Identifikasi Keragaman Gen Growth Hormone Releasing Hormone (Ghrh) Pada Kerbau Lokal (Bubalus Bubalis) Dengan Metode Pcr-Rflp
Teks penuh
Gambar


Dokumen terkait
Pemanfaatan famili gen Hormon Pertumbuhan (GH , GHR, GHRH dan PIT-1) untuk mendeteksi keragaman genetik kerbau di Kabupaten Pandeglang dan Lebak Provinsi
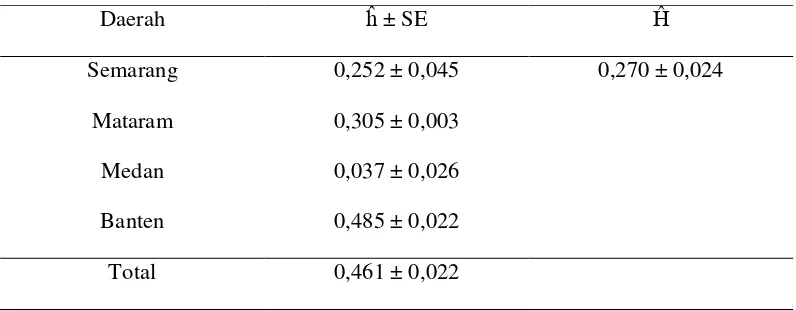
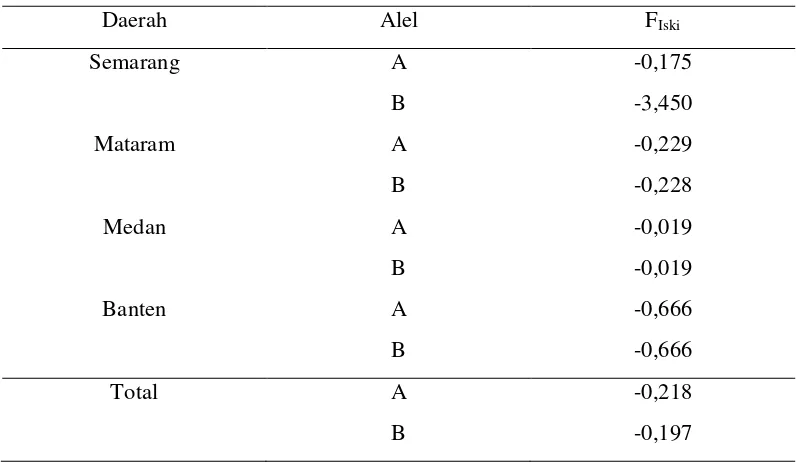
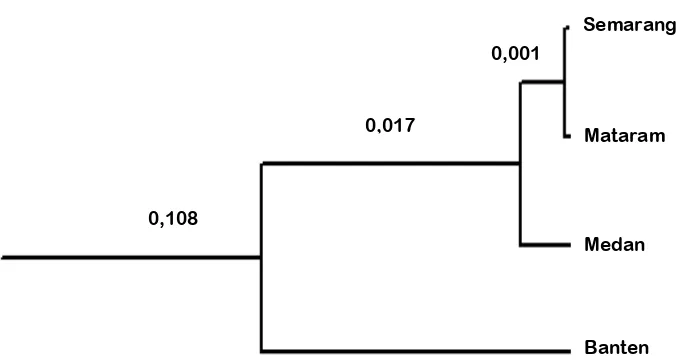
Tujuan penelitian ini adalah mempelajari dan membandingkan keragaman genetik berdasarkan daerah d-loop genom mitokondria kerbau lokal ( Bubalus bubalis ) dan mengkaji
Skripsi yang berjudul Identifikasi Keragaman Gen Follicle Stimulating Hormone Receptor (FSHR|AluI) pada Spesies Sapi Bos javanicus, Bos taurus, dan Bos indicus dengan
Penelitian ini diharapkan dapat memberikan informasi mengenai identifikasi keragaman gen FSHR pada sapi lokal Indonesia yang terdiri atas sapi Bali, Aceh, Pesisir, PO,
Tidak ditemukannya keragaman di sekuen gen FMO3 pada beberapa sampel ayam lokal dapat disebabkan karena teknik PCR-RFLP yang digunakan hanya mampu mengenali satu titik
Penelitian molekuler seperti metode PCR-SSCP untuk mengamati keragaman gen Hormon Pertumbuhan telah banyak dikembangkan untuk mengidentifikasi faktor-faktor yang
Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Desember 2015 ini ialah ayam lokal, dengan judul Identifikasi Kegaraman Gen Growth Hormone
Penelitian ini diharapkan dapat memberikan informasi mengenai identifikasi keragaman gen FSHR pada sapi lokal Indonesia yang terdiri atas sapi Bali, Aceh, Pesisir, PO,
