ALUMINIUM
RADITE TISTAMA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi berjudul”Isolasi dan Introduksi Gen Sitrat Sintase dari Pseudomonas aeruginosa ke dalam Tanaman untuk Meningkatkan Toleransi terhadap Cekaman Aluminium” adalah karya bersama saya dengan pembimbing yang belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2012
RADITE TISTAMA. Isolation and Introduction of Citrate Synthase Gene of Pseudomonas aeruginosa into Plant to Increase Aluminum Tolerance. Supervised by SUHARSONO, UTUT WIDYASTUTI, DIDY SOPANDIE
Commonly, marginal land was known as acid soil with aluminum toxicity as the main problem. Nevertheless, acid soil has the potential for expansion of agricultural lands both for food crops and energy producing plant. Our research target was to develop a biodiesel plant, Jatropha curcas, which has a good growth performance in acid soils. This research divided into four steps: 1) analysis of J. curcas response to Al stress, 2) isolation and characterization of Pseudomonas aeruginosa citrate synthase gene (PaCS), 3) expression of PaCS into model plant, Nicotiana tabacum, and 4) overexpression of PaCS gene into J. curcas. The J. curcas roots that were treated with different concentration of Al showed inhibited root elongation, which was caused by Al accumulation in the roots. Citrate synthase gene was isolated from Pseudomonas aeruginosa SP01 by using specific primer from citrate synthase gene. PaCS gene was introduced into N. tabacum and J. curcas mediated by Agrobacterium tumefaciens. Malate accumulation and malate secretion were increased in J. curcas root under Al stress. The Al-inhibited root elongation was reduced by addition of citrate in culture media. The research showed that J. curcas was less tolerant than Melastoma malabathricum, therefore it was categorized into moderate-Al tolerant. The PaCS gene was consisted of 1287 bp and has 96% similarity with citrate synthase of P. aeruginosa PA01. The N. tabacum transgenic showed more tolerance to Al than that of wild type, and histologically, the N. tabacum transgenic roots accumulated Al lower than that of wild type roots. PaCS gene was successfully introduced into J. curcas, although the transformation efficiency was only about 1%.
RADITE TISTAMA. Isolasi dan introduksi gen sitrat sintase dari Pseudomonas aeruginosa ke dalam tanaman untuk meningkatkan toleransi terhadap cekaman aluminium. Dibimbing oleh SUHARSONO, UTUT WIDYASTUTI, DIDY SOPANDIE
Luas lahan marginal di Indonesia mencapai kurang lebih 102,8 juta ha untuk lahan kering dan lahan rawa masam 34,78 juta ha. Lahan-lahan marginal ini pada umumnya adalah berupa lahan masam dengan toksisitas aluminium (Al) sebagai masalah utama. Meskipun demikian lahan marginal berpotensi untuk perluasan lahan pertanian baik untuk tanaman pangan maupun tanaman sumber energi. Salah satu tanaman sumber energi yang dapat dikembangkan di lahan marginal adalah Jatropha curcas. Tanaman ini kurang kompetitif apabila ditanam di lahan subur. Respon pertumbuhan Jatropha curcas terhadap cekaman Al belum diteliti. Salah satu mekanisme toleransi Al pada tanaman berhubungan dengan kemampuan tanaman dalam mensintesis dan mensekresikan asam organik. Asam organik yang paling kuat mengkelat Al adalah sitrat. Sitrat disintesis oleh enzim sitrat sintase. Ekspresi berlebih gen sitrat sintase di tanaman dapat meningkatkan sintesis dan sekresi sitrat serta meningkatkan toleransi terhadap Al. Gen sitrate sintase dapat diisolasi dari berbagai organisme termasuk bakteri. Tujuan utama penelitian ini adalah untuk mengintroduksi gen sitrat sintase untuk mendapatkan J. curcas lebih toleran terhadap Al. Penelitian ini dibagi menjadi empat tahapan yaitu pertama, pengujian respon J. curcas terhadap berbagai tingkat konsentrasi cekaman Al, kedua, isolasi dan karakterisasi gen sitrat sintase dari Pseudomonas aeruginosa, ketiga, introduksi gen sitrat sintase ke dalam tanaman tanaman model, Nicotiana tabacum dan keempat, introduksi gen sitrat sintase ke dalam J. curcas untuk meningkatkan toleransi terhadap Al.
Pada percobaan pertama, benih J. curcas diperlakukan dengan berbagai konsentrasi aluminium dalam medium hidroponik. Melastoma malabathricum yang diketahui toleran terhadap Al digunakan sebagai tanaman pembanding. Pertumbuhan akar bibit J. curcas yang berumur 2 minggu mulai dihambat pada cekaman 0,4 mM Al. Penghambatan akar tersebut dipicu oleh akumulasi aluminium di ujung akar yang menyebabkan kerusakan sel-sel ujung akar. Cekaman Al menyebabkan akumulasi Al di akar J. curcas dan meningkatkan sekresi malat. Penghambatan pertumbuhan J. curcas berkurang dengan penambahan sitrat pada media. Perbandingan toleransi Al J. curcas dengan M. malabathricum dan beberapa tanaman lain, maka J. curcas termasuk ke dalam tanaman yang moderat toleran Al. Penambahan sitrat ke dalam media meningkatkan pertumbuhan J. curcas yang dicekam Al.
dibandingkan tanaman tipe liarnya. Hasil pengamatan histologis menunjukkan bahwa tanaman tembakau transgenik mengakumulasi aluminium lebih rendah dan kerusakan jaringan ujung akarnya lebih sedikit dibandingkan tanaman liarnya.
Setelah berhasil diekspresikan di tanaman model, PaCS diekspresikan ke dalam J. curcas. Kotiledon berumur 2 minggu digunakan sebagai eksplan dalam proses introduksi gen PaCS diko-kultivasi dengan A. tumefaciens LB 4404 yang mengandung pMSH1-PaCS. Eksplan dinduksi kalusnya kemudian diseleksi pada media yang mengandung 20 ppm kanamisin. Tunas yang dapat tumbuh pada media seleksi disebut sebagai tunas transgenik putatif. Analisis dengn PCR menggunakan kombinasi primer CaMV 35S forward dan PaCS reverse menunjukkan bahwa dua tunas yang tumbuh di media seleksi mengandung PaCS. Efisiensi transformasi J. curcas masih sangat rendah yaitu berkisar 1%.
©Hak Cipta milik IPB, tahun 2012 Hak Cipta Dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar Institut Pertanian Bogor.
MENINGKATKAN TOLERANSI TERHADAP CEKAMAN
ALUMINIUM
RADITE TISTAMA
Disertasi
Sebagian salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Biologi Tumbuhan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup, 13 Juli 2012
1. Dr. Ir. Aris Tjahjoleksono, DEA
Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam IPB
2. Dr. Ir. Memen Surahman, M.Sc.Agr
Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB
Penguji Luar Komisi pada Ujian Terbuka, 19 Juli 2012
1. Prof. Dr. Bambang Sapto Purwoko, M.Sc
Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB
2. Prof. Akiho Yokota, PhD
Terhadap Cekaman Aluminium Nama : Radite Tistama
NIM : G363070011
Disetujui Komisi Pembimbing
Ketua
Prof. Dr. Ir. Suharsono, DEA
Dr. Ir. Utut Widyastuti, M.Si. Prof. Dr. Ir. Didy Sopandie, M. Agr
Anggota Anggota
.
Diketahui
Ketua Program Studi Biologi Tumbuhan Dekan Sekolah Pascasarjana
Dr. Ir. Miftahudin, M.Si. Dr. Ir. Dahrul Syah, M.Sc.Agr.
Alhamdulillah, segala puji hanya kepada Allah SWT atas segala rahmat dan karuniaNya sehingga penulis dapat menyelesaikan karya ilmiah ini. Disertasi ini disusun berdasarkan hasil penelitian yang dilakukan selama 3 tahun di Laboratorium Biotechnology Research Indonesia-The Netherland (BIORIN) Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) IPB dan Nara Institute of Science and Technology (NAIST), Japan. Disertasi ini memuat hasil penelitian tentang isolasi dan introduksi gen sitrate sintase dari Pseudomonas aeruginosa ke dalam tanaman untuk meningkatkan toleransi aluminium, dan selanjutnya diajukan sebagai syarat untuk memperoleh gelar Doktor pada Sekolah Pascasarjana Institut Pertanian Bogor.
Pada kesempatan ini, penulis menyampaikan penghargaan dan terima kasih kepada Prof. Dr. Ir. Suharsono, DEA, selaku ketua komisi pembimbing, Dr.Ir. Utut Widyastuti, M.S. dan Prof. Dr. Ir. Didy Sopandie, M.Agr. sebagai anggota komisi pembimbing yang telah memberikan bantuan dan bimbingan kepada penulis selama penelitian dan penyelesaian disertasi ini. Ucapan terima kasih juga penulis sampaikan pada Program Hibah Kompetensi Ditjen Dikti yang berjudul “Isolation and expression of genes in the frame of obtaining plant tolerant to acid and aluminium stresses” dan Program JSPS-DGHE Joint Research Project 2010 dengan judul “Molecular adaptation of J. curcas to acid soil for reforestation of tropical wasteland” atas nama Prof. Dr. Ir. Suharsono, DEA yang telah memberikan dukungan finansial selama penelitian
dilakukan di NAIST. Penghargaan juga penulis sampaikan kepada Prof. Akiho Yokota, PhD atas fasilitas yang diberikan selama melakukan isolasi gen sitrat sintase serta melakukan analisis fisiologi dan histologi di NAIST, serta Penelitian Kerjasama dan Publikasi Internasional dengan judul: “Genetic engineering of Jatropha curcas by genes responsible for aluminium tolerance and flowering” tahun 2012, atas nama Prof. Dr. Ir. Suharsono, DEA.
Studi Biologi Tumbuhan dan Departemen Biologi atas ilmu dan bantuan administrasi yang diberikan.
Terima kasih kepada Dr. Ir. Tjahjo Leksono, DEA, Dr. Ir. Memen Surahman M.Sc. Agr. Prof. Dr. Bambang Sapto Purwoko, M.Sc. dan Prof. Akiho Yokota, PhD selaku Penguji Luar Komisi, yang telah memberikan masukan-masukan yang berharga untuk perbaikan karya tulis ini.
Penulis juga menyampaikan terima kasih kepada rekan-rekan di Lab. Biorin, Lab. BMST dan semua staf PPSHB IPB Bogor atas segala bantuan, dukungan dan persahabatan selama penulis melakukan penelitian. Masih banyak pihak yang telah membantu kelancaran penelitian, namun tidak dapat penulis sebutkan satu persatu, untuk itu penulis juga mengucapkan terima kasih.
Ucapan terima kasih juga disampaikan kepada istriku Iyen Handayani, SH serta anak-anakku, Naufal Sasotyadena, Puspanendah Sasotyakirana dan Alika Tistama Sasotyahilmi, Ibu Suharni, (alm) Bapak Ngasirun Puspawardaya, dan bapak mertua H. Holil Burhanudin serta seluruh keluarga atas segala dukungan, kesabaran dan doanya.
Semoga karya ilmiah ini bermanfaat.
Penulis dilahirkan di Sleman pada tanggal 22 Juni 1968 sebagai anak ke-3 dari pasangan (almarhum) Ngasirun Puspawardaya dan Suharni. Pendidikan sarjana ditempuh di Fakultas Biologi UGM Yogyakarta. Pada tahun 2003 penulis menamatkan studi S2 di Program Studi Bioteknologi IPB Bogor. Pada tahun 2007 mendapatkan beasiswa dari Balit Sungei Putih Puslit Karet untuk melanjutkan studi S3 di Program Studi Biologi Program Pascasarjana IPB Bogor.
Penulis bekerja sebagai peneliti di Kelompok Penelitian Fisiologi Tumbuhan di Balai Penelitian Sungei Putih sejak 1997 hingga sekarang. Pada tahun 2005-2007, penulis ditugaskan sebagai Kepala Laboratorium Penelitian.
Halaman
Mekanisme Toksisitas Al di Jaringan akar ………... 9
Toleransi Tanaman Terhadap Cekaman Al ……….. 12
Gen-Gen yang Dipengaruhi Cekaman Al ………. 14
Sintesis Sitrat di Siklus Kreb ………... 15
Eksudasi Asam Organik ………. 17
Enzim Sitrat Sintase ……… 19
Biologi Agrobacterium ……….. 20
Peran Agrobacterium dalam Rekayasa Genetik …... 23
BAB III. RESPON FISIOLOGI Jatropha curcas TERHADAP CEKAMAN ALUMINIUM Pendahuluan ……….……….. 27
Metodologi Penelitian ……….……… 29
Hasil Dan Pembahasan ……….………. 31
Kesimpulan ……….……… 39
BAB IV. ISOLASI DAN KARAKTERISASI GEN SITRAT SINTASE Pseudomonas aeruginosa SP01 Pendahuluan ……….. ……… 40
Metodologi Penelitian ……… 41
Hasil Dan Pembahasan ……….……... 44
Kesimpulan ……….……….. 52
UNTUK PENINGKATKAN TOLERANSI TERHADAP ALUMINIUM
Pendahuluan ……….. 65
Metodologi Penelitian ……… 67
Hasil dan Pembahasan ……….. 69
Kesimpulan ……… 74
BAB VII. PEMBAHASAN UMUM ……….. 75
BAB VIII. KESIMPULAN UMUM ……….. 84
DAFTAR PUSTAKA ……….. 85
Halaman
1 Hasil analisis Blastx dari sekuen hasil amplifikasi PCR dengan
menggunakan primer spesifik gen sitrat sintase……….. 48
2
Jumlah tunas dari masing-masing asesi tanpa perlakuan Agrobacterium dan
Halaman
1. Tahapan penelitian…………...……… 7
2 Pengaruh pH terhadap bentuk spesiasi Al………...………. 8
3 Mekanisme toksisitas dan toleransi Al pada tanaman………..……… 13
4 Siklus Kreb……….. 16
5 Perubahan lintasan sintesis sitrat dan isositrat pada sel tanaman ……… 17
6 Struktur tiga dimensi sitrat sintase………... 20
7 Peta fisik plasmid T-DNA……… 21
8 Mekanisme infeksi Agrobacterium ke tanaman ……….. 23
9 Mekanisme penyisipan T-DNA ke genom sel inang………... 24
10 Pengaruh cekaman Al terhadap pemanjangan akar J. curcas……….. 32
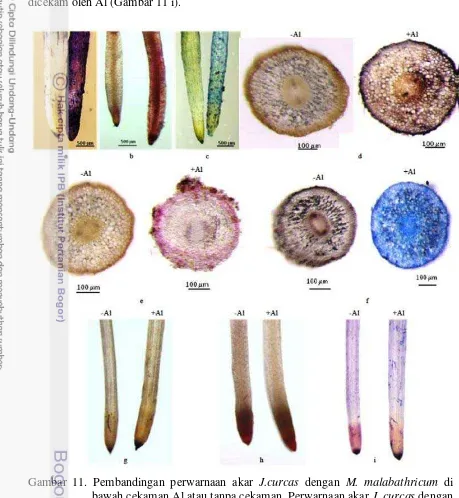
11 Pembandingan perwarnaan akar J.curcas dengan M. malabathricum ………… 34
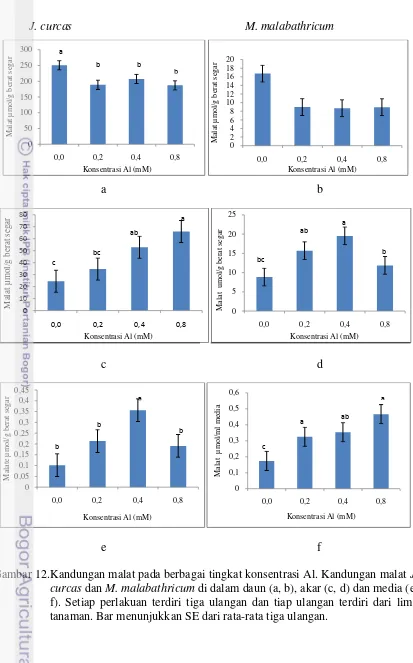
12 Pengaruh cekaman berbagai konsentrasi Al terhadap kandungan malat J. curcas dan M. malabathricum ……… 36 13 Pengaruh pemberian sitrat pada media kultur terhadap persentase berat kering J. curcas. . ………..………. 38 14 Urutan nukleotida fragmen DNA 16S P. aeuroginosa SP01. ………. 45
15 PCR koloni bakteri E. coli DH5α hasil transformasi pGEM-T easy yang mengandung gen sitrat sintase. ……….. 45 16 Deduksi asam amino dari hasil sekuen DNA. Protein PaCS terdiri dari 428 asam amino. ……… 47 17 Pohon filogenetik Pseudomonas sp berdasarkan sekuen gen sitrat sintase……. 49
18 Analisis hidrofobisitas asam-asam amino penyusun sitrat sintase P. aeruginosa SP01 ………..
50
19 Penjajaran sekuen asam amino penyusun sitrat sintase bakteri dan tanaman………..
21 Hasil PCR terhadap tembakau yang diduga transgenik……….. 60
22 Seleksi tanaman tembakau menggunakan 50 µg/ml higromisin ... 60
23 Pemanjangan akar N. tabacum tipe liar (TL) dan tanaman transgenik T1 ... 62
24 Fenotipik tanaman tembakau tipe liar (TL) dan tanaman trangenik T1 ……… 63
25 Proses introduksi PaCS ke J. curcas……….…. 70
BAB I
PENDAHULUAN
Latar Belakang
Potensi pengembangan lahan marginal untuk ekstensifikasi tanaman pangan dan tanaman komersial lainnya masih sangat besar mengingat luasan lahan marginal mencapai kurang lebih 102,8 juta ha untuk lahan kering dan lahan rawa masam 34,78 juta ha (Mulyani 2004). Luas lahan marginal di Kalimantan mencapai 30,51 juta ha atau meliputi 57,22% dari luas pulau (Suharta 2010). Pembentukan lahan masam di Indonesia disebabkan oleh adanya variasi iklim dan curah hujan yang relatif tinggi di sebagian besar wilayah Indonesia. Kondisi tersebut mengakibatkan pencucian kandungan basa dalam tanah sehingga tanah menjadi asam (Subagyo et al. 2004). Lahan-lahan marginal ini pada umumnya memiliki pH rendah yang menyebabkan meningkatnya toksisitas aluminium (Al) di dalam larutan tanah dan miskin unsur hara (Samac & Tesfaye 2003).
Ultisol merupakan salah satu jenis tanah yang mempunyai sebaran luas di Indonesia. Sebaran tanah ultisol paling luas di Kalimantan (21,9 juta ha), diikuti Sumatra (9,5 juta ha), Maluku dan Papua (8,9 juta ha), Sulawesi (4,3 juta ha), Jawa (1,2 juta ha) dan Nusa Tenggara (53 ribu ha). Sebagian besar tanah ultisol bersifat masam hingga sangat masam (pH 5,0-4,0). Kapasitas tukar kation tanah ultisol yang berasal dari granit, sedimen dan tufa tergolong rendah yaitu masing-masing berkisar antara 2,9-7,5 cmol/kg, 6,11-13,68 cmol/kg, dan 6,10-6,80 cmol/kg. Nilai kejenuhan Al tanah ultisol yang berasal dari sedimen dan granit lebih dari 60%, dan yang berasal dari tufa bagian bawah sekitar 37-78% (Prasetyo & Suradikarta 2006).
Di pH larutan yang lebih rendah dari 5, ion Al tersedia dalam bentuk Al(H2O)63+ atau Al3+, dan seiring dengan meningkatnya pH, maka Al3+ mengalami
deprotonisasi menjadi Al(OH)2+
Keracunan Al mempengaruhi pertumbuhan dan produktifitas tanaman. Gangguan yang ditimbulkan oleh keracunan Al umumnya dibagi ke dalam dua kelompok yaitu gangguan jangka pendek dan gangguan jangka panjang. Gangguan jangka pendek mulai kelihatan hanya beberapa jam saja setelah tanaman mendapat
cekaman Al seperti penghambatan pemanjangan akar, kerusakan tudung akar, pembentukan kalose, adanya deposit lignin dinding sel dan penurunan pembelahan sel. Aluminium dalam jangka panjang menyebabkan penurunan biomasa akar dan pucuk, abnormalitas morfologi akar, penurunan penyerapan dan translokasi hara, gangguan penyerapan dan transport air dan supresi fotosintesis (Miyasaka et al. 2006)
Gangguan pertumbuhan akar diawali dengan adanya akumulasi Al di ujung akar. Genotipe-genotipe yang sensitif Al mengakumulasi Al yang lebih intensif dibandingkan genotipe yang toleran (Matsumoto & Motoda 2012). Akumulasi Al di ujung akar terutama terjadi di zona transisi (ZT), yaitu zona di antara zona sel-sel yang aktif membelah dengan zona pemanjangan sel (Poschenrieder et al. 2008). Aluminium juga mempengaruhi arsitektur akar. Arsitektur akar menjadi pendek tebal dan bersisik jika tanaman mendapat cekaman Al.
Gangguan Al terhadap pertumbuhan akar meliputi perubahan proses fisiologis dan biokimia. Perubahan tersebut disebabkan afinitas Al yang tinggi terhadap protein, fosfat inorganik, fosfolipid, nukleotida, DNA, RNA dan antosian (Taylor 1991; Delhaize & Ryan 1995). Respon fisiologis dapat berupa gangguan pertumbuhan ujung akar (Samac & Tesfaye 2003), gangguan pembentukan dinding sel akar, pengambilan dan transport membran terhadap unsur esensial (Ca, Mg, K dan P), serta gangguan aktivitas beberapa enzim (Rout et al. 2001).
Salah satu tanaman model yang mengandalkan mekanisme internal adalah
Melastoma malabathricum. Tanaman ini menggunakan oksalat untuk menetralisir Al di sitosol sel-akar. Oksalat mengkelat Al dan membawanya ke xilem. Al yang dilepaskan di xilem akan dikelat oleh sitrat. Komplek sitrat-Al dibawa menuju jaringan daun karena pengaruh transpirasi. Di dalam jaringan daun Al akan dikelat oksalat dan dikurung di epidermis dan mesofil (Watanabe & Osaki 2002).
Penelitian mengenai mekanisme toleransi tiap spesies penting dilakukan terutama pada tanaman yang potensial dikembangkan di lahan masam. Spesies tanaman tersebut meliputi tanaman sebagai sumber pangan, sumber energi atau sumber biomasa lainnya. Salah satu sumber energi alternatif adalah minyak Jatropha curcas. J. curcas
mempunyai beberapa keunggulan yaitu mudah diperbanyak, masa pembentukan biji cepat, mempunyai kandungan minyak tinggi, pertumbuhannya cepat, dan mampu tumbuh di agro-klimat yang bervariasi (Divacara et al. 2010). Namun demikian, J. curcas kurang kompetitif dibandingkan dengan tanaman perkebunan lainnya jika ditanam di lahan yang subur. Tanah yang subur lebih menguntungkan untuk tanaman pangan atau perkebunan lainnya. Areal alternatif untuk pengembangan perkebunan J. curcas adalah lahan-lahan marginal seperti sebagian Pulau Sumatra dan Kalimantan. Pemahaman mengenai toksisitas aluminium dan respon J. curcas diperlukan untuk mengembangkan pemuliaan tanaman jarak ke depan dan menyusun protokol kultivasi tanaman jarak di lahan masam. Informasi yang lengkap mengenai respon terhadap cekaman Al juga diperlukan untuk melakukan rekayasa genetik yang berkaitan dengan toleransi terhadap cekaman Al.
Toleransi terhadap cekaman Al berkorelasi positif dengan aktivitas eksudasi asam organik pada beberapa spesies tanaman (Pellet et al. 1995; Ma 2000; Kochian et al. 2004; Ryan et al. 2004). Eksudasi asam organik berupa asam sitrat, malat, dan oksalat ke daerah rizosfer merupakan salah satu mekanisme toleransi tanaman terhadap Al (Ryan et al. 1995, Ma et al. 2001).
2006). Tanaman gandum (Li et al. 2000) dan Cassia tora L (Yang et al. 2004) merespon cekaman Al dengan cara meningkatkan konsentrasi dan aktivitas enzim sitrat sintase 3 jam setelah perlakuan Al. Peningkatan sintesis sitrat tersebut diikuti oleh meningkatnya efluks sitrat seiring dengan meningkatnya konsentrasi Al (Yang et al.
2004). Tanaman kedelai mengakumulasi sitrat dan malat di dalam jaringan akar. Akar tanaman kedelai mengandung sitrat lebih rendah dibandingkan malat, tetapi mensekresikan lebih banyak sitrat dibandingkan malat (Kasim et al. 2001). Contoh tanaman yang juga meningkatkan sintesis dan sekresi sitrat jika dicekam oleh Al adalah
Hordeum vulgare L (Furukawa et al. 2004) dan Triticum aestivum (Ryan et al. 2009). Sitrat mempunyai kemampuan mengkelat Al yang paling tinggi dibandingkan asam organik lainnya, diikuti oleh oksalat, malat dan suksinat (Hue et al. 1986; Ma et al.
2001). Sitrat akan membentuk komplek Al-sitrat dengan perbandingan 1:1 dan Al-malat dengan perbandingan 1:3 (Kochian 1995). Bentuk komplek ini merupakan bentuk yang tidak beracun bagi akar serta dapat meningkatkan pH dalam sel (Appanna et al. 2002). Sedangkan oksalat membentuk komplek dengan Al dengan perbandingan 1:1, 1:2 dan 1:3, dan hanya komplek Al:oksalat 1:3 saja yang tidak beracun (Ma & Hiradate 2000; Ma et al. 2001). Bentuk kompleks Al-sitrat yang diambil oleh akar akan segera dimetabolisme di dalam sel (Appanna et al. 2002).
Sitrat disintesis dalam siklus asam trikarboksilat oleh enzim sitrat sintase (EC 4.13.7) di dalam mitokondria dan peroksisomal (Ma et al. 2001; Anoop et al. 2003). Asam sitrat ini digunakan untuk respirasi asam lemak dan perkecambahan (Pracharoenwattana et al. 2005). Sebagian sitrat dikeluarkan dari mitokondria dan dieksudasikan ke dalam media yang mengandung Al (Yang et al. 2006). Eksudasi sitrat difasilitasi oleh suatu protein kinase pada kanal anion (Shen et al. 2004) dan protein transporter sitrat (Yang et al. 2006). Shen et al. (2005) menyebutkan bahwa eksudasi sitrat meningkat dengan meningkatnya aktivitas enzim H+
Teknik rekayasa genetik telah banyak digunakan para peneliti untuk memperbaiki sifat-sifat genetik suatu tanaman seperti peningkatan toleransi Al. Penelitian tersebut memerlukan koleksi gen-gen yang berkaitan dengan cekaman aluminium dan kemudian -ATPase (Li et al. 2000; Yang
menguji ekspresinya. Gen-gen dapat diisolasi dari tanaman yang diperlakukan dengan cekaman Al kemudian dibuat pustaka cDNA. Gen-gen yang diinduksi oleh cekaman Al dapat diketahui dengan cara membandingkan antara pustaka cDNA tanaman yang diperlakukan cekaman Al dengan tanaman kontrol (Anwar 1999). Teknik mutakhir yang digunakan untuk mengetahui ge-gen yang dipengaruhi Al saat ini adalah dengan menganalisis total transkrip terhadap tanaman yang dicekam oleh Al, yang dikenal analisis transkriptomik. Analisis ini menggambarkan pola ekspresi tiap-tiap gen (Kumari et al. 2008; You et al. 2011). Gen-gen yang diinduksi oleh Al termasuk gen-gen yang berkaitan dengan sintesis asam organik dapat dimanfaatkan untuk mendapatkan tanaman yang toleran.
Ekspresi berlebih gen sitrat sintase meningkatkan kandungan sitrat di dalam jaringan akar maupun yang disekresikan (Samac & Tesfaye 2003). Overproduksi sitrat pada tanaman transgenik terbukti meningkatkan toleransi tanaman tembakau dan pepaya terhadap Al (de la Fuente et al. 1997). Introduksi gen sitrat sintase ke tanaman wortel juga meningkatkan kemampuan menyerap unsur fosfor (P) dalam tanah masam (Koyama et al. 2000). Demikian pula dengan transformasi khamir dan kanola dengan gen sitrat sintase meningkatkan akumulasi sitrat dua kali lipat yang diikuti peningkatan toleransi kedua spesies tersebut terhadap Al (Anoop et al. 2003). Meskipun Delhaize et al. (2001) melaporkan tidak mendapatkan tanaman transgenik yang meningkat toleransinya terhadap cekaman Al, tetapi Baron et al. (2008) membuktikan bahwa sitrat sintase bakteri dapat meningkatkan kandungan sitrat sekaligus meningkatkan toleransi tanaman terhadap Al.
Upaya meningkatkan toleransi Al dengan meningkatkan sintesis dan sekresi sitrat pada Jatropha curcas belum diteliti. Upaya tersebut memerlukan sumber gen penyandi sitrat sintase dan diekspresikan dengan promoter kuat. Sebagai sumber gen dapat berasal dari organisme eukariotik maupun prokariotik seperti bakteri. Gen sitrat sintase telah diisolasi dari beberapa bakteri. Ukuran gen sitrat sintase termotoleran berukuran 1612 pb (Schendel et al. 1992), dan dari Pseudomonas aeruginosa berukuran 1258 pb (Donald et al. 1989).
biodegradasi bahan-bahan organik (Idise et al. 2010), sebagai agen antimikroba patogen (Chin-A-Woeng et al. 2003), untuk mengatasi cekaman oksidatif (Hassett et al. 1993), pengkelat logam (Lemire et al. 2010), dan dimanfaatkan gennya untuk meningkatkan toleransi tanaman terhadap cekaman aluminium (de la Fuente et al. 1997). Beberapa spesies dalam genus ini juga dimanfaatkan sebagai bakteri pelarut fosfat, yaitu dengan mensekresikan asam organik, terutama sitrat. P. fluorescens menunjukkan peningkatan sekresi sitrat apabila mendapat cekaman Al (Mailloux et al. 2008) dan sitrat diketahui sebagai pengkelat aluminium (Al) utama di bakteri ini (Lemire et al. 2010).
Salah satu strategi untuk meningkatkan toleransi tanaman terhadap Al adalah dengan mengurangi serapan dan kontak sel dengan Al (Poschenrieder et al. 2008). Beberapa peneliti mengintroduksikan gen sitrat sintase yang berasal dari bakteri (de la Fuente et al. 1997) ataupun dari tanaman lain (Koyama et al. 1999, 2000; Anoop et al.
2003) ke dalam tanaman target untuk mengurangi serapan dan kontak langsung sel dengan Al.
Tujuan Penelitian
Tujuan utama penelitian ini adalah untuk merakit tanaman jarak pagar Jatropha curcas yang toleran terhadap cekaman Al. Penelitian tersebut perlu didukung informasi mengenai respon pertumbuhan tanaman jarak pagar J. curcas terhadap cekaman aluminium (Al). Secara khusus penelitian ini bertujuan :
1. Menganalisis respon fisiologis dan biokimia J. curcas terhadap cekaman aluminium.
2. Mengisolasi dan mengkarakterisasi gen sitrat sintase dari bakteri Pseudomonas aeruginosa.
3. Mengintroduksikan gen sitrat sintase dari bakteri ke dalam tanaman model
Nicotiana tabacum untuk meningkatkan toleransi terhadap cekaman Al.
4. Mengintroduksikan gen sitrat sintase ke dalam tanaman biodisel J. curcas
melalui perantara Agrobacterium tumefaciens.
Manfaat Penelitian
cekaman aluminium. Penelitian ini juga bermanfaat untuk perbaikan genetik J. curcas
dalam kaitannya dengan cekaman Al. Secara khusus penelitian ini bermanfaat untuk mendapat gen sitrat sintase yang dapat digunakan untuk rekayasa genetik berbagai tanaman serta memantapkan protokol introduksi gen ke J. curcas dengan perantara
Agrobacterium tumefaciens.
Strategi Penelitian
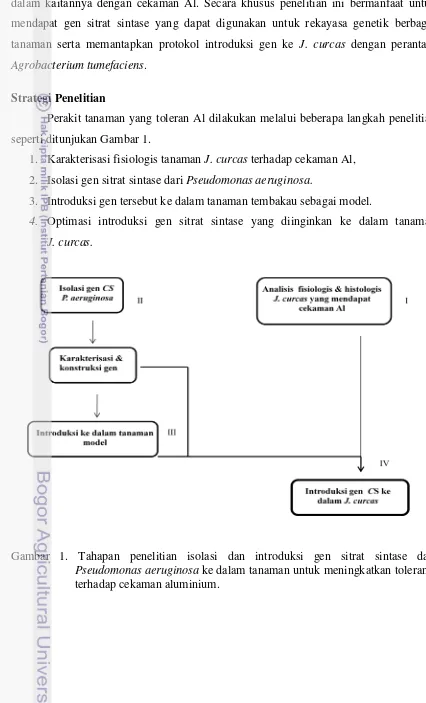
Perakit tanaman yang toleran Al dilakukan melalui beberapa langkah penelitian seperti ditunjukan Gambar 1.
1. Karakterisasi fisiologis tanaman J. curcas terhadap cekaman Al, 2. Isolasi gen sitrat sintase dari Pseudomonas aeruginosa.
3. Introduksi gen tersebut ke dalam tanaman tembakau sebagai model.
4. Optimasi introduksi gen sitrat sintase yang diinginkan ke dalam tanaman
J. curcas.
Gambar 1. Tahapan penelitian isolasi dan introduksi gen sitrat sintase dari
BAB II
TINJAUAN PUSTAKA
Toksisitas Aluminium
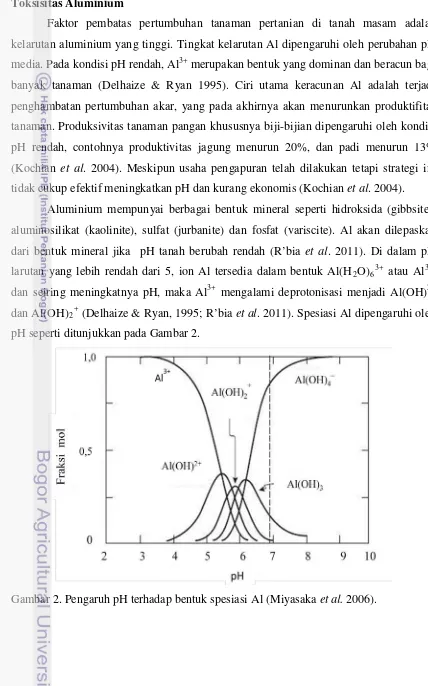
Faktor pembatas pertumbuhan tanaman pertanian di tanah masam adalah kelarutan aluminium yang tinggi. Tingkat kelarutan Al dipengaruhi oleh perubahan pH media. Pada kondisi pH rendah, Al3+
Aluminium mempunyai berbagai bentuk mineral seperti hidroksida (gibbsite), aluminosilikat (kaolinite), sulfat (jurbanite) dan fosfat (variscite). Al akan dilepaskan dari bentuk mineral jika pH tanah berubah rendah (R’bia et al. 2011). Di dalam pH larutan yang lebih rendah dari 5, ion Al tersedia dalam bentuk Al(H
merupakan bentuk yang dominan dan beracun bagi banyak tanaman (Delhaize & Ryan 1995). Ciri utama keracunan Al adalah terjadi penghambatan pertumbuhan akar, yang pada akhirnya akan menurunkan produktifitas tanaman. Produksivitas tanaman pangan khususnya biji-bijian dipengaruhi oleh kondisi pH rendah, contohnya produktivitas jagung menurun 20%, dan padi menurun 13% (Kochian et al. 2004). Meskipun usaha pengapuran telah dilakukan tetapi strategi ini tidak cukup efektif meningkatkan pH dan kurang ekonomis (Kochian et al. 2004).
2O)63+ atau Al3+,
dan seiring meningkatnya pH, maka Al3+ mengalami deprotonisasi menjadi Al(OH)2+ dan Al(OH)2+ (Delhaize & Ryan, 1995; R’bia et al. 2011). Spesiasi Al dipengaruhi oleh
pH seperti ditunjukkan pada Gambar 2.
Keberadaan Al3+ di dalam larutan tanah merngubah permukaan senyawa inorganik dan juga subtansi organik di tanah. Al3+ adalah logam yang reaktif yang dapat membentuk komplek dengan berbagai ligan organik maupun anorganik termasuk karboksilat, (Kochian et al. 2002), fosfat, sulfat, grup karboksil, asam organik, protein dan lemak (Delhaize & Ryan, 1995; Poschenrieder et al. 2008). Beberapa bentuk spesies Al mempunyai toksisitas yang rendah misalnya AlF, AlSO4 dan AlP (Miyasaka et al. 2006). Karakter reaktif Al3+
Mekanisme Toksisitas Al di Jaringan Akar Tanaman
ini juga mempengaruhi penyerapan beberapa hara penting bagi tanaman seperti P, Ca, Mg, dan K (Matsumoto & Yamaya 1988).
Fokus penelitian dalam satu dekade terakhir masih berkisar mengenai toksisitas Al di daerah apoplasma. Proses penghambatan akar yang berlangsung cepat diduga kuat karena adanya Al di apoplasma. Al3+ fitotoksik merupakan logam trivalen yang paling beracun dibandingkan logam trivalen lainnya seperti La3+, Cr3+ atau Ga3+
Gangguan Al terhadap pertumbuhan akar diawali dengan akumulasi Al di ujung akar. Akumulasi Al di akar dapat dideteksi dengan pewarnaan hematoksilin. Genotipe yang sensitif akan menunjukkan warna hematoksilin yang lebih intensif dibandingkan genotipe yang toleran (Matsumoto & Motoda, 2012). Proses pembelahan dan pemanjangan sel-sel di ujung akar dihentikan oleh keberadaan Al di jaringan akar, sehingga akar menjadi rapuh, perkembangan rambut akar menjadi berkurang dan membengkak (Panda et al. 2009). Aluminium tidak hanya berkaitan dengan berkurangnya pertumbuhan akar tetapi juga arsitektur akar. Aluminium mempengaruhi struktur akar yang menyebabkan akar menjadi pendek tebal dan bersisik. Akumulasi Al di daerah ujung akar terutama terjadi di zona transisi paling sensitif, yaitu zona antara zona sel-sel yang aktif membelah dengan zona pemanjangan sel (Poschenrieder et al. 2008).
. Toksisitas Al dapat mengganggu pertumbuhan akar meskipun dalam konsentrasi rendah dan dalam waktu beberapa jam saja. Absorbsi Al berlangsung cepat walaupun dalam jumlah kecil. Aluminium sudah dapat mencapai vakuola dalam waktu 30 menit (Miyasaka et al.
2006). Berbagai molekul menjadi target utama Al, yaitu karboksil pektin pada dinding sel, fosfolipid plasma membran, ATP dan nukleotida di bagian dalam sel (Poschenrieder
Perubahan struktur sel-sel akar di bawah cekaman aluminium meliputi reduksi jumlah butir pati dalam leukoplas, inti sel tersegmentasi dan adanya kondensasi kromosom pada inti (Nagy et al. 2004), reduksi sisterna diktiosom, dan kerusakan pada membran plasma. Di samping itu juga terjadi pembengkakan mitokondria dan reduksi jumlah cristae, serta penebalan pada dinding sel (Konarska 2008). Proses penebalan dinding sel atau lignisasi endodermis akar tanaman sensitif lebih cepat dikarenakan adanya akumulasi Al di jaringan korteks (Silva et al. 2010).
Di dalam sel, Al akan memicu akumulasi polisakarida dinding sel utamanya hemiselulosa sehingga terjadi penebalan dinding sel. Pengikatan Al terhadap material penyusun dinding sel yang baru akan menyebabkan perubahan susunan material, kekuatan dinding sel, elastisitas dan viskositas dinding sel yang baru terbentuk (Ma et al.
2004). Interaksi aluminium dengan sel-sel zona pemanjangan akar tidak hanya menghambat pemanjangan sel-sel tetapi menyebabkan penghambatan pembelahan sel yang dapat dilihat dari perubahan morfologi ujung akar. Faktor utama perubahan ini adalah gangguan pada dinding sel yang mengikat Al dan menempatkan di apoplas. Pektin metilesterase yang berperan dalam demetilasi pektin meningkat dengan adanya cekaman Al. Gangguan Al menyebabkan inisiasi akar baru di bagian distal zona pemanjangan. Inisiasi ini disebabkan adanya hambatan transport auksin ke ujung akar dengan adanya zat penghambat asam naftilptalamat (Poschenrieder et al. 2008).
Laju influk Al3+ ke dalam sitoplasma relatif rendah. Diduga Al masuk ke simplas secara pasif yang difasilitasi oleh protein tertentu yang belum diketahui. Gradien Al ini terjaga dengan pengkelatan Al yang baru saja masuk oleh sitrat asam organik. Di simplas, pH netral menyebabkan aktivitas Al3+ menjadi sangat rendah. Walaupun konsentrasi hanya nanomolar, Al sudah sangat efisien berkompetisi dengan Mg2+
Interaksi Al dengan membran plasma menyebabkan terganggunya fluiditas dan potensial membran sehingga merubah sifat permeabilitasnya (Poschenrieder et al. 2008). Selain itu, interaksi Al dengan membran plasma mengganggu sistem transport, homeostasis kalsium, transduksi signal dan penghambatan mikrotubul (Ma 2000). Aluminium menghambat efluk H
dalam mengikat ATP (Ma 2000; Poschenrieder et al. 2008).
+
Peroksidasi lipida merupakan gejala permulaan keracunan Al (Yamamoto et al.
2001). Reaksi ini dipicu oleh senyawa radikal (Reactive oxygen species/ROS) sebagai akibat cekaman Al di dalam jaringan akar (Yamamoto et al. 2002). Degradasi lipid ini mengakibatkan membran plasma kehilangan integritasnya (Yamamoto et al. 2001), yang selanjutnya memicu gangguan fungsi akar dalam penyerapan hara dan air (Mossor-Peitraszewska, 2001), sehingga menyebabkan defisiensi unsur hara. Tanaman yang keracunan Al, pada beberapa kasus daunnya menunjukkan gejala yang mirip dengan defisiensi fosfor (P), kalsium (Ca) atau besi (Fe) (Rout et al. 2001).
Proses penghambatan pertumbuhan akar oleh Al terjadi dalam waktu beberapa jam saja. Beberapa peneliti berargumen bahwa penghambatan tersebut disebabkan adanya gangguan dalam transduksi signal (Jones & Kochian 1995). Sebagai buktinya, aluminum menghambat aktivitas fosfolipase C (FLC) yang berperan melepaskan inositol 1,4,5-trifosfat ke sitoplasma dan diasil gliserol (DAG) ke membran (Jones & Kochian 1995). Selain FLC, Al juga menghambat calmodulin yang berperan mengaktifkan beberapa protein kinase (Panda et al. 2009). Transport kalsium oleh akar juga terganggu dengan keberadaan Al di dalam larutan. Sementara Ca2+ diketahui mengatur berbagai proses dalam pertumbuhan sel dan metabolisme. Target downstream
(aliran hilir) signal Ca2+ adalah kanal ion, protein kinase, dan sitoskeleton yang dikaitkan dengan pembelahan dan pertumbuhan sel (Srivastava 2002). Gangguan homeostasis Ca2+ akan mereduksi pembelahan sel dan pemanjangan akar (Panda et al. 2009). Al dapat merubah fungsi membran dengan mengikat fosfolipid. Pengikatan tersebut menginduksi terbentuknya senyawa oksigen reaktif yang menyebabkan peroksidasi lipid (Yamamoto et al. 2001). Toksisitas Al ini juga memicu perubahan potensial membran. Perubahan ini mempengaruhi penyerapan kalsium (Ca2+
Penelitian yang mengkaitkan toleransi Al dengan selektifitas membran dilakukan dengan mempelajari kanal Ca
). Kalsium di dalam sitoplasma telah diketahui mempunyai peran penting dalam pertumbuhan sel dan metabolisme (Panda et al. 2009).
2+
membuktikan tidak terdapat perbedaan antara tanaman yang toleran dibandingkan dengan tanaman sensitif dalam hal peran dari kanal Ca2+
Toleransi Tanaman Terhadap Cekaman Al
. Perbedaan toleransi antar kedua genotipe dikarenakan adanya perubahan aktivitas di rizosfer yang mereduksi toksisitas Al (Huang et al. 1995).
Mekanisme toleransi tanaman dibagi menjadi dua kelompok yaitu eksternal dan internal. Mekanisme toleransi eksternal meliputi imobilisasi Al dinding sel, selektifitas membran plasma terhadap Al, induksi pH di daerah perakaran atau apoplasma, eksudasi senyawa-senyawa pengkelat Al. Mekanisme toleransi internal meliputi pengkelatan Al di sitosol, pengurungan Al di vakuola, pengikatan Al oleh protein, akumulasi protein tertentu (Taylor 1991).
Ma et al. (2001) mengajukan dua tipe mekanisme ekslusi asam organik dalam kaitannya dengan cekaman Al. Tipe pertama, asam organik disekresikan sesaat 15-30 menit setelah Al ditambahkan di media. Hal ini telah dilaporkan oleh Delhaize et al.
(1993) pada tanaman Triticum aestivum yang mensekresikan malat dalam 15 menit setelah ditambahkan Al. Tipe kedua, sekresi asam organik ditunda beberapa jam, seperti di Casia tora di mana akumulasi sitrat di ujung akar meningkat pada 3 jam setelah dicekam oleh Al, sedangkan eksudasi terjadi pada 6 jam setelah perlakuan Al (Yang et al. 2004).
Kecepatan sekresi asam organik pada tipe pertama karena tidak memerlukan induksi protein baru, tetapi cukup mengaktifkan protein tranporter tertentu yang ada di membran (Ma et al. 2001). Sebaliknya pada tipe sekresi kedua memerlukan aktivasi gen tertentu seperti protein tranporter atau enzim-enzim yang terlibat dalam biosintesis asam organik (Kochian et al. 2004).
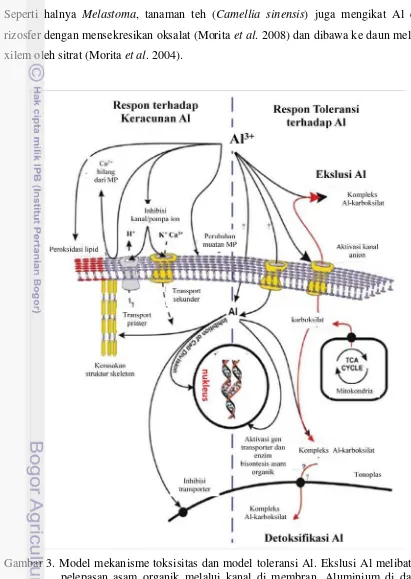
Kochian et al. (2004) mengilustrasikan peran asam organik dalam mendektosifikasi Al (Gambar 3). Mekanisme eksklusi Al melibatkan kanal yang ada pada membran plasma. Dalam mekanisme ekslusi ini ada tiga kemungkinan yaitu 1) Al langsung mengaktifkan protein kanal, 2) Al masuk ke dalam sitosol dan mengaktifkan protein kanal dan 3) Al mengaktifkan transduksi signal mengirimkan pesan untuk mengaktifkan protein kanal (Gambar 3).
akumulator Al mensekresikan asam organik terutama oksalat. Al yang terikat oksalat dibawa dan dilepaskan ke xilem. Di jaringan ini Al diikat oleh sitrat dan dibawa ke daun, dan di daun Al diikat oleh oksalat untuk ditimbun di vakuola atau sitosol epidermis. Seperti halnya Melastoma, tanaman teh (Camellia sinensis) juga mengikat Al dari rizosfer dengan mensekresikan oksalat (Morita et al. 2008) dan dibawa ke daun melalui xilem oleh sitrat (Morita et al. 2004).
Gambar 3. Model mekanisme toksisitas dan model toleransi Al. Ekslusi Al melibatkan pelepasan asam organik melalui kanal di membran. Aluminium di dalam sitosol dikelat oleh asam organik dan dikurung di vakuola (Kochian et al.
Sitrat diketahui sebagai asam organik yang paling kuat dalam mengikat Al. Al terikat dengan sitrat dengan perbandingan molekul 1:1, Al-oksalat 1:2, dan dengan malat 1:3 (Ma dan Hiradate 2000). Setiap spesies memiliki jenis asam organik yang disekresikan yang berbeda-beda. Tanaman yang meningkatkan sintesis dan sekresi sitrat adalah seperti Casia tora (Yang et al. 2004), Hordeum vulgare L (Furukawa et al.
2004) dan Triticum aestivum (Ryan et al. 2009).
Gen-Gen yang Ekspresinya Dipengaruhi Cekaman Al
Karakterisasi gen-gen yang ekspresinya diinduksi Al dilakukan dengan cara membuat pustaka cDNA tanaman yang dicekam oleh Al, kemudian dibandingkan dengan pustaka cDNA tanaman yang tidak dicekam oleh Al (Anwar 1999). Gen-gen yang diinduksi oleh cekaman Al telah diisolasi dari tanaman gandum dan Arabidopsis, yaitu glutation-S-transferase (GST), blue copper binding protein (BCB), superoxide dismutase (SOD), katalase dan reticuline oxygen oxidoreductase (Snowden & Gardner 1993; Richards et al. 1998). Ezaki et al. (1995) juga berhasil mengisolasi tiga gen dari tembakau yang ekspresinya diinduksi cekaman Al yaitu GST, peroksidase (PER) dan
GDP dissociation proteinase inhibitor (DGI). Gen-gen yang diisolasi tersebut lebih banyak berkaitan dengan reduksi senyawa-senyawa radikal. Anwar (1999) telah mengisolasi gen-gen yang diinduksi Al yang berkaitan dengan perkembangan sel atau sistem transport seperti protein histon H3, H+
Karakterisasi mekanisme seluler di bawah cekaman Al saat ini dilakukan dalam skala besar yaitu dengan analisis transkriptomik dan proteomik. Analisis transkriptomik terhadap Arabidopsis yang dicekam oleh Al selama 48 jam menunjukkan perubahan ekspresi gen-gen yang berkaitan dengan jalur stres oksidatif, dinding sel, dan metabolisme polisakarida. Enzim-enzim yang terlibat dalam siklus Kreb tidak dipengaruhi oleh cekaman Al, kecuali malat dehidrogenase (Kumari et al. 2008). You et al. (2011) melaporkan bahwa cekaman Al pada kedelai selama 4 jam meningkatkan ekspresi 561 gen dan menurunkan ekspresi 78 gen. Hampir separuh gen-gen yang responsif Al belum diketahui fungsinya. Gen-gen yang diketahui meningkat ekspresinya adalah faktor ekspresi, famili transporter MATE, STOP dan deposit lignin (You et al. 2011).
Analisis proteomik terhadap kedelai yang toleran Al diketahui bahwa protein yang meningkat jumlahnya adalah heat shock protein, glutathione S-transferase,
chalcone-related synthetase, GTP-binding protein and ABC transporter ATP-binding protein (Zhen et al. 2007). Beberapa protein yang berhubungan dengan ROS meningkat dengan adanya Al yaitu superokside dismutase, dan mondehydroascorbate reductase yang berperan dalam sintesis asam askorbat. Selain itu, dua protein diasosiasikan dengan sintesis prolin juga meningkat oleh paparan Al yaitu arginin tRNA ligase dan glutamate dehidrogenase (Zhou et al. 2009). Duressa et al. (2010) juga melakukan analisis proteomik terhadap genotipe kedelai sensitif dibandingkan dengan proteomik yang toleran Al. Setelah 72 jam diperlakukan dengan Al, protein-protein yang meningkat konsentrasinya adalah m alat dehidrogenase, enolase, malat oksidoreduktase, dan piruvat dehidrogenase. Peningkatan protein-protein tersebut hanya terjadi pada genotipe toleran saja.
Sintesis Sitrat di dalam Siklus Kreb
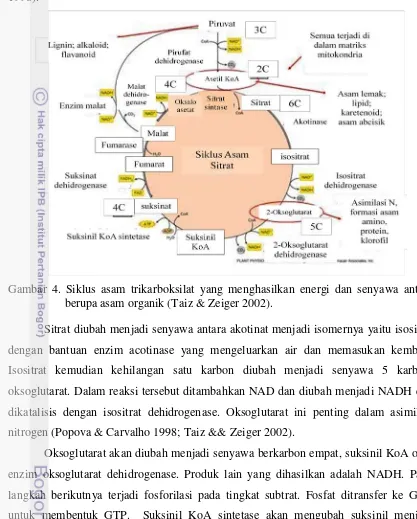
Asam sitrat dihasilkan dari siklus asam trikarboksilat (TCA) yang dikenal dengan Siklus Kreb atau siklus asam sitrat. Siklus ini merupakan pusat jalur metabolik untuk semua proses aerobik dalam kehidupan organisme. Selama siklus TCA unit C2 asetil KoA yang merupakan derivat karbohidrat dan lipida diproses menjadi karbon dioksida dan air. Dalam siklus ini dihasilkan senyawa antara, NADH dan FADH2 sebagai energi reduktif. Meskipun siklus TCA terjadi di mitokondria namun produk antara diakumulasi di vakuola dan energi reduktif masuk ke rantai transport elektron untuk proses fosforilasi di bagian dalam membran mitokondria. Siklus TCA diawali dengan pengubahan pirufat menjadi asetil KoA dengan melepaskan CO2
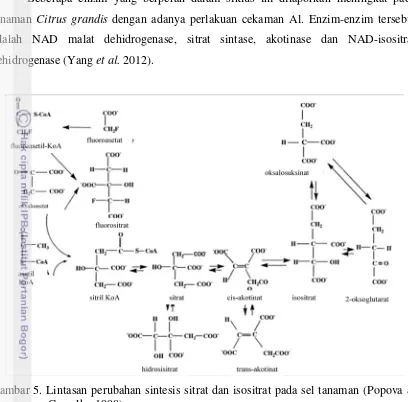
Selama kondensasi oksaloasetat dengan asetil KoA, asetil dari kelompok asetil KoA ditambahkan ke mioti keto oksaloasetat. Sitril asetat adalah senyawa antara dalam reaksi ini (Gambar 5). Sitrat bukan hanya sebagai metabolit dalam siklus TCA saja, tetapi bisa juga berperan menjadi donor asetil oleh ATP-sitrat liase. ATP-sitrat liase
mengkatalisis sitrat dan KoA menjadi oksaloasetat dan asetil KoA (Popova & Carvalho 1998).
Gambar 4. Siklus asam trikarboksilat yang menghasilkan energi dan senyawa antara berupa asam organik (Taiz & Zeiger 2002).
Sitrat diubah menjadi senyawa antara akotinat menjadi isomernya yaitu isositrat dengan bantuan enzim acotinase yang mengeluarkan air dan memasukan kembali. Isositrat kemudian kehilangan satu karbon diubah menjadi senyawa 5 karbon, oksoglutarat. Dalam reaksi tersebut ditambahkan NAD dan diubah menjadi NADH dan dikatalisis dengan isositrat dehidrogenase. Oksoglutarat ini penting dalam asimilasi nitrogen (Popova & Carvalho 1998; Taiz && Zeiger 2002).
Beberapa enzim yang berperan dalam siklus ini dilaporkan meningkat pada tanaman Citrus grandis dengan adanya perlakuan cekaman Al. Enzim-enzim tersebut adalah NAD malat dehidrogenase, sitrat sintase, akotinase dan NAD-isositrat dehidrogenase (Yang et al. 2012).
Gambar 5. Lintasan perubahan sintesis sitrat dan isositrat pada sel tanaman (Popova & Carvalho 1998).
Eksudasi Asam Organik
Asam organik dieksudasikan ke rizosfer dengan bantuan protein transporter. Sasaki et al. (2004) mengidentifikasi pertama kali transpoter malat yang diinduksi Al yang disebut ALMT1. Kanal ALMT1 ini merupakan faktor kunci eksudasi malat pada akar tanaman gandum yang toleran. Penelitian sebelumnya menyebutkan bahwa transporter malat diaktifkan oleh sebuah protein kinase yang berukuran 42 kDa (Osawa & Matsumoto 2001). Ekspresi berlebih gen ALMT1 ke tanaman tembakau dan barley diketahui meningkatkan toleransi terhadap cekaman Al seiring meningkatnya eksudasi malat (Sasaki et al. 2004; Delhaize et al. 2004).
Asam organik lain seperti sitrat juga mempunyai transporter sendiri yaitu
dan menyandikan 555 asam amino (Furukawa et al 2007). Ekspresi berlebih MATE
pada tanaman tembakau meningkatkan toleransi tanaman tembakau terhadap cekaman Al (Furukawa et al. 2007). Selain MATE, sitrat juga dieksudasikan melalui H+ATPase yang berkorelasi dengan efluk H+ (Ohno et al. 2003; Tomasi et al. 2009). Gen penyandi H+ATPase telah diisolasi dari Melastoma malabathricum. Gen tersebut berukuran 2871 pb yang menyandikan 956 asam amino. Pembungkaman terhadap ekspresi gen tersebut menurunkan toleransi tanaman Melastoma terhadap cekaman Al (Muzuni 2010). Mutasi H+
Transporter lain yang juga berperan dalam eksudasi asam organik adalah
transporter ABC yaitu transporter yang mengikat ATP selama mengalirkan larutan
organik maupun anorganik. Baik tanaman yang toleran maupun yang sensitif
mempunyai transporter asam organik, namun yang membedakan antara keduanya
adalah terekspresi lebih tinggi atau lebih aktif pada tanaman toleran (Kochian et al
2004). Salah satu famili gen transporter ABC adalah gen for sensitive Al rhizotoxic
(STAR) yang telah diketahui berperan dalam toleransi terhadap Al (Huang et al. 2010). ATPase meningkatkan sekresi sitrat 20 kali lipat dibandingkan tipe liarnya (Ohno et al. 2003). Penghambatan transporter sitrat dengan anthracene-9-carboxylic acid sebuah inhibitor kanal anion, menurunkan eksudasi sitrat (Yang et al. 2006).
Gen Succinate Fumarate Carrier (SFC) menyandikan protein carrier yang tersusun dari 320 asam amino yang berperan untuk transport suksinat dan fumarat keluar dari mitokondria. Aktivitas transporter suksinat ada di dalam sitosol (Linka & Weber 2010). Pada bakteri juga ditemukan transporter suksinat yang aktivitasnya optimal pada pH 5,6 dan dihambat oleh monokarboksilat (Terpe et al. 1999). Transporter oksalate juga sudah diisolasi dan dianalisis struktur tiga dimensi. Transporter ini berukuran 44 KDa disandikan oleh OxT. Protein ini mempunyai 12 heliks dan 11 heliks diantaranya melintasi membran sel (Hirai et al. 2002). Belum ada penelitian yang menguji respon transporter-transporter tersebut terhadap cekaman Al.
Brassica napus (Ligaba et al. 2004) dan dua spesies jeruk (Yang et al. 2011) jika dicekam oleh Al.
Enzim Sitrat Sintase
Sintesis sitrat diyakini sebagai tahapan pertama dalam Siklus Kreb dengan mengkondensasi asetil KoA dengan oksaloasetat. Pada organisme eukariotik, enzim sitrat sintase ditemukan terutama di dalam mitokondria dan peroksisom pada perkecambahan (Beeckmans 1984). Sitrat sintase juga ditemukan di dalam peroksisom non glioksomal di umbi kentang. Berbeda dengan organel glioksisom, organel non glioksisom tidak mengandung isositrat liase dan malat dihidrogenase (Papke & Gerhard 1996). Berat molekul sitrat sintase mitokondria lebih besar dibandingkan sitrat sintase glioksisom dan masing-masing terdiri dari dua subunit (Beeckmans 1984). Saat ini telah diketahui lima gen yang menyandi sitrat sintase pada Arabidosis thaliana. Dua dari gen tersebut ditargetkan ke dalam mitokondria, sedangkan tiga gen lainnya ditargetkan ke peroksisom. Ketiga sitrat sintase tersebut berperan baik dalam respirasi asam lemak maupun pada saat perkecambahan (Pracharoenwattana et al. 2005). Aktivitas sitrat sintase terdeteksi di peroksisomal hipokotil mung bean dan mesokarp tanaman avokad. Aktivitas sitrat sintase peroksisom lebih rendah daripada enzim sitrat sintase mitokondria (Papke & Gerhardt 1996).
Gambar 6. Struktur kuarterner sitrat sintase bakteri dalam kondisi tertutup (kiri), dan kondisi terbuka (kanan) (Protein Data Bank, 2007).
Biologi Agrobacterium
Agrobacterium adalah bakteri tanah yang termasuk ke dalam famili Rhizobiaceae. Bakteri ini bersifat pathogen bagi tanaman terutama tanaman dikotil dan beberapa spesies Gymnosperm (Srivastava 2002). Interaksi antara
tanaman-Agrobaterium secara alami hanya terjadi di dalam Kelas Dikotil. Secara alami monokotil bukan termasuk tanaman inang bagi Agrobacterium tetapi bagian meristem dapat diinfeksi bakteri ini (Smith & Hood 1995). Saat ini dikenal tiga biotipe yaitu
Agrobacterium tumefaciens yang menyebabkan crown gall, A. rubi yang menginfeksi
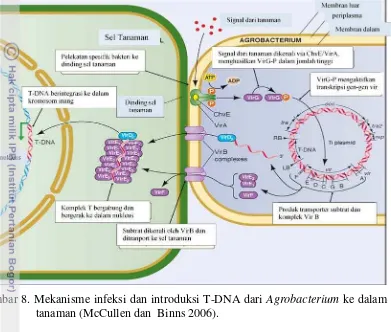
Rubus idaeus dan A. rhizogenesis yang menginfeksi jaringan akar (Otten et al. 2008). Kemampuan Agrobacterium dalam menginduksi tumor adalah karena adanya peningkatan aktivitas sitokinin dan indol acetic acid (IAA). Kedua hormon tersebut diperuntukkan menghasilkan opin sebagai sumber makanan (McMullen & Binns 2006). Pada proses ini, DNA dari Agrobacterium tipe liar ditransfer ke nukleus. Ekspresi T-DNA di dalam jaringan tanaman menyebabkan terbentuknya tumor. Gen-gen di dalam T-DNA terlibat dalam sintesis hormon pertumbuhan dan produksi opin yang merupakan kondensasi antara asam amino dan gula (Sheng & Citovsky 1996)
Ada tiga komponen genetik Agrobacterium yang diperlukan untuk transformasi sel tanaman. Komponen pertama adalah T-DNA yang diperlukan untuk mentransfer gen dari sel bakteri ke dalam sel tanaman. T-DNA terdapat dalam Ti plasmid
Kemampuan Agrobacterium menginfeksi sel inang berkaitan dengan plasmid yang berukuran besar sebagai penginduksi tumor atau tumor-inducing (Ti) atau penginduksi akar atau rhizogenic inducting (Ri). T-DNA dan daerah virulensi (vir) terdapat di tumor-inducing plasmid (pTi) (Gambar 7). Selain di plasmid T-DNA juga bisa terdapat di dalam kromosom dengan cara rekombinasi dengan bantuan protein VirD2 (Gelvin 2008). Ukuran plasmis Ti berkisar 200-800 Kpb dan daerah T kurang dari 10% dari Ti (Gelvin 2003). Ekspresi gen vir diinduksi oleh eksudat yang dikeluarkan dari luka tanaman. Eksudat tersebut berupa senyawa fenolik seperti asetosiringon yang diperkuat komponen gula dan pH (Zupan et al 2003).
Di samping gen-gen vir yang ada di plasmid, beberapa gen virulensi di kromosom juga diperlukan untuk menginfeksi se-sel inang. Gen-gen virulensi tersebut adalah chv A, chvB, chvD, chvE, chvG, chvH, dan chvI, serta gen-gen yang mempengaruhi virulensi di kromosom yaitu acvB dan pgm(exoC), glgP, mia dan ros. Urutan DNA virulensi di dalam kromosom dan plasmid Ti identik (Suzuki et al. 2001).
Gambar 7. Peta plasmid Ti yang terdiri dari T-DNA, daerah virulensi, katabolisme opin dan daerah origin of replication (Hooykas & Beijersbergen 1994).
Ada dua proses yang saling independen dalam proses pengenalan ini yaitu aktifasi gen virulen dan pengikatan sel inang (McMullen & Binns 2006). Pengenalan terhadap sel inang melalui senyawa kimia yang dikeluarkan sel-sel inang yang terluka. Senyawa kimia ini akan dikenali oleh komplek reseptor VirA/VirG yang ada di membrane plasma Agrobacterium. Senyawa opin dikenali oleh gen virA, yang kemudian akan ditransduksikan melalui VirG untuk mengaktifkan salinan gen yang akan ditranfer ke sel inang. Mekanisme ini merupakan karakteristik bakteri konjugatif (Zupan et al. 1996). Bakteri mengikat permukaan sel dan diikuti dengan sintesis filamen selulosa untuk memperkuat pengikatan (Zupan et al. 1996).
Gen virA dan virG mempunyai homologi dengan gen yang menyandikan dua sensor kinase untuk menerima respon dan melanjutkannya dalam status fosforilisasi. VirG dalam kondisi tidak aktif menempel pada protein VirA sebagai penerima signal. Jika terdapat signal berupa opin, gula, PO4
Bersama dengan aktivasi gen vir, Agrobacterium melekatkan diri di sel inang. Terdapat dua tipe pelekatan Agrobacterium ke sel inang yaitu: pelekaran nonspesifik dan spesifik. Pelekatan nonspesifik dapat diketahui dengan cara pencucian menggunakan bufer garam. Jumlah bakteri yang melekat di satu sel inang berkisar 200-1000 bakteri. Pelekatan spesifik terjadi dengan terbentuknya T-pilus yang tersusun dari
β1-2 glukan. Gen-gen di kromosom chvA, chvB dan pscA (exoC) (McMullen &Binns 2006), serta virB di Ti plasmid (Judd et al. 2005) terlibat dalam pembentuk sintesis dan lokalisasi pilus.
dan pH rendah opin maka VirG menjadi bentuk yang terfosforilisasi dan terlepas dari VirA. Pemberian opin termasuk didalamnya oktopin, nopalin, leucinopin, dan sucinamopin meningkatkan induksi 2-10 kali. Asetosiringon dapat mengaktifkan gen virA dan virG (Veluthambi et al. 1989). Asetosiringon dan ADP-glucosa meningkatkan efesiensi transformasi pada embrio muda tanaman gandum (Cheng et al. 1997). VirG akan mengaktifkan operon gen vir
yang berperan mengatur transport T-DNA ke dalam genom tanaman (Zupan et al. 2000).
dengan VirE2 untuk menghindari pemotongan nuklease (Citovsky et al. 1989). VirD1 kemudian dilepaskan dari potongan utas tunggal T-DNA dan virD2 tetap melekat di utas tunggal T-DNA (Zupan et al. 2000).
Gambar 8. Mekanisme infeksi dan introduksi T-DNA dari Agrobacterium ke dalam sel tanaman (McCullen dan Binns 2006).
T-DNA ditransfer ke sel inang melalui protein kanal yang disandikan oleh gen
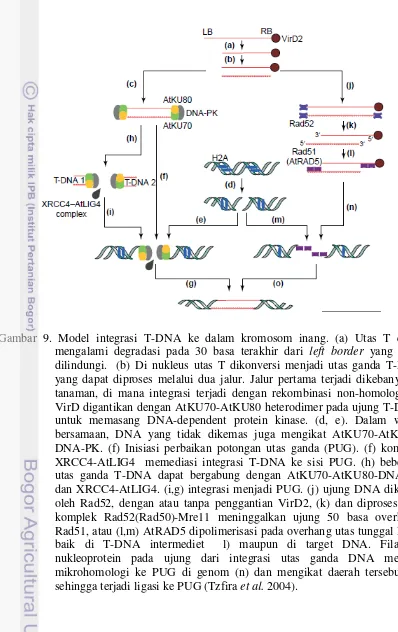
virB dan virD4 (Christie & Vogel 2000). Protein VirD2 tetap berikatan dengan T-DNA dan menjadi faktor penting dalam membawa T-DNA. Mutasi di daerah terminal C VirD menyebabkan kegagalan transfer (van Kregten et al. 2009) Integrasi T-DNA ke dalam kromosom inang diilustrasikan seperti pada Gambar 9 (Tzfira et al. 2004).
Peran Agrobacterium dalam Rekayasa Genetik
target memiliki keunggulan dibandingkan dengan metode lain yaitu lebih ekonomis dan jumlah kopi transgen yang terintegrasi adalah rendah (Paz et al. 2004).
Infeksi Agrobacterium dipengaruhi beberapa faktor yaitu mulai pengenalan terhadap inang hingga integrasi gen target ke dalam genom inang (Zupan et al. 2000). Cahaya mempengaruhi kemampuan Agrobacterium dalam menginfeksi sel inang. Dalam kondisi ada cahaya kemampuan bakteri tersebut menginfeksi tanaman berkurang dikarenakan penurunan jumlah flagela. Penurunan pembentukan flagela disebabkan adanya penghambatan operon flaABC oleh cahaya sehingga kemampuan pembentuk flagelin menurun (Oberpichler et al. 2008). Agrobacterium mampu tumbuh optimum baik di kondisi pH netral, asam dan sedikit basa. Patogenitas bakteri tersebut meningkat pada kondisi asam (Li et al. 2003).
Untuk keperluan uji ekspresi, gen target harus disisipkan di T-DNA di antara promoter dan terminator. Selain gen target, T-DNA juga harus mengandung gen untuk menyeleksi sel-sel transgenik dari sel-sel non transgenik. Gen penyeleksi tersebut dapat berupa gen resistensi terhadap antibiotik (Chee et al. 1989), atau gen resistensi terhadap herbisida (Fang et al. 2004). Sebagai gen pelapor ekspresi dapat digunakan gen β
glucuronidase (GUS) (Batra & Kumar 2003) atau green fluorescent protein (GFP) (Yancheva et al. 2006).
Beberapa peneliti telah melakukan introduksi gen untuk meningkatkan toleransi tanaman terhadap cekaman Al dengan bantuan Agrobacterium. Ezaki et al. (2000) mengintroduksikan beberapa gen glutation S-transferase, peroksidase, inhibitor disosiasion GDP dan blue copper -binding protein dengan bantuan A. tumefaciens LB 4404. Beberapa galur trangenik dilaporkan mampu mengurangi stres Al dan beberapa logam berat. Ekspresi gen super oxyde dismutase (MnSOD) mitokondria dari Triticum aestivum di dalam Brassica napus mampu meningkatkan toleransi tanaman tersebut terhadap cekaman Al. Enzim ini berperan mengontrol konsentrasi 1O2 dan H2O2
Ekspresi gen yang berkaitan dengan asam organik dan sekresinya mampu meningkatkan toleransi tanaman terhadap aluminium. Introduksi gen ALMT, suatu transporter malat yang difasilitasi oleh Agrobacterium juga berhasil mendapatkan galur tanaman sorgum transgenik. Sekresi malat dari akar transgen tersebut meningkat beberapa kali lipat yang sekaligus meningkatnya toleransinya terhadap cekaman Al (Delhaize et al. 2004). Ekspresi gen multidrug and toxic compound (MATE) di dalam
BAB III
RESPON FISIOLOGI Jatropha curcas TERHADAP CEKAMAN ALUMINIUM
Pendahuluan
Kebutuhan energi yang semakin meningkat seiiring meningkatnya jumlah penduduk dunia dan ditambah dengan eskalasi harga minyak bumi dunia tidak menentu, menyebabkan pencarian sumber energi alternatif (Raguskas et al. 2006). Minyak
Jatropha curcas merupakan sumber alternatif untuk pengganti minyak bumi. Tanaman ini mempunyai beberapa keunggulan yaitu mudah diperbanyak, masa pembentukan biji cepat, mempunyai kandungan minyak tinggi, pertumbuhan cepat, dan mampu tumbuh di agro-klimat yang bervariasi (Divacara et al. 2010). Meskipun potensi tanaman ini cukup menjanjikan, namun J. curcas kurang kompetitif jika ditanam di areal yang subur. Tanah yang subur lebih menguntungkan untuk tanaman pangan atau perkebunan. Areal alternatif untuk pengembangan perkebunan J. curcas adalah lahan-lahan marginal seperti sebagian Pulau Sumatra dan Kalimantan. Lahan-lahan marginal umumnya bersifat masam dengan keracunan aluminium (Al) sebagai masalah utama.
Keracunan Al dapat menghambat pertumbuhan akar pada berbagai spesies tanaman (Nagy et al. 2004), mereduksi biomasa akar dan tunas (Poscherieder et al. 2008), dan dalam jangka panjang akan mereduksi hasil panen. Pisum sativum yang dicekam oleh Al menunjukkan bermacam-macam respon seperti penghambatan pertumbuhan akar, akumulasi Al, peroksidasi lipid, dan kehilangan integritas membran plasma (Yamamoto et al. 2001). Reactive oxygen species (ROS) menjadi faktor utama yang menyebabkan penghambatan pertumbuhan akar yang dicekam oleh Al (Yin et al. 2010).
Peroksidasi lipid dapat dideteksi dengan pewarnaan dengan Schiff’s reagent (Yamamoto
et al. 2001), yang mendeteksi adanya aldehid derifat peroksida hasil degradasi lipid oleh ROS (Yin et al. 2010). Gejala keracunan berikutnya adalah sel-sel akan kehilangan integritas membran plasma sehingga pewarna Evan’s blue dapat berdifusi ke dalam sel (Yamamoto et al. 2001;Tamas et al. 2006).
Asam organik dapat mengkelat Al dan berperan penting dalam mendetoksifikasi Al, baik eksternal maupun internal sel (Ma et al. 2000). Eksudasi asam organik seperti sitrat, malat dan oksalat ke area rizosfer telah diajukan sebagai mekanisme toleransi pada beberapa tanaman dalam menghindari Al (Ryan et al. 1995). Aluminium menstimulasi eksudasi sitrat, malat dan suksinat dari jaringan ujung akar, dan sitrat merupakan asam organik yang paling banyak dieksudasikan (Osawa & Kojima 2006). Asam organik di rizosfer dapat mengkelat Al di luar membran plasma, sehingga mencegah Al masuk ke dalam jaringan akar (Miyasaka et al. 1991). Secara in vitro, asam organik dapat mereduksi toksisitas Al, di mana kemampuan sitrat mereduksi lebih efektif dibandingkan malat dan suksinat (Hue et al. 1986). Oryza sativa varietas toleran dilaporkan mempunyai aktivitas sitrat sintasenya yang lebih tinggi dibandingkan varietas peka (Kang & Ishii 2006). Selain mampu mereduksi toksisitas Al, sitrat juga bisa melarutkan fosfor dari bentuk Al-fosfat menjadi bentuk yang tersedia bagi akar tanaman (Miyasaka et al. 1991). Varietas gandum (Triticum aestivum L.) yang toleran menghasilkan malat empat kali lipat dibandingkan varietas yang peka setelah 2 jam
dicekam dengan 200 μM Al (Delhaize et al. 1993).
Ekspresi berlebih gen yang terlibat metabolisme asam organik seperti sitrat sintase ke dalam tanaman tembakau dan pepaya (de la Fuente et al. 1997) dan tanaman wortel (Koyama et al. 1999) dapat meningkatkan sintesis dan eksudasi sitrat sekaligus meningkatkan toleransi Al. Ekspresi gen ALMT1 sebuah transporter malat, terbukti meningkatkan eksudasi malat dan toleransinya terhadap cekaman Al (Delhaize et al. 2004).
Metodologi Penelitian
Penelitian ini menggunakan kecambah J. curcas varietas IP-3P. Biji disterilisasi dengan 5% Na hipoklorid ditambah beberapa tetes Tween-20 selama 10 menit lalu dibilas dengan air. Biji direndam dengan air selama 1 jam lalu dibenamkan di dalam media isolite selama 10 hari. Biji yang mempunyai vigor tinggi dipilih untuk uji cekaman Al menggunakan sistem kultur hidroponik. Media yang digunakan untuk cekaman Al adalah ½ konsentrasi media Murashige & Skoog (MS) (Murashige & Skoog 1962). Biji Melastoma malabathricum dikecambahkan pada media tanah selama 1 bulan. Kecambah tersebut digunakan sebagai pembanding spesies yang toleran Al dengan mekanisme internal detoksifikasi. Kecambah M. malabathricum ditranfer ke dalam media hidroponik pH 5,8 selama 1 minggu sebelum diperlakukan dengan Al.
Penghambatan Pertumbuhan Akar
Kecambah berumur 10 hari diperlakukan dengan berbagai tingkat konsentrasi Al yaitu 0, 0,2 mM, 0,4 mM dan 0,8 mM AlCl3
Akumulasi Al di dalam Akar
. Aluminium tersebut ditambahkan ke dalam ½ konsentrasi media MS dan pH ditetapkan pada 4,0. Cekaman dilakukan selama 7 hari, dan selama perlakuan cekaman kultur diberi aerasi. Media cekaman diganti dengan media baru setiap 2 hari sekali. Tiap perlakuan terdiri dari lima tanaman dan diulang lima kali. Semua panjang akar diukur dengan penggaris sebelum dan sesudah perlakuan cekaman Al. Melastoma digunakan sebagai pembanding dalam percobaan ini dan mendapatkan perlakuan cekaman yang sama dengan tanaman J. curcas.
Percobaan untuk pengamatan histologis dan fisiologis dilakukan dengan menggunakan dua konsentrasi Al yaitu tanpa Al dan 0,8 mM AlCl3 selama 7 hari di
dalam kultur hidroponik. Setelah perlakuan cekaman Al berakhir, akar di rendam dengan akuades selama 1 jam, lalu direndam dengan pewarna hematoksilin (0,2% (w/v) hematoksilin dalam 0,02% (w/v) KIO3) selama 10 menit. Akar dibilas dengan aquades
Degradasi Membran Plasma
Degradasi membran plasma oleh ROS dideteksi dengan menggunakan reagen Schiff’s (Yamamoto et al. 2001). Akar direndam dalam reagen tersebut selama 10 menit dan dibilas dengan 0,5% (w/v) K2S2O5
Gangguan Integritas Membran Plasma
di dalam 0,05 M HCl hingga intensitas warnanya berkurang.
Akar direndam di dalam 10 ml larutan Evan’s blue (0,025% (w/v) Evan’s blue
dalam 100 mM CaCl2 pH 5,6) selama 10 menit. Untuk mengurangi intensitas warna
biru, akar dibilas tiga kali dengan larutan 100 mM CaCl2
Kandungan Malat di dalam Akar, Daun dan Media
. Pengamatan intensitas warna akar utuh maupun irisan akar dilakukan dengan menggunakan mikroskop cahaya (Tamas et al. 2006).
Pengukuran kandungan malat dilakukan berdasarkan metode yang digunakan oleh Delhaize et al. (1993). Kecambah J. curcas dicekam dengan berbagai tingkat konsentrasi Al, yaitu 0, 0,2, 0,4 dan 0,8 mM. Aluminium tersebut ditambahkan ke dalam ½ konsentrasi media MS, dan diatur pada pH 4,0. Tiap-tiap unit percobaan diulang tiga kali, dan setiap ulangan terdiri dari lima tanaman. Percobaan dilakukan selama 6 hari. Media kultur diganti setiap dua hari sekali dan selama percobaan kultur diberi aerasi. Akar dan daun yang telah terbuka penuh dicuci dengan dengan aquades, lalu dikeringkan dengan kertas saring. Akar sepanjang 1 cm dari ujung dipotong menggunakan gunting. Ujung akar dan daun pucuk dibekukan di dalam nitrogen cair lalu disimpan di lemari pendingan pada suhu -80OC sampai diekstraksi. Sampel jaringan seberat 0,3 g digerus di dalam mortal dengan menambahkan N2 cair hingga halus.
Ekstrak tersebut ditambah dengan 0,6 N asam perklorat lalu digerus kembali. Ekstrak dipindahkan ke dalam tabung 1,5 ml, kemudian disentrifugasi dengan kecepatan 15000 rpm (Tomy MX-300) selama 5 menit. Sebanyak 0,9 ml supernatan diambil lalu ditempatkan ke dalam ke tabung baru dan dinetralkan dengan 80 μl K2CO3 (69 g/100
Malat diukur dengan cara mencampurkan 0,25 ml sampel dengan 0,75 ml buffer (0,5 M glisin, 0,4 M hidrasin, pH 9,0), dan 0,05 ml 4 mM NAD. Reaksi dipreinkubasikan terlebih dahulu selama 30 menit hingga mendapatkan nilai yang stabil pada pembacaan A340, sebelum ditambahkan 5 μl malate dehidrogenase. Peningkatan
absorban pada A340
Pengaruh Sitrat terhadap Berat Kering Bibit J. curcas.
dihitung sebagai produksi NADH yang sebanding dengan kandungan malat di dalam sampel. Analisis malat juga dilakukan pada Melastoma malabathricum yang diperlakukan sama dengan J. curcas.
Kecambah berumur 10 hari diperlakukan dengan 1 mM Al dan diperlakukan dengan berbagai konsentrasi sitrat yaitu 0, 0,4 mM, 0,8 dan 1 mM. Media yang digunakan mengikuti komposisi media Watanabe et al. (2005). Setiap perlakuan terdiri dari lima tanaman dan diulang tiga kali. Percobaan dilakukan selama 30 hari. Setiap 6 hari, media diganti dengan yang baru. Berat basah masing-masing individu ditimbang sebelum dan sesudah perlakuan. Pada akhir perlakuan, tanaman dikeringkan dengan cara dioven 60O
Hasil dan Pembahasan
C selama 12 jam sebelum ditimbang.
Penghambatan Pemanjangan Akar
Pertumbuhan akar utama J. curcas berkurang setelah tujuh hari dicekam Al. Pada tingkat cekaman 0,2 mM Al, pemanjangan akar J. curcas belum nyata terhambat jika dibandingkan dengan akar yang tidak dicekam Al. Penurunan pemanjangan akar mulai terjadi pada konsentrasi cekaman 0,4 mM dan 0,8 mM Al, berturut-turut 36,17% dan 48,02% (Gambar 10 a). Akar M. malabathricum tidak terhambat pemanjangannya di semua tingkat cekaman Al. Pemanjangan akar M. malabathricum justru meningkat pada tingkat cekaman Al 0,4 mM Al, dan tidak terjadi reduksi pemanjangan akar pada cekaman 0,8 mM Al (Gambar 10 b). Aluminium membantu penyerapan Ca dan Mg oleh akar Melastoma. Akar Melastoma mensekresi oksalat secara terus menerus sehingga dapat menyebabkan presipitasi Ca dan Mg. Jika di dalam media terdapat Al maka oksalat akan mengikat Al, sementara Ca dan Mg dalam bentuk terlarut dan dapat diserap oleh akar Melastoma (Watanabe et al. 2008).
et al. 2004), dan Leucaena leucocephala (Osawa & Kojima 2006) pemanjangan akarnya menurun drastis meskipun hanya dicekam Al konsentrasi rendah. Penurunan pemanjangan akar kedua tanaman tersebut mencapai 40% saat dicekam 0,01 mM Al. Sebuah penelitian melaporkan bahwa hanya dengan menggunakan kurang dari 0,2 mM Al dapat untuk membedakan suatu tanaman peka dan toleran (Silva et al. 2010).
a b
Gambar 10. Panjang akar J. curcas (a) dan M. malabathricum (b) yang diperlakukan dengan berbagai tingkat konsentrasi Al.
Pemanjangan akar Picea abies yang toleran Al menurun 40% pada cekaman 0,5 mM Al selama 2 hari (Nagy et al. 2004). Akar Eucaliptus sp baru mengalami penurunan yang nyata jika dicekam dengan konsentrasi Al di atas 0,6 mM (Silva et al. 2004). Beberapa varietas kedelai telah dievaluasi dengan tingkat cekaman 0,7 mM Al. Pemanjangan akar varietas peka menurun 46,5%, sedangkan pada varietas toleran, pemanjangan akar hanya berkurang 19,6% (Sopandie et al. 2003). Tanaman jarak mengalami penghambatan akar hingga 48% jika diperlakukan dengan 0,8 mM Al. Penghambatan tersebut lebih tinggi dibandingkan penghambatan akar pada M. malabathricum, Eucaliptus dan varites kedelai toleran Al. Data tersebut mendukung bahwa J. curcas dapat dikategorikan sebagai tanaman agak toleran.
Akumulasi Aluminium di dalam Akar
Akar J. curcas mengakumulasi warna hematoksilin secara intensif terutama di sepanjang 1 cm dari jaringan ujung akar yang dicekam oleh Al (Gambar 11 a). Irisan melintang di sepanjang jaringan tersebut juga menunjukkan adanya warna hematoksilin di dalam sel-sel epidermis hingga mencapai endodermis (Gambar 11 d). Tanaman tanpa perlakuan Al tidak menunjukkan adanya jaringan akar yang terwarnai oleh hematoksilin. Ini menunjukkan bahwa Al yang diberikan di media diserap dan diakumulasi oleh akar