TINJAUAN PUSTAKA
Karakterstik Wilayah Penelitian
Geografi dan Wilayah Administratif
Provinsi Maluku Utara merupakan hasil pemekaran dari wilayah provinsi Maluku. Ibukota Provinsi Maluku Utara yang definitif adalah di Sofifi. Mempertimbangkan berbagai aspek daya dukung prasarana dan sarana pemerintahan yang ada di Sofifi belum memadai untuk menjalankan pemerintahan, maka dalam rangka menjalankan roda pemerintahan provinsi, untuk sementara ditempatkan di kota Ternate dan berjalan sampai dengan saat ini.
Secara geografis wilayah Provinsi Maluku Utara berada pada posisi koordinat 30 Lintang Utara sampai 30 Lintang Selatan dan 1240 sampai 1290 Bujur Timur, dengan batas – batas wilayah sebagai berikut :
• Sebelah Timur berbatasan dengan Laut Halmahera • Sebelah Barat berbatasan dengan Laut Maluku • Sebelah Utara berbatasan dengan Samudera Pasifik • Sebelah Selatan berbatasan dengan Laut Seram (Maluku).
Luas total wilayah provinsi Maluku Utara mencapai 140 255.36 km2,
dengan luas wilayah perairan 106 977.32 km2 (76.27%), dan daratan seluas 33
278 km2 (23.73 %). Terdiri dari 395 buah pulau besar dan kecil. Dari jumlah itu, sebanyak 64 pulau telah di huni, sedangkan 331 pulau lainnya tidak dihuni. Jumlah penduduk tahun 2004 sebanyak 910 656 jiwa, rata-rata laju pertumbuhan sebesar 2.16% per tahun.
Sebagai wilayah kepulauan, provinsi Maluku Utara terdiri dari pulau besar dan pulau kecil. Pulau yang tergolong relatif besar ialah Pulau Halmahera (18 000 km2), Pulau-pulau yang relatif sedang besar ialah Pulau Obi (3 900 km2), Pulau Taliabu (3 295 km2), Pulau Bacan (2 878 km2) dan Pulau Morotai (2 325 km2) dan pulau – pulau yang relatif kecil antara lain pulau Ternate, Tidore, Moti, Makian, Kayoa, Gebe dan sebagainya.
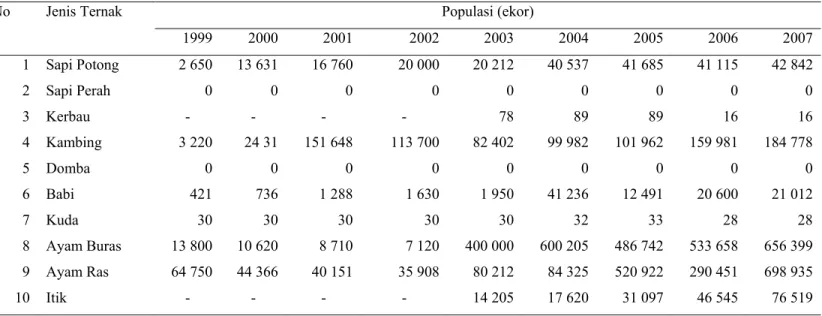
Secara administratif pemerintahan, provinsi Maluku Utara terdiri dari 6 Kabupaten dan 2 Kota, yaitu Kabupaten Halmahera Tengah, Kota Ternate, Kabupaten Halmahera Barat, Kabupaten Halmahera Utara, Kabupaten Halmahera Selatan, Kabupaten Kepulauan Sula, Kabupaten Halmahera Timur, dan Kota Tidore Kepulauan (Tabel 1).
Tabel 1 Luas wilayah kabupaten/kota di provinsi Maluku Utara No. Kabupaten/Kota Jumlah
kecamatan Jumlah desa/kelurahan Desa Kelurahan Luas wilayah (km2)
1 2 3 4 5 6 1. 2. 3. 4. 5. 6. 7. 8. Halmahera Tengah Kota Ternate Halmahera Barat Halmahera Utara Halmahera Selatan Kepulauan Sula Halmahera Timur Tidore Kepulauan 3 4 5 9 9 6 4 6 31 - 130 174 194 86 41 21 - 63 - - - - - 20 2 276.65 1 122.98 2 897 575 10 493.73 8 977 497 4 977 497 6 506.20 1 797.18 Total 45 676 83 140 255.36
Sumber : Bappeda Provinsi Maluku Utara (2006)
Geologi dan Fisiografi
Berdasarkan struktur dan tektonik serta litologinya, geologi sebagian besar Provinsi Maluku Utara bagian Tengah dan Utara merupakan daerah pegunungan dengan bahan induk bervariasi. Bagian Utara dan Timur Laut semenanjung Halmahera didominasi oleh pegunungan, semenanjung Utara disusun oleh formasi gunung api (Andesit dan bahan batuan beku Andesit). Pada semanjung Timur Laut ditemukan batuan beku asam, basa, dan ultra basa serta bahan sedimen.
Di semenanjung utara Halmahera terdapat barisan gunung api aktif dengan bentuk dan struktur yang sangat khas. Pada bagian ini, dataran alluvial tidak ditemukan, tetapi memasuki daerah Kao ditemukan dataran alluviasi yang luas pada daerah pedalaman, dataran vulkanik yang berombak dan dataran berawa secara lokal. Pulau Morotai memiliki banyak kesamaan dengan Pulau Halmahera bagian Utara dan Timur yang dicirikan oleh gunung-gunung yang berkembang dari batuan sediment dan batuan beku basa. Pada semenanjung bagian Selatan Halmahera lebih didominasi oleh daerah gunung yang terutama berkembang dari
bahan-bahan sedimentasi dan batu gamping , dimana bagian ini terbentang dataran sempit alluvial arah Timur-Barat.
Kawasan sepanjang pantai Barat Halmahera terbentang sejumlah pulau-pulau besar dan kecil yang dimulai dari pulau-pulau Ternate bagian Utara sampai Obi di bagian Selatan. Pulau-pulau kecil di bagian Utara umumnya merupakan daerah vulkanik yang tersusun dari bahan andesit, dan batuan beku basaltik dengan lereng curam (30 – 45 %) sampai sangat curam (> 45%).
Kelompok pulau-pulau Bacan mempunyai bentangan lahan pegunungan yang sama dengan Halmahera Utara yaitu batuan beku basa dan batuan metamorfik. Batuan metamorfik walaupun menyebar secara lokal tetapi merupakan batuan induk dominan pada daerah ini. Sepanjang pesisir terdapat dataran pantai yang sempit, dan bagian tengah dari pusat pulau Bacan dibentuk oleh daratan alluvial.
Bentang lahan pulau Obi mengikuti pola yang sama, dimana bagian tengah didominasi oleh daerah pegunungan dengan bahan penyusunnya batuan beku basa dan diapit oleh deretan perbukitan dari batuan sediment.
Kelompok kepulauan Sulabesi mempunyai struktur yang sama tetapi memiliki susunan bahan induk yang berbeda sebagian besar pulau. Taliabu dan Pulau Sanana merupakan daerah pegunungan dengan puncak tajam dan lereng yang curam, berkembang terutama dari batuan metamorfik. Bagian Barat pulau Sanana juga ditemukan bahan induk granit.
Topografi
Provinsi Maluku Utara dibentangkan oleh relief-relief besar dimana palung Oceanis dan punggung pengunungannya saling bergantungan dengan kemiringan lahannya. Sebagian besar bergunung – gunung dan berbukit – bukit yang terdiri dari pulau – pulau Vulkanis dan pulau karang, sedangkan sebagian lainnya merupakan hamparan dataran. Pulau Halmahera mempunyai banyak pegunungan yang rapat mulai dari teluk Kao, teluk Buli, teluk Weda, teluk Payahe dan Dodinga. Di setiap daerah terdapat punggung gunung yang merapat ke pesisir, sedangkan pada daerah sekitar teluk Buli, pesisir barat mulai dari
teluk Jailolo ke Utara dan teluk Weda ke Selatan ditemui daerah hamparan dataran yang luas. Pada bagian lainnya terdapat deretan pegunungan yang melandai dengan arah pesisir. Pulau – pulau yang relatif sedang (Obi, Morotai, Taliabu dan Bacan) umumnya memiliki dataran luas yang diselingi pegunungan yang bervariasi.
Jenis Tanah
Tanah yang terdapat di wilayah provinsi Maluku Utara menunjukkan sifat–sifat yang berbeda, mulai dari pulau Morotai di bagian Utara sampai pulau Sulabesi di bagian selatan perbedaan ini disebabkan oleh faktor klimatologi (curah hujan, suhu dan angin ) yang tinggi.
Selain itu, yang membedakan sifat-sifat tanah adalah tipe batuan/bahan induk dan kemiringan lereng yang berkorelasi dengan kedalaman efektif perakaran serta vegetasi dimana tanah itu berkembang. Selain iklim dan vegetasi, kompleks geologi provinsi Maluku Utara sangat erat hubungannya dengan penyebaran sifat-sifat tanah. Keadaan geologi dibarengi pula dengan proses pelapukan dan pencucian di bawah kondisi suhu dan curah hujan yang bervariasi. Oleh karena itu, tanah di daerah Maluku Utara berada dalam suatu perkembangan dan kedalaman yang bervariasi dengan drainasi baik, tekstur tanah halus, kesuburan yang relatif rendah pada daerah-daerah perbukitan dan pegunungan yang berlereng curam sampai sangat curam dengan penutupan vegetasi yang jarang. Ini secara relatif juga mempengaruhi erosi permukaan, sehingga sering ditemukan tanah-tanah dengan kedalaman solum dangkal sampai sedang dengan tingkat perkembangan lemah sampai sedang. Adapun jenis tanah yang tersebar di daerah Maluku Utara antara lain, adalah:
1. Jenis tanah mediteran terdapat di pulau Morotai bagian Barat, Timur dan Selatan, pulau Doi, dan kecamatan Loloda .
2. Jenis tanah podsolik merah kuning terdapat di pulau Halmahera dari Utara ke Selatan, Tobelo, Ibu, Obi, bagian Timur, Sanana, pulau Taliabu, Oba, Weda, Patani dan Maba.
3. Jenis tanah kompleks terdapat di pulau Morotai bagian Barat dan Timur ,Obi bagian tengah , pulau Halmahera bagian Tengah sampai Timur.
4. Jenis tanah latosol terdapat di Loloda, Jailolo bagian Selatan, Gane Timur, Gane Barat, Bacan, Oba, Wasile, Weda dan Maba
5. Jenis tanah regosol terdapat Loloda, Galela, Sahu, Kao, pulau Ternate, pulau Makian, Pulau Obi di pesisir Utara .
6. Jenis tanah alluvial terdapat di pulau Obi bagian Barat, pulau Taliabu, bagian Utara dan Tenggara, Oba, Wasile, Weda, Patani dan Maba.
Klimatologi
Secara Umum iklim di kepulauan Maluku Utara hampir sama. Temperatur rata-rata tahunan yang diukur dari stasiun Duma Galela, Ternate dan Tobelo antara 25.60 C – 26.10 C dengan curah hujan rata-rata tahunan antara 2.138 mm – 3.693 mm.
Wilayah Maluku Utara dipengaruhi oleh iklim laut tropis dan iklim musim. Oleh karena itu iklimnya sangat dipengaruhi oleh lautan dan bervariasi antara tiap bagian wilayah, yaitu Halmahera Utara, Halmahera Tengah/Barat, Bacan dan Kepulauan Sula.
a. Daerah iklim Halmahera Utara terdiri atas dua musim yaitu : ● Musim hujan pada bulan Desember sampai Februari
● Musim kemarau pada bulan Agustus sampai dengan bulan Desember, yang diselingi pancaroba pada bulan November – Desember.
b. Daerah iklim Halmahera Tengah/Barat yang dipengaruhi oleh dua musim yaitu :
● Musim Utara pada bulan Oktober – Maret dan musim pancaroba pada bulan April.
● Musim Selatan pada bulan April – September yang diselingi angin Timur dan pancaroba pada bulan September.
c. Daerah iklim Bacan yang dipengaruhi oleh dua musim yaitu :
● Musim Utara pada bulan Oktober – Maret yang diselingi angin Barat dan pancaroba pada bulan April.
● Musim Selatan pada bulan September diselingi angin Timur dan pancaroba pada bulan September.
d. Daerah iklim kepulauan Sula yang terdiri dari dua musim yaitu :
● Musim Utara pada bulan Oktober – Maret diselingi angin barat dan pancaroba pada bulan April.
Sebagaimana umumnya daerah Maluku Utara yang didominasi wilayah laut. Kota Ternate sangat dipengaruhi oleh iklim laut karena mempunyai tipe iklim tropis yang terdiri dari dua musim (Utara-Barat dan Timur-Selatan), yang seringkali diselingi dengan dua kali masa pancaroba disetiap tahunnya. Temperatur rata bulanan 26.96°C (maksimum 30.68 °C – minimum 24.12 °C), dengan rata-rata kelembaban 86.42 % dan penyinaran matahari 54.42 % dengan kecepatan angin 4.25 km/jam.
Berdasarkan klasifikasi iklim Schimdt dan Ferguson, daerah Maluku Utara umumnya bertipe iklim B, dengan rata-rata curah hujan per tahun 1869.4 mm. Bulan basah adalah bulan dengan curah hujan lebih tinggi atau sama dengan 1000 mm dan bulan kering adalah bulan dengan curah hujan lebih rendah atau sama dengan 600 mm. Bulan November dan bulan Agustus adalah bulan dengan curah hujan yang tertinggi selain itu bulan April juga bulan dengan curah hujan yang tinggi yaitu 293.3 mm (Tabel 2). Periode curah hujan rendah berlangsung pada bulan September dan Oktober dengan curah hujan terendah 50.8 mm pada bulan September lihat Tabel 3.
Adapun curah hujan di propinsi Maluku Utara adalah sebagai berikut :
● Curah hujan antara 1000 – 2000 mm, meliputi Pulau Tobelo, Pulau Mangole, Pulau Sulabesi, Pulau Obi dan sekitarnya, Pulau Bacan dan sekitarnya dan Pulau Halmahera bagian Selatan.
● Curah hujan antara 2500 – 3000 mm, meliputi Pulau Halmahera bagian Utara, sebagian Kecamatan Ibu, Galela dan Loloda.
Tabel 2 Curah hujan, bulan basah dan bulan kering di provinsi Maluku Utara
Kabupaten Curah Hujan per Tahun Pola Tipe Iklim Bulan Hujan Kota Ternate 3000 - 4000 Berfluktuasi Basah CH ≤ 100 = 0 mm/bln
CH ≤ 100 - 150 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kota Tidore Kepulauan 2000 - 3000 Ganda
(double wafe) Basah CH CH ≤ 100 - 150 = ≤ 4 mm/bln ≤ 100 = ≤ 4 mm/bln CH ≤ 150 - 200 = ≤ 5 mm/bln CH > 200 = 6 – 8 mm/bln Kabupaten Halmahera Barat 3000 - 4000 Berfluktuasi Basah CH ≤ 100 = 0 mm/bln
CH ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kabupaten Halmahera Utara 1000 - 2000 Berfluktuasi
(multiple wafe) Kering CH CH ≤ 100 = 0 mm/bln ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kabupaten Halmahera Timur 1000 - 2000 Berfluktuasi
(multiple wafe) Kering CH CH ≤ 100 = 0 mm/bln ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kabupaten Halmahera Tengah 2000 - 3000 Ganda
(double wafe) Basah CH CH ≤ 100 = 0 mm/bln ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kabupaten Halmahera Selatan 2000 - 3000 Ganda
(double wafe) Basah CH CH ≤ 100 = 0 mm/bln ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln Kabupaten Kepulauan Sula 1000 - 2000 Berfluktuasi
(multiple wafe) Kering CH CH ≤ 100 = 0 mm/bln ≤ 100 = 0 mm/bln CH ≤ 150 - 200 = ≤ 2 mm/bln CH > 200 = 9 – 12 mm/bln
Tabel 3 Suhu udara, kelembaban dan rataan penyinaran matahari pada stasiun meteorologi Babullah Ternate 2005
No.
Bulan Suhu Udara (
OC) Kelembaban (%) Penyinaran Matahari Rata-rata (Jam) Maks Min Rataan
1. Januari 30.1 24.9 26.9 83 41 2. Pebruari 30.5 23.9 26.5 84 62 3. Maret 31.4 24.5 27.2 82 68 4. April 30.6 24.3 26.9 85 56 5. Mei 31.3 24.0 27.0 84 61 6. Juni 30.8 24.1 27.0 83 59 7. Juli 30.0 24.0 27.0 82 41 8. Agustus 31.1 24.0 27.0 77 59 9. September 31.4 23.7 27.1 77 71 10. Oktober 30.8 23.9 26.9 90 49 11. Nopember 30.0 24.0 27.0 86 44 12. Desember 27.0 30.2 27.0 86 37
Sumber : Badan Pusat Statistik Maluku Utara (2006)
Keadaan Umum Peternakan Provinsi Maluku Utara
Pembangunan pertanian termasuk didalamnya sub sektor peternakan sebagai bagian integral dari Pembangunan Daerah mempunyai peranan yang strategis dalam pemulihan ekonomi daerah. Peranan strategis tersebut khususnya adalah dalam peningkatan pendapatan daerah, penyediaan pangan, penyediaan bahan baku industri, peningkatan ekspor, penyediaan kesempatan kerja dan kesempatan berusaha, peningkatan pendapatan petani dan kesejahteraan masyarakat.
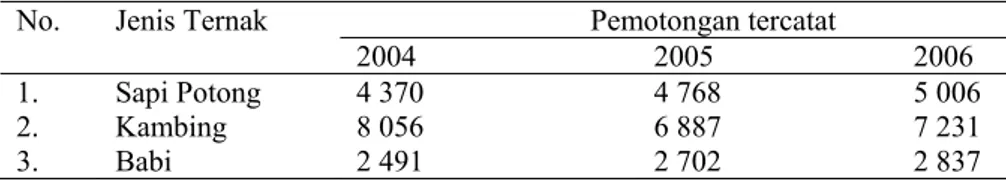
Namun di era pasca konflik yang bernuansa sara diikuti dengan krisis moneter yang dialami bangsa Indonesia, kondisi subsektor peternakan di wilayah Maluku Utara terkena imbasnya yang mengakibatkan rusaknya sarana prasarana peternakan baik milik pemerintah maupun aset-aset peternakan yang dimiliki masyarakat di daerah ini. Puluhan ribu ternak hilang, baik ternak ruminansia ( sapi dan kambing), maupun ternak unggas (ayam ras, buras dan itik). Hal ini dapat dilihat dari jumlah populasi ternak dari tahun 1999 – 2007 di Maluku Utara pada Tabel 4.
Tabel 4 Populasi ternak tercatat (ekor) pada 1999 – 2007 di provinsi Maluku Utara
No Jenis Ternak Populasi (ekor)
1999 2000 2001 2002 2003 2004 2005 2006 2007 1 Sapi Potong 2 650 13 631 16 760 20 000 20 212 40 537 41 685 41 115 42 842 2 Sapi Perah 0 0 0 0 0 0 0 0 0 3 Kerbau - - - - 78 89 89 16 16 4 Kambing 3 220 24 31 151 648 113 700 82 402 99 982 101 962 159 981 184 778 5 Domba 0 0 0 0 0 0 0 0 0 6 Babi 421 736 1 288 1 630 1 950 41 236 12 491 20 600 21 012 7 Kuda 30 30 30 30 30 32 33 28 28 8 Ayam Buras 13 800 10 620 8 710 7 120 400 000 600 205 486 742 533 658 656 399 9 Ayam Ras 64 750 44 366 40 151 35 908 80 212 84 325 520 922 290 451 698 935 10 Itik - - - - 14 205 17 620 31 097 46 545 76 519
Sumber : data sekunder rangkuman berbagai sumber
Dari data populasi pada Tabel 4 terjadi pertambahan populasi yang signifikan pada ternak sapi tahun 2004, hal ini disebabkan beberapa program pemerintah untuk pengadaan ternak sapi di tahun 2003 melalui program peningkatan ketahanan pangan melalui sub kegiatan Inpres No. 6 tahun 2003 dan Program Peningkatan kesejahteraan petani di provinsi Maluku Utara. Hal yang sama juga dilakukan terhadap ternak kambing pada tahun 2001 sampai dengan tahun 2003, ternak unggas (ayam ras, buras dan itik) terjadi pertambahan populasi yang sangat signifikan pada tahun 2003. Jumlah populasi ternak sapi dan kambing terendah terjadi pada tahun 1999, hal ini disebabkan karena konflik yang terjadi di masyarakat, dimana pada tahun 1998 untuk ternak sapi di Maluku Utara sebanyak 86 213 ekor, sedangkan ternak kambing sebayak 133 261 ekor, penurunan populasi yang sangat drastis terjadi di tahun 1999 untuk kedua jenis ternak ini disebabkan karena peternak membiarkan ternak sebagian masuk ke hutan dan sebaian besar dibawa ke provinsi Sulawesi Utara.
Tingkat pertumbuhan populasi pertahun dari masing-masing ternak di provinsi Maluku Utara dapat disajikan pada Tabel 5.
Tabel 5 Tingkat pertumbuhan populasi ternak (2000 -2005) No. Jenis Ternak Tingkat Pertumbuhan pertahun (%)
1. Sapi potong 25.05
2. Kambing 33.2
3. Babi 76.17
4. Ayam Buras 114.8
5. Ayam Ras 63.6
Sumber : Dinas Pertanian dan Ketahanan Pangan Daerah Prov. Maluku Utara (2006)
Tingkat pemotongan untuk beberapa komoditas ternak yang tercatat pada tahun 2004-2006 disajikan pada Tabel 6.
Tabel 6 Pemotongan ternak tercatat (ekor) tahun 2004-2006 No. Jenis Ternak Pemotongan tercatat
2004 2005 2006 1. 2. 3. Sapi Potong Kambing Babi 4 370 8 056 2 491 4 768 6 887 2 702 5 006 7 231 2 837 Sumber : Dinas Pertanian dan Ketahanan Pangan Daerah Prov. Maluku
Sedangkan untuk tingkat produksi/hasil peternakan pada tahun 2005 dapat di lihat pada Tabel 7.
Tabel 7 Produksi hasil ternak di Maluku Utara (2005)
No. Jenis Produksi Jumlah (kg) 1. 2. 3. 4. 5. 6. 7. Daging Sapi Daging kambing Daging Ayam Buras Daging Ayan Ras Telur Ayam Ras Telur Ayam Buras Telur Itik 978 751 344 350 705 776 3 334 943 57 672 204 432 145 532 Sumber : Dinas Pertanian dan Ketahanan Pangan Daerah (2006)
Pola peternakan di wilayah Maluku Utara untuk semua komoditas ternak sebagian besar (80-90 persen) masih bertahan dalam bentuk usaha rakyat. Pola peternakan rakyat ini mempunyai ciri-ciri antara lain (Yusdja 2004), tingkat pendidikan rendah, pendapatan rendah, penerapan manajemen dan teknologi konvensional, lokasi ternak menyebar luas, ukuran skala usaha relatif sangat kecil, penggunaan tenaga kerja keluarga, penguasaan lahan hijauan makanan ternak (HMT) yang terbatas.
Kambing Liar dan Kambing Domestikasi
Kambing tergolong genus Capra. Ellerman dan Morrison-Scott (1951) yang dikutip oleh Devandra dan Burns (1970) membagi genus ini atas lima spesies yaitu Capra hircus (termasuk bezoar), Capra ibex, Capra caucasica, Capra pyrenaica (Ibex Spanyol) dan Capra falconeri. Menurut Herre dan Rohrs (1973) yang dikutip oleh Devendra dan Nozawa (1976) kambing liar (Capra aegagrus) yang hidup dan menyesuaikan diri terutama di lingkungan pegunungan dan lingkungan yang agak kering (semi-arid) dapat dibagi atas tiga spesies yaitu: - Bezoar atau Psaang (C.a.aegagrus) yang hidup liar di Asia Barat,- Ibex (C.a..ibex) hidup di Asia Barat, Afrika Timur dan Eropah, - Markhol (C.a.falconeri) hidup liar di Afganistan dan Kashmir – Karakorum. Tiap spesies terdiri dari beberapa
sub spesies karena terpisah secara geografik, namun persilangan antar sub spesies adalah subur.
Menurut Murtidjo (1993), umumnya kambing merupakan hewan yang hidup di lereng-lereng pegunungan, bukit-bukit yang curam ataupun tempat-tempat yang curam, selain tempat-tempat yang tandus dan sedikit ditumbuhi rumput atau tanaman. Kambing yang kita kenal sekarang merupakan hasil domestikasi manusia yang diturunkan dari tiga jenis kambing liar, yaitu: Capra hircus, merupakan jenis kambing liar yang berasal dari daerah sekitar perbatasn Pakistan-Turki; Capra falconeri, merupakan jenis kambing liar yang berasal dari daerah sepanjang Kashmir, India; Capra prisca, merupakan jenis kambing liar yang berasal dari daerah sepanjang Balkan. Dari ketiga jenis kambing liar tersebut, kini dikenal beberapa bangsa kambing yang tersebar di seluruh dunia, seperti: kambing Kacang, kambing Etawah, kambing Saanen, kambing Kashmir, kambing Angora, kambing Toggenburg, Nubian dan lain-lain.
Penyebaran Kambing ke Asia Tenggara
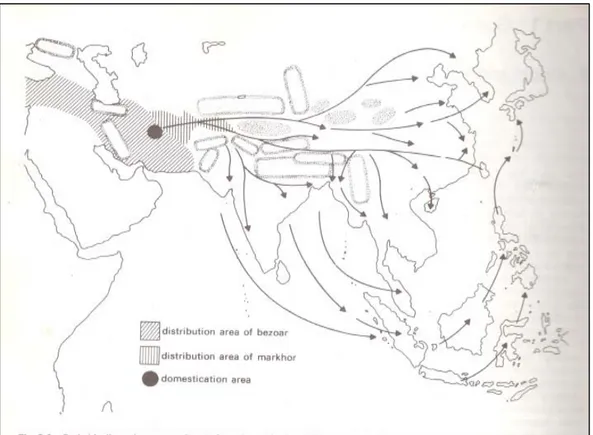
Devendra dan Nozawa (1976) mengemukakan bahwa kambing piara dari Asia Barat menyebar ke Timur melalui dua jalan utama. Pertama, dari Persia dan Afganistan melalui Turkestan ke Mongolia atau Cina Utara, yang dinamakan lintasan sutera, yang terjadi pada sekitar 2000 tahun sebelum Masehi. Kedua, kearah anak benua India melalui Khyber Pass. Jalan ini sangat tua, yaitu sejak orang Indo-Aryan mengetahuinya pada sekitar 2000 tahun sebelum Masehi. Dengan demikian, Mongolia, Cina dan India menerima kambing piara dari Barat dengan perantaraan para pengembara.
Gambar 2 menunjukkan jalan migrasi kambing asli Asia dari wilayah penjinakan. Jalan ini diduga atas konfirmasi dari peninggalan-peninggalan lama dari hasil penelitian. Dari anak benua India, kambing piara ini menyebar ke pulau Sumatera, Jawa, Kalimantan, Sulawesi, Philipina dan terus ke Jepang. Thailand menerima kambing piara dari Utara. Perkembangan selanjutnya, kambing pendatang ini menjadi kambing asli di wilayah penyebarannya dan di Indonesia
kita kenal dengan kambing kacang tersebar di pulau-pulau Indonesia terutama Jawa dan Sumatera.
Kambing-kambing asli yang sekarang terdapat di Negara di kawasan Asia Tenggara berasal dari turunan bangsa kambing yang sama dapat dilihat terutama dari kesamaan morfologinya, terutama warna bulu. Warna bulu coklat dengan garis punggung berwarna hitam (ciri dari Bezoar) dan warna bulu hitam merupakan jumlah terbanyak dari kambing asli di Malaysia Barat dan Timur (Shotake et al. 1976). Hal yang serupa juga terdapat di Philipine, di Tahiland dan Taiwan. Penelitian yang dilakukan oleh Abdulgani et al. (1981) di Sumatera Barat, Jawa Barat dan Sulawesi Selatan menunjukkan bahwa kambing yang berwarna bulu coklat dan hitam merupakan jumlah terbanyak.
Murtidjo (1993) menyatakan bahwa kambing kacang merupakan bangsa kambing lokal asli Indonesia. Tubuh kambing kacang relatif kecil, kepala ringan dan kecil, telinga pendek dan tegak lurus mengarah ke atas depan. Kehidupannya sangat sederhana, memiliki daya adapasi yang tinggi terhadap kondisi alam setempat dan reproduksinya dapat digolongkan sangat tinggi. Jenis kambing ini juga terdapat di Filipina, Myanmar, Thailand, Malaysia dan sekitarnya. Kambing tipe kecil yang disebut kambing kacang merupakan kambing pendatang pertama di Malaysia dari India dan akhirnya menjadi kambing asli Malaysia (Devendra 1966).
Selanjutnya dikatakan bahwa kambing yang tipenya serupa telah menyebar di bagian lain dari Asia Tenggara sampai Taiwan dan kepulauan Jepang bagian Selatan. Imigrasi orang-orang Pakistan ke Thailand menyertakan kambing tipe dwiguna sehingga menyebabkan variabilitas genetik yang tinggi pada kambing-kambing asli Thailand, namun tidak dapat dikesampingkan kemungkinan telah terjadi persilangan dengan bangsa-bangsa kambing Eropah seperti Seaanen atau lainnya.
Gambar 2 Postulat penyebaran kambing piara ( ●) ke Asia Timur dan Tenggara Sumber Devendra dan Nozawa (1976)
Klasifikasi Bangsa Kambing
De Haas dan Horst (1979) mengelompokkan kambing atas tiga tipe berdasarkan tinggi pundak dan bobot badan hidup (Tabel 8).
Tabel 8 Klasifikasi bangsa kambing (dewasa) menurut tinggi Pundak dan Bobot badan Hidup
Tipe kambing Tinggi pundak (cm) Bobot badan hidup (kg) Besar
Sedang
Kecil (dan Kerdil)
65 50-65 50 30-60 20-45 9-30 Sumber : De Haas dan Horst (1979)
Fungsi utama kambing tipe kecil adalah penghasil daging, tipe sedang untuk penghasil daging dan susu, sedangkan tipe besar ditujukan untuk penghasil
susu. Tipe kerdil (dwarf) sama sekali tidak ideal sebagai penghasil daging karena pertumbuhannya yang sangat lambat.
Chai (1961) dikutip Gunawan (1982) menyatakan bahwa peningkatan bobot badan dapat dilakukan melalui kawin silang dengan bangsa lainnya yang superior karena gen-gen yang mengendalikan ukuran-ukuran badan yang lebih besar dominant terhadap gen-gen yang mengendalikan ukuran-ukuran badan yang lebih kecil.
Berdasarkan tinggi pundak dan bobot badan Devendra dan Burns (1970) menyimpulkan bahwa kambing yang tergolong tipe besar diantaranya adalah Jamnapari, Beetal, Barbari, Malabar, Damascus, Syrian Mounain, Sardinian, Benadir, Angora, Sahel, Maradi, Mudugh, Sudanese Nubian, Sudanese Shukria, Soviet Mohair dan Moxoto; yang tergolong tipe kecil diantaranya adalah Ma T’ou, kambing kacang, Kigezi, Arab angora, Melteze dan Moxoto; yang tergolong tipe kerdil (dwarf) di antaranya adalah , South China, Bengal, East African , South Sudan, Congo dwarf, West African Dwarf dan Kosi.
Selanjutnya dinyatakan bahwa faktor lingkungan sangat berpengaruh terhadap ukuran-ukuran serta bobot badan kambing. Dengan demikian suatu bangsa kambing yang tergolong tipe besar pada suatu lokasi akan tergeser ke tipe kecil pada lokasi lainnya, atau suatu bangsa kambing yang tergolong tipe kecil pada suatu lokasi akan tergeser ke tipe kerdil (dwarf) pada lokasi lainnya.
Beberapa Ciri Kambing Kacang
Menurut Murtidjo (1993), kambing kacang merupakan kambing lokal asli Indonesia. Tubuh kambing kacang relatif kecil, kepala ringan dan kecil, telinga pendek dan tegak lurus mengarah ke atas depan, dengan kehidupan yang sederhana, memiliki daya adaptasi yang tinggi terhadap kondisi alam setempat dan reproduksinya dapat digolongkan sangat tinggi. Jenis kambing ini juga terdapat di Filipina, Myanmar, Thailand, Malaysia dan sekitarnya.
Lebih lanjut dikatakan bahwa kambing kacang memiliki warna tunggal, yakni: putih, hitam dan coklat, serta adakalanya warna campur dari ketiga warna tersebut. Kambing Kacang kelamin jantan maupun betina mempunyai tanduk 8 –
10 cm. Berat tubuh kambing kacang dewasa rata-rata sekitar 17 – 30 kg. Betina umumnya memiliki bulu pendek pada seluruh tubuh, kecuali pada bagian ekor dan dagu. Gambaran beberapa ciri kambing kacang dapat disajikan pada Gambar 3.
Gambar 3 Jenis kambing kacang (Sumber dok: Hoda 2006)
Damshik (2001) mengemukakan bahwa kambing kacang berbadan relatif kecil dengan tinggi pundak dewasa rata-rata 50 cm dan bobot badan 30 kg. Bila dibandingkan dengan bagian-bagian lainnya maka kepala mempunyai proporsi yang sangat baik dan seimbang; ukuran telinga sedang, selalu bergerak, tidak tergantung tetapi tegak. Tanduk terdapat baik pada yang jantan maupun pada betina dan ukurannya relatif pendek. Janggut tumbuh dengan baik pada kambing jantan, namun juga terdapat pada yang betina dewasa walaupun tidak begitu lebat. Leher pendek dan memberi kesan tebal dan tegap. Punggung lurus dan pada beberapa kasus terlihat agak melengkung dan memeberi kesan makin kebelakang makin tinggi sampai pinggul. Devendra dan Burns (1970) menyatakan bahwa profil kambing kacang berbentk lurus. Ekor kelihatan kecil dan tegang. Ambing kecil dengan konformasi baik dengan puting yang besar. Bulu pendek serta kasar pada yang betina, tetapi pada yang jantan lebih panjang. Kambing kacang tahan hidup pada keadaan kondisi lingkungan yang sangat beragam dan sanggup beradaptasi pada metode manajemen yang berubah-ubah dan sangat beragam. Umur ketika mencapai pubertas sekitar enam bulan pada yang jantan. Umur beranak pertama dicapai ketika umur 12 – 13 bulan.
Siklus Reproduksi Ternak Kambing
Pada umumnya, ternak kambing mulai dewasa kelamin pada umur 5 – 10 bulan. Dewasa kelamin sangat tergantung dari rasa tau tipe, jenis kelamin dan lokasi pemeliharaan. Kambing tipe kecil lebih cepat mengalami dewasa kelamin dibandingkan kambing tipe besar. Perkawinan induk kambing betina sebaiknya dilakukan pada umur 9 – 12 bulan, karena pada umur ini secara fisik kambing sudah tumbuh dewasa sehingga mampu memproduksi susu dan menjalani masa kebuntingan. Menurut Devendra dan Burns (1970) menyatakan bahwa kebanyakan bangsa kambing daerah tropis biasa melahirkan pada umur satu tahun dan dapat digunakan sebagai produsen anak sampai kambing berumur 5 – 6 tahun. Umur dini pada beranak pertama mengurangi biaya pemeliharaan calon induk dan meningkatkan pendapatan ekonomi, serta menunjang perbaikan genetik yang cepat, dan oleh karenanya hal itu sangat diinginkan.
Siklus birahi seekor kambing betina antara 20 – 24 hari. Masa birahinya berlangsung selama 1 – 2 hari. Kambing betina tidak akan bunting bila dikawinkan dalam keadaan tidak sedang birahi. Kambing yang sedang bunting tidak mengalami masa birahi lagi. Mishra dan Biswas (1966) yang mempelajari penyebaran birahi pada kambing lokal di India, yang melibatkan 12 081 ekor kambing betina, menunjukkan bahwa rata-rata lama birahi sekitar 38 jam. Pretorius (1977) mempelajari usap vagina dari induk kambing Angora yang sedang mengalami siklus birahi dan yang tidak (anestrus) mencirikan perubahan yang terjadi. Selama birahi, terjadi aliran lendir jernih dan encer yang membentuk pola kristalisasi seperti pakis. Setelah ovulasi dan pada fase birahi akhir, lendir tersebut menjadi masa putih yang kental, mengandung banyak elemen sel bertanduk, sedangkan pada fase luteal dan anestrus ditandai dengan sekresi lender yang sedikit dan tidak membentuk pola kristalisasi.
Kambing pejantan bisa dikawinkan pada umur 10 bulan tetapi tidak dibiarkan melayani lebih dari 20 ekor induk betina sebelum umurnya genap satu tahun. Pada tenggang waktu dua bulan itu, kambing jantan hanya kawin 16 – 20 kali atau maksimal dua kali kawin dalam seminggu. Pejantan dapat digunakan sebagai pemacek sampai umur 7 – 8 tahun. Penjelasan mengenai perkembangan
reproduksi ternak kambing jantan telah banyak dilakukan. Skinner (1975) menyembelih pejantan kambing boer pada berbagai interval dari saat lahir sampai berumur 196 hari. Selama waktu itu, berat testis meningkat secara lambat 1.3 g pada saat lahir menjadi 9.9 g pada umur 84 hari, dan selanjutnya secara cepat menjadi 25 g pada umur 140 hari, ketika spermatozoa untuk pertama kali tersedia melalui saluran eferens. Spermatogenensis mulai pada umur 84 hari dan pada umur 120 hari, spermatozoa ada dalam epididimis.
Hasil penelitian menunjukkan bahwa lama bunting pada kambing ditemukan agak konstan sekitar 146 hari, meskipun kisaran yang dilaporkan antara 143 – 153 hari. Otchere dan Nimo (1975) mengatakan bahwa lama bunting pada kambing kerdil Afrika Barat dilaporkan rata-rata 141,3 ± 4.7 hari. Shelton (1978) menduga bahwa lama bunting yang singkat merupakan ciri bangsa kambing kecil. Penyebab keragaman dalam periode bunting tidak diketahui secara rinci, tetapi seperti pada spesies hewan lainnya, hal itu dipengaruhi oleh jenis kelamin janin, kondisi habis beranak, dan keragaman lingkungan lainnya, khususnya makanan, dan oleh faktor keturunan. Mishra et al. 1979 menemukan bahwa lama bunting rata-rata 146,42 ± 0.24 hari, dan berkorelasi secara kuat dengan berat lahir anak (0.33) dan berat induk saat dikawinkan (0.41).
Periode perkawinan merupakan faktor penentu interval beranak yang paling penting karena beda dalam lama bunting kecil dan manfaat praktisnya sedikit. Lama periode perkawinan ini tergantung pada seberapa cepat induk tersebut bunting lagi setelah beranak, yang pada gilirannya tergantung pada timbulnya siklus birahi. Kondisi ini dipengaruhi oleh bangsa kambing dan oleh beberapa faktor lingkungan. Kondisi hewan yang buruk, makanan yang tidak memadai, atau sebab lain yang dapat menunda timbulnya birahi setelah melahirkan. Devendra(1962) melaporkan bahwa periode perkawinan pada kambing Anglo-Nubia murni di Malaysia rata-rata 327 hari, sedang pada kambing lokal (kacang) rata-rata 92 hari.
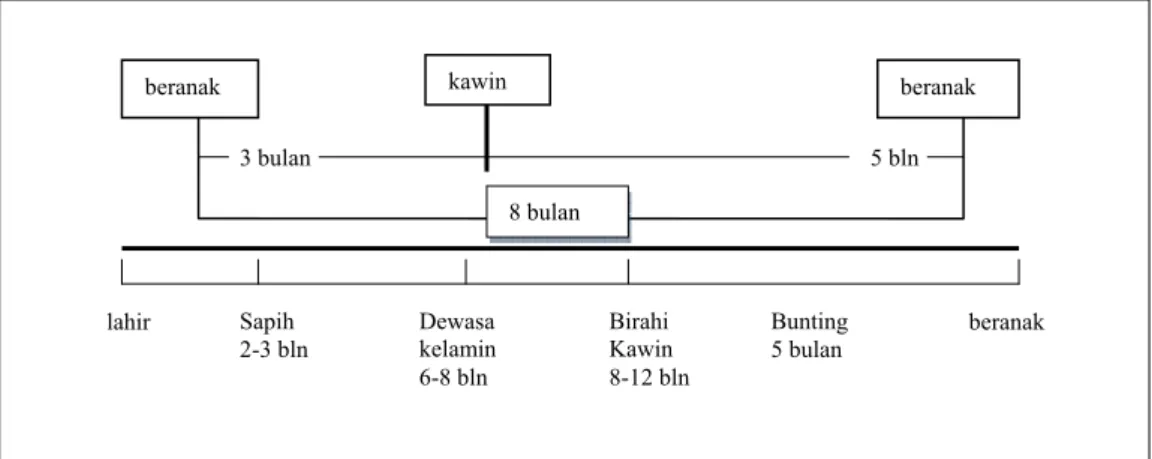
Mengawinkan kambing sesuai dengan waktunya, baik waktu kelahiran, penyapihan, serta mengawinkan kembali, perlu dilakukan pengaturan siklus reproduksi ternak seperti pada Gambar 4 berikut ini.
Gambar 4 Siklus reproduksi kambing betina agar dapat beranak 3 kali dalam 2 tahun atau beranak setiap 8 bulan (Mulyono 2004).
Produktivitas Ternak Kambing
Produktivitas semua bangsa hewan ternak dipengaruhi oleh faktor genetik dan lingkungan hidup ternak bersangkutan (Johansson et al. 1968 yang disetir Abdulgani 1981). Bersama-sama dengan kedua faktor di atas, peranan peternak dalam mengelola ternaknya menentukan pula tinggi rendahnya produksi yang akan dicapai.
Beberapa literatur umumnya dinyatakan bahwa pelaksanaan manajemen untuk meningkatkan keefisienan produksi ternak antara lain : (1) berusaha agar kondisi badan serta kesehatannya baik, (2) bebas dari gangguan penyakit, (3) pelaksanaan flushing (pemberian makanan yang cukup dan bermutu menjelang dan selama masa birahi), (4) berusaha agar dikawinkan pada umur muda dan tepat waktunya, (5) lekas dikawinkan kembali setelah beranak, dan (6) cukup tersedia pejantan unggul yang selalu dijaga kondisi makanannya (Ray dan Smith 1966; Guha et al. 1967; Turner dan Young 1969 dikutip Abdullgani (1981). Selanjutnya dinyatakan bahwa dalam bidang pemuliaan ternak, usaha untuk meningkatkan keefisienan produksinya dilakukan dengan cara menyeleksi ternak-ternak yang tingkat kesuburannya tinggi, menyisihkan ternak-ternak yang memiliki sifat produksi buruk, dan cara-cara perkawinan yang tepat.
Menurut Devendra dan Nozawa (1976), usaha peningkatan produktivitas ternak kambing di suatu wilayah perlu diperhatikan hal-hal sebagai berikut: (1) penggunaan bibit ternak yang baik mutunya; (2) peningkatan banyaknya cempe
beranak kawin beranak
8 bulan 3 bulan 5 bln lahir Sapih 2-3 bln Dewasa kelamin 6-8 bln Birahi Kawin 8-12 bln Bunting 5 bulan beranak
yang dilahirkan serta memperpanjang kehidupan induk yang produktif; (3) usaha pengendalian terhadap banyaknya ternak yang dipelihara; (4) meningkatkan penggunaan bibit ternak yang telah terbukti keunggulannya; (5) manajemen yang lebih efisien, terutama dalam hal pemberian bahan makanannya; (6) usaha pengendalian penyakit; (7) penelitian terhadap ternak bersangkutan serta penerapannya dalam praktek melalui pendidikan.
Untuk mendapatkan gambaran tentang tingkat produktivitas ternak kambing adalah dengan menghitung banyaknya cempe yang lahir dalam kelompok kambing tersebut selama waktu tertentu, selang beranak atau interval kelahiran, umur mulai dikawinkan, masa bunting, umur beranak pertama, bobot lahir cempe, bobot kambing pada umur tertentu, bobot badan kambing dewasa, tingkat kematian dan heritabilitasnya.
Jumlah Anak Kambing yang Lahir per Kelahiran (Litter Size)
Faktor-faktor yang mempengaruhi jumlah anak lahir perkelahiran (litter size) pada ternak kambing diantaranya umur induk, tingkat nutrisi maupun pengaruh lingkungan lainnya. Astuti (1984) melaporkan bahwa rataan jumlah anak lahir per kelahiran kambing Peranakan Etawah di Yogyakarta sebesar 1.7 relatif tinggi bila dibanding pengamatan Setiadi et al. 1984 di Bogor yakni sebesar 1.3 ekor.
Wilson (1983) yang mengamati kambing lokal di Sudan dan Mali mendapatkan jumlah anak lahir per kelahiran sebesar 1.52 di Sudan dan 1.21 di Mali. Astuti (1984) melaporkan bahwa pada Kambing Kacang di Imogiri Jogyakarta, rata-rata banyaknnya cempe per kelahiran adalah 1.73 ekor. Data kelahiran kambing di fakultas peternakan IPB selama periode 1954-1973 rata-rata menunjukkan banyaknya cempe per kelahiran adalah 1.49 ± 0.07 ekor.
Jumlah anak lahir per kelahiran pada ternak kambing bervariasi baik dalam satu bangsa maupun antar bangsa kambing. Dari 3 914 ekor anak kambing Beetal di India, 24 persen lahir tunggal, 63 persen kembar dua, 11.5 persen kembar tiga dan 1.5 persen kembar empat dan dilain pihak pada kambing Beetal
di Zambia, lahir kembar dua jarang terjadi dan hanya kelahiran tunggal yang sering terjadi (Mc Dowell dan Bove 1977).
Umur Beranak Pertama
Umur beranak pertama sangat erat hubungannya dengan umur mulai dikawinkan yang bergantung pada kondisi badan atau makanan ternak bersangkutan. Kambing dara yang masak dini akan lebih cepat beranak pertama daripada yang masak lambat. Ini berarti bahwa bangsa kambing yang tergolong masak dini akan lebih cepat mampu menyumbangkan hasil produksinya bagi masyarakat.
Budihardi (1972) melaporkan bahwa umur beranak pertama pada kambing kacang di Imogiri Jogyakarta, adalah 1.5 tahun. Di fakultas Peternakan IPB rata-rata umur beranak pertama adalah 979.50±115.85 hari (29 bulan). Pengaruh keadaan lingkungan tempat kambing hidup dan berkembang diperlihatkan oleh adanya perbedaan dalam mencapai umur beranak pertama. Galeon (1951) yang dikutip Abdulgani (1981) melaporkan bahwa kambing di Pilipina rata-rata umur beranak pertamanya adalah 459.75 hari. Kambing Saanen yang diternakan di Israel beranak pertama pada umur 2 tahun (masak lambat).
Bobot Lahir Cempe
Bobot lahir adalah salah satu faktor penting di dalam dunia peternakan, karena merupakan titik awal pengukuran perkembangan selanjutnya. Epstein dan Herz (1964) dan Scott (1970) disetir Setiadi (1987) menyatakan bahwa bobot lahir dapat digunakan sebagai petunjuk terhadap perkembangan setelah lahir.
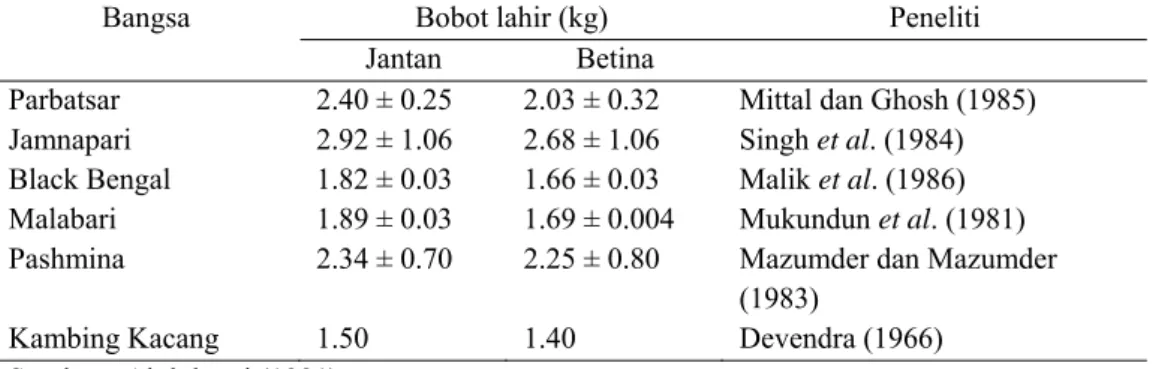
Banyaknya anak per kelahiran dan jenis kelamin berpengaruh terhadap bobot lahir. Setiadi (1987) melaporkan bahwa bobot lahir kambing Peranakan Etawah (PE) di pedesaan sebesar 2.5±0.6 kg. Bobot lahir anak jantan lebih berat dari betina, yaitu 2.6 kg vs 2.2 kg. Selanjutnya Singh et al.(2002) melaporkan bahwa bobot lahir kambing Jamnapari jantan sebesar 2.92±1.06 kg dan betina sebesar 2.68±1.06 kg. Jenis kelamin berpengaruh terhadap bobot lahir, dimana
bobot lahir anak jantan lebih tinggi daripada bobot lahir anak betina seperti disajikan dalam Tabel 9.
Tabel 9 Pengaruh jenis kelamin terhadap bobot lahir beberapa bangsa kambing Bangsa Bobot lahir (kg) Peneliti
Jantan Betina Parbatsar Jamnapari Black Bengal Malabari Pashmina Kambing Kacang 2.40 ± 0.25 2.92 ± 1.06 1.82 ± 0.03 1.89 ± 0.03 2.34 ± 0.70 1.50 2.03 ± 0.32 2.68 ± 1.06 1.66 ± 0.03 1.69 ± 0.004 2.25 ± 0.80 1.40
Mittal dan Ghosh (1985) Singh et al. (1984) Malik et al. (1986) Mukundun et al. (1981) Mazumder dan Mazumder (1983)
Devendra (1966) Sumber : Abdulgani (1981)
Mittal (1979) menyatakan bahwa faktor musim berpengaruh terhadap bobot lahir anak kambing (Tabel 10). Penelitiannya meliputi dua bangsa kambing dengan dua faktor musim yaitu musim panas (summer) dan musim dingin dan hujan (monsoon). Anak-anak kambing yang tetuanya kawin pada musim dingin dan hujan mempunyai bobot lahir lebih tinggi dari anak-anak kambing yang tetuanya kawin pada musim panas. Hal ini berlaku pada kedua bangsa, baik yang terlahir tunggal maupun yang terlahir kembar.
Tabel 10. Pengaruh musim kawin terhadap bobot lahir anak kambing Bangsa Musim Panas (Summer) Musim Dingin (Winter)
Jantan Betina Jantan Betina
Barbari Tunggal Kembar Dua Jamnapari Tunggal Kembar dua 1.98 ± 0.30 1.84 ± 0.16 2.69 ± 0.04 2.32 ± 0.02 1.80 ± 0.18 1.72 ± 0.17 2.38 ± 0.09 2.12 ± 0.08 2.41 ± 0.09 2.38 ± 0.04 3.46 ± 0.13 3.20 ± 0.02 2.18 ± 0.04 1.98 ± 0.11 2.94 ± 0.14 2.85 ± 0.12 Sumber : Mittal (1979)
Mittal (1979) juga menyatakan bahwa umur induk berpengaruh terhadap bobot lahir anak kambing. Ada kecendrungan bahwa makin tua umur induk maka makin besar pula bobot lahir anaknya, baik pada tipe kelahiran tunggal maupun pada tipe kelahiran kembar dua Tabel 11.
Tabel 11 Pengaruh umur induk terhadap bobot lahir kambing Bangsa Tipe Kelahiran Jenis
Kelamin
Umur Induk (tahun)
2 2-4 4-6 Barbari Jamnapari Tunggal Kembar dua Tunggal Kembar dua Jantan Betina Jantan betina Jantan Betina Jantan Betina 2.22 ± 0.30 1.89 ± 0.13 1.75 ± 0.23 1.53 ± 0.12 3.02 ± 0.13 2.79 ± 0.07 2.98 ± 0.13 2.51 ± 0.06 2.20 ± 0.12 2.05 ± 0.13 2.11 ± 0.23 1.77 ± 0.24 3.28 ± 0.07 2.88 ± 0.03 3.12 ± 0.20 2.66 ± 0.08 2.38 ± 0.17 2.28 ± 0.11 2.01 ± 0.32 1.71 ± 0.10 3.44 ± 0.12 2.80 ± 0.06 3.14 ± 0.08 2.58 ± 0.15 Sumber : Mittal (1979)
Potensi Ekonomi Ternak Kambing di Indonesia
Peternakan kambing di Indonesia yang masih berskala kecil perlu diusahakan secara komersial. Hal ini perlu dikembangkan karena adanya pertumbuhan penduduk sekitar 2% dan kenaikan tingkat daya beli masyarakat. Kebutuhan daging saat ini belum mencukupi permintaan. Dengan jumlah penduduk tahun 2005 mencapai sekitar 220 juta jiwa dengan rata-rata konsumsi daging sekitar 1.75 kg/kapita/tahun, maka total kebutuhan daging domestik mencapai sekitar 384.81 ribu ton. Sementara itu total produksi daging dalam negeri hanya mencapai 271.84 ribu ton sehingga masih ada kekurangan sekitar 112.97 ribu ton atau 29.36% dari total kebutuhan dalam negeri (Ditjend Peternakan 2005)
Selain pertimbangan di atas, ternak kambing juga mempunyai potensi ekonomi yang baik. Potensi ekonomi ternak kambing ini antara lain menurut Mulyono (2004):
1) Ternak Kambing mempunyai badan yang relatif kecil dan pertumbuhan yang cepat sehingga tingkat reproduksi dan produksi lebih tinggi,
2) ternak kambing tidak memerlukan lahan yang luas, apalagi dapat dilakukan kemitraan dengan pihak pengadaan pakan hijauan,
3) suka bergerombol sehingga dalam hal tenaga kerja, sistem pengembalaan lebih efisien,
4) skala usaha pembibitan ternak kambing yang dianjurkan adalah 8-12 ekor induk dengan harapan setiap kali melahirkan akan diperoleh anak sapih sekitar 12 – 18 ekor.
5) memiliki sifat toleransi yang tinggi terhadap bermacam-macam hijauan pakan ternak,
6) memiliki daya adaptasi yang baik terhadap berbagai keadaan lingkungan sehingga dapat diternakkan dimana saja dan dapat berkembangbiak sepanjang tahun.
Jumlah ternak kambing di Indonesia dibandingkan dengan ternak ruminansia lainnya pada tahun 2005 relatif lebih banyak Tabel 12 dan untuk populasi ternak kambing per provinsi dapat dilihat pada Tabel 13.
Tabel 12 Populasi Ternak Tahun 2001-2005 (000 ekor) No Jenis Ternak Tahun 2001 2002 2003 2004 2005 (1) (2) (3) (4) (5) (6) (7) 1. 2. 3. 4. 5. Sapi potong Sapi perah Kerbau Kambing Domba 11 137 347 2 333 12 464 7 401 11 298 358 2 403 12 549 7 641 10 504 374 2 459 12 722 7 811 10 533 364 2 403 12 781 8 075 10 680 374 2 428 13 182 8 307 Sumber : Dirjen Peternakan (2005)
Tabel 13 Populasi kambing tahun 2003-2007 per provinsi di Indonesia Provinsi Tahun 2003 2004 2005 2006 2007 NAD 637 188 647 089 655 242 787 708 894 199 Sumut 712 566 717 196 721 858 643 86 712 753 Sumbar 256 230 195 176 148 467 223 334 259 058 Riau 231 757 203 999 215 793 238 043 250 527 Jambi 126 863 132 369 138 114 137 989 155 169 Sumsel 436 607 435 504 453 926 463 72 582 534 Bengkulu 166 589 108 619 112 964 102 855 106 969 Lampung 810 456 824 235 868 133 798 816 846 122 DKI Jakarta 5 351 6 971 7 313 9 333 10 373 Jabar 930 066 1 144 102 1 235 973 1 148 547 1 393 190 Jateng 2 984 845 2 993 138 3 007 593 3 165 040 3 193 842 DIY 241 007 256 417 258 981 280 182 282 984 Jatim 2 334 554 2 359 375 2 382 969 2 414 350 2 457 059 Bali 61 958 44 418 44 418 70 785 70 833 NTB 282 500 300 280 315 294 376 13 394 937 NTT 435 151 462 101 490 721 496 766 509 23 Kalbar 96 360 99 010 104 960 107 762 118 54 Kalteng 23 160 37 398 37 398 41 046 43 098 Kalsel 84 442 91 911 96 507 107 873 111 733 Kaltim 74 335 72 071 73 512 53 105 54 167 Sulut 45 910 44 234 44 375 42 759 43 399 Sulteng 161 920 163 090 163 948 188 362 171 723 sulsel 555 927 403 505 511 895 433 495 543 672 Sultra 73 927 82 160 90 080 97 976 102 645 Maluku 156 406 168 719 171 755 149 405 152 394 Papua 41 969 55 069 57 066 37 226 41 822 Babel 11 377 2 450 2 499 3 561 3 846 Banten 522 380 566 716 564 463 681 253 817 732 Gorontalo 137 879 92 944 131 618 96 568 111 098 Malut 82 402 70 695 74 229 139 981 149 776 Kepri - - 21 558 20 238 20 643 Irjabar - - 12 923 11 708 13 163 Sulbar - - 209 694 220 179 254 286 Total 12 722 082 12 780 961 13 182 064 13 789 954 14 873 516 Sumber: Dirjen Peternakan (2008)
Dinamika Populasi
Dinamika populasi menjelaskan suatu keragaman di dalam ukuran populasi serta komposisi umur-kelamin, laju reproduksi dan mortalitas di dalam suatu populasi satwa. Dinamika populasi mempelajari bagaimana keragaman tersebut muncul akibat adanya interaksi antara individu maupun individu dengan lingkungannya (Bailey 1984).
Berbagai populasi satwa liar sebagai contoh dapat berubah ukurannya dalam jangka waktu tertentu (satu musim, satu tahun, ataupun beberapa tahun). Perubahan-perubahan anggota populasi ini sangat penting diketahui agar dapat mengatur populasi untuk memperoleh suatu jumlah yang optimum sesuai dengan daya dukung habitatnya. Perubahan ukuran populasi yang tidak beraturan menurut skala waktunya (irregular) disebut fluktuasi, sedangkan perubahan yang beraturan dan tetap skala waktunya (regular) disebut siklis. Fluktuasi dapat disebabkan karena adanya perubahan cuaca, perubahan ketersediaan makanan, ataupun karena adanya pengaruh perburuan untuk konsumsi manusia. Menurut Sihombing (1977) bahwa semua makhluk hidup dalam populasi akan berubah karena pengaruh perubahan lingkungannya.
Menurut Krebs (1978), populasi dapat dikelompokkan ke dalam deme-deme atau populasi lokal, yang dapat melakukan perkawinan antara organisme. Setiap populasi memiliki karakteristik kelompok yang beragam. Secara umum, karakteristik populasi yang paling mendasar adalah ukuran atau kepadatan. Empat parameter yang mempengaruhi kepadatan adalah natalitas, mortalitas, emigrasi dan imigrasi. Karakteristik sekunder dari populasi adalah sebaran umur, komposisi genetik, dan pola sebaran (penyebaran secara individu di dalam suatu ruang). Karakteristik yang terakhir adalah karakteristik yang dimiliki secara individual.
Parameter populasi yang utama adalah struktur populasi, yang terdiri dari seks rasio, distribusi kelas umur, tingkat kepadatan dan kondisi fisik (Van Lavieren 1982). Kepadatan populasi adalah besaran populasi dalam suatu unit luas atau volume. Nilai kepadatan diperlukan untuk menunjukkan kondisi daya dukung habitatnya.
Natalitas merupakan jumlah individu baru (anak) yang lahir dalam suatu populasi, yang dapat dinyatakan dalam beberapa cara yaitu produksi individu baru dalam suatu populasi (Odum 1971). Natalitas juga dapat dinyatakan dalam laju kelahiran kasar (crude birth rate), yaitu perbandingan antara jumlah individu yang dilahirkan dengan jumlah seluruh anggota populasi pada suatu periode waktu.
Keanekaragaman Genetik
Keanekaragaman genetik seringkali dihubungkan dengan tingkah laku reproduktif dari individu dalam suatu populasi. Populasi adalah satu kelompok individu yang kawin satu sama lain dengan menghasilkan keturunan; satu spesies mungkin mencakup satu atau lebih populasi yang terpisah. Populasi mungkin terdiri dari satu individu atau jutaan individu (Primack et al. 1998).
Individu di dalam suatu populasi biasanya berbeda secara genetik satu sama lain. Variasi genetik muncul karena individu memiliki gen yang berbeda, unit kromosom yang memberi kode untuk protein tertentu. Perbedaan bentuk dari suatu gen disebut alel, dan perbedaan ini muncul karena mutasi yang terjadi di dalam deoxyribonucleic acid (DNA) yang membentuk komponen-komponen individu kromosom. Beragam alel dari satu gen mungkin akan mempengaruhi perkembangan dan fisiologis suatu organisme secara berbeda. Pengembangbiakan tanaman dan hewan mengambil keuntungan dari variasi genetik ini untuk menghasilkan strain yang lebih tahan penyakit terutama untuk spesies yang sudah dipelihara seperti gandum, jagung dan ternak.
Variasi genetik meningkat sewaktu keturunan menerima kombinasi unik gen dan kromosom dari induknya melalui rekombinasi gen yang muncul selama reproduksi seksual. Gen dipertukarkan antara kromosom selama miosis, dan kombinasi baru diciptakan sewaktu kromosom dari kedua induk dikombinasikan untuk membentuk keturunan yang unik secara genetik.Walaupun mutasi memberi materi dasar bagi variasi genetik, namun kemampuan untuk menghasilkan spesies secara seksual menjadi alel yang disusun secara acak di dalam kombinasi yang berbeda-beda meningkatkan potensi untuk variasi genetik (Primack et al. 1998). Lebih jauh dikatakannya, bahwa jumlah variabilitas genetik di dalam suatu
populasi ditentukan oleh jumlah gen pool (larikan total gen dan alel di dalam suatu populasi) yang memiliki lebih dari satu alel (disebut juga gen polymorphik) dan oleh jumlah alel dari setiap gen yang bersifat polimorfik. Keberadaan gen polimorfik memungkinkan individu di dalam populasi menjadi heterozygous bagi gen tersebut untuk menerima alel gen yang berbeda dari setiap induknya.
Pelestarian Sumberdaya Genetik Ternak
FAO memprediksi bahwa paling sedikit satu bangsa ternak tradisional punah setiap minggu dan lebih dari 30% ternak Eropa sekarang ini diperkirakan dalam keadaan terancam kepunahan (FAO 1995). Banyak bangsa ternak tradisional sudah menghilang karena para petani lebih fokus pada bangsa/breed baru. Sekitar 16% dari bangsa sapi tradisional telah punah dan kurang dari 15% bersifat jarang (FAO 2000) Semakin berkembangnya ilmu pengetahuan dan teknologi pemuliaan ternak, bioteknologi, permintaan pasar, mekanisme pertanian dan produksi ternak, akan mendorong eksploitasi ternak melalui persilangan, penggantian breed baru (Subandriyo dan Setiadi 2003; Sodhi et al. 2006), maupun pengurasan stock secara berlebihan, dan pada gilirannya akan mengancam keragaman genetik ternak. Di lain pihak pelestarian keragaman genetik ternak akan selalu diperlukan dalam pemuliaan di masa datang, karena tanpa adanya keragaman genetik, pemuliaan ternak tidak mungkin dilaksanakan untuk mengantisipasi keperluan di masa mendatang (Subandriyo dan Setiadi 2003)
Pelestarian terhadap sumberdaya genetik ternak lokal sebagai bagian dari komponen keanekaragaman hayati adalah penting untuk memenuhi kebutuhan pangan, pertanian dan perkembangan sosial masyarakat di masa yang akan datang. Ada beberapa alasan untuk ini, antara lain (1) lebih dari 60 persen dari bangsa-bangsa ternak di dunia berada di negara-negara sedang berkembang; (2) konservasi bangsa ternak lokal tidak menarik bagi petani; (3) secara umum tidak ada program monitoring yang sistematis dan tidak tersedianya informasi deskriptif dasar sebagian besar sumber daya genetik hewan ternak; serta (4) sedikit sekali bangsa-bangsa ternak asli yang telah digunakan dan dikembangkan secara aktif (FAO 2001).
Ada tiga metode utama program pelestarian plasma nutfah ternak yang telah dilaksanakan masyarakat atau pemulia: (1) mempertahankan populasi ternak hidup, (2) penyimpanan beku materi genetik berupa haploid (n) seperti gamet yakni semen dan oocyte atau berupa diploid (2n) seperti embrio, dan (3) penyimpanan DNA (deoxyrybonucleic acid). Metode bioteknologi dapat digunakan untuk mengkarakterisasi gen-gen ternak dan plasma nutfah suatu populasi. Metode ini akan membantu dalam pembuatan keputusan tentang pelestarian plasma nutfah yang unik. Studi mengenai struktur dan fungsi gen-gen pada tingkat molekuler suatu populasi ternak dapat membantu menentukan kesamaan material genetik yang dibawa oleh dua atau lebih populasi dan keragaman genetik dalam populasi ternak yang diamati. Identifikasi gen-gen dari individu ternak akan membantu program pemuliaan (genetik) ternak, yang membedakan dari penampilan (fenotipe) yang tampak, yang dapat menentukan proses pemilihan tetua untuk generasi yang akan datang (seleksi buatan). Jika gen-gen untuk sifat produksi dapat diidentifikasi, ternak-ternak tersebut dapat diseleksi walaupun tidak diekspresikan oleh individu ternak yang bersangkutan. Sebagai alternatif, jika mereka dapat diikatkan dengan gen-gen yang diketahui lokasinya dalam kromosom (marker lokus-lokus), seleksi dapat dilaksanakan berdasarkan acuan tersebut (Subandriyo dan Setiadi 2003).
Filogeografi Intraspesies
Sebagai prinsip atau proses yang mengarahkan distribusi geografi spesies tertentu, filogeografi menekankan pada kajian variasi geografi yang mencakup berbagai tipe karakter seperti morfologi, fisiologi, etologi dan genetik (Lougheed and Handford 1993). Kajian tersebut memegang peranan penting dalam memahami proses-proses evolusi, karena variasi-variasi yang bersifat menurun adalah materi dasar yang terlibat dalam proses seleksi alam dan adaptasi. Penyebaran populasi suatu spesies dimulai dari variasi-variasi geografi yang semakin lama semakin berkembang. Filogeografi pada tingkat intraspesies dapat mengungkap keragaman di dalam spesies, mulai dari tingkat populasi, subpopulasi, subspesies sampai spesies.
Selain dapat menjelaskan pola-pola biogeografi (Mustrangi dan Patton 1997), filogeografi juga memberikan kontribusi pada pemahaman respon populasi terhadap perubahan lingkungan yang sangat cepat (Avise 1997). Pertama, populasi konspesies yang berbeda (khususnya yang terpisah dalam kurun waktu panjang) kemungkinan memberikan respon yang tidak sama terhadap perubahan lingkungan. Adaptasi terhadap lingkungan menyebabkan gen-gen yang terakumulasi berbeda antara populasi satu dengan populasi lainnya. Kedua, perkembangan pesat dalam metode analisis genetik memungkinkan dilakukannya identifikasi respon populasi terhadap modifikasi lingkungan pada tingkat intraspesies. Ketiga, analisis filogeografi dapat membuktikan hubungan dekat antara demografi populasi dan genetik. Hal ini akan berimplikasi pada konservasi khususnya dalam periode perubahan lingkungan yang sangat cepat. Keempat, analisis filogeografi dapat menunjukkan catatan-catatan perubahan genetik sebagai akibat dari perubahan kejadian demografi yang aneh.
Gambar 5 terlihat hubungan asal-usul genetik antara dua populasi konspesies yang semula berasal dari satu daerah geografi tetapi kemudian terpisah dalam dua daerah geografi ( daerah I dan II). Adanya Barrier menyebabkan tidak adanya aliran gen antara populasi B dan C. Filogeni antara keturunan populasi B dan C semakin berbeda akibat kurun waktu isolasi yang semakin lama. Perbedaan tersebut dapat ditunjukkan dengan jarak genetik (genetic disttance) yaitu ukuran perbedaan genetik dari dua takson ( Hillis et al. 1996).
Masa lalu, perspektif filogeografi intraspesies jarang diaplikasikan karena kekurangan pendekatan genetik molekuler untuk mendapatkan hubungan historis. Selain itu, juga masih terdapat persepsi luas bahwa pada populasi-populasi yang masing-masing dapat melakukan silang antar (interbreding), filogeni tidak memiliki pengertian yang nyata (Avise 1979). Hal tersebut disebabkan analisis filogeografi dilakukan berdasarkan karakter morfolgi saja sehingga hanya sedikit dapat menunjukkan keragaman intraspesies.
Gambar 5 Hipotesis Avise tentang asal-usul DNA (Avise 1997 dalam Surata 2000). (A) populasi nenek moyang ; (B) populasi yang merupakan keturunan dari populasi A pada daerah I; (C) populasi yang merupakan keturunan dari populasi A pada daerah II ; ( → ) arah divergensi dari populasi induk kepada populasi-populasi keturunannya.
Seleksi dan Breeding
Seleksi dari segi genetik diartikan sebagai suatu tindakan untuk membiarkan ternak-ternak tertentu berproduksi, sedangkan ternak lainnya tidak diberi kesempatan berproduksi (Noor 2004). Lebih jauh dikatakannya bahwa terdapat dua kekuatan yang menentukan apakah ternak-ternak pada generasi tertentu bisa menjadi tetua pada generasi selanjutnya. Kedua kekuatan itu adalah seleksi alam dan seleksi buatan.
Seleksi dalam pemuliaan ternak adalah memilih ternak yang baik untuk digunakan sebagai bibit yang menghasilkan generasi yang akan datang. Untuk bidang peternakan, yang diseleksi adalah sifat-sifat terukur seperti kecepatan pertumbuhan, bobot lahir, produksi susu dan bobot sapih (Falconer 1972). Sifat-sifat ini memberikan manfaat secara ekonomi disamping harus mempunyai kemampuan mewarisi yang tinggi yang dapat ditentukan dari nilai heritabilitasnya. B1 A B14 B11 B12 B13 B2 B C C14 C13 C12 C11 C2 C1 Barrier Waktu
Falconer (1972) yang dikutip oleh Hardjosubroto (1994) mengemukakan bahwa ada tiga metode seleksi yang sederhana, yaitu:
1. Seleksi individu (individual selection) adalah seleksi per ternak sesuai dengan nilai fenotipe yang dimilikinya. Metode ini adalah yang paling sederhana daripada umumnya dan menghasilkan respon seleksi yang cepat.
2. Seleksi keluarga ( family selection) adalah seleksi keluarga per keluarga sebagai kesatuan unit sesuai dengan fenotip yang dimiliki oleh keluarga yang bersangkutan. Individu tidak berperan dalam metode seleksi ini. 3. Seleksi dalam keluarga (within-family selection) adalah seleksi tiap
individu di dalam keluarga berdasarkan nilai rata-rata fenotip dari keluarga asal individu bersangkutan.
Dasar pemilihan dan penyingkiran yang digunakan dalam seleksi adalah mutu genetik seekor ternak. Mutu genetik ternak tidak tampak dari luar, yang tampak dan dapat diukur dari luar adalah performansnya. Performans ini sangat ditentukan oleh dua faktor, yaitu faktor genetik dan lingkungan. Oleh karena itu, harus dilakukan suatu pendugaan atau penaksiran terlebih dahulu terhadap mutu genetiknya atas dasar performnsnya (Hardjosubroto 1994).
Menurut Widodo dan Hakim (1981) dalam Rahmat (2006), pada saat melakukan seleksi, ternak yang mempunyai performans di atas dari performans yang telah ditentukan terlebih dahulu akan dipilih, sedangkan yang lebih rendah dari performans tadi akan disingkirkan. Ternak yang terpilih akan memiliki nilai rerata performans yang lebih tinggi dari performans keseluruhan sebelum
seleksi. Menurut Hadjosubroto (1994), perbedaan antara rerata performans dari ternak yang terseleksi dengan rerata performans populasi sebelum seleksi disebut sebagai diferensial seleksi (selection differential) yang dirumuskan sebagai:
S = ( s - )
Keterangan : S = Diferensial seleksi (selection differential) = rerata fenotip populasi
s= rerata fenotip sesudah adanya seleksi.
Selanjutnya dikatakan bahwa proporsi dari diferensial seleksi yang dapat diwariskan kepada generasi berikutnya adalah hanya yang bersifat genetik saja,
yaitu sebesar angka pewarisannya (heritability). Jadi, besarnya differensial seleksi yang diwariskan adalah sebesar h2
S dan ini disebut sebagai tanggapan (respon) seleksi yang akan muncul pada generasi berikutnya.
Pola Pemuliaan (Breeding Scheme)
Pemuliaan ternak adalah usaha jangka panjang dengan suatu tantangan utamanya adalah memperkirakan ternak macam apa yang menjadi permintaan di masa mendatang serta merencanakan untuk menghasilkan ternak-ternak yang diharapkan tersebut (Warwick et al. 1990). Peran pemuliaan dalam kegiatan produksi ternak sangat penting diantaranya untuk menghasilkan ternak-ternak yang efisien dan adaptif terhadap lingkungan. Produksi ternak yang efisien bergantung pada keberhasilan memadu sisitem manajemen, makanan, kontrol penyakit, dan perbaikan genetik.
Salah satu cara untuk perbaikan genetik pada ternak dilakukan melalui seleksi dalam kelompok ternak lokal dengan tujuan untuk meningkatkan frekuensi gen yang diinginkan. Kegiatan seleksi akan efektif bila jumlah ternak yang diseleksi banyak, walaupun catatan performan individu dari jumlah yang banyak akan sangat mahal. Salah satu cara untuk mengatasi hal ini adalah, seleksi atau peningkatan mutu genetik dilakukan pada kelompok-kelompok tertentu kemudian disebarkan pada kelompok lain (Wiener 1999). Struktur ternak bibit umumnya berbentuk piramida yang terbagi menjadi tiga strata (tiers) yaitu pada puncak piramida kelompok elite (nucleus), kelompok pembiak (multiplier), dan paling bawah kelompok niaga (Nicholas 1993; Warwick et al. 1990; Wiener 1999).
Pola pemuliaan pada dasarnya ada dua bentuk yaitu pola inti tertutup (Closed nucleus breeding scheme) dan pola inti terbuka (Open nucleus breeding scheme). Pada pola tertutup aliran gen hanya berlangsung satu arah dari puncak (nucleus) ke bawah tidak ada gen yang mengalir dari bawah ke nucleus. Perbaikan genetik pada commercial stock terjadi bila ada perbaikan pada nucleus. Peningkatan mutu genetik pada nucleus tidak segera tampak pada strata di bawahnya. Perlu waktu untuk meneruskan kemajuan genetik pada suatu strata ke strata berikutnya. Perbedaan performans antara dua strata yang berdekatan
biasanya diekspresikan dengan jumlah tahun terjadinya perubahan genetik yang ditunjukkan oleh perbedaan performan antara strata yang berdekatan (Nicholas 1993).
Pola inti terbuka adalah suatu sistem dimana inti (nucleus) tidak tertutup. Oleh karena itu, aliran gen tidak hanya dari strata atas ke bawah saja tetapi juga dari bawah ke atas. Karena itu setiap perbaikan genetik yang diperoleh dari hasil seleksi di tingkat dasar akan memberikan kontribusi pada peningkatan genetik inti, yang besarnya kontribusi bergantung pada laju aliran gen dari dasar ke inti. Dengan masuknya ternak bibit dari kelompok lain ke inti, hubungan kekerabatan antara induk dengan jantan makin jauh sehingga laju inbreeding berkurang. James (1979) mengemukakan bahwa kemajuan genetik pada sistem terbuka lebih tinggi dibandingkan dengan sistem tertutup. Pada sistem terbuka respon seleksi meningkat 10 sampai 15%, dengan laju inbreeding lebih rendah 50% bila dibandingkan dengan sistem tertutup pada kondisi dan ukuran sama.
Kosgey (2004) mengemukakan bahwa pola inti terbuka cocok digunakan untuk pemuliaan domba di negara berkembang (tropik). Selanjutnya dinyatakan bahwa pola pemuliaan yang digunakan di negara berkembang berbeda-beda sesuai dengan kondisi lingkungan dan sosial budaya setempat Pola-pola tersebut antara lain pola tiga strata terdiri atas inti (nucleus), kelompok pembiak (multiplier) dan populasi dasar; pola dua strata (inti dan peternak); dan pola hanya inti saja, yang programnya hanya menyeleksi jantan saja serta program seleksi jantan dan betina.
Karakteristik Ternak
Setiap sifat yang diekspresikan seekor hewan disebut fenotipe. Seekor hewan atau ternak menunjukkan fenotipenya (P) sebagai hasil pengaruh seluruh gen atau genotipenya (G), lingkungan (E) dan interaksi antara genotype dan lingkungan (IGE) (Martojo 1992; Hardjosubroto 1994) Sifat kuantitatif dan kualitatif pada hewan atau ternak merupakan fenotipe.
Pada program pemuliaan, prediksi perbedaan genetic di antara hewan dapat berdasarkan observasi fenotipe yang bergantung pada faktor genetik dan
lingkungan (Muladno 2006). Fenotipe ternak dapat diketahui melalui ukuran-ukuran tubuh (Otsuka et al. 1980; Karthickeyan et al. 2006), warna dan pola warna tubuh, pertumbuhan tanduk (Wiley 1981; Warwick et al. 1990). Warna termasuk sifat kualitatif seekor ternak. Warna tubuh ternak dianggap sebagai character displacement untuk membewdakan satu bangsa dengan bangsa lainnya.
Sifat kuantitatif adalah kharakteristik dari makhluk hidup yang dapat diukur, dihitung atau diskors. Karakter ini ditentukan oleh banyak pasang gen (poligenik) dan sangat dipengaruhi oleh lingkungan (Wiley 1981). Sedangkan sifat kualitatif seperti warna, pola warna, sifat bertanduk atau tidak bertanduk dapat dibedakan tanpa harus mengukurnya. Sifat kualitatif biasanya hanya dikontrol oleh sepasang gen (Noor 2004).
Sifat-sifat kualitatif adalah karakteristik yang pada umumnya dijelaskan dengan kata-kata atau gambar (Warwic et al. 1990). Spesies-spesies sering ditandai oleh warna atau pola warna tertentu. Pola warna apabila ada kemungkinan lebih berguna daripada warna itu sendiri. Hal ini biasanya dianggap sebagai character displacement untuk menghindari kesalahan saat membedakan bangsa ternak (Wiley 1981).
Balai Penelitian Ternak Ciawi sudah memulai mengkarakterisasi kambing Kosta (tahun 1995) dan Gembrong (tahun 1997) serta dilanjutkan oleh Loka Penelitian Kambing Potong Sei Putih pada tahun 2000-2006 untuk penelitian/karakterisasi kambing Marica (Sulawesi Selatan), kambing Muara (Kabupaten Tapanuli Utara-Propinsi Sumatera Utara) dan kambing Samosir (Kabupaten Samosir- Propinsi Sumatera Utara). Sampai saat ini sudah 7 bangsa kambing yang sudah dikarakterisasi karakteristik penotipenya, dan akan dilanjutkan untuk melaksanakan penelitian di beberapa daerah lain lagi (seperti kambing Benggala- Propinsi Nusa Tenggara Timur, Kambing Wetar- Propinsi Maluku). Diperkirakan masih banyak lagi bangsa kambing lokal Indonesia yang belum dapat dikarakterisasi dan sebagian mungkin sudah hampir punah atau jumlah populasinya sudah mendekati punah padahal kita belum sempat mengekplorasi potensi keragaman genetiknya untuk dimanfaatkan sebagai sumber peningkatan mutu genetik kambing di Indonesia
Struktur Kimia DNA
Setiap nukleotida tersusun oleh tiga komponen, yaitu molekul gula pentosa (deoxyribose untuk DNA dan ribose untuk RNA), gugus phospat, dan basa nitrogen. Dua komponen pertama terdapat di semua nukleotida dengan susunan dan bentuk yang identik, sedangkan komponen ketiga (basa nitrogen) mempunyai susunan dan bentuk yang berbeda di dalam suatu nukleotida dengan nukleotida lainnya. Namun demikian, hanya terdapat macam basa nitrogen penyusun setiap nukleotida (Muladno 2002).
Berdasarkan bentuk molekulnya basa nitrogen dikelompokan menjadi dua, yaitu purin dan pyrimidin. Basa purin terdiri atas basa Adenin (A) dan Guanin (G), sedangkan basa pyrimidin terdiri atas basa Cytosin (C), Uracyl (U) dan Thymin (T). Perbedaan struktur molekul lima basa nitrogen tersebut disajikan pada Gambar 6.
Gambar 6 Struktur kimia nukleotida pembawa basa nitrogen adenine (A), guanine (G), citosine (C), thymine (T) dan uracil (U) (Alberts et al. 1994).
Untai ganda DNA tersusun oleh dua rantai polinukleotida yang berpilin. Kedua rantai mempunyai orientasi yang berlawanan (antiparalel), rantai yang satu mempunyai orientasi 5’ 3’, sedangkan rantai yang lain berorientasi 3’ 5’. Kedua rantai tersebut berikatan dengan adanya ikatan hidrogen antara basa
adenine (A) dengan thymine (T), dan antara guanine (G) dengan cytosine (C). Ikatan antara A dan T berupa dua ikatan hidrogen, sedangkan antara G dan C berupa tiga ikatan hidrogen sehingga ikatan G dan C lebih kuat (Gambar 7). Spesifitas pasangan basa semacam ini disebut sebagai komplementaritas. Proporsi basa A dan T serta G dan C selalu sama sehingga komposisi DNA dapat dinyatakan dengan kandungan G + C (G + C content) yang berkisar dari 26% sampai 74%. Hal ini dikenal sebagai hukum Chargaff.
Gambar 7 Ikatan hidrogen antarnukleotida. Ikatan antara adenine (A) dengan thymine (T) dilakukan melalui dua ikatan hidrogen, sedangkan pada ikatan antara guanine (G) dan cytosine (C) ada tiga ikatan hidrogen sehingga ikatan G-C lebih kuat diabandingkan dengan ikatan A-T.
Berkembangnya biologi molekuler dan teknik DNA rekombinan menjadikan DNA inti dan mitokondria semakin banyak dapat diungkapkan. Sejalan dengan itu, penggunaan kajian genetika molekul telah mulai banyak dilakukan dalam dunia Genetika Populasi. Genom mitokondria akhir-akhir ini sering dijadikan alat dalam genetika populasi seperti penelusuran perjalanan kolonisasi populasi, pemisahan biogeografi populasi, hubungan filogeni, dan penelusuran asal-usul hewan (Hayashi et al. 1995; Harihara et al. 1996; Ming 1999; Perwitasari-Parajallah 1999). Selain itu, beberapa penyakit degeneratif, penuaan, dan kanker sering diimplikasikan dari kerusakan (defek) mitokondria
(Wallace, 1999). Cukup strategisnya eksistensi genom mitokondria menyebabkan biologi mitokondria perlu dipahami secara lebih mendalam.
Penciri DNA
Sejak era Mendel sampai tahun 1980-an, para ahli genetika hanya mendapatkan penciri genetik lokus tunggal berupa tampilan fenotipe (Crawford et al. 2000 dalam Margawati 2005). Penciri tersebut diantaranya seperti warna mata Drosophila atau polimorfisme protein seperti dalam penggolongan darah. Lebih lanjut dijelaskan bahwa penciri tersebut pada beberapa peta keterpautan genetik secara rinci telah dikembangkan pada model jenis seperti mencit dan drosophila. Namun demikian terdapat beberapa keterbatasan untuk penyusunan peta keterpautan pada persilangan jenis hewan domestik. Kehadiran teknologi DNA rekombinan, terutama teknik Polymerase Chain Reaction (PCR) telah mengubah secara mendadak hambatan dalam penyediaan penciri DNA. Dengan demikian seperti sekarang ini dapat dilihat banyak proyek pemetaan keterpautan untuk jenis ternak apapun dapat direncanakan dan diimplementasikan. Selama lebih dari satu dasawarsa terakhir ini, terdapat sejumlah penciri DNA yang secara rinci telah dideskripsikan dalam hubungannya dengan pencarian Quantitative Trait Loci (QTL), peta keterpautan perbandingan (comparative linkage mapping) dan pengukuran keragaman genetik dan pengukuran keragaman genetik.
Secara garis besar penciri DNA ini dapat dibedakan menjadi dua, yaitu Multilocus marker dan single locus marker (Crawford et al. 2000 dalam Margawati 2005). Termasuk dalam multilocus marker yaitu minisatelit atau variable number tandem reapeat (VNTR), randem amplified polymorphic DNA fragment (RAPD) dan amplified fragment length polymorphisms (AFLP). Sedangkan yang termasuk single locus markers adalah restriction fragment lenghth polymorphisms (RFLPs), Mikrosatelit dan single nucleotide polymorphisms (SNPs).
DNA Mitokondria
Organisme eukariot termasuk ternak domestik, sumber DNA dapat diperoleh dari organel-organel sitoplasmik antara lain DNA mitokondria (mtDNA). DNA mitokondria memiliki karakteristik sebagai molekul DNA yang diturunkan secara utuh tanpa adanya rekombinasi, memiliki molekul dengan ukuran kecil/pendek yang susunannya berbeda dengan DNA inti, dan memiliki variasi basa nukleotida yang lebih tinggi dibandingkan DNA inti. Tingginya variasi basa nukleotida disebabkan DNA mitokondria memiliki laju perubahan 5 – 10 kali lebih tinggi dibandingkan DNA inti (Muladno 2006).
DNA mitokondria mempunyai beberapa kelebihan yang menjadikannya banyak digunakan untuk mengidentifikasi keragaman genetik dan dinamika populasi . Beberapa kelebihan tersebut adalah: (1) memiliki ukuran yang kompak dan relatif kecil (16 000 - 20 000 pasang basa), tidak sekompleks DNA inti sehingga dapat dipelajari sebagai satu kesatuan yang utuh; (2) berevolusi lebih cepat dibandingkan dengan DNA inti; (3) hanya sel telur yang menyumbangkan material mitokondria sehingga mitokondria DNA hanya diturunkan dari induk betina; dan (4) bagian-bagian dari genom mitokondria berevolusi dengan laju yang berbeda, sehingga dapat berguna untuk studi sistematika dan penelusuran kesamaan asal-usul (Park dan Moran 1995).
Dewasa ini sekuen lengkap basa-basa penyusun genom mitokondria beberapa jenis organisme, terutama hewan, telah banyak diketahui. Beberapa diantaranya, bahkan telah banyak dimanfaatkan dalam analisis keragaman genetik dan biologi populasi pada hewan. Selain dapat melahirkan suatu rekonstruksi filogenetik dari beberap spesies yang saling berdekatan, hasil analisis tersebut juga dapat digunakan untuk menelusuri proses pemecahan dari spesies yang satu terhadap spesies lainnya.
Umumnya DNA mitokondria hewan berbentuk sirkuler dan berutas ganda, yakni terdiri dari utas berat (Heavy strand) dan utas ringan (Light strand). Berdasarkan jenis gennya, genom mitokondria ini dapat dibagi menjadi dua bagian, yaitu: daerah penyandi (coding region) dan daerah bukan penyandi (noncoding region). Daerah penyandi terdiri dari 37 gen, yaitu: 13 gen penyandi