Bab II Tinjauan Pustaka
II.1 Gajah
Gajah adalah hewan mamalia, merupakan satu-satunya famili yang tersisa dari ordo Proboscidea. Gajah merupakan hewan darat terbesar di dunia. Sepanjang 55 juta tahun terdapat 500 spesies gajah yang dikenal, tetapi saat ini di dunia hanya terdapat 2 spesies gajah, yaitu gajah Afrika dan gajah Asia.
II.1.1 Gajah Afrika (Loxodonta africana).
Gajah-gajah dari genus Loxodonta, yang lebih dikenal dengan gajah Afrika, sekarang ini ditemukan di 37 lokasi di Afrika. Gajah Afrika berbeda dengan gajah Asia dalam beberapa hal, yang paling nyata terlihat adalah telinganya yang lebih besar dan berbentuk konkaf terbalik. Gajah Afrika jantan dan betina keduanya memiliki gading dan biasanya memiliki rambut yang lebih sedikit dibandingkan dengan gajah Asia. Gajah Afrika diklasifikasikan menjadi dua subspesies, yaitu gajah savanna (Loxodonta africana africana) dan gajah hutan (Loxodonta africana cyclotis) [Eggert et al., 2002], analisis DNA terbaru menyatakan bahwa keduanya merupakan species yang berbeda [Roca et al., 2001], namun pernyataan ini belum sepenuhnya diterima secara univesal. Gajah savanna merupakan binatang terbesar didunia, tingginya mencapai 4 m sampai batas bahu dan berat rata-ratanya 7.000 kg (7.7 tons). Gajah savanna paling sering ditemukan di padang rumput, rawa-rawa, dan tepi danau. Kebanyakan gajah savanna tinggal di bagian selatan Sahara. Jenis lain dari gajah Afrika adalah gajah hutan (Loxodonta cyclotis). Dibandingkan dengan gajah savanna, bentuk telinga biasanya lebih kecil dan lebih bulat, dan gadingnya lebih kurus dan tidak terlalu banyak mengarah keluar. Gajah hutan dapat mencapai berat 4.500 kg dan tingginya hingga 3 m. Gajah hutan lebih sulit dipelajari karena lingkungan hidup mereka yang sulit dijangkau. Secara normal mereka hidup di hutan hujan di pusat dan barat Afrika, namun ada kalanya mereka menyisiri tepian hutan dan bersinggungan dengan teritorial gajah savanna dan menghasilkan keturunan dengannya. Di tahun 1979, Iain Douglas-Hamilton memperkirakan populasi Gajah Afrika sekitar 1.3 juta ekor [Douglas et al., 1979].Perkiraan ini kontroversial dan dan berlebihan [Parker
et al., 1983]. Namun telah disitasi secara luas dan menjadi fakta yang salah dalam mengukur jumlah nyata polulasi dari species itu. Ditahun 1980, Loxodonta menerima perhatian yang luas dari dunia luas berkaitan dengan jumlah populasinya yang memprihatinkan di Afrika Timur. Menurut Laporan Status gajah Afrika IUCN [Blanc et al., 2007] di Afrika ada kira-kira antara 470,000 dan 690,000 gajah Afrika di alam liar. Sejauh ini populasi terbesar ditemukan di Afrika Selatan dan Timur. Akibatnya di Afrika Selatan membuat undang-undang bahwa ilegal bila membunuh gajah.
Gambar II.1 Gajah Afrika (Loxodonta africana). Saat ini merupakan gajah terbesar di dunia dengan tinggi mencapai 4 meter dan berat 7.000 kg. Telinganya yang lebih besar dan berbentuk konkaf terbalik
II.1.2 Gajah Asia (Elephas maximus).
Diperkirakan populasi gajah Asia sekitar 60,000 ekor, atau sekitar sepersepuluh dari jumlah gajah Afrika. Lebih rinci diperkirakan gajah liar antara 38.000 ekor hingga 53.000 ekor dan gajah yang domestikasi sekitar 14.500 hingga 15.300 yang tersebar di kebun binatang dan pusat perlindungan hewan di Asia [Asian Elephant distribution, 2007] Asian elephants mengungkapkan bahwa kemungkinan penyebab utamanya adalah perburuan liar dan pengrusakan habitat hidupnya akibat ulah manusia. Gajah Asia berbeda daripada gajah Afrika. Gajah Asia memiliki telinga lebih kecil sedikit daripada gajah Afrika, mempunyai dahi yang rata, dan dua bonggol di kepalanya merupakan puncak tertinggi gajah,
kepala. Selain itu, ujung belalai gajah Asia hanya mempunyai 1 bibir, sementara gajah Afrika mempunyai 2 bibir di ujung belalai. Kedua jenis kelamin gajah Afrika mempunyai gading sementara hanya gajah Asia jantan yang mempunyai gading yang jelas kelihatan. Beberapa subspecies Elephas maximus, yaitu gajah Sri Lanka, gajah India, gajah Sumatera dan gajah Borneo.
II.1.2.1 Gajah Sri Lanka
Subspesies yang pertama adalah gajah Sri Lanka (Elephas maximus maximus). Ditemukan hanya di pulau Sri Lanka, hewan ini merupakan yang terbesar di Asia. Populasinya diperkirakan 3,000–4,500 ekor hingga hari ini di alam liar. Gajah jantan dewasa memiliki bobot badan hingga 5,400 kg dan tinggi mencapai 3,4 m. Gajah Sri Lanka jantan mempunyai cranial besar yang menonjol, kedua jenis kelamin dari gajah ini mempunyai daerah depigmentasi lebih dibanding gajah Asia lainnya. Secara khas, permukaan kulit telinga, muka, belalai dan perut mereka dipenuhi bintik bintik merah muda.Untuk mencegah kepunahan dibangun tempat penangkaran gajah di Pinnawala Sri Lanka.
II.1.2.2 Gajah India
Gajah India (Elephas maximus indicus), jumlah populasinya sekitar 36,000 ekor, gajah ini berwarna abu-abu, dengan depigmentasi hanya pada bagian telinga dan belalainya. Gajah jantan dewasa memiliki bobot sekitar 5,000 kg dengan rata-rata tingginya sama dengan gajah Sri Lanka. Gajah ini tersebar di India, Laos, Burma, Kamboja, Thailand, Malaysia dan China.
II.1.2.3 Gajah Sumatera
Gajah dengan postur kecil dibanding gajah yang dibahas sebelumnya adalah gajah Sumatera (Elephas maximus sumatrensis). Populasi gajah ini diperkirakan dari 2,100 hingga 3,000 ekor. Kulitnya berwarna abu-abu terang dan depigmentation yang paling sedikit dibanding gajah Asia lainnya, dengan bintik merah muda hanya pada bagian telinga. Gajah Sumatera dewasa biasanya hanya memiliki tinggi 1.7–2.6 m sampai batas bahu dan bobot badan kurang dari 3,000 kg. Gajah ini hanya berada di pulau Sumatera biasanya di kawasan hutan.
Berikut merupakan klasifikasi gajah Sumatera: Kingdom Phylum Subphylum Class Order Superfamily Family Subfamilia Tribe Subtribe/Supergenus Genus Species Subspecies : Animalia : Chordata : Vertebrata : Mammalia : Proboscidea : Elephantoidea : Elephantidae : Elephantinae : Elephantini : Elephantina/Elephadon : Elephas : Elephas maximus
: Elephas maximus sumatrensis
II.1.2.4 Gajah Borneo
Ditahun 2003 telah diidentifikasikan subspesies baru didaerah Borneo, yaitu Gajah Kerdil Borneo, ini adalah gajah terkecil dibandingkan gajah Asia lainnya selain itu gajah ini juga paling jinak. Gajah Borneo memiliki telinga yang relatif besar, ekor yang panjang dan gading yang lurus.
Gambar II.2 Gajah Asia (Elephas maximus indicus). Warna kulit cenderung abu-abu dengan depigmentasi pada bagian telinga dan belalainya. Bobot badan mencapai 5.000 kg dengan tinggi 3-3,5 meter.
II.1.2 Peta Penyebaran Populasi Gajah
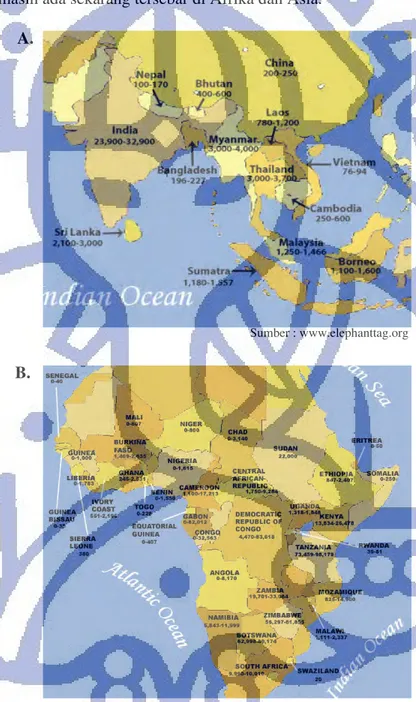
Gajah yang masih ada sekarang tersebar di Afrika dan Asia.
Gambar II.3 Gambar Peta Penyebaran Populasi Gajah. (A) Gambar penyebaran populasi gajah Asia yang menyebar di Indonesia, Malaysia, Kamboja, Thailand, Sri Lanka, India, Burma, China, dll dan (B) gambar penyebaran populasi gajah Afrika yang menyebar di Zambia, Zimbabwe, Republik Congo, Kenya, dll.
A.
B.
Sumber : www.scionline.org Sumber : www.elephanttag.org
II.1.3 Pola Hidup Gajah
Gajah hidup di dalam urutan sosial yang terstruktur. Kehidupan sosial dari jantan dan betina sungguh berbeda. Betina menghabiskan hampir seluruh hidupnya di dalam satu grup keluarga yang terdiri atas ibu, anak perempuan, saudara perempuan, dan bibi. Grup ini dipimpin oleh perempuan tertua. Sedangkan jantan dewasa menghabiskan waktunya dalam kehidupan sendiri (tidak berkelompok). Sebagai anggota dari kelas Mamalia, gajah berkembangbiak dengan cara beranak. Gajah betina hanya mampu melahirkan satu anak gajah dan akan menjaga anaknya sampai mampu berdiri dalam jangka waktu 5 tahun. Periode kehamilan gajah adalah 22 bulan, terlama dibandingkan hewan darat lainnya. Berat anak gajah pada umumnya 120 kilogram. Seekor gajah bisa hidup selama 70 tahun, bahkan lebih lama.
Gajah adalah hewan herbivora. Ia menghabiskan 16 jam sehari untuk mengumpulkan makanan tanaman. Makanannya terdiri atas sedikitnya 50% rumput, ditambah dengan dedaunan, ranting, akar, dan sedikit buah, benih dan bunga. Karena gajah hanya mencerna 40% dari yang dimakannya, mereka harus mengonsumsi makanan dalam jumlah besar. Gajah dewasa dapat mengonsumsi 300 hingga 600 pon (140-270 kg) makanan per hari. Enam puluh persen dari makanan tersebut tertinggal dalam tubuh gajah tidak tercerna.
II.2 Mitokondria
Mitokondria merupakan organel yang berperan dalam metabolisme energi dalam sel eukariot. Mereka bertanggung jawab hampir pada semua penggunaan energi yang berasal dari pemecahan karbohidrat dan asam lemak, dimana dikonversi dalam bentuk ATP melalui proses fosporilasi oksidatif. Mitokondria sangat unik karena memiliki DNA sendiri, dimana mengkode tRNA, rRNA, dan beberapa protein dalam mitokondria [Cooper, 2000]
II.2.1 Struktur Mitokondria
Mitokondria berbentuk elips dengan diameter ~0,5 m dan panjang 0,5 – 1,0 m, serta berfungsi sebagai penghasil energi. Mitokondria terdiri dari 4 bagian
penting, yaitu: (1) membran luar, (2) membran dalam, (3) ruang antarmembran, dan (4) matriks. Membran luar adalah tempat berlangsungnya reaksi fosforilasi oksidatif, membran dalam adalah tempat berlangsungnya sintesis fosfolipid, ruang antarmembran adalah tempat dihasilkannya nukleotida kinase, dan matriks adalah tempat terjadinya oksidasi piruvat dan siklus asam sitrat (siklus Krebs). Struktur mitokondria dapat dilihat pada Gambar II.4. Mitokondria didukung oleh hipotesis endosimbiosis yang mengatakan bahwa pada tahap awal evolusi sel eukariot bersimbiosis dengan prokariot (bakteri) (Margullis, 1981). Kemudian keduanya mengembangkan hubungan simbiosis dan membentuk organel sel yang pertama. Adanya DNA pada mitokondria menunjukkan bahwa dahulu mitokondria merupakan entitas yang terpisah dari sel inangnya. Hipotesis ini ditunjang oleh beberapa kemiripan antara mitokondria dan bakteri. Ukuran mitokondria menyerupai ukuran bakteri, dan keduanya bereproduksi dengan cara membelah diri menjadi dua. Hal yang utama adalah keduanya memiliki DNA berbentuk lingkar. Oleh karena itu, mitokondria memiliki sistem genetik sendiri yang berbeda dengan sistem genetik inti. Selain itu, ribosom dan rRNA mitokondria lebih mirip dengan yang dimiliki bakteri dibandingkan dengan yang dikode oleh inti sel eukariot (Cooper, 2000).
Gambar II.4. Struktur Mitokondria. Mitokondria berbentuk lonjong atau bulat, dibalut oleh sistem membran ganda, yang terdiri dari membran luar dan membran dalam. Lipatan membran dalam (krista)
II.2.2 Fungsi Mitokondria
Fungsi utama mitokondria adalah memproduksi energi kimia dalam bentuk ATP yang akan dipergunakan untuk aktivitas seluruh sel-sel tubuh manusia. Secara garis besar, reaksi pembentukan ATP yang berlangsung di mitokondria dapat dibagi menjadi 3 tahap yaitu (1). Reaksi oksidasi piruvat (atau asam lemak) menjadi CO
2. Reaksi ini terkait dengan reduksi NAD +
dan FAD menjadi NADH dan FADH
2 yang berlangsung dalam ruang matriks mitokondria. (2). Transfer
elektron dari NADH dan FADH
2 ke O yang berlangsung pada membran dalam
dan terkait dengan pembentukan proton motive force atau gradien elektrokimia lintas membran dalam mitokondria. (3). Pemanfaatan energi yang tersimpan dalam bentuk gradien elektrokimia untuk memproduksi ATP. Reaksi ini dikatalisis oleh kompleks enzim F
0-F1 ATP sintetase yang berlokasi pada
membran dalam (Gambar II.5).
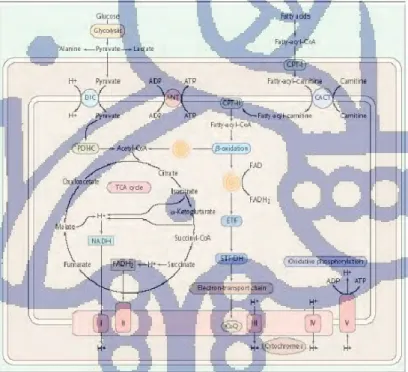
Gambar II.5 Jalur metabolik dalam mitokondria. Spiral menunjukkan reaksi oksidasi yang menghasilkan pelepasan acetyl-coenzim A dan penurunan flavoprotein. ANT adenine nucleotide translocator, CACT carnitine-acyl carnitine translocase, CPT carnitine palmitcyltransferase, DIC dicarboxylate carrier, ETF electron-transfer flavoprotein. ETH-DH electron electron-transfer dehydrogenase, PDHC pyruvate dehydrogenase complex.
II.2.3 DNA Mitokondria
Mitokondria memiliki genomnya sendiri yang dikenal sebagai DNA mitokondria (mtDNA) dan berbeda dengan genom inti. Struktur mtDNA dapat dilihat pada Gambar II.6. DNA gajah berbentuk sirkuler dan memiliki untai ganda yang terdiri atas untai Heavy (H) dan untai Light (L). Penamaan ini didasarkan pada perbedaan densitas tiap untai dalam gradien denaturan cesium klorida (CsCl), dimana untai H memiliki berat molekul yang lebih besar dibandingkan dengan untai L karena untai H memiliki lebih banyak basa-basa purin yang memiliki dua buah cincin pada strukturnya. Untai L memiliki komposisi basa sebagai berikut T 24,7%, C 31,2%, A 30,9%, dan G 13,2%. Dapat dilihat bahwa komposisi basa purin (A+G) lebih kecil (44,1%) dibandingkan dengan basa pirimidin (T+C), yaitu 55,9%.
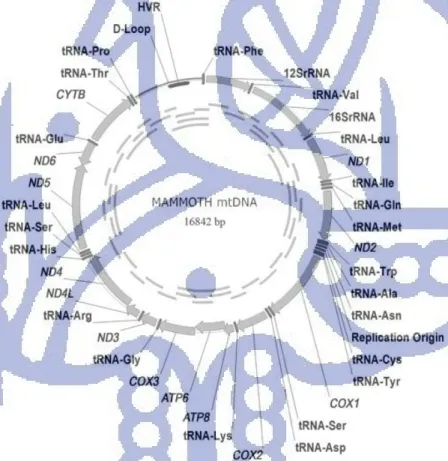
Gambar II.6 DNA mitokondria Mammoth (Mammuthus primigenius)
MtDNA berbentuk sirkular, beruntai ganda (untai H untuk Heavy dan untai L untuk Light), berukuran 16842 pb yang terdiri dari daerah pengode (coding region) yang mengode 2 rRNA, 22 tRNA, 13 polipeptida dan daerah yang tidak mengode (non coding region) atau daerah pengontrol yang mengandung D-loop. [Rogaev et al.,
DNA mitokondria Mammoth berukuran 16842 pb, mtDNA gajah Asia berukuran 16902 pb dan mtDNA gajah Afrika berukuran 16866. Urutan lengkap gajah Afrika (Loxodonta africana) ditentukan pertama kali oleh Hauf et al. (2000). Urutan lengkap nukleotida Mammoth (Mammuthus primigenius) pertama kali ditentukan oleh Krause et al. (2005). Urutan lengkap gajah Asia (Elephas maximus indicus) ditentukan oleh Rogaev et al. (2006). MtDNA gajah mengandung 37 gen pengode untuk 2 rRNA, 22 tRNA, dan 13 polipeptida yang merupakan subunit kompleks enzim yang terlibat dalam fosforilasi oksidatif, yaitu subunit 1, 2, 3, 4, 4L, 5, dan 6 dari kompleks I (NADH dehidrogenase); subunit b (sitokrom b) dari kompleks III (koenzim Q - sitokrom C reduktase); subunit I, II, dan III dari kompleks IV (sitokrom oksidase); serta subunit 6 dan 8 dari kompleks V (ATP sintase). Selain gen pengode tadi, DNA mitokondria memiliki daerah yang tidak mengkode (noncoding region) sepanjang 1420 pb, mulai dari nukleotida 15422 sampai 16842 dan terletak di antara gen tRNApro dan tRNAphe. Daerah non coding ini mengandung daerah dengan variasi tinggi yang disebut dengan displacement loop (D-loop). D-loop merupakan daerah beruntai tiga (triple stranded), untai ketiga lebih dikenal sebagai 7S DNA. Selain D-loop, daerah non coding juga mengandung origin of replication untuk untai H (OH) dan
promoter untuk untai H dan L (PL dan PH), oleh karena itu sering disebut daerah
pengontrol (control region).
II.2.4 Sifat DNA Mitokondria
II.2.4.1 Pola Pewarisan Secara Maternal
DNA mitokondria berbeda dengan DNA inti karena mtDNA diwariskan melalui garis keturunan ibu (Browning et al., 1982, Giles et al.,1980). Sel telur memiliki jumlah kopi mtDNA yang tinggi (≥100000) sementara sel sperma memiliki jumlah kopi mtDNA yang rendah (100-1500) (Chen et al., 1995, Manfredi et al., 1997) dan terdapat paling banyak pada bagian ekornya. Dalam sel sperma mitokondria banyak terkandung dalam bagian ekor karena bagian ini yang sangat aktif bergerak sehingga membutuhkan banyak ATP. Pada saat terjadi pembuahan sel telur, bagian ekor sperma dilepaskan sehingga hanya sedikit atau hampir tidak ada mtDNA yang masuk ke dalam sel telur. Karena tidak terjadi rekombinasi,
maka mtDNA bersifat haploid, diturunkan dari ibu ke seluruh keturunannya (Cann et al., 1987, Giles et al., 1980, Wallace, 1997).
Gambar II.7 Pola pewarisan mtDNA
Gambar ini menunjukkan bagaimana mtDNA (berwarna biru) diwariskan dari ibu ke seluruh anaknya. Lelaki direpresentasikan oleh kotak dan perempuan direpresentasikan oleh lingkaran. (Cann et al., 1987, Giles et al., 1980, Wallace, 1997).
II.2.4.2 Laju Mutasi mtDNA
DNA mitokondria juga bersifat unik dan berbeda dengan DNA inti karena memiliki laju mutasi yang tinggi, yaitu sekitar 10-17 kali DNA inti (Wallace, et al., 1997). Hal ini disebabkan karena mtDNA tidak memiliki mekanisme reparasi yang efisien [Bogenhagen, 1999], tidak memiliki protein pelindung seperti histon, dan terletak berdekatan dengan membran dalam mitokondria tempat berlangsungnya reaksi fosforilasi oksidatif yang menghasilkan radikal oksigen sebagai produk samping (Richter, 1988). Tidak adanya aktivitas ini menyebabkan mtDNA tidak memiliki sistem perbaikan yang dapat menghilangkan kesalahan replikasi. Replikasi mtDNA yang tidak akurat ini akan menyebabkan mutasi mudah terjadi. Dalam mitokondria sendiri, daerah D-loop memiliki laju mutasi
yang lebih tinggi dibandingkan dengan daerah pengode karena pada posisi inilah awal replikasi dimulai (Howell et al., 1996).
II.2.4.3 Kode Genetik mtDNA
Salah satu bentuk keunikan lain dari mtDNA adalah kode genetik mitokondria menunjukkan perbedaan dalam hal pengenalan kodon yang berbeda dengan kode universal. Perbedaan ini ditunjukkan pada II.1
Tabel II.1 Perbedaan kodon mtDNA manusia dengan kodon universal. Universal MtDNA Manusia
UGA STOP Triptofan
AUA Isoleusin Metionin
AUU Isoleusin Metionin
AGG Arginin STOP
AGA Arginin STOP
Ada tiga jenis mutasi yang dapat tejadi pada DNA mitokondria, yaitu substitusi, delesi, dan inseri. Substitusi terjadi ketika ada perubahan basa pada nukleotida. Substitusi terbagi menjadi dua jenis, yaitu (1) substitusi transisi, yaitu perubahan nukleotida menjadi nukleotida lain yang jenis basanya sama, dari purin menjadi purin atau dari pirimidin menjadi pirimidin, misalnya A G, C T, dan sebaliknya, (2) substitusi transverse, yaitu perubahan nukleotida menjadi nukleotida lain yang jenis basanya berbeda, dari purin menjadi pirimidin atau sebaliknya, misalnya dari A C dan C G. Insersi terjadi akibat adanya penyisipan nukleotida, sedangkan delesi terjadi akibat adanya pengurangan nukleotida.
DNA polimerase untuk replikasi DNA mitokondria adalah DNA polimerase yang tidak mempunyai aktivitas proofreading (eksonuklease). Tidak adanya aktivitas ini menyebabkan mtDNA tidak memiliki sistem perbaikan yang dapat menghilangkan kesalahan replikasi. Replikasi mtDNA yang tidak selalu akurat ini akan menyebabkan mutasi mudah terjadi. Mutasi ini akan diturunkan dari satu generasi ke generasi selanjutnya sehingga semakin jauh hubungan kekerabatan antara dua individu, makin besar pula jumlah perbedaan mutasi. Variasi basa atau polimorfisme yang disebabkan oleh mutasi ini disebut dengan Single Nucleotide
Polymorphism (SNP). SNP, yang dapat terjadi pada daerah pengkode (coding region) maupun daerah bukan pengkode (noncoding region), misalnya pada daerah D-loop, dapat digunakan untuk membedakan satu individu dengan individu lain.
II.3 Polymerase Chain Reaction
PCR merupakan teknik in vitro untuk DNA yang dibatasi oleh sepasang primer (oligonukleotida pendek) menggunakan enzim 17 DNA polimerase dan dNTP sebagai monomernya [Newton dan Graham, 1997; Innis dan Gelfand, 1990]. Pada umumnya proses PCR berlangsung dalam tiga tahap yaitu: (1). Denaturasi, yaitu pemisahan DNA untai ganda menjadi untai tunggal karena terjadi pemutusan ikatan hidrogen basa-basanya pada suhu tinggi (94-96 °C). (2). Annealing, yaitu tahap penempelan primer pada templat DNA. Suhu annealing dapat dihitung berdasarkan nilai melting temperatur (Tm) dari primer-primer yang digunakan. (3). Extension, yaitu tahap reaksi polimerisasi oleh enzim DNA polimerase menggunakan dNTP sebagai monomernya dan dimulai dari ujung 3’ primer sepanjang DNA templatnya hingga terbentuk untai DNA baru. Tahap ini berlangsung pada temperatur saat enzim DNA polimerase bekerja optimal. Waktu yang dibutuhkan pada tahap ekstensi tergantung pada panjang fragmen yang diamplifikasi dan kecepatan reaksi (processity) dari enzim DNA polimerase yang digunakan [Cheng dan Kolmodin, 1997]. Ketiga tahap tersebut merupakan siklus yang berlangsung secara terus menerus. Untuk menghasilkan produk yang banyak dibutuhkan sekitar 25-30 siklus. Secara teori jumlah fragmen DNA yang dihasilkan selama n siklus PCR dirumuskan dengan (2
n
– 2n)x, dimana n = jumlah siklus, dan x = jumlah templat DNA [Newton dan Graham, 1997; Innis dan Gelfand, 1990].
II.4 Direct Sequencing dengan Metode Dideoksi-Sanger
Direct sequencing adalah proses sekuensing menggunakan templat DNA hasil PCR secara langsung tanpa melalui proses kloning. Dideoksi Sanger adalah metode penentuan urutan nukleotida yang didasarkan pada terminasi basa spesifik saat dilakukan sintesis DNA secara in vitro oleh enzim DNA polimerase
menggunakan satu primer. Basa spesifik yang digunakan adalah ddNTP yaitu dideoksinukleosida trifosfat yang tidak memiliki gugus hidroksil pada karbon 3’nya. Hilangnya gugus hidroksil ini menyebabkan DNA polimerase tidak dapat membentuk ikatan fosfodiester dengan dNTP atau ddNTP berikutnya, sehingga terjadi proses terminasi sintesis rantai DNA setelah reaksi dengan ddNTP. Terminasi berlangsung secara acak sehingga dihasilkan untai DNA yang panjangnya berbeda [Newton dan Graham, 1997]. Elektroforesis gel poliakrilamida berfungsi untuk memisahkan masing-masing fragmen berdasarkan ukuran nukleotidanya. 18 Detektor akan mendeteksi masing-masing fragmen berlabel dengan bantuan program komputer sehingga akhirnya bisa disusun urutan nukleotidanya.
Beberapa faktor mempengaruhi kualitas hasil sekuensing yaitu jumlah templat DNA, kemurnian DNA, kualitas primer, serta kontaminan seperti EDTA, fenol, dan kadar garam yang tinggi. EDTA pada konsentrasi di atas 0,5 mM dapat mengganggu ion Mg
2+
sebagai kofaktor enzim DNA polimerase. Adanya fenol dapat mengganggu dye fluorescent. Konsentrasi garam yang tinggi dapat menginhibisi enzim [Robertson, 1996]. Direct sequencing telah banyak digunakan dalam berbagai bidang penelitian misalnya bidang forensik, antropologi, dan medis.