluler.
yeast pada Drosophila memiliki diversitas yang tinggi tetapi frekuensinya rendah karena
Drosophila mendatangi substrat yang berbeda-beda.
3.5 Dinamika Populasi Killer Yeast dan Perannya di dalam Habitat
Keberadaan killer yeast berbeda pada masing-masing jenis habitat (baik di alam maupun makanan). Di alam, diversitas killer yeast terbanyak ditemukan pada buah. Studi ini sejalan dengan J. Pintar dan Starmer (2003) yang menyatakan bahwa pada buah yang membusuk,
killer yeast dapat lebih unggul karena memiliki kondisi yang optimal untuk aktivitas killer
toxin. Di lain sisi, studi tersebut juga menunjukkan bahwa aktivitas produksi dari faktor killer
menjadi indikator dari kompetisi dan suksesi yeast dalam suatu fermentasi alami. Oleh karena itu, killer yeast dinyatakan sebagai salah satu faktor yang bertanggungjawab terhadap hilangnya strain sensitive selama suksesi yang terjadi pada buah. Selain itu, diversitas yang tinggi juga berhubungan dengan peran serangga (lalat). Serangga memiliki kebiasaan berpindah-pindah substrat dan membawa berbagai yeast dari satu substrat ke substrat lain.
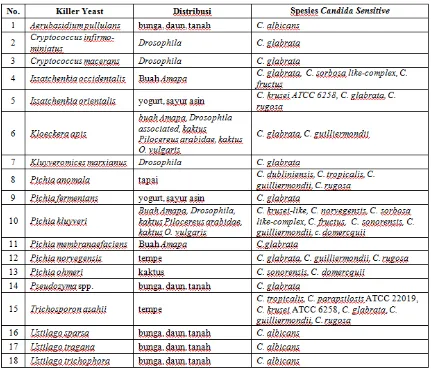
Tabel 3. Beberapa Killer Yeast yang memiliki Daya Anti-Candida(*)
*Keterangan : Data diolah dari Tan, H.W. & Tay, S.T (2010 dan 2011) dan Abranches, J. et al. (1997)
yeast pada wine memengaruhi populasi Saccharomyces yang menjadi yeast utama dalam
proses fermentasi. Namun begitu, killer yeast dalam proporsi tertentu dapat menambah citarasa pada wine karena killer yeast ternyata juga memengaruhi komposisi senyawa yang ada di dalam wine(13).
Berbagai killer yeast pada berbagai habitat di daerah tropis memiliki aktivitas
anti-Candida. Candida dikenal sebagai organisme patogen yang dapat menyebabkan infeksi pada
KESIMPULAN
Killer yeast di daerah tropis dapat ditemukan buah, bunga, daun, kaktus, bromelia,
Drosophila, serta pada makanan fermentasi (tape, tempe, kecap, rice wine, yogurt, cuka,
fermentasi kacang dan miso, seta fermentasi buah dan sayuran). Killer yeast yang sering ditemukan, antara lain genus Aureobasidium pullulans, Cryptococcus infirmo-miniatus,
Cryptococcus macerans, Issatchenkia occidentalis, Issatchenkia orientalis, Kloeckera apis,
Kluyveromices marxianus, Pichia anomala, Pichia fermentans, Pichia kluyveri, Pichia
membranaefaciens, Pichia norvegensis, Pichia ohmeri, Pseudozyma spp., Trichosporon asahii,
Ustilago sparsa, Ustilago tragana, Ustilago trichophora. Diduga mereka berperan dalam
DAFTAR PUSTAKA
1. Schmitt, J.M., Breinig, F. (2006). Yeast viral killer toxin: lethality and self protection. Nature Publishing Group. 4:212-220
2. Daecon, Jim. The microbial world: yeast-like fungi
http://archive.bio.ed.ac.uk/jdeacon/microbes/yeast.htm. Institute of Cell and Molecular Biology, The University of Edinburgh. Diakses pada 26 November 2013]
3. Northrop, J.H., (1917). The role of yeast in the nutrition of an insect (Drosophila). J. Biol. Chem. 1917, 30:181-187
4. Marquina, D., Santos, A., Peinado, J.M. (2002). Biology of killer yeast. Int Microbiol 5: 65 71
5. Sugisaki, Y., Gunge, N., Kenji, S., Yamasaki, M., Tamura, G. (1985). Transfer of DNA killer plasmids from Kluyveromyces lactis to Kluyveromyces fragilis and Candida
pseudotropicalis. Journal of Bacteriology, Dec. 1985, p. 1373-1375.
6. Vondrej.V., Janderova B., Valasek L., (1996).Yeast killer toxin k1 and exploitation in genetic manipulations. Folia Microbiol. 41(5), 379-394.
7. Pintar, J., Starmer, T., (2003). The cost and benefits of killer toxin production by the yeast
Pichia kluyvery. Antonie van Leeuwenhoek 83: 89–98, 2003.
8. Honigberg, Saul M., (2011). Cell signals, cellcontacts, amd the organization of yeast communities. Eukaryot Cell. 2011 April; 10(4): 466–473.
9. K.A., Hamby, Hernández A, Boundy-Mills K, Zalom FG. (2012). Associations of yeasts with spotted-wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in cherries and raspberries. Appl Environ Microbiol. 78(14):4869-73.
10. Heard, G.M., Fleet, G.H., (1987). Occurrence and growth of killer yeast during wine fermentation. Applied and Environmental Microbiology. 53(9):2171-2174 11. K.A., Hamby, Hernandez A, Boundy-Mills K, Zalom FG, (2012). Association of yeast with