TINJAUAN PUSTAKA
Virus Hepatitis B dan Antigen Permukaan
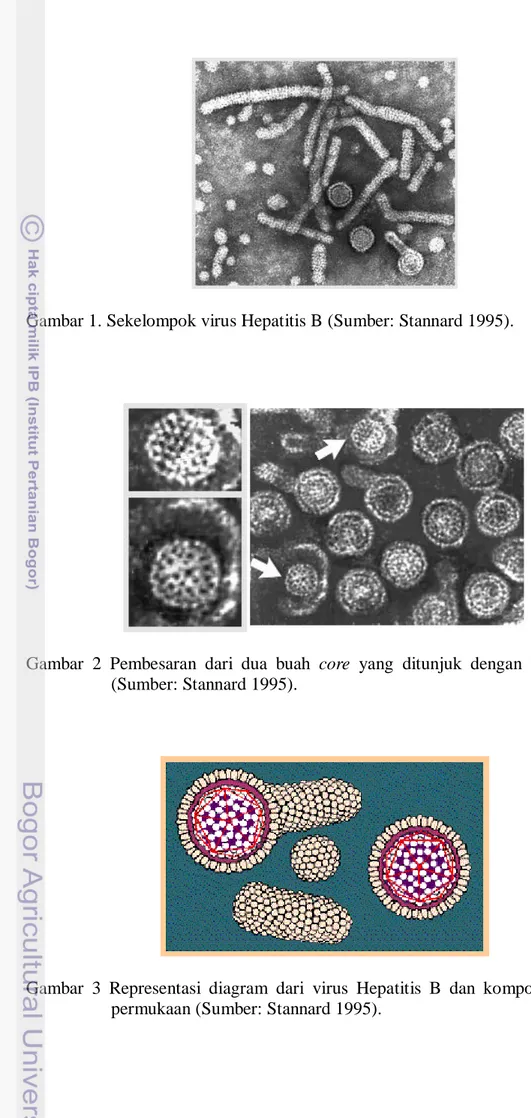
Menurut Dayal dan Maldonado (1998), Virus Hepatitis B masuk dalam famili virus Hepadna dan mempunyai ukuran genome yang terkecil diantara semua virus DNA hewan, yaitu dengan ukuran panjang 3,2 kb. Menurut Stannard (1995), virus Hepatitis B menyebabkan infeksi kronis dan akut pada hati manusia. Infeksi akut biasanya sampai beberapa bulan saja, sedangkan infeksi kronis bisa mencapai bertahun-tahun bahkan bisa sampai selama hidupnya. Diameter total dari virus Hepatitis B adalah 42 nm, sedangkan diameter core atau nucleocapsidnya adalah 27 nm. Core dilapisi oleh mantel (outer coat) yang tebalnya sekitar 4 nm. Protein yang terdapat pada mantel disebut surface antigen atau HBsAg. HBsAg ini kadang-kadang berkembang memanjang membentuk ekor (tubular) pada salah satu sisi dari partikel virus tersebut. Gambaran virus Hepatitis B yang lebih jelas, diilustrasikan pada Gambar 1, Gambar 2, Gambar 3 dan Gambar 4.
Virus Hepatitis B (VHB) telah menginfeksi sampai ratusan juta orang di seluruh dunia dan sekitar 20 juta orang terinfeksi setiap tahun, dan sekitar 78% penderita berdomisili di Asia (Joshi dan Kumar 2001). Di Indonesia, jumlah penderita penyakit tersebut mencapai sekitar 15 juta orang. VHB merupakan penyebab utama sirosis (pengerasan hati) dan kanker hepatoseluler (Human Hepatocellular Carcinoma) yang merupakan salah satu penyakit terganas penyebab kematian di seluruh dunia. Jenis kanker ini telah menyebabkan kematian lebih dari 1 juta orang setiap tahun (Ji et al. 2005).
Gambar 1. Sekelompok virus Hepatitis B (Sumber: Stannard 1995).
Gambar 2 Pembesaran dari dua buah core yang ditunjuk dengan tanda panah (Sumber: Stannard 1995).
Gambar 3 Representasi diagram dari virus Hepatitis B dan komponen antigen permukaan (Sumber: Stannard 1995).
Gambar 4 Ilustrasi virus Hepatitis B dengan capsid dan internal density yang tampak pada irisan melintang (Sumber: Dryden et al. 2006).
Data dari berbagai penelitian menunjukkan bahwa sebagian besar infeksi virus Hepatitis B yang bersifat menetap, timbul sebagai akibat infeksi pada waktu bayi dan anak-anak. Makin muda usia seseorang terkena infeksi virus tersebut, maka akan lebih besar kemungkinannya untuk menderita infeksi virus Hepatitis B yang menetap, sehingga lebih besar jumlah resiko untuk menjadi sirosis hati dan kanker hati primer dikemudian hari (Mulyanto et al. 2002).
Penelitian tentang sebaran geografis virus ini menunjukkan bahwa virus tersebut tersebar di seluruh dunia. Namun, prevalensi tertinggi ditemukan di Asia Tenggara dan Sub-sahara Afrika. Mulyanto et al. (1997) membagi zona distribusi sub tipe virus Hepatitis B di Indonesia berdasarkan perbedaan epitope pada HBsAg menjadi 4 bagian: (1) zona adw yang merupakan sub tipe utama di Indonesia meliputi Sumatera, Jawa, Kalimantan bagian Selatan, Bali, Lombok, Ternate dan Morotai, (2) zona ayw yang meliputi bagian barat dari Nusa Tenggara dan Maluku, (3) zona adr meliputi Papua, dan (4) campuran antara berbagai sub tipe yang terdapat di Kalimantan Selatan dan Sumbawa.
Virus Hepatitis B merupakan virus DNA untai ganda dengan panjang genome mencapai 3.2-3.3 kilo pasangan basa (kpb). Virus yang termasuk famili hepadnaviridae tersebut memiliki genome yang terbungkus oleh glycoprotein. Siklus replikasi virus ini dimulai dengan melekatnya protein selubung tersebut pada hepatosit. Di dalam inti sel hati, sintesis DNA virus disempurnakan dimana genome virus tersebut diubah menjadi cccDNA (covalently closed circular DNA). cccDNA inilah yang akan menjadi template untuk sintesis RNA yang kemudian akan diubah menjadi DNA virus (Lok dan McMahon 2001).
Selubung virus Hepatitis B (Hepatitis B virus envelope) terdiri dari membran glikoprotein dimana terdapat 3 bagian protein permukaan yaitu antigen pre-S1 (119 asam amino), pre-S2 (55 asam aminio) dan S (226 asam amino) (Jaoude dan Sureau 2005; Barrera et al. 2005). Beberapa ahli menggolongkan ketiga protein tersebut sebagai protein kecil (small), sedang (middle) dan besar (large). Antigen pre-S1 memiliki beberapa epitop yang memiliki daya immunogenik (Hu et al. 2004). Antigen bagian pre-S1 ini dibutuhkan oleh virus Hepatitis B untuk melakukan infeksi pada korban (Barrera et al. 2005). Menurut Deng et al (2005), asam-asam amino Pre-S1 pada nomor 21-47 merupakan epitop yang berfungsi untuk melekatnya virus pada jaringan hati. Oleh karena itu, protein bagian ini memiliki peranan yang sangat penting untuk siklus virus Hepatitis B. Antigen pre-S2 diduga mempunyai tingkat imunogenisitas lebih tinggi dibandingkan HBsAg (Ji et al. 2005) terutama 26 asam amino pada ujung N (Joung et al. 2004). Antigen pre-S2 mempunyai peranan sangat penting, hal ini telah dibuktikan secara nyata melalui serangkaian diagnosa (Maruyama et al. 2000). Menurut Hu et al. (2004a), HBsAg telah digunakan secara luas sampai saat ini sebagai vaksin konvensional, asam-asam amino ke 139-147 pada bagian S merupakan epitop utama pada protein S (HBsAg). Namun demikian, penghilangan epitop ini masih tetap dapat memberikan reaksi antigenisitas yang menunjukkan bahwa masih terdapat epitop lain selain epitop yang terletak pada asam-asam amino nomor 139-147 tersebut.
Struktur DNA dan Genome Virus Hepatitis B
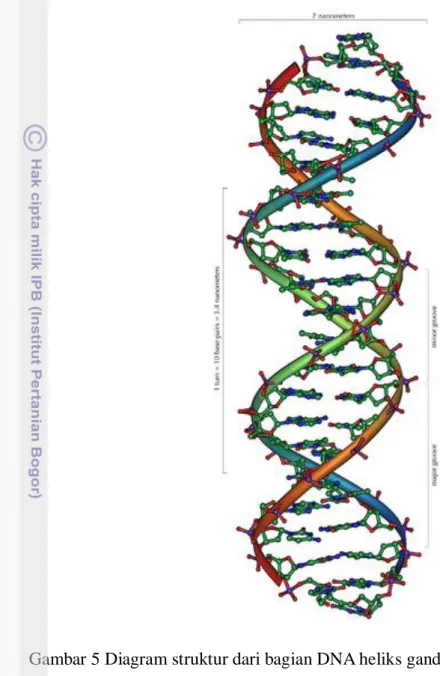
Menurut Winarno dan Agustinah (2007), DNA adalah deoxyribo nucleic acid, yaitu sebuah asam nukleat yang terdiri atas sejumlah nukleotida yang diatur sedemikian rupa sehingga berbentuk single strand. Biasanya dua buah utas DNA saling melingkar satu sama lain untuk membentuk sebuah double helix (heliks ganda), seperti ditunjukkan oleh Andre (2006) pada Gambar 5. Muladno (2002) menegaskan bahwa untuk membentuk rangkaian molekul DNA heliks ganda, basa nitrogen dari setiap nukleotida dalam satu rangkaian akan berpasangan dengan basa nitrogen dari setiap nukleotida pada rangkaian lainnya melalui ikatan hidrogen. Pengikatan basa nitrogen dari masing-masing nukleotida tersebut sangat spesifik. Basa A (Adenine) dari satu nukleotida selalu berikatan dengan basa T (Thymine) dari nukleotida lainnya, sedangkan basa G (Guanine) selalu berpasangan dengan basa C (Cytosine). Pasangan A dan T terbentuk dengan dua ikatan hidrogen, sedangkan pasangan G dan C terbentuk dengan tiga ikatan hidrogen. Oleh karena itu, pasangan G-C lebih stabil daripada pasangan A-T.
Winarno dan Agustinah (2007) menjelaskan, bahwa gugus basa DNA terdiri atas empat senyawa berikut: sitosin, adenin, guanin atau timin. Gugus gulanya adalah deoksiribosa. DNA terdapat di dalam kromosom prokariot dan eukariot dan di dalam mitokondria eukariot. DNA merupakan materi kebakaan atau keturunan di hampir semua organisme hidup yang mampu memperbanyak dirinya sendiri dalam pembelahan inti. Winarno dan Agustinah (2007) menjelaskan bahwa hipotesa Watson dan Crick merupakan hipotesa berdasarkan X-ray crystallography yang mengusulkan bahwa DNA merupakan suatu heliks ganda dari dua uliran rantai fosfat dan gula yang saling bergantian, dengan gula yang terkait oleh sepasang basa. Selanjutnya Muladno (2002) menjelaskan, bahwa Watson dan Crick akhirnya memperoleh Nobel dalam bidang biologi modern pada tahun 1953 setelah menemukan bukti bahwa struktur DNA adalah heliks ganda. Penemuan ini merupakan tonggak sejarah yang penting terhadap munculnya teknologi rekayasa genetika yang lahir 20 tahun kemudian, yaitu tahun 1973. DNA bersama-sama protein (histone) dan molekul ribo nucleic acid (RNA), terdapat di dalam inti sel. Ketiga materi tersebut saling kait mengkait dalam suatu
susunan yang sangat rumit membentuk kromosom yang merupakan komponen penting dalam semua sel mahluk hidup.
Gambar 5 Diagram struktur dari bagian DNA heliks ganda (Sumber: Andre 2006).
Melalui suatu proses kimia, kromosom dapat dikeluarkan dari inti sel. Selanjutnya, protein yang berikatan dengan DNA dilisiskan dengan enzim proteinase, sedangkan RNA yang masih berada di sekitar DNA dikatalisis atau diurai dengan enzim RNAse. Selanjutnya, DNA yang telah terbebas dari protein (histone) dan RNA siap direkayasa atau dimanipulasi dalam teknologi rekayasa genetika (Muladno 2002). Old dan Primrose (1989) menjelaskan, bahwa istilah manipulasi gen dapat diterapkan pada beberapa macam teknik genetika in-vivo maupun in-vitro yang canggih. Di negara-negara Barat terdapat definisi resmi
yang tepat untuk istilah manipulasi gen sebagai akibat adanya peraturan Pemerintah untuk mengendalikannya. Di Inggris, manipulasi gen didefinisikan sebagai pembentukan kombinasi baru materi yang dapat diturunkan dengan melakukan penyisipan (insertion) molekul-molekul asam nukleat, yang dihasilkan dengan cara apapun di luar sel, ke dalam suatu virus, plasmid bakteri atau sistem pembawa lainnya yang memungkinkan terjadinya penggabungan ke dalam organisme inang secara tidak alami tetapi selanjutnya mampu melakukan penggandaan lagi. Definisi resmi ini menekankan penggandaan molekul asam nukleat asing (asam nukleat ini hampir selalu DNA) di dalam tubuh organisme inang yang berbeda. Kemampuan untuk melintasi penghalang spesies alami dan memasukkan gen-gen dari organisme apapun ke dalam suatu organisme inang yang tidak berhubungan merupakan satu ciri penting manipulasi gen. Ciri penting kedua berupa kenyataan bahwa relatif sepotong kecil DNA tertentu digandakan dalam tubuh organisme inang.
Setiap organisme mempunyai sebuah genome yang mengandung semua informasi biologik yang diperlukan untuk membangun dan memelihara kehidupan organisme tersebut. Informasi biologik yang terkandung di dalam genome dikode oleh DNA yang terkandung di dalam genome yang dibagi ke dalam unit-unit khusus yang disebut gen (Barnum 2005). Menurut Winarno dan Agustinah (2007), genome dalam arti sederhana berarti satu set lengkap mengandung informasi genetika yang dimiliki oleh suatu organisme. Selanjutnya dijelaskan oleh Muladno (2002), bahwa setiap mahluk hidup mempunyai sel yang di dalam inti selnya terdapat kromosom dengan jumlah berbeda-beda untuk setiap mahluk hidup. Manusia mempunyai 23 pasang kromosom dalam setiap intinya. Sapi mempunyai 30 pasang kromosom, lalat buah Drosophila mempunyai empat pasang kromosom, bakteri E. coli mempunyai satu kromosom. Beberapa peneliti lain menambahkan, bahwa virus hepatitis B mempunyai 1 kromosom (Dayal dan Maldonado 1998; Mason et al. 1998; Burda et al. 2001; Muljono dan Soemohardjo 2003; Anzola 2004; Wagner et al. 2004; Beck dan Nassal 2007; GenBank 2008; Nurainy et al. 2008).
Menurut Muladno (2002), total kromosom dalam inti sel dinamakan genome atau lebih tepatnya disebut genome inti karena berasal dari inti sel. Jadi
bisa dikatakan bahwa genom manusia terdiri atas 23 pasang kromosom, genom sapi terdiri atas 30 pasang kromosom, dan seterusnya. Apabila DNA dari genom tersebut direntang secara linear, maka ukuran panjang rentangan DNA pada genom tersebut berbeda-beda pada setiap organisme seperti dijelaskan pada Tabel 1. Rentangan DNA dari genom tersebut disebut genomic DNA.
Berdasarkan data pada Tabel 1, maka dapat dijelaskan bahwa besarnya ukuran genom tidak mencerminkan besarnya ukuran makhluk hidup, seperti apabila membandingkan ukuran tubuh manusia dengan ukuran tubuh tikus yang sangat jauh berbeda, tetapi keduanya mempunyai ukuran genom yang hampir sama. Demikian pula, ukuran tubuh cacing yang jauh lebih besar dari ukuran tubuh lalat buah Drosophila, tetapi ukuran genome cacing lebih sedikit daripada ukuran genome lalat buah Drosophila.
Tabel 1 Ukuran genome dari beberapa makhluk hidup dalam bentuk haploid
Jenis mahluk hidup Ukuran genome (dalam pb) Virus Hepatitis B 3 215* Virus T4 160 000** Bakteri E.coli 4 200 000** Jamur 20 000 000** Cacing 80 000 000**
Lalat buah Drosophila 140 000 000**
Tikus 3 000 000 000**
Manusia 3 300 000 000**
Sumber: *Nurainy et al. 2008, **Lewin 1990.
Muladno (2002) menyatakan, bahwa virus dan bakteri merupakan makhluk hidup yang sederhana dan mempunyai ukuran genome yang kecil sehingga organisasi genomenya juga sederhana. Oleh karena itu, kedua makhluk hidup ini menjadi “bahan utama” untuk penelitian-penelitian dalam bidang genetika molekuler. Seperti telah diuraikan di atas, bahwa virus Hepatitis B mempunyai ukuran genome yang terkecil diantara semua virus DNA, yang menurut beberapa peneliti (Dayal dan Maldonado 1998; Burda et al. 2001; Muljono dan Soemohardjo 2003; Anzola 2004; Wagner et al. 2004; Beck dan Nassal 2007; Nurainy et al. 2008), virus Hepatitis B mempunyai ukuran panjang genom sekitar
3.2 kb. Dayal dan Maldonado (1998) menjelaskan bahwa virus hepatitis B mempunyai ciri-ciri sebagai berikut:
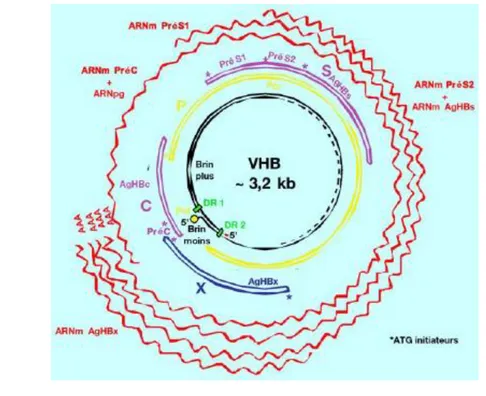
1 Genome virus hepatitis B yang menurut mereka ditemukan oleh William S. Robinson dari Stanford University School of Medicine merupakan genome yang terkecil dengan ukuran panjang 3,2 kb. Diagram organisasi genome virus Hepatitis B diilustrasikan pada Gambar 7.
2 Informasi genetik dibawa melalui DNA rantai ganda.
3 DNA penyusun genome terdiri atas rantai yang lebih panjang yang disebut rantai negatif (-) lengkap, dan rantai positif (+) tidak lengkap, terdapat sekitar 15-50% merupakan rantai tunggal.
4 Genome sirkuler dipertahankan dalam bentuk longgar oleh pasangan basa dari sekuen overlap 240 nukleotida dan Direct Repeats (DR) pendek antara kedua rantai pada ujung 5’.
5 Ujung 5’ dari rantai negatif (-) mengikat protein secara kovalen, sedangkan ujung 5’ dari rantai positif (+) mengikat oligoribonukleotida sebagai RNA primer.
6 Nukleokapsid berbentuk icosahedral. 7 Nukleokapsid dikelilingi oleh amplop lipid. 8 Diameter Nukleokapsid 27 nm.
9 Melalui gambar mikrograf elektron, virus tampak spherical (bentuk bola) dengan diameter total 42 nm.
10 Mengandung 4 Overlapping Open Reading Frames (ORF): S, C, P, dan X yang disandi pada rantai DNA (-).
11 Nomer Triangulasi=3 (T=3).
12 Gen S terdiri atas 3 bagian: yaitu pre-S1, pre-S2, dan S. Tiga macam protein yang berbeda diproduksi oleh kombinasi dari ke tiga gen tersebut dalam kombinasi yang berbeda: protein L (large protein), yaitu protein pada amplop yang dibuat berdasarkan pengkodean dari gen-gen pre-S1, pre-S2, dan S; protein M (middle-sized proteins/medium surface protein) pada amplop yang dikode oleh gen pre-S2 dan gen S; dan protein S yang merupakan komponen mayor (major component) dari HBsAg.
Gambar 6 Diagram organisasi genome virus hepatitis B (Sumber: Wagner 2004).
Nurainy et al. (2008) melalui risetnya yang berjudul Genetic Study of Hepatitis B Virus in Indonesia Reveals a New Subgenotype of Genotype B in East Nusa Tenggara dengan menggunakan isolat Virus Hepatitis B 2059Java, berhasil menemukan sekuen genom lengkap Virus Hepatitis B (Lampiran 2). Selanjutnya dijelaskan, bahwa sekuen genome lengkap tersebut tersusun atas 4 overlapping open reading frames yang masing-masing mempunyai sekuen sebagai berikut: 1 Gen P (sekuen:2307-3215, 1-1623); yang menyandi pembentukan enzim-enzim
DNA polymerase, reverse trancriptase, dan RNAse.
2 Gen S yang terdiri atas 3 bagian, yaitu: gen pre-S1, gen pre-S2, dan gen S. Tiga jenis protein amplop VHB yang berbeda diproduksi berdasarkan pengkodean dari kombinasi tiga gen-gen ini dalam kombinasi yang berbeda. (1) Protein L (large protein) dikode oleh kombinasi dari gen-gen pre-S1, pre-S2, dan S (sekuen: 2848-3215, 1-835). (2) Protein M (medium protein) dikode oleh kombinasi dari gen pre-S2 dan gen S (sekuen: 3205-3215, 1-835). (3) Protein S (HBsAg) dikode oleh gen S (155-835).
3 Gen X (sekuen: 1374-1838); yang menyandi pembentukan protein X yang peranannya belum diketahui secara pasti, tetapi diduga berperan dalam aktivasi transkripsi.
4 Gen C (sekuen: 1814-2452); yang berperan mengkode pembentukan HBcAg (antigen core).
DNA Rekombinan dan Kloning DNA
DNA rekombinan adalah suatu DNA buatan atau hasil rekayasa yang berasal dari satu sumber atau lebih yang tergabung ke dalam satu molekul rekombinan (Barnum 2005). Selanjutnya, Winarno dan Agustinah (2007) menjelaskan, bahwa teknologi rekayasa genetika merupakan kegiatan bioteknologi modern dengan teknologi DNA rekombinan (rDNA) untuk melakukan pemindahan atau transfer suatu sifat tertentu yang dibawa gen, yang tersusun dalam DNA, dari suatu spesies yang sama atau berbeda untuk menghasilkan spesies baru yang lebih unggul. Muladno (2002) juga menyatakan, seiring dengan kemajuan teknologi molekuler, perpindahan gen dapat terjadi antar organisme yang sama sekali tidak berkerabat dekat, misalnya gen manusia dipindahkan ke bakteri atau gen manusia dipindahkan ke hewan ternak. Perpindahan gen tersebut mengakibatkan terbentuknya molekul DNA yang berasal dari sumber yang berbeda dapat digabungkan menjadi DNA rekombinan. Teknik menggabungkan molekul DNA tersebut dikenal sebagai Teknik DNA Rekombinan.
Biasanya DNA rekombinan merupakan gabungan antara DNA vektor yang merupakan molekul DNA yang dapat mereplikasi diri dan DNA asing yang biasanya berupa gen dari suatu mahluk hidup. Vektor tersebut berfungsi sebagai pembawa DNA asing yang berasal dari suatu organisme untuk dipindahkan ke dalam organisme lain. Gen yang terkandung pada DNA rekombinan di dalam organisme resipien diharapkan dapat diekspresikan untuk menghasilkan protein (Muladno 2002). Menurut Glick dan Pasternak (1994), teknologi DNA rekombinan, juga disebut kloning gen atau kloning molekuler, adalah suatu istilah yang mencakup sejumlah protokol percobaan yang bertujuan untuk transfer informasi genetik (DNA) dari suatu organisme ke organisme yang lain. Percobaan DNA rekombinan biasanya mengikuti prosedur sebagai berikut:
1 DNA target (DNA asing, DNA insert, DNA klon) dari organisme donor diekstrak, dipotong secara enzimatik, kemudian disambung dengan plasmid
atau DNA vektor (vektor kloning) untuk membentuk suatu bentuk baru yang disebut molekul DNA rekombinan (rDNA).
2 DNA rekombinan tersebut ditransfer ke dalam sel inang. Proses introduksi DNA rekombinan ke dalam suatu sel inang atau bakteri disebut transformasi. 3 Selanjutnya sel-sel bakteri tersebut dikultur dan diseleksi atau dimurnikan,
kemudian diisolasi.
4 Sel-sel bakteri inang yang telah dimurnikan dan diisolasi, siap dikultur atau dikembangbiakkan untuk memproduksi protein spesifik yang dikode oleh DNA klon yang terkandung dalam DNA rekombinan.
Pembuatan DNA rekombinan memerlukan bantuan dua macam enzim. Pertama, enzim endonuclease (restriction enzyme) berperan sebagai pemotong molekul DNA. Kedua, enzim ligase berfungsi untuk menggabungkan molekul-molekul DNA yang telah dipotong oleh enzim restriksi (Muladno 2002; Barnum 2005).
Ilustrasi tahapan pembuatan DNA rekombinan pada Gambar 8 menunjukkan bahwa enzim endonuclease yang digunakan untuk memotong kedua sumber DNA adalah BamHI. Enzim restriksi ini memotong kedua molekul DNA tersebut pada lokasi yang sama dengan membentuk potongan sticky end atau kohesif. Selanjutnya enzim ligase DNA menggabungkan kedua molekul DNA tersebut dengan ikatan kovalen menjadi satu molekul DNA rekombinan.
Muladno (2002) menegaskan bahwa pada prinsipnya kloning DNA adalah proses penggandaan jumlah DNA rekombinan melalui proses perkembangbiakan sel bakteri (biasanya E. coli). Hal ini dilakukan dengan memasukkan DNA rekombinan yang dihasilkan dari proses penggabungan tersebut di atas ke dalam sel E. coli. Selanjutnya sel ini diinkubasi pada suhu optimal sehingga sel dapat berkembangbiak secara eksponensial. Masuknya molekul DNA rekombinan ke dalam sel akan mengubah fenotip sel tersebut, sehingga proses pemasukan molekul DNA ke dalam sel juga disebut transformasi. Sel yang digunakan dalam proses transformasi ini biasanya disebut dengan sel kompeten. Plasmid merupakan molekul kecil yang berukuran sekitar lima ribu pasang basa yang terdapat di dalam sel bakteri Escherichia coli, posisinya terpisah dengan kromosom dan mampu mereplikasi sendiri tanpa harus bergantung kepada
kromosom, kebanyakan berupa rangkaian molekul DNA untai ganda dan biasanya berbentuk bulat. Selanjutnya Glick dan Pasternak (1994) menerangkan, bahwa ori adalah sekuen nukleotida yang merupakan tempat diawalinya atau dimulainya sintesis DNA pada saat replikasi.
Barnum (2005) menjelaskan, ada beberapa cara dalam melakukan kloning DNA, dan metode yang digunakan bervariasi tergantung pada tipe DNA, tipe sel inang, dan tujuan akhir dari kloning DNA. Contohnya, tipe vektor yang digunakan untuk kloning akan tergantung kepada apakah DNA klon akan tetap berada di dalam vektor atau akan disisipkan ke dalam kromosom sel inang.
Vaksin Hepatitis B
Menurut Dayal dan Maldonado (1998), vaksin hepatitis B yang pertama kali mendapat lisensi (pada tahun 1981) adalah Heptavax-B dan telah dipasarkan oleh Merck Sharp dan Dhome. Vaksin tersebut diperoleh dari hasil ekstraksi dan pemurnian antigen HBsAG dari serum penderita hepatitis B kronis. Selanjutnya dijelaskan bahwa vaksin rekombinan atau sebagai hasil rekayasa genetika yang telah berhasil diproduksi secara komersial adalah Recombivax HB Chiron Corp dan Merck serta Engerix-B oleh SmithKline Biologicals. Vaksin rekombinan tersebut diproduksi dari hasil kloning gen HBsAg yang terdapat di dalam yeast. Joung et al. (2004) menyatakan, hampir semua vaksin hepatitis B konvensional yang sudah mendapat lisensi saat ini adalah vaksin yang dihasilkan dari plasma. Namun, keberhasilan program imunisasi menyebabkan pasien hepatitis B yang akan menjadi sumber vaksin tersebut semakin berkurang yang berakibat pada semakin terbatasnya darah yang dapat digunakan sebagai sumber vaksin. Oleh sebab itu produksi vaksin hepatitis B dengan menggunakan plasma semakin sulit dilakukan. Kekhawatiran terhadap adanya kontaminan pada darah terutama oleh virus berbahaya seperti HIV, menimbulkan kekhawatiran tersendiri untuk menggunakan vaksin yang bersumber dari plasma tersebut.
Menurut Mulyanto et al. (1997), Indonesia merupakan daerah endemik sedang sampai tinggi untuk penyakit hepatitis B, sehingga WHO menghimbau untuk segera melaksanakan usaha pencegahan. Pengobatan terhadap penderita penyakit hepatitis B yang sangat mahal menyebabkan tindakan preventif melalui
vaksinasi merupakan tindakan yang lebih tepat. Vaksinasi secara besar-besaran dinilai efektif untuk mencegah terjadinya penyakit ini. Selanjutnya Mulyanto et al. (2002) menyatakan, bahwa pada tahun 1987, Pemerintah Indonesia menjadikan Pulau Lombok sebagai model immunisasi massal Hepatitis B pertama di dunia. Hal ini disebabkan karena tingkat endemik penyakit tersebut di Pulau Lombok sangat tinggi. Hasil proyek percontohan tersebut cukup menggembirakan sehingga pemerintah mulai memperluas program immunisasi ke 4 propinsi yang lain di tahun 1991 dan kemudian ke 6 propinsi lainnya pada tahun 1992. Saat ini pemerintah Indonesia telah mengintegrasikan vaksinasi Hepatitis B untuk balita ke dalam Program Pengembangan Imunisasi (Extended Program of Immunization).
Berbagai upaya untuk menghasilkan vaksin yang mengandung antigen rekombinan telah dilakukan, diantaranya dengan memproduksi antigen menggunakan sel tanaman maupun ragi. Keberhasilan ekspresi antigen tersebut telah dilaporkan melalui kultur sel tembakau (Kumar et al. 2003), maupun ragi (Maruyama et al. 2000; Lu et al. 2002; Ddeman & Zyl 2003; Hu et al. 2004). Meskipun vaksin tersebut memberikan hasil yang sangat memuaskan, sistem produksi menggunakan hewan maupun tanaman terhambat oleh lamanya waktu yang diperlukan, terjadi variasi pada produk akhir, terkontaminasi oleh bahan-bahan kimia pertanian serta kesulitan dalam meningkatkan hasil produk akibat rumitnya faktor-faktor regulasi. Penggunaan bakteri E. coli menjadi pilihan terakhir karena waktu yang lebih singkat, harga media lebih murah serta teknologi pembiakan maupun komponen-komponen yang dibutuhkan untuk optimalisasi transkripsi maupun translasi telah dikuasai (Ali 2006).
Produksi bagian yang sederhana dari antigen permukaan Hepatitis B dengan menggunakan bakteri E. coli bisa dilakukan, namun demikian berbeda dengan hasil ekspresi yang dilakukan dengan induk semang hewan maupun tumbuhan yang dapat menghasilkan produksi yang tinggi, ekspresi menggunakan E. coli menghasilkan produk yang sedikit. Hal ini disebabkan oleh sifat antigen permukaan Hepatitis B yang toksik bagi E. coli. Maeng et al. (2001) melakukan percobaan ekspresi gen virus Hepatitis B secara parsial yang diikuti dengan menggabungkan gen tersebut dengan gen penyandi enzim
gluthathion-S-transferase (GST), dengan demikian ekspresi gen dan kelarutan antigen permukaan Hepatitis B pada E. coli tersebut bisa ditingkatkan. Ekspresi gen tersebut dilakukan dibawah kontrol promoter tac. Hasil penelitian tersebut menunjukkan bahwa terjadi peningkatan tingkat ekspresi antigen pre-S1 yang digabung dengan GST. Koschorreck et al. (2005) juga melaporkan terjadi peningkatan solubilitas protein rekombinan yang digabung dengan GST. Terjadinya peningkatan kelarutan ini sangat penting untuk proses pemurnian antigen rekombinan dengan menggunakan kolum yang telah tersedia secara komersial.
Aplikasi Rekayasa Genetik di Bidang Peternakan
Menurut Winarno dan Agustinah (2007), WHO telah meramalkan bahwa populasi dunia akan berlipat dua pada tahun 2020 sehingga jumlahnya akan lebih dari 10 milyar. Oleh karena itu, produksi pangan juga harus ditingkatkan. Kendalanya adalah jumlah sisa lahan dunia yang belum termanfaatkan saat ini sangat kecil dan terbatas. Teknologi rekayasa genetika atau GMO akan memainkan peranan yang sangat penting dalam mengatasi kendala tersebut. Teknologi rekayasa genetika dapat menjadi strategi yang sangat bagus untuk meningkatkan produksi pangan. Wiryosuhanto dan Sudradjat (1992) menjelaskan, bahwa teknologi rekayasa genetika merupakan alat yang dapat membantu pengusaha industri peternakan untuk meningkatkan status kesehatan ternaknya, meningkatkan feed convertion, dan memperpendek waktu pemeliharaan sampai mencapai berat badan yang sesuai dengan permintaan pasar. Teknologi rekayasa genetik memberi harapan untuk diaplikasikan di dalam pembangunan peternakan di masa datang, yaitu bioteknologi molekuler yang berkaitan dengan keberhasilan teknologi kloning, ternak transgenik, dan vaksin. Winarno (2004) menyatakan, bahwa teknologi rekayasa genetika juga mampu membantu masalah yang dihadapi oleh industri susu dalam mengatasi masalah turunnya suplai renin. Renin, yang sekarang lebih dikenal dengan nama chymosin, adalah enzim proteolitik yang digunakan dalam pembuatan keju oleh industri susu. Secara konvensional, renin diproduksi dari abomasum (lambung ke-4) anak sapi. Timbulnya kekurangan renin menyebabkan para industri keju mencari alternatif
lain dengan memproduksi renin mikroba yang diekstraksi dari fungi, namun sifat biokimianya tidak sama dengan renin dari abomasum anak sapi. Hal ini bisa diatasi dengan cara mentransfer gen dari anak sapi yang mengkode pembentukan renin ke dalam bakteri, selanjutnya bakteri tersebut digunakan sebagai mesin pembentuk renin yang sifat biokimianya sama dengan renin dari abomasum anak sapi.