I would especially like to thank Sujay and Toshiki, who were invaluable to me as I learned the electrophysiology of patch clamps. I would also like to thank my committee members, Dave Weaver, Rich Breyer, David Jacobson, Fred Lamb, and Craig Lindsley, who through their insightful questions and suggestions encouraged me to think about my project critically and produced the most best possible for my work. work. I would like to acknowledge my financial support from grants to Jerod, my NIH F31 fellowships, training grants, and travel awards that allowed me to conduct research as well as travel and share it with my peers.
I also have to thank all my friends back home in Arizona, across the country and in Nashville. I would also like to thank all my friends I made during my time in the Department of Pharmacology, the student association for Pharmacology graduates, including Aparna, Mark, Sheryl, Jamal, Kayla, Bre, Kristin and James. Finally, I would like to thank my immediate and extended family at home for their support throughout my time in college, especially to Paty, Mason, Alyce, Jackie and the Tinoco family.
I would like to thank my parents, Maximino and Alejandra, for instilling in me a strong work ethic, dedication to learning, and kindness to others that have prepared me to succeed in my endeavors. I also thank my parents along with my brothers, Max and Adrian, for their love, encouragement and support whenever I needed them.
INTRODUCTION
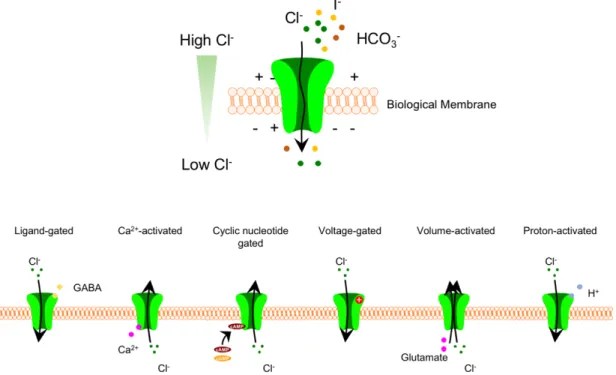
These mutations in CLC-1 caused changes in voltage sensitivity, as well as changes in the relative anion/cation permeability of the channel [27, 28]. Furthermore, iodide flux assays in combination with siRNA screening have led to the discovery of the genes encoding VRAC and PAC, again highlighting the utility of these powerful ion-sensitive dyes [51, 52]. I have summarized many of these discoveries below, as well as discoveries made before the discovery of the LRRC8 gene.
Expression of the LRRC8D subunit has been shown to be important for the transport of platinum-based anticancer drugs, taurine and blasticidin [98]. The final homomeric LRRC8D structure had a resolved N-terminal helix, providing further evidence that the N-terminal region of LRRC8 subunits forms part of the channel pore [ 66 ]. VRAC has also been proposed to act as a regulator of the cell cycle and cell proliferation.
However, the lack of specificity of these compounds raises concerns about potentially confounding off-target effects of the drugs [173–177]. The differences in potency between DCPIB suggest that the length of the oxy-carboxylic acid group is important for VRAC inhibition. With the recent discovery of the genes encoding VRAC, many of these roles have been verified using genetic approaches.
The recent cloning of genes encoding VRACs provides unprecedented opportunities to study the molecular physiology of these important channels.
CYSTL1 RECEPTOR ANTAGONISTS PRANLUKAST AND ZAFIRLUKAST
In this study, we performed a high-throughput screen using HEK293 cells for novel small molecule modulators of VRAC. Interestingly, CysLT1R has been implicated in VRAC modulation [216], but we have shown that HEK293 cells do not functionally express the CysLT1 receptor. HEK293 cells were grown in 75 cm2 flasks with Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (P/S).
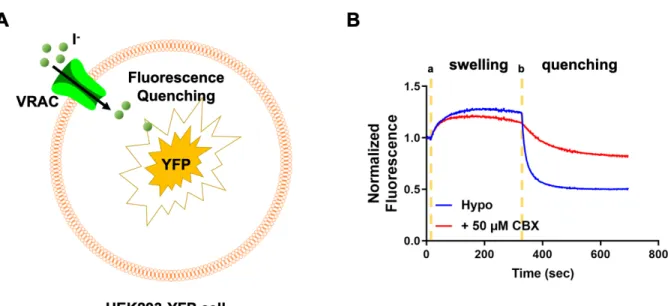
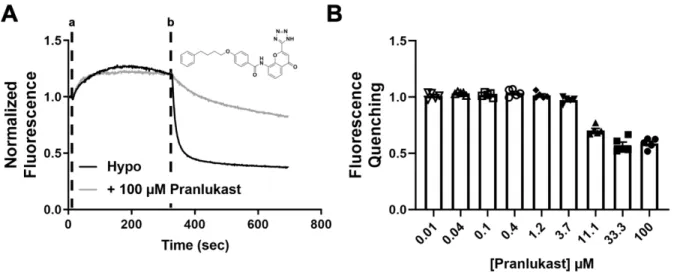
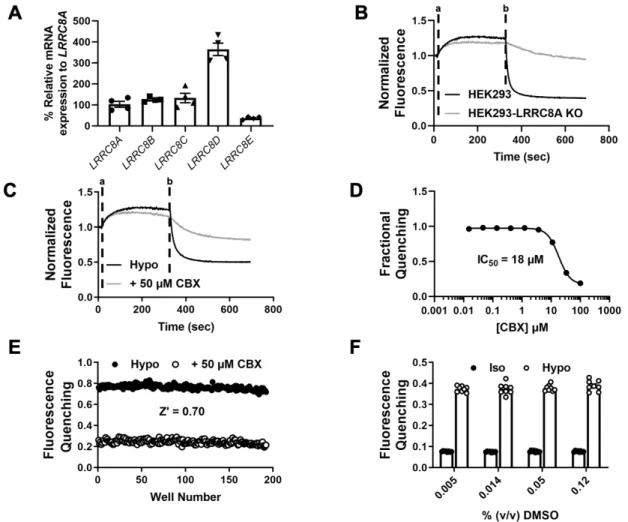
We evaluated the ability of the assay to quantify the inhibition of VRAC by the known inhibitor carbenoxolone (CBX). Hypotonic solutions (hypotonic (hypo) bath; hypotonic bath + 100 μM Pranlukast) were added at time “a” and NaI was added at time “b”. .. B) Dose-dependent inhibition of VRAC using Ozzy quenching assay (n = 5). We determined whether, despite the low abundance of CysLT1R mRNA in HEK293 cells (Fig. 8), sufficient receptor protein expression is present to mediate the effects of Pranlukast on VRAC activity.
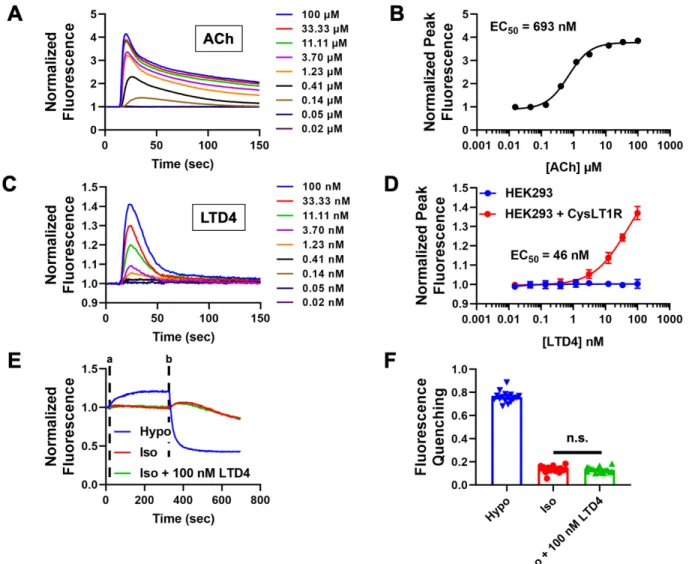
HEK293 cells were plated in 384-well plates, loaded with the fluorescence indicator dye Fluo-8 AM, and then treated with increasing doses of acetylcholine (ACh). It has been postulated that cell swelling leads to the synthesis of leukotrienes and potentiation of VRAC via the CysLT1 receptor [216]. Taken together, the results of these experiments are consistent with the interpretation that HEK293 cells lack both CysLT1R mRNA and protein expression required for receptor-mediated inhibition of VRAC by Pranlukast.
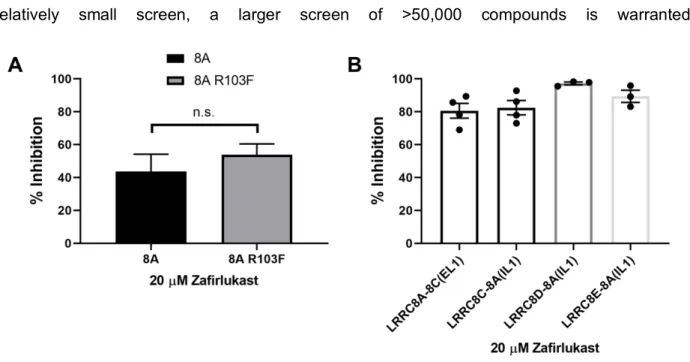
HEK293 cells were transfected with either GFP (control) or CysLT1R and GFP and patch clamped in the whole-cell configuration. Representative traces of HEK293 cells treated with hypotonic, isotonic, or isotonic + 100 nM LTD4 solutions using Ozzy quenching assay. Given that VRAC plays key roles in both of these physiological processes [6], we determined whether Zafirlukast inhibits VRAC in HEK293 cells.
We took a molecular target-based HTS approach to discover novel small-molecule modulators of VRAC. Our results agree with another study that found undetectable levels of CYSLTR1 mRNA expression in HEK293 cells [ 227 ]. Taken together, these data support the conclusion that VRAC inhibition by Pranlukast does not require either CysLT1 receptor expression or function in HEK293 cells.
ZINC PYRITHIONE POTENTIATES THE VOLUME-REGULATED ANION
Cell swelling activates a process known as regulatory volume depletion (RVD) in which chloride and potassium ions, organic osmolytes (eg, glutamate, taurine), and osmotically forced water are transported out of the cell, returning cell volume. in its normal state. VRAC can be functionally reconstituted by heterologous expression of the obligate subunit LRRC8A together with at least one other LRRC8 subunit LRRC8C/D/E [ 51 ]. Current activation is defined as the point for which the current increases continuously above the baseline current (ie, the current measured with little fluctuation since the start of the experiment).
The fluorescence values from each well were normalized to baseline readings (i.e., the average of the first five readings before adding the hypotonic solution). Peak fluorescence was calculated by taking the average trace uptake of each compound screened on the plate and subtracting it from each trace of the compound on the same plate. We recently reported the discovery of the CysLT1 receptor antagonist Pranlukast as a novel VRAC inhibitor in a screen of the SelleckChem collection of 1,184 FDA-approved drugs [45].
The HTS assay used in this screen reports the quenching of the YFP Ozzy variant by iodide when the anion enters osmotically swollen HEK293 cells through endogenously expressed VRACs (see Methods). Traces were normalized to the mean of five baseline fluorescence measurements taken at the start of the experiment. A hypotonic solution (130 mOsm) containing DMSO (vehicle) or the drug compound was added 10 seconds after the start of the experiment.
Because the subunit composition of heteromeric VRAC channels is thought to vary between cell types [235], which may alter the regulatory properties of the channel, we tested whether PTZ could also enhance VRAC currents expressed in HCT116 colorectal carcinoma cells. In all experiments described thus far, ZPT was applied together with a hypotonic bath, raising the possibility that ZPT simply potentiates the same regulatory process that activates VRAC during hypotonic cell swelling. To test this hypothesis, we evaluated the effects of the broad-spectrum ROS scavenger N-acetylcysteine (NAC) and the general NAD(P)H oxidase inhibitor, diphenylene iodonium chloride (DPI) on ZPT-dependent activation of VRAC under isotonic conditions.
Genetic approaches have revealed that fusion of fluorescent proteins to the C-terminal end of LRRC8 proteins can activate heteromeric channels expressed heterologously, bypassing the importance of the C-terminal leucine-rich repeat region of LRRC8 proteins [103]. While these studies have provided insight into the regions of VRAC proteins that contribute to activation, these constructs do not exist in nature. Through reanalysis of the existing data set, we set out to identify compounds that increased the I-induced fluorescence quenching in the window immediately after the addition of I (ie, increasing the rate of fluorescence quenching).

CONCLUSIONS AND FUTURE DIRECTIONS
Riordan, J.R., et al., Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Bear, C.E., et al., Purification and functional reconstitution of the cystic fibrosis transmembrane conductance regulator (CFTR). Voss, F.K., et al., Identification of LRRC8 heteromers as essential components of the volume-regulated anion channel VRAC.
Qiu, Z., et al., SWELL1, a plasma membrane protein, is an essential component of the volume-regulated anion channel. Gaitan-Penas, H., et al., Investigation of LRRC8-Mediated Volume-Regulated Anion Currents in Xenopus Oocytes. Sabirov, R.Z., et al., Reduction of ionic strength activates single volume-regulated anion channels (VRAC) in endothelial cells.
Trouet, D., et al., Caveolin-1 modulates volume-regulated chloride channel activity [In Process Citation]. Fujii, T., et al., Inhibition of gastric H+,K+-ATPase by 4-(2-butyl-6,7-dichloro-2-cyclopentylindan-1-on-5-yl)oxybutyric acid (DCPIB), an inhibitor of a volume-regulated anion channel. Afzal, A., et al., The LRRC8 volume-regulated anion channel inhibitor, DCPIB, inhibits mitochondrial respiration independently of the channel.
Minieri, L., et al., The volume-gated anion channel inhibitor DCPIB activates TREK potassium channels in cultured astrocytes. Wang, R., et al. A volume-regulated anion channel (LRRC8) in nodose neurons is sensitive to acidic pH. Yamaguchi, T., et al., [Preventive effect of the novel leukotriene antagonist ONO-1078 on human bronchial smooth muscle contraction induced by leukotriene C4 and D4].