VOLUME
72.NUMBER
12A STUDY OF THE BODY TEMPERATURE OF BIRDS
BY
ALEXANDER WETMORE
(Publication 2658)
CITY
OF WASHINGTON
PUBLISHED
BY THE SMITHSONIAN
INSTITUTIONDECEMBER
30, 1921BALTIMORE,MD., U. S. A.
By ALEXANDER WETMORE,
BIOLOGICAL SURVEY, U. S. DEPARTMENT OF AGRICULTURE
CONTENTS
Introduction i
Method of securingavian temperatures 3
Diurnal rhythmin temperature • 5
Table i.
Time
ofmaximum
andminimum
temperatures in birds 5Variation in temperature in relation to sex g
External temperature in relation to bodily heat 12 Diverse miscellaneous factors intheir relation to body temperature 16
Temperature of young 18
Table 2. Temperatures of young or nestling birds 19
Method of temperature control in birds 21
Significance of temperature control 28
Discussion of differences in average temperatures 30
Explanation of tables 32
Table 3.
Summary
of temperature records 36Table 4. Average temperature of families of birds, summarized from
Table3 47
Table 5. Temperatures of species of birds not included in Table 3,
taken from available literature 48
Bibliography 51
INTRODUCTION
The
subject of thebody
temperatures of themany and
varied species thatcompose
the great vertebrate class of birds is one that in the past has been rather shghtly treatedby
those interested in avianphysiology. Statements to the effect that the bodily tempera- tures of birds are higher than in others in thegroup
of classescom-
posingthe Vertebrata, are current inmany
zoologiesand
text books, buton
the whole, literature givesfew
definite statements of facton
the subject,and
observationshave
been restricted to a comparatively smallnumber
ofspecies. In thecourse of otherwork many hundreds
of birds have been handled in the fleshby
the present writer,and
aftersome
thoughtmethods were
devised for the takingand
record- ing ofbody
temperatures in as accurate amanner
as possible. Thiswas
alittleknown
fieldof endeavorand
atfirstno
guidesastomethod
Smithsonian Miscellaneous Collections,Vol. 72, No. 12
2
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 'J2.were
available, so that mistakeswere made, and
early recordshad
to be abandoned. Continued experimentation led to definitemethods
thatpromisedaccuracy,and
itisupon
these that thefollowing accountis based.
The
results of studies covering the periodfrom
January, 1912, to October, 1919, are included herewith, giving a totalof 1,558 records covering observationson
327 species of birds distributedamong
50 families. If thenumber
of observationsseems
smallcompared
to the length of time involved itmust
be considered that the taking of temperatureswas
not always feasible as the difficulties attending the taskwere
manifoldand
varied.The
results of thiswork
areembodied
in a series of tableswhich form
the foundationupon which
this report is based. In addition to the 327 species of birdson which
observations regardingbody
temperaturehave
beenmade by
thewriter personally, a supplementary compilation has been given (in table 5)embodying
published recordson
this subject for other species, in order to render this account of thebody
temperatures of birds as complete as practicable. In this final table records are given forj 89 forms, so that in the following pagesmay
befound
definite statements regarding the temperatures of416
species of birds. It ishoped
that thework
of securing such recordsmay
be continued until amuch
greateramount
of informa- tion coveringmany more
species is available.It
was
the original intention to incorporate in this paper a table giving in detail the individual recordsupon which
thework
has been based.Such
a coursewas
desirable as itwould
have furnished amass
of data far greater thanany
hitherto available for thosewho may
be interestedin using suchinformationin linesof researchother thanthatinwhich
ithasbeen utilized here. In additionitwould have
given opportunity to checkup more
carefully the deductions that Ihave
made.
Cost of printing of tabular matter has proved so high at the time of publication that it has been necessary to omit such a detailedstatementand
to supplytherecords only inacondensed form.The
table itself is, however, deposited in the files of the Smithsonian Institutionand may
be consultedby
those interested in using it.From
his studies in the subject of avianbody
temperatures the writer has ventured tocome
to certain conclusionsand
deductions,some
inaccordanceand some
at variancewithmodern
ideas as previ- ously accepted. If theseseem
sufficiently substantiated tomeet
the approval of others, thenmay
the timeand
labor required in the compilation of these data bedeemed
justified.The
following pages are respectfully submitted for attention, with thehope
that part ifnot all of this labor
may
gain thestamp
of approvalfrom
thosewho may
be interested in such lines of research.In carrying on these investigationsthe writer has
been
indebted to the Smithsonian Institution for a grant of seventy-five dollarsfrom
theHodgkins Fund,
for the purchase of specially constructed ther-mometers
required,and
for certain otherminor
expenses incurred.METHOD OF SECURING AVIAN BODY TEMPERATURES
In recording the
body
temperatures of birds itwas
necessary towork
withthermometers
that permitteda considerablerange in regis- tration as itwas found
that the degree of bodily heatwas
high orlow
according to the species in hand. All recordswere made
in the Fahrenheit scaleby means
of clinicalthermometers
similar inform
to those used
by
physicians.For
the present investigation itwas
necessary to have these instruments speciallymade
to give accurate registration rangingfrom 95°+
to 115°+
.Ordinary
clinical ther-mometers do
not register aboveiio°+
so that they could not be utilized, for certainpasseriform species frequently have temperatures ranging above that figure.While engaged
in fieldwork
these specialthermometers were
carried constantly in suitable carrying cases.When
a birdwas
shot a temperature readingwas
takenwhen
the specimen could be reached immediately. If therewas
delay in retrieving forany
reason,an
accurate temperature record could not bemade,
so thatitwas
possible to secure recordsin lessthanone-half of the birds thatwere
collected.In taking temperatures of specimens of small or
medium
size thethermometer was
placed in theopened mouth
of thebird,and worked down
untilthe lowerend was
entirely withinthe cavityof thethorax, usually until it reached the proventriculus.With
birds of larger size the reading w^as taken through the anus, with thethermometer
thrust inthrough the cloaca well into the canal of the large intestine.In either case it
was
necessary topenetrate thebody
cavity to secure a correct reading.The
peripheral circulation of birds is poor, so thatoften theremay
bea dififerenceoftwo
tofivedegreesbetween
the temperatureof theupper
part of the oesophagusand
that of thebody
cavity proper.A
similar variationwas
notedbetween
the tempera- ture of the cloacaand
the large intestine above it.The thermometers
usedwere
self-registering,and were
so con- structed thatthey reacted immediately toany
increase in heat.The
highest point of a reading
was
reached in a veryfew
seconds, but to insure accuracy the instrumentwas
held in position for a period4 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. ^2 rangingfrom
half aminute
to a minute. After each reading thethermometer was
shakendown
to be ready for instant usewhen needed
again.Temperatures were
read to tenths of a degree,and were
tran- scribed atoncein a notebook
opposite thename
of thebird, together with the dateand
otherneeded
information. In each case the time ofday was
noted to the nearest quarter hour.Thus
a temperature taken at five minutes past ten in themorning was
recorded as lo.oo a.m. and
one at ten minutes past ten as 10.15 a.m.
Further refine-ment
in recording the timeofday was
considered useless, as save on certain meridians there is always slight variationbetween
the actual time asshown by
the height of the sun,and
the time adopted for universal localuse as a matter of convenience.The
time of observa- tions recorded during the continuance of the so-called "daylightsaving"
regulations of 1918and
1919 has been corrected in each instance to normal.In order to be prepared at all times to secure temperature records of birds it
was
necessary to havethermometers
constantly athand
while in the field.The
physical labor involved in fieldwork
at timesis arduous, so that though carrying cases of an
improved
typewere
usedmany
instrumentswere
destroyed in spite of every precaution.Some were
broken through accident, otherswere
crushedby
birds while recordswere
being secured,and
afew were
lost. In all a considerablenumber were
used in securing the records given.The
records of temperatures of birds secured have been taken throughout the year, but the localitieswhere
thiswork
has been carriedonhave allbeen inthelimitsof theUnited
Statesin temperate regionswhere extreme
cold in winter has not been encountered.As
a matter of fact very
few
of the observations have beenmade
at a timewhen
the temperature of the airwas below
-f25° Fahr.For
this reason it has not been necessary to adopt
means
forwarming
the thermometers, or for keepingthem warm,
imm'ediately previous to their use.During
the early course of the observations recorded herein experimentswere made
in heatingthermometers by
holdingthem
in themouth
of theobserverwhen
itseemed
probably that theywere
to be used in a short time. In thisway
the temperature of the bulband
the glass for a short distance abovewas warmed
to -1-98°,more
or less (dependingupon
the condition of themouth).
Itwas
soonfound however
that the results gained withwarm and
coldthermometers were
so nearly identical that itwas
impossible to dis- tinguishbetween
them.Where
observationson
thebody
tempera- tures of birds are tobemade
at timeswhen
the airis at zero Fahren-NO. 12
BODY TEMPERATURE
OF BIRDSWETMORE
heit, oreven lower, it
would seem
probablethat itwould
be necessary to takesome
steps to usethermometers
thathad
been previously heated at least toaslightdegree. This factmay
bereadily determinedby
observation. It is probable that save in themost extreme
cold weather errorfrom
usingunwarmed
instrumentswould
notamount
to
more
than afew
tenths of a degree.The
observations recorded in the tables at the close of this paper are to be regarded at best as approximations to the actual tempera- ture of the birds handled. In treating data of this nature takenby
forcefrom
the lower animalswe
can onlyassume
thatwe
are correct in our observations.Through
experiencewe may
establishwhat we
consider as thenormal
limitof variation,butinmany
casesit isdifficulttobe assuredthatwe
are absolutely correct.Through
long experiencethewriter believes that the recordspresentedareaportrayal of theconditions as nearly correctasmay
be with theinstrumentsused.DIURNAL RHYTHM IN TEMPERATURE
The
question of the diurnal riseand
fall of temperature in afew
speciesof birdsand
animals hasbeen
carefully investigatedby Simp-
sonand
Galbraith.^These
authors, ontwo
dififerentoccasions,secured records of thebody
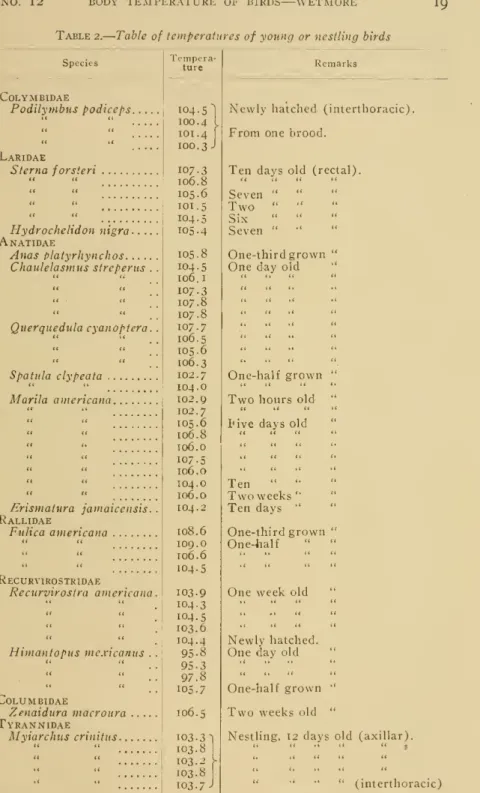
temperature of living gulls, starlings, sparrow-Table I. Time of
maximum
andminimum
temperatures in birds (Taken from Simpson andGalbraith)Species
Domesticfowld Domesticfowl 9
Bantam
fowld" .Bantam
fowl ?.Domestic duckd"
Domesticduck$
Domesticpigeond Domesticpigeon9
"Seagull"
"Seagull"
Jackdawd Jackdaw9
Timeof average
maximum
temp.
6 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. ']2of high
and low
temperature in individuals belonging- to thesame
species, as
shown
in the table on page 5, are probably tobe explainedon
thegrounds
of individualtemperament
in thebirdsunder
experi- ment.Thus
a bird that soonbecame
accustomed to handlingwould
give a slightly different series of readingsfrom
one thatremained
wildand
that struggled violentlywhenever
approached.)Inthecase ofthose species normallyactive duringthehoursofday- light a constant diurnal
rhythm was
indicated, with a gradual rise until late in the afternoonand
then a corresponding decrease until early in the morning. In owls, species of nocturnal habit, the tem- perature curveswere
reversed, the highest point being observed late at nightand
the lowest during the day. Itwas found
on thewhole
that the temperature curves of daylight-loving specieswere
similar to those of diurnalmammals
save that the point of highest tempera- ture in the birdscame
earlier in the afternoonand
that of low tem- perature earlier in the morning.Hilden
and
Stenback' recorda series ofexperiments inwhich
birdswere
confined in a darkroom and
their activities regulatedby means
of artificial light. Lightwas
turnedon from
6 p.m. to 6 a. m., with amore
brilliant illuminationfrom
9 p.m.
to 3 a.m.
to correspond to the brighter portion of the middle of thenormal
day. After the second day the diurnal birds studied in general adapted themselves to this change incondition in suchaway
thatthetemperaturerhythm was
reversed, the highest point in the record for each twenty-four hourscoming
aftermidnight instead of afternoon.When
the experi-ments were
terminatedand
the birdsagain led anormal
lifeinrelation to daylight the temperature curves at once adjusted to thenormal
rhythm.In studies
made by
the present writer therangeof diurnal tempera- ture is wellshown by
records for certain of the owls.Thus
amale
barn owl (Ahico pratincola) killed at 3.00 p.m., as it flewfrom
a perch in acottonwood
treehad
a temperature of 101.9°.The day was
brightand
clearand
the birdhad
in all probability been at rest since early morning.Another
male, shotat 8.30 p.m.,while quarter- ing backand
forth across a level flat near the GilaRiver in Arizona,showed
abody
heat of 105.0°. Thisbirdwas
seencoursing about for several minutesand had
evidently been hunting for food forsome
time.
The
variation in the screech owls(Otus
a^io) also is instruc- tive in connection with thesame
points.A
maleand
a female takenby hand
at 3.30 p.m.,from
alow
pine at the edge of aswamp
inSkandinavisches Arcli. fiir Phys. Bd. 34, 1916, pp. 382-413.
7
western Florida,gave temperatures of ioi.8°
and
102.7° respectively.A
secondmale
shot anhour
later registered 100.7°.These
birdshad
been resting quietly for the entire day. In direct contrast is the conditionfound
in amale
killedby
moonlight at 8.15p.m.
in the ChiricahuaMountains
in southeastern Arizona.On
this evening screechowlshad
beenactivelycalling forover anhour beforethis onewas
secured.The
bird takenhad
a temperatureof 105.4°.A
female secured in theDragoon
Mountains, Arizona, at 8.45 a.m., not long after ithad
retired for the day, registered abody
heat of 105.3°.In the case of those species active during the hours of daylight the differenceisless
marked, due
inlargeparttothefactthat themajority of birdsexamined had
been killed during the period of theirnormal
activity
when
little or no temperature rangewas
evident that could be correlated directly withtime.However,
it is possibleto citeafew
casesshowing
this regular variation.The
seriesofmourning
doves{Zcnaidura macroura)
obtainedmay
be discussed as pertinent in the matter of diurnal variation in birds of diurnal habit. In four males secured
between
5.00 a.m. and
5.45 a.m., the range in temperature isfrom
108.0° to 109.8°, the four records givingamean
of 108.9°. Five other birds shotbetween
5.00 p.m. and
6.00 p.m. under
similar conditions give a variation offrom
109.3° to 110.4° with amean
of 109.6°.An
average difference of .7° is thus shown. All of these birdswere
feeding but in the first set killed early inmorning
thebody
heatwas
still comparatively low.As
stated above,most
of the recordswere
secured at hourswhen
the temperature
was
naturally high so that little time variation is shovvoi.However,
the followingmay
be quoted in addition to the above.An
ash-throated flycatcher (Myiarchiis cinerascens) taken at 7.30 a.m. had
a temperatureof 108.6°.Another
taken at 10.00 a.m.
registered 110.0°
and
a third shot at 11.00 a.m. showed
abody
tem- perature of 111.8°. Allof these birdswere
males.A male Canadian
warbler (JVilsonia canadensis) shot at 7.30a.m.
registered 106.7°and
another takenat9.45a.m.
gaveatemperatureof 107.6°.Females
of thesame
species taken at 7.30and
9.00on
thesame morning had
a bodily heat of 107.7°^"^
107.0° while a third bird of the latter sex killed at 12.00 p.m. showed
108.3°.The
daily increase in bodily temperature in our smaller birdsmay
be exemplified also
by
the following:On September
12, 1919, atPlummers
Island,Maryland,
the writer spent the day in observing smallbirds,many
ofwhich were
in theirsouthward
migration.There
had been aheavy
rain the previous evening,and
laterhighwind had
come up
with a considerable fall in temperature.The
earlymorning
8 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 'J^was damp and
cold so that insectivorous birdswere more
or less inactive until the sunwas
wellup
in the sky.As
the airbecame warmer
the birds,correspondingly,became more
sprightly.A
female magnolia warbler{Dendroica
magnolia) secured at 8.30 a. m.,had
a bodily temperature of 107.0°.At
this time birds of similar habitwere
justcommencing
tomove
aboutand
feed.A male
of thesame
species shot at 9.30 a. m.had
a temperature of 108.2°and
a secondmale
taken at 10.15a.m. had
a temperature of 108.3°.Other war-
blers takenbetween
9.00 a.m. and
10.30 a.m.
gave records similar to thelast.Thus
a female bay-breasted warbler (Dendroicacastanca) shot at 9.15a.m.
registered 108.8°and
amale
killed at 10.00a.m.
registered 108.7°.
A male
blackand
whitewarbler {Mnioiilta varla) secured at 9.15 registered 108.5°. In the observations of this fore-noon
itwas
noted that the red-eyed vireo(Vireosyha
olivacea)was
astir
much
earlier intheday
than the warblers. This activity appar- entlywas
reflected in thebody
temperatureas a female taken at 9.15gave
a record of 109.7°, a reading distinctly higher than that of the warblers taken at thesame
time, although average temperatures for thetwo
groupson
thewhole
are about the same.Inperusal of the data used in preparing this paper
many
cases arefound
of range in temperature not correlated with time. These, however, cannot be taken as destroying the value of those instanceswhere
earlyand
late readingswere
available thatshow
a distinctrhythm
or increasefrom
early to late.As
stated above, the bulk of the records aremade
during the period of high activitywhen
the bodily temperature approaches amaximum. Range
in temperature then is to be attributed to other causes.Had
recordsmade
early in themorning
been available in all of these cases there can beno
ques- tionbutthatan
increase inbody
heat correlated with timewould
have been shown.The
daily variation in temperature ismuch more
in bi,rds of small size than in those of greater bulk.Thus Simpson and
Galbraith'
foundthe daily range inthe "thrush"
(Turdus
nierula?) tobefrom
6.8° to 7.5°, in starlings {Stiirnus vulgaris)from
5.2° to 7.2°, while inthedomestic fowl itamounted
to 1.9°and
in thedomesticatedduck from
1.6° to 1.8°.The marked
differencein these casesis noticeable.The
diurnal variation in temperature that has been notedmay
be attributed directly to the metabolism of the individual as reflectedby
its activities.With movement and
the digestion of food heat isgenerated.
Although
a part of this is dissipated through the usual*Journ. of Phys., Vol. XXXIII, 1905,p.237.
channels there is a gradual accumulation that
warms
the tissuesand
thus is held. This accumulation reaches its height near the close of the day's activities.At
night thebody
is inactive so that at oncethe production of heat is lessened.The
storedup
heat energy of theday
therefore is slowly dissipatedand
is replaced only in part, so that there is a steady lessening of thebody
temperature until the beginning of anew
period of activity.^Xkh
this there is a suddenjump
again in the production of heatand
a corresponding increase inbody
temperature. In the case of our small birds that migrateby
night the increased activity inducedmust
of necessitymake
a decided breakinthe dailytemperature rhythm.No
dataon
this point isavail- able, but itmust
be supposed thatthe prolongedflights that aremade
tendtobringthetemperature abovethenormal
for individuals atrest.A
part of theaccumulated heat secured during the daymay
beattrib- utedtothe continual ingestionand
digestion of food, as the nutriment thus securedwould
be provocative ofrenewed
energy.As
digestionis rapid the
stomach
is soonempty when no
food is being taken in so that the temperature of night-flying migrantsmay
be helddown
tosome
extentby
a decrease without constant renewal in the stored nutriment in the tissues.Although
in birds there seems at presentno
indication of a m?.rked seasonal variation indegree of temperature yetwe may
suppose that thetotalamount
of heatproduced by
thebody may
be slightlymore
insummer
thanin winter.As
theperiod of daily activity is oneof high temperature (consideredon
the basis of diurnal species) it will be readily seen that that period of activity ismuch
longer, in species living north or south ofthe tropics, duringa day inJune
than during a corresponding period inDecember. There
is therefore a contrast inmany
birds inthe totalamount
of heat produced in the seasons ofsummer and
winter eventhough no
birds areknown
to hibernate.Although
itmay
be supposed that the increased activity in simimeramong
our small birdsmay
be ofiFset in partby
brief siestas taken in theheat of the day, still itwould
notseem
that the diflFerencebetween
the
two
seasonswould
beam'where
near compensated.The
quantity ofbody
heatproduced
during thesummer
periodmay
therefore be considered greater than for the winter.VARIATION IX TEMPERATURE IX RELATION TO SEX
Variation in
body
temperature correlated with sex has been well establishedinmammals, where
thetemperatureof the femaleisstated to average slightly higher than in the male.Thus Roger*
records a^Richet'sDiet, de Phys., Vol. Ill, 189^, p. g6.
10
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 72 slightlyhighertemperaturein girlsthan inboys,althoughthe subjects of experimentwere
tooyoung
to exhibit striking sexual differences.Martins
^ notes a higher temperature in females than inmales of the domestic duck,and Simpson and
Galbraith^make
similar observa- tions with regard to other birds in those cases inwhich
the sex of the individuals inhand was known
at thetime of experiment orwas
determined later by dissection.A few
observationsmade by Hilden and
Stenback''do
not bear this out, as they cite higher temperatures in males than in females. Apparently these authors misunderstoodSimpson and
Galbraith as they state that these observers also record higher temperatures in males than in females although the reverseis true.
In
my own
observations there isfound
insome
species a convinc- ingagreement
with the findings ofSimpson and
Galbraith, Martinsand Roger
in thismatter, aswhere
a sufficientamount
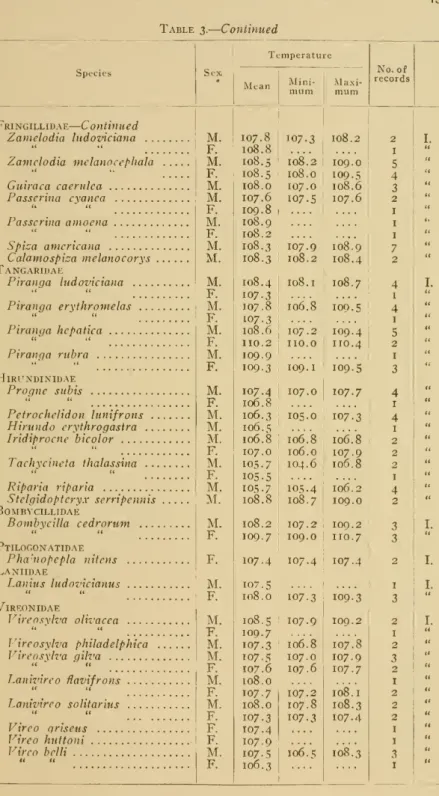
of dataisavail- able the average temperature of the female is usually slightly higher than that in the male. Certain exceptions to this rule will be notedlater.
The
difference in favor of the femalewhere
present is rather slight, being usually only a part of a degree.Thus
in the green-winged
teal (Ncttion carolinensc) the temperature of themale
(19 records) averaged 106.1°, of the female (8records) 106.6°, a differ- enceof .5° in favor of the female. InTraill's flycatcher(Empidonax
trailli) males (6 records) averaged 108.0°
and
females (4 records) 108.6° a difference of .6°.Numerous
other instances will be notedin table 3 but need not be cited here as the
two
given will serve to exemplify the statementmade
above.The
present work, however, hasemphasized
the fact that in cer- tain groups the temperature of themale on
the average is distinctly higher'than in the female. Apparently this is true in the herons (Ardeidae) as in three species in thatgroup we
have the following averages: Great blue heron(Ardea
herodias)male
(2) 104.8°, female (2) 103.7° ;snowy
heron (Egretta candidissima)male
(5) 104.8°,female(8) 104.0° ;and
black-crownednightheron(A^ychVora.*- naevius)male
(3) 103.5° ^-^d female (2) 102.6°.A
similar differ- ence is bestshown
perhaps in certain of the shore-birds, as the phal- aropes,where
it is indeed striking. In the northern phalarope (Lobipes lobatus) males (9 records) averaged 107.6°and
females (17) only 106.6°.Males
(10) of the Wilson's phalarope (Sfegano- pus tricolor) gave 106.3°,and
females (18) only 105.7°.The same
'Journ. de Phys., 1858, p. 19.
^Journ. ofPhys., 1905,p. 237.
^Skandinavischcs Arch, fiirPhys., Bd. 34, 1916, pp. 382-413.
holds true of the avocet {Kecurvirostra amcricana)
where
14 males averaged 106.6°and
12 females only 104.9°. ^^good
series of read- ings for the black-necked stilt {Hinianfopusmcxicanus) showed
thesame
average (105.8°) for both malesand
females.Through
the family Scolopacidae the general averageon
thewhole showed
a balance in favor of the males save in afew
instances.Among
the Charadriidaetwo
species, the black-bellied plover {Squatarolasqiiata- rola)and
the killdeer{Oxyechus
vociferus)show
a balance in favor of the female,while in thesnowy
plover (AEgialitis nivosa)and
themountain
plover(Podasocys montanus)
the reverseis true. Insome
passerines the
two
sexes average about thesame
in degree of bodily heat.Thus
in the yellow-headedblackbird {Xanthocephalus xantho- cephalus) males (29 records) averaged 108.3°&"d
females (18 records) 108.2°. Similarlyinthehousefinch(Corpodacus mexicanus)
males (20 records) averaged 108.9° ^.nd females (7 records) 108.8°.In these species
we
have, therefore practically anagreement
in both sexes as the difiference noted,amounting
to only one-tenth of one degreemay
well resultfrom
accident in securing the records.It appears therefore that in
many
species of birds temperatures of females are higher than those of males. In a considerablenumber,
however, thetwo
sexes average about the same,though
withmore
information itmay
befound
that there is a slight difiference in favor of the females. In afew
cases there isfound
a higher temperature in males than in females. This last is true inthe Ardeidae, the Pha- laropodidae, the Scolopacidaeand
in part in the Charadriidae.Inthe average run of species of
homoiothermal
animalsthe differ- ence in temperaturebetween male and
femalemay
be ascribed to the needs of sexual activityand
reproduction.At
least this higher tem- peratureseems
correlated with certain phases of reproduction. Inmany
birds the care of theyoung
devolvesupon
the femaleand
she hasthe higherbody
temperature. In the Phalaropodidae, onthe con- trary, it has long beenknown
that the duties of incubationand
the rearing of theyoung
fall to the lot of the male, so that in connection with this itwould
appear that he has developed a higherbody
tem- perature than the female. In the case of the Scolopacidae the writeris prepared to state
from
hisown
observations that incubationand
brooding of theyoung may
fall largelyupon
the male in the willet (Catoptrophonis inoniafus)and
it is suspected (though not yet proven) that thismay
be true of the majority of the species in this family.Males
of thisgroup examined
in breeding season often exhibit areasupon
the lower surface of thebody
bare of featherswhere
the skin is thickenedand
vascular as in birds that have been12
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. ^2 incubating. This is true also in the Recurvirostridae. In the case of theheronsdefiniteknowledge
is lacking so that I ventureno
state-ment
in regard to their sexual aberration inbody
temperature.Admitting
that the incubation of eggsand
the brooding ofyoung
necessitatea higher temperaturein theparent, itappears thatin those specieswhere
this duty fallsupon
the female shehasa higher average temperature than the male.Where
themale performs
these duties the reverse is true.As
inmany
cases, in particularamong
the pas- serines, these cares are shared about equallyby
both parents,we may
expect in such species a close approximation in
body
temperature inthetwo
sexes,a supposition thatis wellborne outby
thedatagiven in table 3.On
the basis of this reasoning itmay
be permissible to theorize further with regard to the shore birds. Inmany
species here the cares of the family are undoubtedly sharedby
both parents though as has been said, inagood many forms
this duty falls on the males alone.However,
in those that have been investigated, the greater partshow
a higher temperature in males than in females.In this
group
then,where
themale
is thehome drudge
taking overallthefamilycares
and
leaving the femaleinfreedom
after the deposi- tion of the eggs the conditionmay
beassumed
tobe aprimitive one.Males
of other species havebecome
emancipated in partfrom
this domesticyoke
so that the task of rearing offspring is shared in partby
theirspouses,though
thishas occurred so recently thatadjustmentis not complete
and
thebody
of themale
still develops a higher average temperature.Such
statementshowever must
be taken with reserveand
cannot be considered asapplyingtoother groupsofbirds.EXTERNAL TEMPERATURE IN RELATION TO BODILY HEAT
In the cold-blooded vertebrates heat generation within the
body
isslow, while the processes that act in controlling radiation are imper- fectly active. In consequence the animalchills or is
warmed
in closeharmony
with fluctuations in heat of its surrounding element.Such
creatures of necessity are sluggishwhen
they encounterlow
tempera- turesand become more
activewhen
wellwarmed. When
cooledbelow
a certain pointtheybecome
torpidand
dormant. Itwould seem
that animals of such habithave
themeans
utilized in equalizing or resist- ing high temperatures better developed than those thatmight
assistthem
inovercoming
cold.Otherwise
turtles, frogs, or lizardswould
bekilledwhen
basking inthe intense heat of amidsummer
sun. This equalization of heatmust be
accomplished largelyby
the lungs in Reptilia, as skinglands thatmight
serve this purposeare absent.The
amphibians, with their poorly developed lungs, gain the
same end
by evaporationand
radiation through their moist glandular skins.There
isadefinitelimit, however,tothe degreeof heat that these cold- blooded animalsmay
endure. InArizona
Ihave
seena Gilamonster {Heloderma suspectum)
freshfrom
the desert perishwhen
the sack inwhich
itwas
confinedwas
inadvertently left exposed in the noon-day
sun for a period of fifteen or twenty minutes.Although
in these cold-blooded animals there is a direct reaction toexternalcold,withbirds the caseisentirelydifferent.Some
investi- gatorsinmaking
studiesof avianbody
temperatures have beencareful to record the temperature of theatmosphere and
to cite this data in connection with their other records. Afterdue
consideration Ihave
notdone
this as Ido
notconsider that there isany
constant relationbetween
thenormal
temperatureof the surroundingmedium and
that of thebody
cavity in birds. Aftermaking
careful records of avianbody
temperatures at all seasons of the year, Iam,
in light of the records available at present, unable to recognizeany
constant dififer- encebetween body
temperaturesmade
in thesame
speciesat seasons ofmarked
heat ormarked
cold.Where
the individual is innormal
healthand
is sufficiently suppliedwith food, the agencies of tempera- ture control will tend to maintain an evenbody
heat.Any
variation thatmay
occur, other than that incident to the daily rhytlimic riseand
fall ofbody
heat,may
be attributed tosome
other condition thatunder normal
conditionswould
disappear within a comparatively short period through a readjustment of the bodily functions.Any
bird
may,
through inclement exposure,become
thoroughly chilledand
sohave
a greatly reduced temperature but such a condition cannot be considered normal.Thus
animmature
white-faced glossy ibis(Plcgadis guaraiina)
exposed
for half anhour
to a severe rainand
hail storm
became
so chilled that it could scarcely standand shook
violently with cold.When warmed
with hot towelsand
dried out oncemore
itwas
restored to itsnormal
conditionand
soonwas
run- ning abouton
the floor of the laboratory so far recovered that itmischievously
began
to torment other smaller birds confined with it.Birds,however,
may
be dividedroughlyintotwo
classeswith regard totheir ability to adjust to external temperature.The
first category includes those able to withstandany
reasonable degree of cold, while in thesecondare included those species that migrate to regionswhere
cold inany
degree is not encountered at the approach of inclement weather.Broadly
speaking, the question of differencebetween
thesetwo
groupsis not somuch
one ofchange inexternaltemperatureas it isoffoodsupply.Thus
species that feedon
flyingorcrawlinginsects,14
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 72 oron
fresh fruits,must
leave before a supply of this food fails.Others that search out insects in hibernation, dried berries, or live
on
seeds,pay
littleattention totheapproach
of chilling weather.The
question
seems on
thewhole one
of adequate food supply, that theorganism may
receive its life-giving elements constantly.However, forms
that habitually experience cold weather during a part of the yearmust
have a greater development of reaction for temperature regulation thando
those of the other groups.Some
speciesfrom
the category of thoseaccustomed
only to a hot climatemay
experience severe cold withoutharm
if supplied with proper food. Others suc-cumb under
theseunusualconditions. Thus,Mr. N.
Hollister, Super- intendent of the National Zoological Park, informsme
that red, blueand
yellowmacaws (Ara
niacao) confinedinlargeflightcages, remain outdoors atWashington,
D.C,
during the winter in perfect health.An
alliedspecies however,theblueand
yellowmacaw {Ara
araramia),
was
unableto withstandthe coldand
perished,though
itthroveduringwarmer
weather.From
information available itseems
that both of thesemacaws
in theirnormal
range inhabit the tropical zone,and
are subjected to thesame
general conditions of life.There
is amarked
decrease inbody
temperaturewhere
food is not obtained in suitableamount
while thebird is subjected to cold. Thismay
be seenreadilyamong
oursmaller insectivorous birdswhere
they arecaughtby
asudden
return of cold weather duringtheirnorthward
migration in spring. Decrease in bodily temperaturefrom
this causemay
be illustratedby
the following:During
the latter part ofMay,
1916, Iwas
stationed at a small fieldlaboratory inUtah
near the pointwhere Bear
River enters Great Salt Lake.For
several days preceding the evening ofMay
23 the weatherhad
been mild,and
small migrant birds that nested in themountains had
left the middleof the valley for the uplands.On
the night in question a coldwind
with a driving raincame on and
con- tinued until ten the following morning,and
therewas
little rise in temperature of theair until late afternoon.A few
Audubon's, pileo- latedand
yellow warblersand
an occasional small flycatcher appeared in the willows,and
untilnoon
therewas
a steady flight of swallowsdown
the rivertoward some haven
on the flats below.To
escapethe drivingwind, the latter flewlow
over the river or beat along behind shelter of the willows that fringed the stream.Hundreds
passed, travellinginlittleflocksso that for atimethereseemed
tobeno end
of the constant procession ofpassing birds.These
small birdswere
not obtaining food asno
insectswere
to behad and
in consequencemany
were
sufferingfrom
a loweredvitality. Thiswas
reflected inthebody
temperatures of individuals that I collected. Thus,
two male
violet- greenswallows (TacJiycincta thalassina lepida) registered 103.8°and
106.8°,respectively,a femaleblack-throatedgray warbler (Dendroica nigresccns) 105.4°, a femaleAudubon's
warbler {Dendroica audu- honi) 105.6°and two
purple martins(Prognc
subis) 104.8°and
105.0°.Such abnormal
records, due manifestly to lack of sufficient food,were
not included in the register giving thenormal
averageand
range of temperature.That
theywere abnormal may
be ascer- tainedby
referring to other records given for these species.Birds that remain in regions
where
they areexposed
to cold, be-come more
heavily feathered before thewinter season so thatthere is less radiation of heat externally. Correspondingly, insummer
the feathered covering is thinner,and
the feathers themselves oftenbecome more worn
so they arelessburdensome. With
increased cold there is apparentlysome
readjustment to maintain the bodily heat at itsnormal
point.Were
this not so the individualwould become
affected so unfavorablythat withprolonged exposureitwould
perish.We must
suppose amore
rapid metabolismand
a conservation of the resultant energy in order toovercome
this.Such
acondition is notdifficult toimagine in hawks, crows,
and
other birds of largesize, butis
wonderful when
such feathered mites as the kinglets,creepersand
chickadees are considered.The
part that the feetand
tarsi of birds play in equalizing thebody
temperature is difficult to state. In the majority of birds the spacefrom
the lowerend
of the tibio-tarsus to the tips of the toesiscovered withskin in
which
aredevelopedmore
orlessperfecthorny
scutes (thewhole forming one
of themost
evident reptilian features visible intheliving bird.)The
blood supplyis of fairquantity clear to the tips, of the toes, as blood tricklesfrom
slightwounds
in the foot.There
isno
evidenceof a forced circulation yet inmany
caseswarm
bloodmust
beconveyed
constantlytothese parts toavoid frost- bite.Many
ducks, grebesand
other aquatic birdsremain
during winter just south of the line of ice.Swans,
phalaropesand
loons appear in theArctic regions withthefirstbreakingup
of ice. I have seen auklets, puffins,and
murreletsswimming and
diving for hours in BeringSea
withno
apparent discomfort at a timewhen
the tem- perature of the water registered -^39° Fahrenheit. Mallardsand
other wildducks frequentlyclamber out of the waterand
standabout on ice for considerable periods without visible hardship.How
these birds
overcome
or avoid the effect of coldupon
their feet is amystery
as yet.Although
supplied Avith a certainamount
of blood, as has been stated, the feetand
tarsi of birds aremore
often cold orl6
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. ^2 cool to the touch than otherwise in spite of the highbody
heat. In fact they are seldomwarm
save in the hottest weather.Frozen
feetand
toes are notuncommon among
domestic fowls but areseldom
encountered in wild birds.The
skin of the feetand
tarsi issmooth and
oily, so that in the case of aquatic species'water does not adherewhen
the extremities are exposed to the air, thus preventing danger through the formation of ice. Aquatic birds at rest frequentlydraw up
thefeet oneata timebeneaththelong featherscoveringtheflanks,and
ducks oftenreston
frozenground
with both feetdrawn up
inthismanner,
so that relief is availablewhen
needed.In
some
groups of birds the tarsiand
at times the toes are well protectedby
a covering of feathers that prevent the radiation of heat.Such
adevelopment
isfound
in ptarmiganand
certain other grouse, in our owls of northern habitat,and
in the sand grouseand
rough-legged hawks.A
similar covering, though less denseand
heavy, is found,,however, in other birds that never encounter severe cold.Thus
thetarsusis featheredinwhole
orinpartinsome
trogons, in whippoorwills, in certain species of edible-nest swiftlets {Collo- calia)and many
others.The
entire tarsiand upper
surfaces of the toes are feathered in theOld World
martin (Delichon urbica). Al-though
this covering is present in Delichon, in thebank
swallow (Riparia riparia) a species that also breeds regularly far north in Arctic regions, the feathering is restricted to a small tufton
the posterior face of the tarsus near its lower end.Where
this covering of the legsand
feet is found in species that at presentdo
notseem
to require it for protection it
may
be supposed that it has persisted afteran
ancient need causing thegrowth
has disappeared, or thatit has developed as a correlated structure, perhaps ornamental in nature.
Thus
in tropical owls the feathering of tarsiand
toes isgreatly reduced, although in northern species it is very heavy.
On
thewhole
itwould
appear that radiation of heat through the lower extremities is comparatively slight.DIVERSE MISCELLANEOUS FACTORS IN THEIR RELATION TO BODY TEMPERATURE
Previous sections have covered various phases of variations in temperature
due
to sex, daily temperaturerhythm and
other condi- tions. It remains to consider afew
miscellaneous factors thatafifect this matter.Some
of these arenormal and some
abnormal.The
ingestion of large masses of food will frequently cause asudden
decrease inbody
temperature in a bird of small size.The
matter swallowed if cold will absorbwarmth
until it has acquired adegree of heat equivalent to that of the tissues inclosing it. In this
way
a distinct lowering of internal heatmay
be occasioned.On
Sept. 12, 1919, while watching
two
yellow-throated viroes {Lanivireo flavirons) thatwere
feeding incompany
Isaw
one aftermuch
effort swallow a very large caterpillar.Both
of these birdswere
collectedand were found
to l)eimmature
females.The
bird thathad eaten the caterpillar five minutes before itwas
killed registered abody
tem- perature of 107.2°, while the other gave a reading of ioS.i°. This difference of .9°was
attributed to heat absorbed by the largemass
ofrecentlyingested food. Thismay
be consideredanormal
variation.In small birds bathing
may
also occasion a slight decrease inbody
temperaturewhere
theplumage becomes
thoroughly wet.The
heat takenup
during evaporation incident to drying the feathersmay
occasion
an
appreciabledrop
inbody
heat.As an example
of this,a verdin {Auriparus flaviceps) that
had
just bathed, takenJune
16, 1919, near Arlington, Arizona, gave a temperature of only 106.0°, while others of this species rangedfrom
106.5° to 107.6°. Variationfrom
this cause is slight, however,and would
not be appreciable save in species of very small size.Many
persons withwhom
the writer has discussed the question of the takingand
recording of thebody
temperatures of birds have expressed the belief that the shockproduced
in, the birdwhen
it isshotissufficienttoincrease the bodilytemperaturetoa
marked
degree.Such
statements havecome
in particularfrom
physiciansand
others of similar training.Experiment and
observationhave
shown,how-
ever,that this isnot true.
On
various occasionsby
accident or inten- tion birds have been killed in such away
that theywere
instantly riddled by shot, so that all functions of the body, nervous as well as circulatorymust
have ceased instantlywhen
the birdwas
struck,and
thison
occasionswhen
the individual in questionhad no
reason to suspect danger.Temperatures
of such specimensshow
no varia- tionfrom
those of birds taken in amore normal
way.As
a matter of factithastranspired that theshockofwound
inbirds serves rapidly to reduce theirbody
heat after a period offrom
thirty to sixty seconds.Thus
a wing-tipped bird, withan
injury that iscompara-
tively slight, will be
found
usually tohave
a temperaturebelow normal
after a period oftwo
minutes has elapsedfrom
the time thatit
was
injured.With more
serious injuries the fall inbody
heatmay
be so great that a record
made on
a living bird four or five minutes afteritwas
shotissolowthatitmust
oftenbe consideredasabnormal.As
anexample
of this Imay
cite thecase of acinnamon
teal(Ouer-
quedula cyanoptcra) thatwas
struck in such away
that the sight ofl8
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL.'J2
both eyes
was
destroyed though other injurywas
not present. This bird rested quietly on the water, while Iwent
for a boat in order to retrieve it.On
reaching the spot fifteen minutes later thebody
tem- perature of theduck was found
to register only 102.0°, a reading considerablybelow
normal.Many
otherexamples
of a similar sort in the case of birds bearing only slightwounds
havecome under
observation.The
rapid reduction inbody
heat isdue
perhaps to anabnormal exchange and
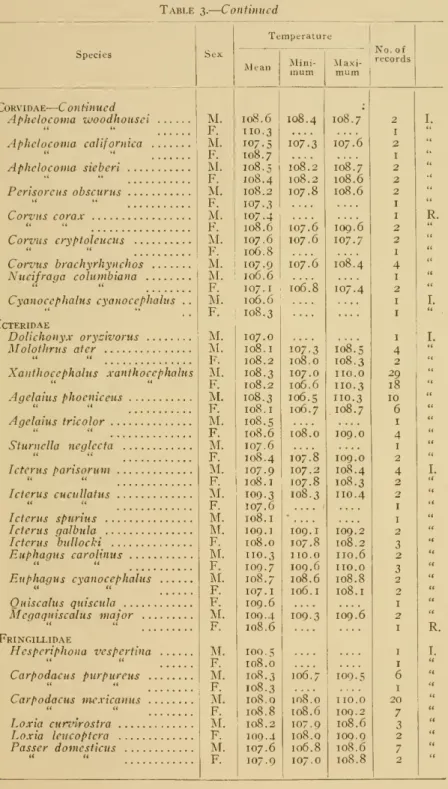
radiation through the air-sacs.TEMPERATURE OF YOUNG
During
the course of this investigation occasional opportunitieswere
presented for securing temperatureson
nestling birds oron young
of species that leave the nest as soon as hatched.These
have not been used in securing the average temperatures for each species given in table 3 or for the family records in table 4, as theyshowed some
variationsfrom
readings foradults.The
results obtained are, however, of considerable interestand
are presented in tabularform
herewith in order that theymay
be discussed briefly. In this table thefew
species included aregrouped under
family headings. In the secondcolumn
is given the temperatureand under remarks
isincluded astatementof the
approximate
ageof the individualand
themanner
inwhich
the temperaturewas
taken. In certain passerine species,where
axillar temperatureswere
taken, the end of the ther-mometer was
held closely in the hollowbetween
the foldedwing and
thebody. Birds utilizedfrom
thesame
nestor brood aregrouped
in brackets.Of
the species listed those belonging to the families Colymbidae, Laridae, Anatidae, Rallidaeand
Recurvirostridae are precocial while those listedunder
Columbidae, Tyrannidae, Hirundinidae, Mniotilti- dae,Mimidae,
Paridae,and Turdidae
arealtricial.A
differenceinthe temperature records in thetwo
groups is readily apparentupon examining
the table. In thegroup
of precocial birds there ison
thewhole
less variationand
the temperatures given closely approxi-mate
those of adults of thesame
species.The
onlywide
divergence in thisgroup
is in the case of the three-day oldyoung
of the black- necked stilt {HivianfopHs incxicanus). It so happened, however, that the temperatures of these three birdswere
takenupon
a coldraw
day,when
theyoung were
veryevidently affected by the external cold, perhaps through lack of sufficient food.Inthe case of the altricial species considerable variation is present
and
these birdsshow
amuch
lower average temperature than adults.NO. 12
BODY TEMPERATURE
OF BIRDSWETMORE
Table2. Tableoftemperaturesofyoungor nestling birds
19
Species
20 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 72 Table 2.—
Continued Species
trol is not fully
matured
until theyoung
leave the nest.The
earlydevelopment
of this faculty in precocialyoung
is in line with theiradvanced
stage as regards securing food, general activity,and
ability to care for themselves.According
toPembrey/
similar statements regardingyoung
birds have beenmade by
Edwards.^METHOD OF TEMPERATURE CONTROL IN BIRDS
Bodilyheat in all animals is causedby
tissue changes during activework performed by
various organsor parts. Mills'states thatbodily heat,though
arising in great partfrom
actual oxidations that take place in the system, is in its entireamount
best defined as the out-come
of all chemical processes that take place in the organism. In so-calledcold-bloodedvertebratesthecombined
energyor rateof these chemicalchanges is slow, so thatheat is given offby
thebody
almost as rapidly as it is generated. In the groups thatwe
class aswarm-
blooded,
Aves and Mammalia,
these changes aremore
rapidand
intense so that heatgenerationmay
beinexcess of heat radiation. In thewarm-blooded group
thereis alsoamore
orlessperfect control of heat radiationwhen
thebody
isnormal
in health. Inhomoiothermal
animals there is, therefore,an
approximation to the maintenance of a fairlyuniform
internal temperature,and
the animal remains inde- pendent of the ordinary riseand
fall of the degree of heat of itsexternal
medium.
Bayliss^considers control of heatproduction (probablyinmuscles) as the primitive
method
of temperature control.Among homoio-
thermal animals, theMonotremes
(bothEchidna and
Ornithorhyfi- chus) have the lowestbody
temperatures, as the average for these species is only 85.6° Fahr. Inthe case ofEchidna
all regulation of temperature appears to be throughchange
inheat production as this animalpossessesno sudoriferous glandsand shows no
apparentchange
in respiration athigh temperatures. Incoldweather ithibernatesand
maintains a temperature only slightly above that of the air.The
duck-bill (Ornithorhynchus) has the
power
of regulating both heat lossand
heat production so that its temperature is maintained at amore even
level.The
marsupials are intermediate in this respectbetween monotremes and
highermammals.
'Schafer, E. A., Text-bookof Phys., London,Vol. I, 1898, p.804.
*"
De
I'influence des agens physiques sur la vie, Paris, 1824. (Not seen by the present writer.)*Animal Physiology, p. 461.
*Principles of General Physiology, 1915,pp. 458-459.
22
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 72It has been estimated
by Helmholtz
^ that in thehuman body
heat lost through transpired airamounts
to 5.2^, through the water of respiration to I4.7;»and
through the skin to 77.5^.The
remainingamount
disseminated isgivenoff inegesta orisconsumed
inwarming
ingesta.
The
partplayedby
theskinglands inregulatingtemperature inthemammal
is readilyseen.The
distribution of skinglands varies in different groups,though
such glands areknown
in all save Cetaceans, elephants. Echidna,and some
others. It is claimed, for example,that inthedog
skinglandsare present onlyinthelegs or feet butin thiscase theopen mouth and
protrudingtongueactasorgansfor reducing excessbody
heat.The
presence of skinglands in thehorseis readily observed in
an
animalthat is hard driven inwarm
weather.Amphibians
agree withmammals
in the presence ofmany
integu-mentary
glands, though in thisgroup
the use of these structures is insome ways
different in purpose. In reptilesand
in birds, thetwo
classes joined in the supergroup Sauropsida, skin-glands are practi- cally
wanting and no
case isknown
inwhich
glands similar to those inmammals
are found. In birds thedevelopment
of feathers with their filamentous barbsand
barbules, as abody
covering,would
nothave
been possiblehad
sudoriferous glands been present in the skin.Excretion of fluid through such glands inevitably
would have
soiled such delicate structures as feathersand
ultimatelyhave
destroyed them.The
diffusion of heat through the skin in birds is confined to theamount,
notably small in quantity, that is, given offby
direct radiation. Itisa facteasyof verificationthattheskinin birdsisdefi- cient inbloodsupplywhen compared
withmammals. Only
acompara-
tively small
amount
of blood, therefore, can becooledtoany
extensive degreethrough the agency of the skin.As
a matter of fact the feathers thatform
a loose covering over the bodiesof birdsare notadaptedto the radiation ofheat buton
the contrarytend to conserve itand
hold it within.Though
thecontour feathers lie smoothly oneupon
the other yet they arepermeated and
separatedby
innumerable air-spaces varying in sizefrom
the tiny intersticesbetween
barbs, barbules,and
barbicels in individual feathers, to the broader areas separating one featherfrom
another.These
allgo
tomake up
series ofmore
orless closedair cells that act efficiently as non-conductorsand
serve to retain the bodily heat within.The
use of so-called "dead " air-spacesbetween
walls as a protection againstconduction of heatand
cold is too wellknown
tomake
furtherexplanation ofthis factor necessary.'Smith, R. M., Physiology of Domestic Animals, 1889, p.696.
This lack of heat regulation
by means
of the skinwould throw
the vitalwork
oftemperaturecontroldirectlyupon
the respiratory system.In this factthen
we
have a ready explanation forthe presence of the great series ofpulmonary
air-sacs that are developed throughout the avian class as a whole. Birds in order to maintain a high rate of metabolism, necessarytocontinuedactivitywithoutreferenceto shiftsand
changes inthetemperatureof their surrounding media,have
been forced to develop an auxiliary to the smallamount
of heat thatmay
be
thrown
ofifthrough the lungs. This has led to the evolution of the air-sacs that,while connectedby
ostia directlywith the lungs, radiate throughout thecoelom and
penetrate the bones toserve asan
agency of temperature control. In other words, safety to theorganism demanded
that if activitybe
greatand
continued, there besome
safe release for the excess heat developed during rapidmuscular move-
ment.The
proper function of the air-sacshas been amoot
point formany
years
and
has given rise to considerable discussion.Some have
con- sidered thatthese sacs acted as reservoirs toreplenish air inthe lungs, as containersthat, balloonlike, raised the weight of the birdinflight,orthat the presenceof these
open
spaces reduced the relative specific gravity of the body.While
the idea of the true use of theair-sacs inbirds asorgansoftemperaturecontrolwas
arrivedatindependentlyby
the present writer, subsequentlyan
admirable exposition of thesame
fact hasbeenfound
in an accountby
J.-M. Soum."" This author inturnbelieved that thediscoveryofthis factoriginated inan
hypoth- esis firstadvanced by De
Vescovi.^W.
P. Pycraft' also has adopted thisview
as he states that "the air stored in these reservoirs serves not only for respiratorypurposes, but also as regulators of tempera- ture, therebycompensating
for the lack of sweat glands."With
thiscomment,
however, he goes nofurther, ashe
givesno
detailstosupport this statement.*M. Soum,
however,made
an admirable exposition of his hypothesis.He
pointed out that all birds possess air-sacs, have a covering of feathers,and
lack skin glands,and
allhave
a high temperature.To
correlate these factshe
believed it necessary to consider the air-sacs asameans
oftemperature control.The
addi- tional facts that Iam
abletobring forth leaveno
doubt as tothe cor- rectness of this belief.*Soc. Linn, de Lyon, Vol.XLII, 1805, pp. 153-157.
^Res Zoologicae, Ann. i, No. i, Rome. (This publication I havenot seen.)
'Historyof Birds, London, 1910, p. 17.
*Withregardto statementsbyotherauthors consultalso Headley, Structure
andLife of Birds,<