(End
ofVolume)
Cijarlesi JB. anD iHarp "Paux OTalcott
i^esiearcf) Jf unb
THE LOWER EOCENE KNIGHT FORMATION OF WESTERN WYOMING
AND ITS MAMMALIAN FAUNAS
(With U
Plates)BY
C.
LEWIS GAZIN
Curator, Divison ofVertebrate Paleontology UnitedStates National Museum
(Publication 4097)
CITY OF WASHINGTON
PUBLISHED BY THE SMITHSONIAN INSTITUTION
DECEMBER
9, 1952FORMATION OF WESTERN WYOMING
AND ITS MAMMALIAN FAUNAS
(End
ofVolume)
Cljarlesi ®.
antiiWarp "^aux OTalcott
I^esiearcf) Jf unb
THE LOWER EOCENE KNIGHT FORMATION OF WESTERN WYOMING
AND ITS MAMMALIAN FAUNAS
(With
11 Plates)BY
C.
LEWIS GAZIN
Curator, Divisonof Vertebrate Paleontology UnitedStatesNationalMuseum
per\ /orb
(Publication 4097)
CITY OF WASHINGTON
PUBLISHED BY THE SMITHSONIAN INSTITUTION
DECEMBER
9, 1952BAliTIMORE, MS., V.8. A.
Introduction i
Acknowledgments 2
History of investigation 3
Occurrence and preservation of material 4
The Knight faunas 7
Correlation and age of faunas 9
Isolated occurrences to the east of the Bridger basin 13
Geologic relations 15
Systematic description ofthe Mammalia 17
Marsupialia 17
Didelphidae 17
Insectivora 19
Leptictidae 19
Mixodectidae 20
Tillodontia 21
Esthonychidae 21
Primates 22
Adapidae 22
Anaptomorphidae 24
Taeniodonta 26
Stylinodontidae 26
Edentata 32
Epoicotheriidae? 32
Rodentia 46
Ischyromyidae 46
Carnivora 50
Arctocyonidae 50
Mesonychidae 50
Oxyaenidae 50
Limnocyonidae 51
Hyaenodontidae 53
Miacidae 54
Condylarthra 59
Hyopsodontidae 59
Phenacodontidae 61
Meniscotheriidae 61
Pantodonta 63
Coryphodontidae 63
Dinocerata 64
Uintatheriidae 64
Perissodactyla 65
Equidae 65
Brontotheriidae ^7
Helaletidae 69
Hyrachyidae 70
Artiodactyla 70
Dichobunidae /O
References
n
v
vi
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7ILLUSTRATIONS
Plates
(All platesfollowingp.82.)
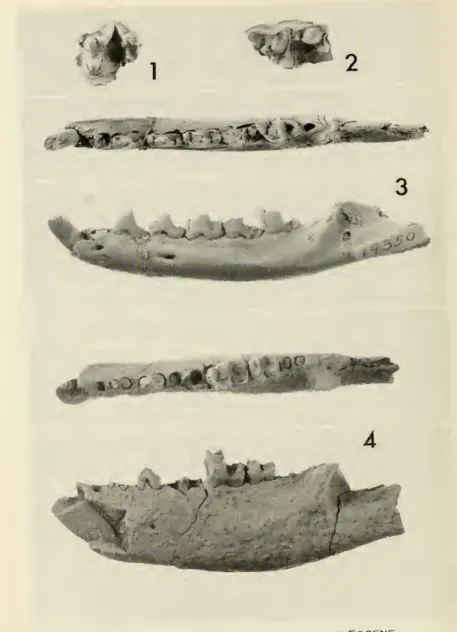
1. Marsupials, insectivore, and primates from the Knight lower Eocene.
2. Insectivore andtaeniodont from the Knight lower Eocene.
3. Taeniodontfoot fromthe Knight lower Eocene.
4. Taeniodont footfrom the Knight lower Eocene.
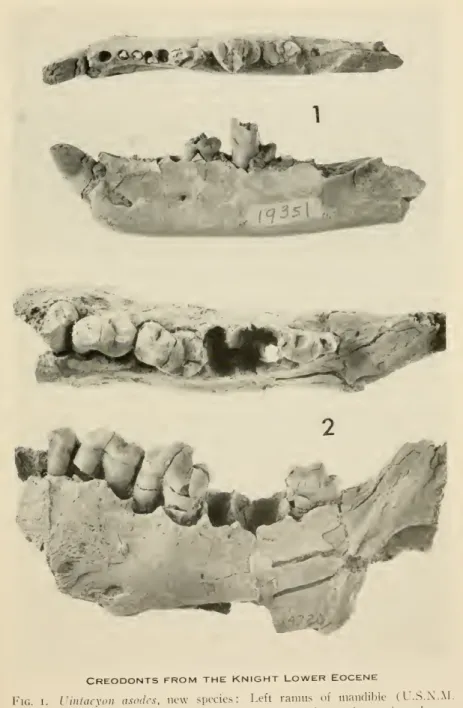
5. Creodonts from the Knight lower Eocene.
6. Creodonts fromthe Knight lower Eocene.
7. Meniscotheriiimfrom the Knight lower Eocene.
8. Meniscotherium from the Knight lower Eocene.
9. Coryphodon from the Knight lower Eocene.
ID. Lamhdotherium from the Knight lower Eocene.
II. Artiodactyles from the Knightlower Eocene.
Figures
1.
Map
ofsouthwesternWyoming
and portions of adjacent States 5 2. Pentapassalus pearcei,newgenus and species 33 3. Pentapassaluspearcei, newgenus andspecies 404. Pentapassahispearcei, newgenus and species 41
5. Tillomys senior, new species 48
6. Viverraviis lutosits, new species 56
Cfjarlefi!
©. anb iHarp ^aux
Halcott i^egeacJj jFunbTHE LOWER EOCENE KNIGHT FORMATION OF WESTERN WYOMING AND ITS
MAMMALIAN FAUNAS
By
C.lewis GAZIN
Curator, Division of Vertebrate Paleontology United States National
Museum
(With
ii Plates)INTRODUCTION
The
variegated beds of theWasatch group
in southwesternWyo- ming
have been ofmuch
interest tobothgeologistsand
paleontologists since their descriptionby Hayden
in 1869.To
vertebrate paleontolo- gists significance lies largely in their having yielded the firstknown
lowerEocene mammals from North
America. Unfortunately,how-
ever,the
term "Wasatch" was
toobroadlydefined for precise geologic usageand
included strata ofmore
than one geologic age. In conse- quence of this,Veatch
in 1907 redefined the term as a groupand
divided it in ascending order into theAlmy, Fowkes, and
Knight formations.The
last-named formation, including the fossiliferous lowerEocene
horizons, is the primary concern of this report.Investigation
by
the Smithsonian Institution of themammalian
faunas of theKnight
formationwas
largely an outgrowth of the discoveryby U.
S. GeologicalSurvey
parties of Paleocene vertebrate remains in underlyingAlmy
deposits (Gazin, 1942), Collectionsby
J. B. Reeside, Jr.,
W. W. Rubey, and
B.N. Moore
in 1936and by Rubey and John Rogers
in 1939 ledus in 1941 to a careful examina- tion of theAlmy
beds as exposedon La Barge Creek
in Lincoln County,Wyo., and
to asomewhat
desultory search of the nearbyKnight
exposures alongGreen
River in Sublette County.Examina-
tion of thetwo
sets of bedswas renewed
in 1948,and
an intensive search of theKnight
exposureswas made
alongthe east side ofGreen
Riverfrom
the vicinity of the"Three
Bridges" southeast of Big Pineyto 10or 11 miles southon
FogartyDraw.
In 1949searchwas
extended to both sides of theGreen
River, on the west side as far southasLa
Barge,and
toabout 12 milesnorthofBig Piney. In 195SMITHSONIAN MISCELLANEOUS COLLECTIONS, VOL. 117, NO. 18
2
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7 re-examinationwas made
of all themore
profitable localitiesand
in- vestigationwas
carriedupward
intotheNew Fork member,
a tongue ofWasatchian
strata interfingering with the lowermostGreen
River beds in thesame
area.Collections
made from
the upperGreen
River basin, together with materialsfrom
theKnight
beds in the vicinity of Fossiland
those oftheearlier collectionsfrom
theKnight on Bear
River, result in ournow
recognizing at least four horizons for fossilmammals
in pre- Bridgerian Tertiary of southwesternWyoming. These
include the Clarkforkian Paleocene stage represented bymammalian
remainsfrom
theAlmy
onLa Barge
Creek, equivalent topart of the originalWasatch;
a Lysite or aboutmiddle-Wasatchian stage represented by collectionsfrom
the type locality for the Knight,and
probably theKnight
at Fossil;and
apparentlytwo
horizons of general Lost Cabin equivalence supportedby
the collectionsfrom
theupper Knight
bedsand New Fork
beds along theGreen
River in Sublette County.The
lowerWasatchian
orGray
Bull has not been certainly recognized, except in the adjacentHoback
basin (see Dorr, 1952).ACKNOWLEDGMENTS
In addition to the
above-named members
of theU.
S. Geological Survey, I wish toacknowledge
helpful information givenme by
Dr. G. E. Lewis,who made
a small collectionfrom Knight
beds near theGreen
River in 1947 whileworking
with Dr.Rubey on
the Big Piney Quadrangle.George
Shea, of Billings, Mont., very graciously aided in locating forme
materialswhich
heand
G. E.Lewis
ob- served in 1947.Acknowledgment
is alsomade
of the courtesies ex- tended in connection withmy work
in 1951by
the geologists of the General Petroleum Co., theMountain
Fueland Supply
Co.,and
the Shell Oil Co. doing exploratorywork
in thesame
area.My
field assistants in 1941 included G. F. Sternbergand
F. L. Pearce. In 1948 Iwas
aidedby my
wife, Elisabeth,and
son, Chester Gazin;
both
accompanied
F. L. Pearceand
myself in 1949.In the faunal study I
am
indebted to Dr.Glenn
L. Jepsen for per- mittingme
toexamine and
describeKnight
materialsinthe Princeton Universitycollectionsand make
comparisons with typeand
otherspeci-mens
there. Drs. G. G. Simpson, E.H.
Colbert,and Bobb
Schaefifer extended tome
facilities oftheAmerican Museum
of Natural Historyand
permittedme
to studyKnight
materialsfrom
thetypelocalityand
various type specimens of lowerEocene mammals
in theAmerican
Museum
collections.The
drawings depictingnew
forms infigures 2-6were
prepared by William D. Crockett.The
photographs of specimens in plates i, 2 (fig. i), 5-8, 10,and
IIwere made
through the kindness of Dr.G.
Arthur
Cooper. Photographs in text figure i, plate 2 (figs. 2-5),and
plates3, 4,and 9 were made by
the photographiclaboratoryoftheU.
S. NationalMuseum.
HISTORY OF INVESTIGATION
Specialhistoric interest,ashas beennoted, isinvolvedin considera- tion of the
Knight
formationand
its faunas. Fossilmammalian
re-mains from
exposures of this formation alongBear
River, near Evanston,Wyo.,
are apparently the first to be describedfrom
the lowerEocene
ofNorth
America.From
hereHayden,
or Cleburne accordingtoMarsh
(1893,p. 321), in 1871 secured theremains thatCope
(1872a) described as"Bathmodon"
radiansand "Bathmodon"
semicinctus,
and
part of the material later described as"Bathmodon"
latipes. In 1872 Cope, assisted
by Carman, and
whileworking
for theHayden
Survey, collected the type ofHyracotherium
vasacciense as well as materials thatbecame
the typesof various turtles (1872b).It is of interest that in 1872 Cope'stravels took
him
to themouth
ofLa Barge Creek
inthegeneralareaofupperKnight
bedsthatproduced the materials forming the basis of this investigation. It is unlikely, however, that hemade any
collection there.The same
year he ob- tainedfrom
thevicinity of BlackButtes, eastof theRock
Springs up-lift, teeth
from two
badlydecomposed Coryphodon
skulls that henamed
(1873b)"Metalophodon"
armatus. Apparently again in 1873Cope went
over theexposures alongBear
River, southeast of Evans- ton,and
collected the type materials ofHyracotherium
indexand Phenacodus primaevus
(1873a),and
additional material ofCorypho- don
radiansand Coryphodon
latipes.Cope's investigation in the
Knight
ended with his transfer of in- terest to the lowerEocene
ofNew Mexico and
our next record of activity in these beds is Marsh's description ofCoryphodon hamatus
in 1877. This
was
collected in about 1874,from
exposureson Bear
River,about "35mileswestof Bridger,"by
R.Veltman
of Evanston,Wyo.,
formerly a storekeeper at Fort Bridger.Veltman
first wrote^Marsh
about having discovered largebones onBear
RiverinJune
of1871.
Marsh
apparently did not followup
this information untilVeltman
wroteagaininNovember
of 1874, atwhich
time he reported1Information kindly furnished
me
by Dr. J. T. Gregory from the file of Marsh's correspondence.4 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. 11/finding a skull
and
other bones of a very large animal, offeringthem
for sale.These were
evidently of theCoryphodon hamatus
skeleton thatMarsh
eventually obtained.About
themiddleof thesame
decadeMarsh
acquired material of Hyracotheriiim,from
the vicinity of Black Buttes, thatin 1876 henamed Eohippus
pernix.William
Cle- burne, oftheUnion
Pacific Railroad, in 1875 collected the skull andjaws
thatThorpe
(1934) described asMeniscotherinm
robustum.The
latterwas found
in a cut on the old grade of the railroad about 2 miles west ofAspen
(old location).I have
no
further informationon
early paleontological exploration in these beds until 1906,when Granger and
Miller collected about a dozen specimens of smallmammals
approximately200
feetup on
the blufifs ofBear
River south ofKnight
stationand
in the vicinity of the railroad bridge.These
are specifically mentionedby Granger
in 1914 (p. 203)and
regarded as Lysite in age.More
recent active interest in theKnight was
recorded by Bonillaswhen,
in1936, withhisdescription ofthe dentitionofLambdotherium from
depositsnearLa
Barge,Wyo.,
he mentionedthefieldexploration of the California Institute ofTechnology
in these bedsand
cited briefly the fauna obtained. Their collectingwas done
largely in the vicinity ofLa Barge and
included searchon
both sides of theGreen
River. Fieldwork by
partiesfrom
Princeton University intheKnight
formation resulted in the discovery in 1939and
1940 of fragmentarymammalian
remainsinKnight
beds40
to 100feet belowGreen
River strata about 3 miles northwest of Fossil,Wyo.
Also, ajaw
of Ani- bloctonus as well asan
isolated tooth ofOxyaerm were found by them
southeast ofLa Barge
in 1941. These, togetherwiththe above- noted finds of theU.
S. GeologicalSurvey and
themore
extensive explorationand
collectingby
the Smithsonian Listitution, complete the history of paleontologicalwork
in theKnight
of the Fossiland
upperGreen
River basins, as far as it isknown
to me.OCCURRENCE AND PRESERVATION OF MATERIAL For
practical purposes theKnight
collectionsmay
be regarded ascoming from
three general localities or areas of exposurem
thewesternmost part of the State: one in the northwestern part of the upper
Green
River or Bridger basinand two
in the adjacent, small but elongate structural basin to the westwhich
includes the type section. In addition to these are afew
isolated occurrences either at remote localities tentatively regarded asKnight
or in exposures sep- aratedfrom
the generallocalitiesby
significant faulting.Fig I
-Map
ofsouthwesternWyoming
andportions ofadjacent Statesshow- ing Eocene sedimentary basins, with fossil localities in Knight formation num- bered as follows: i, Knight station, type Knight; 2, Fossil; 3. La Barge-Big Piney; 4,New
Fork; 5, Rock Springs; 6, Vermilion Creek; 7, Black Buttes,8, Dad-Baggs; 9, Great Divide basin; 10, Red Desert.
Map
reproduced from Osborn, U. S. Geol. Surv. Monogr. 55, figs. 9, 49, 1929-6 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7 Historically, themore
important of the localitieswould
be the ex- posures alongBear
River, in general betweenEvanston and Knight
station. Granger's collection
came
mostlyfrom
the bluffs south ofKnight
station,and
theonly detailed statementmade by Cope
(1872b, p. 473) regarding his collectionrefers to "upper redand
white strataon
thebluffseleven miles S.E. of Evanston, or nearthebend
ofBear
River."Except
for theCoryphodon hamatus
skeleton collected forMarsh,
the materials so far recoveredfrom
this area have been very fragmentary.Those
collectedby Granger and
Miller occurred in a yellowish or buff sandstone,somewhat
conglomeratic, henceconditions of depositionwere
not themost
favorable for preservation of the smallermammals. The bone
portion of thejaws
of the smallermammals
isvery poorly preservedand
only the teeth arein reasonablygood
condition.The
exposures near Fossil,Wyo., from which
Princeton obtained acollection, areabout 45 miles due north ofKnight
station butin thesame
basin of deposition.The
Princeton collectionwas made
in variegated bedsfrom 40
to 100 feet beneath theGreen
River forma- tion about 3 miles north of Fossil.However,
ajaw
ofPhenacodus was
obtainedby
Dr.Dunkle
for the Smithsonian Institution in gray exposures beneath the red beds just 3 miles to the east of Fossil.The
materialsfrom
near Fossil are also fragmentary but the bone hasmore
the appearance of thatwhich
occurs in the vicinity ofLa Barge and
Big Piney.The most
prolificcollectingground
inbeds considered to be a part of theKnight
formationare to befound
in the vicinity of theGreen
River in SubletteCounty — from La
Barge, approximately40
miles northeast of Fossil,toapointabout 12 milesnorthofBig Piney,Wyo.
Fossil materials of this area are
found
inmany
placeson
both sides of theGreen
River; however, certain small patches or coves of ex- posures have provenmuch more
productive than others. Materials herewere
encounteredat various levelsfrom
asnearas about 40 feet belowGreen
River strata, to possibly 100 feet lower.A
collectionwas
also obtainedfrom
theNew Fork
tongue ofWasatchian
bedsin the lowerpart of the
Green
River section along AlkaliCreek and
New
Fork, tributaries of theGreen
River to the eastand
southeast of Big Piney.The
Smithsonian collections,which
arealmost exclusivelyfrom
theLa Barge-Big
Piney area,number
a little under600 and
includema-
terials in various states of preservation
and
completeness. Excellentskulls
were
found ofsome
forms,showing
varyingamounts
of dis- tortion, but less in general than exhibitedby
materialsfrom
lowerEocene
deposits in the BigHorn, Wind
River,and San Juan
basins.Not
allspecimensare free ofaniron-oxide coating,but the prevalence of this conditionisfar less than in theBigHorn
basinWillwood ma-
terials.
No
one horizon or lithology can be regarded as exclusively fossiliferous, except locallyand
for short distances, although certain dark reddish to purplish zones to the east of theGreen
River have been relativelyproductive,and
a softblue-gray shale at certain placeson
the west side betweenLa Barge and
Big Piney.A
rather profit- able concentration of materialswas
encountered about 12 miles north ofBig Pineyin reddish-buff layersand
gray sandy shale.Isolated occurrences include the
Aspen
locality for the type of Meniscotheriiimrohustum
(in theGreen
River basin of depositionand
hence not to be confused with the type Knight),and
the occur- rence ofCoryphodon and Hyracotherium
near the upper reaches ofDry
Piney Creek, referred to in 1942. In the eastern part of the Bridger basin,around
theRock
Springs uplift, isolated occurrencespresumably
inthesame
formationincludea Meniscotheriumrohustum
skullcollected
by Roland W. Brown
justsouthwest ofRock
Springs;a smallcollection of fragmentary remains that I
made from
near the southern extremity of the upHfton
a tributary of Vermilion Creek;and
the Black Buttes occurrence tothe eastof the uplift. Stillmore
remote, butnevertheless in thesame
depositional basin,may
bemen-
tioned the occurrenceson
the easternmargin
of theWashakie
basin beneath the Tipton tongue ofGreen
River betweenDad and
Baggs, referredto inWood
el al. (1941, p. 18),and from
northeast of Cres- tonmade by
aU.
S. GeologicalSurvey
party in 1907.THE KNIGHT FAUNAS
The
following tabulation pertains to the principal occurrences of fossilmammals
in theKnight
formation as itis exposed in the upperGreen
River basin (theLa Barge and New Fork
faunas)and
in the adjacent small structural basin west of the OysterRidge which
in- cludes thetypeKnight and
theoccurrenceat Fossil,Wyo. The
figures used referto thenumber
of specimensencountered, giving an indica- tion of extent of materialsupon which
identifications are based,and
asuggestion of the relativeabundance
ofvarious forms in the faunas.8 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7Knight LaBarge- New
MarsUPIALIA: station Fossil Big Piney Fork
Pcratheriiim cdwardi, new species 2
Peratherium chesteri, new species I
Insectivora:
Diacodon pineyensis, new species 3
Diacodon, cf. altictispis Cope i
Cynodontoniys knightensis, new species.. 13
Cynodontomys, species i
TiLLODONTIA:
Esthonyx, cf. acntidens Cope 3
Esthonyx,species i I
Primates:
Pelycodus? species i
Notharctus liniosus, new species 24
Notharctus venticolus Osborn 6
Absarokius noctivagus Matthew 3
Paratetoniusfsublettensis, new species... I
Taeniodonta:
undetermined stylinodont 4
Edentata:
Pentapassalws pearcci, new genus and
species 2
RODENTIA:
Paramys,cf. bticcatus * (Cope) i 2
Paramys, cf. copei *Loomis 8
Sciuratnis, possibly5". depressusLoomis. .
I
Tillomys senior, new species I
Carnivora:
Thryptacodon, near T.antiqmis Matthev/. 2
Pachyaena? species 2
Oxyaena, species I
Ambloctonus, cf. major Denison i I
Prolimnocyon elisabethae, new species... 3
Sinopa vulpeciila Matthew 5
Sinopa,cf. strenua (Cope) i
Didymictis altidcns Cope ? 14
Viverravus lutosus, new species 8
Uintacyon asodes, new species i
Miacis, cf. latidens Matthew 3
Viilpaviis asiiis, new species 6
CONDYLARTHRA:
Hyopsodiis zvortmani Osborn 29 I
Hyopsodus mentclis (Cope) 76
Hyopsodus brozvni Loomis 4
Phenacodtis primaeviis Cope I i**
Meniscotherixmn robustum Thorpe 93
Mcniscothcrimn, cf. chamense Cope 3
*Paramys, cf. buccatiis in Knight isParamys cxcavatus Loomis; and Paramys, cf. copei Loomis includes P. copei,P. major, and anewspecies, as determined by A. E. Wood.
**Recorded from ahorizon stratigraphically lower than other forms cited in this fauna.
NO. 1
LOWER EOCENE KNIGHT FORMATION — GAZIN 9
TJA•^T-t./^r^^^^T.I.A. Kttight LoBatQe- New
rANTODONTA. station Fossil Big Piney Fork
Coryphodon radians (Cope)
Xf
cf.15Coryphodon seniicinctus (Cope) i
Coryphodon latipes (Cope) I
Coryphodon hamatus Marsh i
Coryphodon, species I
DiNOCERATA:
Cf.Bathyopsisfissidens Cope 2
Perissodactyla:
Hyracotherium index (Cope) 2 i 35
Hyracothermn vasacciense (Cope) 4 67
Hyracotherium, cf. venticolum Cope
143
Hyracotherium, possibly H. cristatum
Wortman
iLambdotherimn popoagicum Cope 93
n
Heptodon, cf. ventorum (Cope) i 30
Heptodon, species I 4
Hyrachyus, species I
Artiodactyla:
Bunophorus, cf. macropternus (Cope)... 7
Diacodexis, near D. secans (Cope) I
Diacodexis, species 2%
Hexacoduspelodes,newgenusandspecies. 11
Hexacodus iiintensis, new species i
tNumberof specimensnotknown and should includematerial of at least a part of those specieslistedbelow which areprobablysynonyms.
+Specimensnot seen.
CORRELATION AND AGE OF FAUNAS
Within
the formation thatmay
properly be calledKnight
are clearlythree horizons that can be distinguishedon
the basis of fossilmammalian
remains, including theNew Fork
tongue ofWasatchian
material interfingering in the lower part of theGreen
River series.The
lowerof theseis nearestto Lysite in ageand
the upper aretwo
distinct zones of Lost Cabin age separated locally by the Fontenelle tongue of
Green
River. In the absence of evidence of any lowerWasatchian
orGray
Bull horizon, except in theHoback
basin (see Dorr, 1952), theKnight may
well be regarded as about equivalent to theWind
River series.Knight
station.—
Criticalexaminationof the materials derivedfrom
thetypeKnight
beds alongBear
River leadme
to support Granger's (1914, p. 203) conclusion that thehorizon there represented is about equivalent to Lysite.The Cynodontomys
material, as incompletelyknown,
does not appear to bemore
indicative of a Lysite specieslO
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7 thanofthelaterform now known
tocharacterize theLa Barge
fauna;
however,
Hyopsodus browni
is highly suggestive of Lysiteand
theHeptodon
materialwould
not appear to be older.The
relative fre- quency withwhich Coryphodon
remainshave been encounteredwould
appear to haveno
significance,and
although not recorded in the type Lysite they arefound
in Lost Cabin beds of bothWind
Riverand Green
River basins, as well as in theGray
Bull, so that their middleWasatchian
or Lysite representationmay
well be here.Among
the 14 to 16forms
recognized in the fauna, characteristic or peculiarlyGray
Bull types are missing, particularly such adominant form
asHomogalax.
In a likemanner,
the absence ofany
representation of Lost Cabinforms
asM
eniscotheriumand Lambdotherimn
is equally suggestive. In support of this it should be noted that, with regard tothe Lost Cabin equivalent of theKnight
in the upperGreen
River basin, almost a third of the specimens encountered are of one or the other latter forms. In a cursory examination of the upperKnight
the percentage of theseforms
would, because of their size, appear to be evenmuch
higher.Fossil.
— The
rather limitedfaunarepresentedbythesmallcollectionfrom
near Fossil,Wyo.,
is less clearly diagnostic with respect to the horizonrepresentedthan is theBear
River Knight.Comparison
with horizons of theWind
Riverand
BigHorn
basins leavesmuch
to be desired; nevertheless, its closest affinity is with the type Knight.Heptodon,
taken into consideration with other similarities to the faunafrom
theKnight on Bear
River, tend to restrict comparison to only the middleand
perhaps upperWasatchian
levels.On
the otherhand
the species ofDiacodon and Hexacodus
represented are dis- tinctly not thosefound
in the Lost Cabin equivalent of theKnight
in theGreen
Riverbasin,and Hexacodus
is notamong
the rarerforms
represented in the latter beds.Phenacodus
primaevus, thoughfound
at a
somewhat
lower level than other materialsfrom
the Fossil area, is significantly absentfrom
theLa Barge
fauna. Itmust
be noted, however, the situation with respect toPhenacodus
is reversed in theWind
River basin. Individually, the elements of the Fossil fauna are inconclusive, butcollectively thereis a strongpresumptiontoward
the Lysitelevelasthatlevel isindicatedby
theKnight
fauna obtainedfrom
theBear
River section.La
Barge.— There would
appear to beno
doubt of the generalequivalance of the
La Barge
fauna to the Lost Cabin of theWind
River basin.
There
is a specific difference betweenthem
inmany
of the genera mutually represented, but this
may
])e largely environ- mental, or possibly ofsome
smalltime significance if within arapidly evolvinggroup.Such
atime difference, however,would
notbe of themagnitude
of that between Lysiteand
Lost Cabin.The
Lost Cabin equivalanceis indicatedbythepresenceof such generaas Notharcfits, Meniscotheriiim,and Lambdotherium which
first appear, or reach the indicated development,intheLost Cabin horizon oftheWyoming
lower
Eocene; by
such species asEsthonyx, cf.acittideiis; Notharctus venticolns; Ahsarokius noctivagiis; Ambloctonus, cf. major; Sinopa vulpecula; Miacis, cf. latidens;Hyopsodus
mentaJis;Hyopsodus wortmani; Hyracothermm,
cf. venticoliim;and Lambdotherium
popo- agiciimwhich
are close or identical to such speciesmore
or less characteristic of Lost Cabin in theWind
River or BigHorn
basins;and by
the developmental stage reached in such species asCynodon- tomys
knightensis, Notharctuslimosus,and Prolimnocyon
elisabethaewhich
are otherwise distinctfrom
related Lost Cabin species.Suggestive of perhapsa slightly earlier stagethan type Lost Cabin, although as noted above these differences
may
be nomore
than en- vironmental in nature, are the following:The
smaller size, thovigh equivalent dental development, ofCynodontomys
knightensisincom-
parison withCynodontomys
scottianus in a sequence of species in- creasing in size; the smaller sizeand
relatively smaller premolars in Notharctus limosus than inA^
nunienus; theweaker
hypocone in upper molars of Notharctus venticolns of theLa Barge
fauna; the smaller size (of questionable significance) ofDiacodon
pineyensis;Esthonyx, cf. acutidens; Ahsarokius noctivagiis;
Prolimnocyon
elisa- bethae;Sinopa
vulpecida; Didymictis altidens; Viverravus lutosus;and Vulpavus
asius in comparison with thesame
or equivalent Lost Cabin species;and
the presence ofParatctonius?, Thryptacodon,and Hyracotherium
index.The
abovewould
appear to be of perhapstrifling importance in consideration of relative ages, but
on
the otherhand
there is even less to suggest a later stage than type Lost Cabin, although such might be interpretedfrom
the presence of Sciuravus, Tillomys,and Hexacodus. The
evidence for a slightly older age affordedby
Meniscotheriiim isperhapsthemost
interesting,inasmuch
as the largeM. rohustum
so farnotfound
outside of theGreen
River basin occurs stratigraphically below Meniscotherimn, cf.chamensc
of the
New Fork
tongue.Only
the smaller species, regarded asM.
chamense, is recordedfrom
the Lost Cabin beds of theWind
River basin.
12
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7The
following tabulation is a comparison of generanow known
in theLa Barge and
Lost Cabin faunas:
Lost LaBarge Cabin
Perathcriiim
X
Diacodon
X X
Parictops
X
Didclphodus
X
Palacosinopa
X
Cynodontoniys
X X
Estlwnyx
X X
Notharctus
X X
Loveina
X
Shoshoniiis
X
Tetonius
X
Paratetonius?
X
Absarokius
X X
Styl'modon cf.
X
palaeanodont
X
Tiibid'Odon
X
Pentapassalus
X
Paramys
X X
Sciuravus
X
Tillomys
X
Mysops
X
Thryptacodon
X
Oxyacna
X X
Lost LaBarge Cabin
Protopsalis
X
Ambloctonus
X X
Prolimnocyon
X X
Sinopa
X X
Didymictis
X X
Viverravus
X X
Uintacyon
X
Miacis
X X
Vulpavus
X X
Phenacodus
X
Ectocion
X
Mcniscotherinm ....
X X
Hyopsodus
X X
Coryphodon
X X
Bathyopsis
X
Hyracothcrium
X X
Lambdothcriiim ....
X X
Eotitanops
X
Heptodon
X X
Hyrachyus
X
Bunophorus
X X
Diacodexis
X X
Hexacodiis
X
New
Fork.— The New Fork
tongue, separatedby Green
River beds called the Fontenelle tongue byDonovan
(1950)from
theKnight
along theGreen
River, has yielded a faunal representationwhich compares
equally well with thatof the Lost Cabin, butincludesforms
notfound
in theLa Barge
fauna.The
Lost Cabin age is here indicatedby
Ambloctonus, cf.major; Hyopsodus wortmani; Menis-
cotherium, cf.chamense;
cf. Bathyopsisfissidens;Hyracotherium, cf.venticolum;
and Lamhdotherium
popoagicum.The
significant dif- ferencesto be notedfrom
theLa Barge
faunaare in (i) the species of MeniscotJierium represented, (2) the presence of Bathyopsisf in- steadof
Coryphodon, and
(3) theappearance ofHyrachyus.
I suspect that,were
theNew Fork
betterknown,
Eotitanopswould
be repre- sented, also Trogosus,which
isknown from
the Cathedral Bluff tongue in the northwestern part of theRed
Desert.The most
significant feature of the discovery of theNew Fork
fauna is the clear demonstration that the lower beds of the
Green
River are actually Lost CabinWasatchian
in ageand
that the LostNO. l8
LOWER EOCENE KNIGHT FORMATION — GAZIN
I3Cabin
fauna can be largely brokenup
intotwo
stages. Tentatively thelower stagemay
be regardedas characterizedby
Meniscotheriiimrohustum and Coryphodon and
the upper stage by Meniscotheriiim chamense,Hyrachyus,
species,and
possiblyBathyopsis.Osborn
early regarded the Lost Cabin as comprisingtwo
stages, but believedthem
to be distinguished by
Lamhdotherium and
Eotitanops, respectively;however,
Lamhdotherium
isherefound
inbothlevels.Much more
of theNew Fork
fauna needs to beknown
for amore
complete under- standing of Lost Cabin zones. Thismay come when
the Cathedral Bluff fauna currently under study byWilliam
Morris is placedon
record.ISOLATED OCCURRENCES TO THE EAST OF THE BRIDGER BASIN
Not
included in the foregoing tabulationand
correlations,and
for themost
partomittedfrom
consideration in the systematic treatment of the faunas, are the scattered occurrences in the eastern part of the Bridger basin,around
theRock
Springs uplift, in the eastern part of theWashakie and
Great Divide basins,and
in theRed
Desert.Itis not certainly
known
but entirelyprobable thattheformation rep- resentedby
these occurrences is also Knight.An
examination of the geologicmap
ofWyoming
leads to the conclusion that theEocene
basin of deposition is thesame
for these deposits as for those in the vicinity ofLa Barge and Big
Piney,and
that theformation is continu- ous beneath theGreen
Riverand
Bridger formations,and
around theRock
Springs uplift to thesewidely separated areas.The more
significant of these localities are discussed belowRock
Springs.— West
side ofRock
Springs uplift, near fork ofBitter
Creek and
Little Bitter Creek, about4
miles southwest of thecityof
Rock
Springs, collectedby
R.W. Brown
in sees. 7, 8, 17,and
18,T. 18 N., R. 105
W.
:M
eniscotheriumrohustum and
Coryphodon, species.The
rather fortuitous discovery of theseformsintherelatively thinzoneofWasatchian
beds betweenRock
Springsand Green
Riveris highly indicative of the upper
Knight
orLa Barge
equivalent of Lost Cabin being represented here between Paleoceneand Green
River strata.Vermilion Creek.
— South
ofRock
Springs uplift on a tributaryto Vermilion
Creek
(in type Vermilion Creek ofKing and
type Hia-watha
of Nightingale) near line between T. 11 N.and
T. 12 N., in R. 101W.,
northernmost part of Moffat County, Colo., collectedby
writer: Insectivore, possiblyDiacodon; Par amy
s hicuspis;and Hexa-
codus, cf. pelodes.
The
insectivorefound
here is not significant.14
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7Paramys
hicuspis, accordingtoA. E.Wood, who
identifiedthe speci-men,
is suggestive of Lysite; however,Hexacodus,
cf. pelodeswould
suggest an equivalence with theLa Barge
fauna.BlackButtes.
—
Eastside ofRock
Springs uplift, nearBlack Buttes, collected in part byCope and
in partby
a collector forMarsh
:
Coryphodon armatus and Hyracotherium
pernix.These
give littleinformation other than a
Wasatchian
age.Dad-Baggs. —
East side ofWashakie
basin, exposures along high-way
beneath Tipton tongue,between Dad and
Baggs,Wyo.,
collectedby
the writer:Coryphodon,
large species,and
Hyracotherium, cf.vasacciense.
A
small lot of materialfrom
nearBaggs was
collectedby
Reesideand Eby
in 1924, but this material is notnow
in the col- lections.Another
collectionfrom
nearDad was
designated byWood
etal. (1941,p. 18) as
"Dad
localfauna"and
notedbyMcGrew
(1951,p. 54) as having been collected
by
J.LeRoy Kay
for the CarnegieMuseum. Of
thematerials collectedby Kay
onlyHyracotherium was
mentioned.Of
the materialswhich
I have observednone
are particu- larly diagnostic; however, Iam
inclined to regard the horse as ofKnight
age rather than lower Wasatchian, It should be noted thatWood
et al. (1941, chart) regard the fauna as late Wasatchian. Thiswas
basedon
evidencethat I have not examined.Great Divide basin.
— About
18 miles west of Rawlins,Wyo., and
about 12 miles northeast of Creston, SE^^rSE^^ sec. 32, T. 22 N., R.90 W., Sweetwater
County, collectedby
E. E.Smith and A.
C.Veatch
in 1907 while with theU.
S. GeologicalSurvey on
theNorth Rawlins
Coalfield:Esthonyx,cf.acutidens; cf.Notharctusventicolus;Coryphodon,
species;and
cf.Heptodon,
species.The
materialsfrom
this locality in the Great Divide basin are very fragmentary, but cf.
Heptodon
suggests Knight,and
Esthonyx, cf. acutidens,and
cf.No-
tharctus venticolus
would
appear to place the horizon represented in the upperKnight
or Lost Cabin equivalent. This occurrence is very neartheeasternlimit inWyoming
ofbedswhich may
becalledKnight,still within the
Eocene
basinof depositionwhich
includes the Bridger basin proper.Cathedral Bluff tongue.
— Northwest
part ofRed
Desert,NW.
partof T.
26
N., R.98 W.,
collectedby
R. L.Nace
(1939, pp. 17, 26-27)from
75 feetbelow
top of Cathedral Bluff tongue,and
identifiedby
Simpson
:Trogosus
or Tilloiherium.As Simpson
has indicated, the evidence is rather strong for a Bridger age.However,
Iam
inclined to regard the Cathedral Bluffand New Fork
as equivalent,inasmuch
as somarked
a retreat ofLake
Gosiute as indicatedby
thesetongueswould
effect the entire periphery, unless complicated by tectonic ac-NO. l8
LOWER EOCENE KNIGHT FORMATION — GAZIN
Jtivity involving tilting or folding. In
any
case there is presumptive evidence favoringcontemporaneity of the occurrences of an extensive tongue of variegated beds encountered interfingering with the lower part of theGreen
River series in various parts of this basin. This leads to the conclusion that the Cathedral Bluffwas
deposited during LostCabin timeand
thatTrogosus may
wellhave appeared before the Bridgerian stage,much
asHyrachyus, and
have lived as a contem- porary of Esthonyx.GEOLOGIC RELATIONS
Sequence
of formatiotis.— The
sequence of early Tertiary materials in the southwestern part ofWyoming
includes as its lowest unit theEvanston;
above this theWasatch
group, consisting, as redefinedby Veatch
(1907, p. 88), oftheAlmy, Fowkes, and
Knight, is followedby
theGreen
Riverand
Bridger formations.To
the east of the Bridgerbasin,around
theRock
Springs upliftand
alongthe easternmargin
of the Tertiary basin, asshown by
R.W. Brown
(1949), Paleocene beds generally designated as "FortUnion"
are exposed, butwhose
exactage is notknown,
having so farproduced noknown
fossil
mammal
remains.Washakie
beds, overlyingGreen
Riverin theWashakie
basin, are equivalent to the upper part of the Bridgerand
include beds regardedassomewhat
laterthantypical Bridger.To
the south of theRock
Springs uplift, in the basin of Vermilion Creek, the VermilionCreek
beds of King, as definedby hismap
(1876,atlasmap
2) areapproximately equivalenttotheWasatch
ofHayden. The
VermilionCreek
includes Paleocene in the vicinity of theRock
Springs uplift,and
theTiptonand
CathedralBlufftongues as well as Knight.The Hiawatha
of Nightingale (1930, p. 1023) in thesame
general area is in a generalway
equivalent to the Knight. In the northern part of the Bridger basinDonovan's
(1950) Fontenelle tongue ofGreen
River is likely equivalent to the Tipton tongue farther south,and
as notedabove,hisNew Fork
tongue ofvariegated beds likewisemay
be Cathedral Bluff tongue, althoughtheidentity of theseisyettobeproved. Still farthernorthintheHoback
basin,bedswhich
Schultz (1914)mapped
asEvanston and
undifferentiated Eocene,and
calledHoback
formation by the University ofMichigan
fieldparties, have produceda Tiffanian
and Wasatchian
fauna (Dorr, 1952).The
equivalence of these beds with others in the Bridgerand
Fossil basin to the west has not been established.The
supposition isthatthe Paleoceneportion
may
be equivalenttotheEvanston, or pos- sibly to a part of theAlmy. The Wasatchian
horizonmay
be lower Knight, or possiblyFowkes.
Itis noted, however,that neither Evans-l6
SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7 tonnorFowkes were
recognizedbyVeatch
or Schultzelsewhere along the westernmargin
of the Bridger basin.Should
the beds carrying theWasatchian
fauna in theHoback
basin prove to be a part of theKnight
thiswould
then be the onlyknown
placewhere
bedswhich
appear to be lowerWasatchian
in age are included in the formation.Relations between basins.
— The
geologic relations involved in the present study are largely thosewhich
existbetween
the sequence of continental early Tertiary deposits in the upperGreen
River or Bridger basins,and
those in the small, north-south elongated basinwhich
includes the type Knight.The
latter,which
will be referred toasthe Fossilbasin,liesina generalway
tothewestof OysterRidge and
includes the high plateau areafrom
the upper reaches ofHam's Fork
in the northsouthward beyond
the headwaters ofBear
River.On
the west it isbounded by
the intermittentband
of pre-Tertiary rocks extending in a generalway from Tunp Range
in the north tobeyond Evanston
southward.Early Tertiary deposition in these
two
basins appears to have been entirely separateand
independent.As we may
seefrom
Veatch's (1907)map and
sections, theAlmy,
Knight,and Green
River lap successively onto older rocks along both the easternand
western margins of the Fossil basin,and
along the westernmargin
of the BridgerorGreen
Riverbasin,wherever
therelation isnot actuallyone offaulting.And
althoughthe pre-Tertiaryismuch
foldedand
faulted in the exposedbands
along the eastand
west margins of the Fossil basin, the Tertiary sediments areforthemost
part but gentlywarped
or tilted.However,
it is evident that theLaramide
activity did not completely ceaseby
lowerEocene
time so that itwould
be unsafe toassume
that therewas
entire synchroneitybeween
the deposits of the Fossiland
Bridger basins. This lack of equivalence ismore
notice- ablebetween
the Fossil basinand
that nexttoward
the west, in the absence of theFowkes and
in a scant representation ofEvanston
in the Fossil sequence. It is probable, therefore, that theKnight
to the eastand
that tothe west of OysterRidge
are not entirely equivalentand may
havehad somewhat
earlier or later beginningsand
endings, although I suspecta rather general equivalence.The same may
be said of theGreen
River serieswhich
in thetwo
basinswas
deposited in entirely separate lakes.The
ultimate reason for the existence of thesetwo
lakesmay
bethe same, or their originsmay
be tied to thesame
structural or orogenic control, but again their upper or lower limitsmay
not be entirelycontemporaneous
in thetwo
basins.NO. l8
LOWER EOCENE KNIGHT FORMATION — GAZIN
1The
evidence presentedby
thefossilmammals
in theKnight
beds of thetwo
basins is such as to suggest that the upper limit of theKnight may
have beenasmuch
differentastheintervalbetweenLysiteand
Lost Cabin time.The
Lost Cabin fauna in the upperGreen
Riverbasin extendsfrom
afew
tensof feet belowtheGreen
Riverto 100 ormore
feet,whereas
in the Fossil basin so far only a Lysite fauna has been certainly recognized. In the vicinity of thetown
of Fossil, the fauna represented is thought to be equivalent to that of the Lysite as represented by the typeKnight
farther south,and was
likewise found
40
to 100 feet belowGreen
River beds. This reason- ing also leads to supposition that theGreen
River beds in the Fossil basinmay
includesomewhat
older strata than in theGreen
River basin,and
that this lakehad
its originsomewhat
earlier in lowerEocene
time.Should
a furtherand more
intensive search of the Fossil basin demonstratebeyond
doubt the absence of beds of Lost Cabin age immediately underlyingtheGreen
River, or interfingeringwithGreen
River to the southward, one might argue theadvisability of selecting a distinct formationname
forbeds calledKnight
in theGreen
River basin. Iwould
not, however, venture tomake
such a discrimination, as the lithology is similarand
in all probability there ismore
or less overlap if not precise or complete equivalance in time. Continental deposits are often not clearly definable units, and, of course, notori- ously unreliable both as to timeand
lithologywhen
traced laterally forany
considerable distance, even within thesame
basin of deposition.SYSTEMATIC DESCRIPTION OF THE MAMMALIA
MARSUPIALIA
DIDELPHIDAE
PERATHERIUM EDWARDI,2 new
species Plate I, figures 3, 4Type.
—
Leftramus
of mandible with lasttwo
molars,U.S.N.M.
No.
19200.Horizon and
locality.—Upper Knight
beds,La Barge
fauna,SWi
sec. 33, T. 32 N., R. IllW.,
12 miles north of Big Piney, Su- blette County,Wyo.
2
Named
for G. E. Lewis,who
first calledmy
attention to the particular locality which has since yielded all the Peratherium material from these beds.l8
SMITHSONIAN MISCKLLANEOUS COLLECTIONS
VOL. 11/Description.
—
Size significantly smaller thanPeratherium com-
sfockiCope
(1884, pi. 25a, fig. 15)from
the"Wind
River" beds, nearer that ofPeratherium marsupium
Troxell (1923b)from
the Bridger beds.Cusps
of lasttwo
molars sharpand
well defined, par- ticularly the entoconidand
hypoconulidwhich
in the last molar aremuch
betterdefinedthan inP.marsupium,
as representedby
NationalMuseum
materials.Length M2-M3,
5.5mm.
Transverse diameter of penultimate molar, 1.8mm.
A
leftmaxillaryportion,U.S.N.M. No.
19206, ofa small marsupial found at thesame
locality is referred to Peratherium cdzmrdi.The
specimen retains the lasttwo
molars,which
occlude well with those of thetype.The
teethare rather wellworn and show
little ofdiagnos- tic value.The
externalmargin
of the penultimate toothshows
threesmall, worn, but about equally well-defined stylar cusps between thedistinct parastyleand
metastyle at the angles at the tooth.There
appears, however, buttwo
stylarcusps between themore
conspicuous parastyleand
metastyle on the last molar. InPeratherium
innomina-tum Simpson
(1928) material in the NationalMuseum
collectionsfrom
the Bridger, a relatively greater spread between the metastyleand
theprecedingstylarcuspis notedinthepenultimateand
antepen- ultimate molars.Length
of lasttwo
upper molars combined, 4.3mm.
Transverse diameterofpenultimateuppermolar, 3.1
mm.
PERATHERIUM CHESTERI,- new
species Plate I, figure iType.
—
Rightramus
ofmandible with penultimate molar,U.S.N.M.
No.
19199.Horicon and
locality.— Upper Knight
beds.La Barge
fauna,SW^
sec. 33, T. 32 N., R. Ill
W.,
12 miles north ofBig
Piney, Sublette County,Wyo.
Description.
—
Smaller than hithertoknown
species of Peratheriumfrom
theEocene.A
littlesmallerthantheBridgerspeciesPeratherium innominatum Simpson
(1928). Penultimate molar length about 1.4mm. and
width about0.8mm. The
alveoli of the lastthree molarsmeasure
about4 mm.
Trigonid relatively highand
talonid small with entoconidand
hypoconulidmuch
less prominent than in the larger P. edzvardi.The
tooth has points of resemblance to an in- sectivore but the characterand
position of the hypoconulidand
an- teriorcingulumseem
certainly marsupial.3
Named
for Chester Gazin,who
collected the type specimens of both P. edzvardi and P. chesteri.INSECTIVORA
LEPTICTIDAE
DIACODON PINEYENSIS,* new
species Plate I, figure 2Type.
—
Leftramus
of mandible withMi
toM, and
part of P4,U.S.N.M. No.
19204.Horizon and
locality.-— Upper Knight
beds,La Barge
fauna,SV^\
sec. 33, T. 32 N., R. Ill
W.,
12 miles north of Big Piney, Sublette County,Wyo.
Description.
—
SizenearDiacodon
alticuspisCope
(1875, PP- 11-12) but with lowermolar
teeth slightly shorterand
distinctly broader.P4
much
larger.Length
ofmolartoothseries 8.3mm.,
shorterthan in Parictops multicuspisGranger
(1910, pp. 250-251) orDiacodon
bicuspis(Cope)
(1880a, p. 746; see alsoMatthew,
1918, p. 575).Also, lower
jaw much
shallower than in either.Diacodon
pineyensisis appreciably larger than
Diacodon
tauri-cinerei Jepsen (1930, pp.124-126)
from
the lowerGray
Bull beds.Discussion.
— A
sufficient portion of the heel of P3 is preserved inU.S.N.AL No.
19203 toshow
that theKnight
species isDiacodon
ratherthanParictops. Moreover, absenceofa fourthdiminutivecuspon
the heel of the molars is further evidence supporting reference to Diacodon.In critically
examining
the type ofDiacodon
alticuspis, I find that the preserved portion of P4 is quite unlike this tooth inDiacodon
bicuspis,
D.
tauri-cinerei, orD.
pineyensis.The
decidedly short P4 with its narrow,much
reduced talonid inD.
alticuspis might justify resurrecting the genericname
Pcdaeictops for D. bicuspis, etc.Mat-
thew's reasons for first erecting^and
then dropping thename Pa-
laeictops are not given.
However,
the differences here noted are as greatas, if not greater than, thosebetween Parictops multicuspisand Diacodon
bicuspis. I (1949, p. 221) earlier regarded Parictops as probablyvalid,butthesignificance ofits charactersmay
beno
greater than thosewhich seem
indicated for Palaeictops. It should be noted, moreover, that in the character of P3Diacodon
tauri-cinereimakes
a distinct approachtoward
Parictops multicuspis. Itis anticipated thatnew
collections currently beingmade by
Dr.Simpson from
theSan
Jose formation inNew Mexico
will reveal additional material ofDiacodon
alticuspis so that thisform
will bemore
clearly definedand
the genus better understood.•*
Named
for the town of Big Piney, Wyo.5Granger (1910, p. 250) has incorrectly attributed this genus to Cope.
20 SMITHSONIAN MISCELLANEOUS COLLECTIONS
VOL. II7DIACODON,
cf.ALTICUSPIS
CopeThere
is in Princeton'scollection,from
about lOOfeet belowGreen
River beds, 3 miles northwest of Fossil,Wyo.,
a lowerjaw
portion, P.U.No.
16171, representing a species ofDiacodon
very close toD.
alticuspisCope.
The
speciesis clearlynotDiacodon
pineyensis.Com-
pared to the latter the cheek teeth are narrower, particularly the talonids,
and
the paraconid is placed lower with respect to themuch
elevated protoconid
and
metaconid.The
narrowness of the talonid ismost
emphasized inP4
inwhich
the entoconid ismuch subdued and
thebasin restricted.The
teeth,except for P4,are slightly largerthan inD.
tauri-cinereiand
the trigonids of the molars are higher, except for the paraconid,and
broader.Comparison
withD.
alticuspis sug- gests a close relationship, but onewhich
is inconclusiveowing
to the fragmentary nature ofthetype.MIXODECTIDAE
CYNODONTOMYS KNIGHTENSIS,e new
species Plate 2, figure iType.
—
Leftramus
ofmandible with P4-M3,U.S.N.M. No.
19314.Horizon and
locality.— Upper Knight
beds.La Barge
fauna,SW-^
sec. 33, T. 32 N., R. Ill
W.,
12 miles north of Big Piney, Sublette County,Wyo,
Description.
—
Size close to that ofCynodontomys
latidensCope
(1882a, pp. 151-152),much
smaller thanmore
nearly contemporane- ousCynodontomys
scottianus(Cope)
(i88ia, pp. 188-189).P4
with paraconid distinctand
low,and
with metaconid almost as large asprotoconidand
well separatedfrom
itthough
not somuch
so as in Microsyops. Talonid ofP4
broadand
fully molariform,though
slightly
narrower
than in molars, with protoconid, entoconid,and
hypoconulid sharpand
well defined.P4
distinctlymore
progressive than illustrated(Matthew,
1915c, figs. 45, 46) forCynodontomys
latidens Cope.Discussion.
—
In all there are 13 specimens of this form, all but oneare lowerjaws and, except for two, arefrom
thesame
exposures as thetype.One
lowerjaw and
anisolated uppermolarwere
obtained southeast of Big Piney,on
the east side of theGreen
River. This interesting species, except for its smaller size, is as fully developed asCynodontomys
scottianus in the progressiveness of its fourth lower premolar, asmight
be expected in this horizon.Cynodontomys
scot-^
Named
for the Knightformation.tianus, however, is not represented in this fauna, nor is the relatively gigantic
Cynodontomys
lundeliusiWhite
(1952, pp. 191-192)from
theLost Cabin beds intheBoysen
Reservoir area.Though
C. knight- ensisand
C. scottianus arefrom
distinctly separate basins of de- position duringupper Wasatchian
time, the geographic distance be-tween
the occurrences is not particularly great. It seems probable that the difference inspecies istobe attributedtoecologic differences.MEASUREMENTS OFTYPE LOWER
JAW
OFCynodontomys knightensis,
new
species, u.s.n.m. no. 19314 ram.Pz (alveolus) to Ms, incl 20.5
P4 to Ms, incl 15.2
Ml
to Ms, incl 11.5Depth of jaw beneath
M2
(internally) 7.$CYNODONTOMYS,
speciesA
portionofaleftlowerjaw
includingMg and
M3,inthecollections oftheAmerican Museum, from
thebluffsof theBear
Riversouth ofKnight
stationis identifiedasCynodontomys. M2
inthis specimen has thesame
proportions asin C. knightensis, butM3
is relatively alittle shorter as inCynodontomys
angustidens. This specimen,A.M. No.
12836, has been labeled
Cynodontomys
latidensf but in the absence of P4 the progressiveness of the dentition cannot be clearly deter- mined.An
unusual feature of the specimen is the remarkable flange- like development of the outward-and
forward-turned anterolateralmargin
of the ascending ramus.An
approachto this condition is seen in theprominence
of this crest in certain of the specimens of C.knightensis.
TILLODONTIA ESTHONYCHIDAE
ESTHONYX,
cf,ACUTIDENS
CopeTwo
incomplete, isolated upper molarsfrom
variegated beds be-tween
Big Pineyand La
Barge,Wyo., and
a singlelower molarfrom
12milesNorth
ofBigPiney,may
well represent thespeciesEsthonyx
acutidensCope
(1881a, pp. 185-186).The
l