SELEKSI MENGGUNAKAN PCR BERDASARKAN MARKA
GEN
badh2
PADA PEMBENTUKAN BC2F1
CIHERANG/MENTIK WANGI DAN BC3F1
CIHERANG/PANDAN WANGI
TAUFIQ
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
TAUFIQ. Seleksi Menggunakan PCR Berdasarkan Marka Gen badh2 pada
Pembentukan BC2F1 Ciherang/Mentik Wangi dan BC3F1 Ciherang/Pandan
Wangi. Dibimbing oleh DJAROT SASONGKO HAMISENO dan TRI JOKO
SANTOSO.
ABSTRACT
TAUFIQ. PCR Selection Based on badh2 Gene Marker in the Formation of
BC2F1 Ciherang/Mentik Wangi and BC3F1 Ciherang/Pandan Wangi. Under the
direction of DJAROT SASONGKO HAMISENO and TRI JOKO SANTOSO.
SELEKSI MENGGUNAKAN PCR BERDASARKAN MARKA
GEN
badh2
PADA PEMBENTUKAN BC2F1
CIHERANG/MENTIK WANGI DAN BC3F1
CIHERANG/PANDAN WANGI
TAUFIQ
Skripsi
Sebagai Salah Satu Syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi
: Seleksi Menggunakan PCR Berdasarkan Marka Gen badh2 pada
Pembentukan BC2F1 Ciherang/Mentik Wangi dan BC3F1
Ciherang/Pandan
Wangi.
Nama :
Taufiq
NRP :
G84063560
Disetujui
Komisi Pembimbing
Dr. Djarot Sasongko Hamiseno, MS.
Dr. Tri Joko Santoso, M.Si
Ketua
Anggota
Diketahui
Dr.Ir. I. Made Artika M.App.,Sc
Ketua Departemen Biokimia
PRAKATA
Alhamdulillah, segala puji bagi Allah yang telah berkehendak atas segala
sesuatu yang terjadi di alam semesta sehingga laporan penelitian ini dapat
terselesaikan. Penelitian ini dilaksanakan sejak April 2010 sampai dengan
Oktober 2010 di Laboratorium Biologi Molekuler dan Rumah Kaca Balai Besar
Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian
(BB-BIOGEN) dengan materi Seleksi Menggunakan PCR Berbantuan Marka Gen
badh2 pada Pembentukan BC2F1 Ciherang/Mentik Wangi dan BC3F1
Ciherang/Pandan Wangi.
Penulis mengucapkan terima kasih yang sebesar-besarnya kepada semua pihak
terutama Dr. Djarot Sasongko Hami Seno, MS. dan Dr. Tri Joko Santoso, M.Si
yang telah membimbing penulis selama penyusunan laporan penelitian. Terima
kasih yang sebesar-besarnya juga penulis ucapkan kepada teknisi-teknisi
BB-BIOGEN khususnya Pak Asep dan Pak Iman yang telah banyak membantu
penulis selama penelitian. Tak lupa penulis ucapkan terima kasih kepada keluarga
di rumah yang selalu mendukung dan mendoakan juga kepada Dewi Praptiwi,
Ganti Swara Pratama, Sugihartati, Euis Marlina, dan Joel Rivandi Sinaga selaku
rekan kerja yang telah banyak membantu penelitian ini. Juga kepada teman-teman
Biokimia 43 yang selalu memberi saran dan dukungan untuk kelancaran laporan
penelitian ini.
Akhir kata bahwa tak ada sesuatu yang sempurna, begitu pun laporan
penelitian ini. Oleh karena itu penulis mengharapkan kritik dan saran yang
membangun untuk perbaikan di masa yang akan datang. Semoga laporan
penelitian ini dapat digunakan dengan sebaik-baiknya.
Bogor, Maret 2011
RIWAYAT HIDUP
Penulis dilahirkan di Makkah, Arab Saudi pada tanggal 26 Juli 1987 dari
ayahanda Wardoyo dan ibunda Hartati. Penulis merupakan anak ke dua dari tiga
bersaudara. Tahun 2005 penulis lulus dari SMU Sekolah Indonesia Jeddah dan
pada tahun berikutnya lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui
jalur Undangan Seleksi Masuk IPB (USMI) pada Program Studi Biokimia,
Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA).
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
TINJAUAN PUSTAKA
Deskripsi Tanaman Padi ... 2
Aroma Padi ... 3
Gen Aroma ... 4
Metode Persilangan Terarah (Site-directed Crossing) ... 4
Pengembangan Varietas Padi Aromatik ... 5
Metode Analisis Aroma ... 5
BAHAN DAN METODE
Bahan dan Alat ... 6
Metode Penelitian ... 6
HASIL DAN PEMBAHASAN
Pembentukan Populasi BC2F1 CM dan BC3F1 CP ... 7
Pengujian Kualitatif dan Kuantitatif DNA ... 8
Seleksi Tanaman Padi BC2F1 CM Menggunakan Marka Bradbury ... 9
Seleksi Tanaman Padi BC3F1 CP Menggunakan Marka RM223 ... 10
Pengamatan Karakter Tanaman Padi BC2F1 CM dan BC3F1 CP ... 11
SIMPULAN DAN SARAN
Simpulan ... 12
Saran ... 12
DAFTAR PUSTAKA ... 12
DAFTAR GAMBAR
Halaman
1 Ciherang, Pandan Wangi, dan Mentik Wangi ... 2
2 Jalur pembentukan 2-asetil-1-pirolin ... 3
3 Perbandingan sekuen DNA alel badh2 pada berbagai varietas padi ... 4
4 Diagram metode persilangan terarah (site-directed crossing) ... 5
5 Pembentukan populasi tanaman padi BC2F1 CM maupun BC3F1 CP ... 8
6 Proses amplifikasi dengan marka Bradbury ... 9
7 Amplifikasi DNA padi BC2F1 CM menggunakan marka Bradbury ... 10
8 Proses amplifikasi dengan marka RM223... 10
9 Amplifikasi DNA padi BC3F1 CP menggunakan marka RM223 ... 10
10 Gabah padi yang berbulu pada tanaman BC3F1 CP ... 11
DAFTAR LAMPIRAN
Halaman
1 Diagram alir penelitian ... 16
2 Isolasi DNA dengan metode CTAB ... 17
3 Komposisi bufer ekstraksi DNA ... 18
4 Pengukuran konsentrasi, kemurnian, dan perhitungan pengenceran DNA ... 19
5 Contoh hasil elektroforesis produk PCR ... 21
PENDAHULUAN
Berdasarkan aromanya, padi dapat dibedakan menjadi padi aromatik dan nonaromatik. Umumnya padi nonaromatik lebih tahan terhadap penyakit dan stres, tidak terlalu selektif dalam pemilihan lokasi dan kondisi lahan, penanaman dan pemeliharaan lebih mudah, waktu tanam lebih singkat, dan produktivitasnya lebih tinggi. Namun demikian, rasa dan aroma yang merupakan satu-satunya kelebihan padi aromatik mempunyai aspek komersial yang tinggi. Selain nilai jualnya yang lebih tinggi, permintaan pasar nasional maupun internasional akan padi aromatik sangatlah tinggi dan meningkat terus, terutama pada masyarakat dengan taraf ekonomi yang baik.
Krisnamurthi (2006) menyebutkan bahwa setiap peningkatan pendapatan per kapita Rp 1.000,- menyebabkan kenaikan permintaan beras aromatik sebesar 3.570 kg, sedangkan setiap kenaikan jumlah penduduk 1.000 orang menyebabkan kenaikan permintaan beras nonaromatik sebesar 2.140 kg. Oleh karena itu, dalam rangka ketahanan pangan akan sangat prospektif jika aroma dapat ditambahkan pada padi nonaromatik dengan tanpa merusak kelebihan-kelebihan padi tetua pemulih (host) seperti yang telah disebutkan. Hal ini akan membuat petani mendapatkan produk beras aromatik dengan kemudahan, waktu, resiko dan produktivitas seperti menanam padi nonaromatik.
Hasil penelitian mendapatkan aroma padi disebabkan karena mutasi pada gen badh2
(Bradbury et al. 2005a,b; Borguis et al. 2008). Oleh karena itu karakter aroma dapat diintrogresi (dimasukkan) pada padi nonaromatik melalui inaktivasi gen badh2nya. Inaktivasi dapat dilakukan dengan berbagai metode rekayasa genetik (Wanchana et al. 2004, Vanavichit et al. 2008). Namun metode-metode ini akan menghasilkan produk varietas tanaman transgenik yang pemasarannya terhambat dengan regulasi GMO (Genetically Modified Organisms) yang ketat. Di lain pihak, persilangan acak (random crossing) akan menghasilkan turunan dengan karakter yang sulit diramalkan. Aromatisasi nontransgenik padi nonaromatik dapat dilakukan melalui penggantian alel gen badh2
padi nonaromatik dengan alel gen terkait dari varietas aromatik, menggunakan metode persilangan terarah (site-directed crossing) untuk mendapatkan turunan homozigot resesif (Hami Seno et al. 2009).
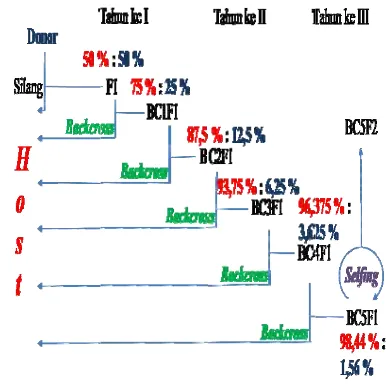
Penelitian ini merupakan bagian dari usaha pengembangan varietas aromatik baru nontransgenik yang karakter agronominya sebaik padi nonaromatik (BC5F2), menggunakan host Ciherang dan donor aroma Mentik Wangi atau Pandan Wangi. Tanaman BC1F1 Ciherang/Mentik Wangi (CM) dan BC2F1 Ciherang/Pandan Wangi (CP) yang diperoleh dari penelitian sebelumnya (Sugihartati 2009) disilangbalikkan (backcross) lebih lanjut dengan Ciherang sehingga menghasilkan BC2F1 CM dan BC3F1 CP. Seleksi dilakukan mengunakan PCR berbantuan marka Bradbury (Bradbury et al. 2005b) untuk CM dan RM223 (Lang dan Buu 2008) untuk CP. Pemilihan Ciherang karena merupakan varietas pengembangan lokal yang akrab dengan petani maupun konsumen, tahan hama dan penyakit, kondisi lahan budidaya tidak memerlukan prasyarat yang ketat sehingga budidaya dapat dilakukan di berbagai lokasi di Indonesia dengan produktivitas yang tinggi (hampir 8 ton/ha) (Krisnamurthi 2006).
Mentik Wangi dan Pandan Wangi juga merupakan varietas lokal yang juga telah akrab dengan petani dan konsumen serta aroma digemari masyarakat Indonesia, bahkan Internasional. Namun demikian, kurang tahan hama dan penyakit, lokasi dan kondisi lahan budidaya spesifik, waktu panen lebih lama, dan produktivitas kurang tinggi (hampir 4 ton/ha) (Krisnamurthi 2006). Diharapkan jika varietas aromatik tersebut disilangkan dengan varietas Ciherang, disilangbalikkan lima kali, kemudian diselfing, seperti pada metode persilangan terarah (site-directed crossing) (Hami Seno et al. 2009), akan diperoleh varietas Ciherang aromatik (BC5F2). Varietas baru dengan karakter agronomi sebaik Ciherang seperti yang telah disebutkan, serta beraroma seperti Mentik Wangi dan Pandan Wangi. Karakter positif dari varietas baru tersebut diharapkan dapat meningkatkan ekonomi petani dan sekaligus devisa negara jika diekspor, serta lebih menggairahkan minat pertanian, baik petani maupun industri.
dengan resiko kegagalan panen, kemudahan tanam, produktivitasnya seperti menanam padi Ciherang sehingga berimplikasi padi aromatik menjadi murah.
TINJAUAN PUSTAKA
Deskripsi Tanaman Padi
Tanaman padi (Oryza sativa L.) diklasifikasikan dalam divisi Spermatophyta, subdivisi Angiospermae, kelas Monocotyledone, ordo Poales/Glumiflorae, famili Graminae, genus Oryza, dan spesies
Oryza sativa (Siregar 1981). Padi yang termasuk ke dalam genus Oryza, Oryza sativa
L. merupakan tanaman padi yang banyak dibudidayakan di Asia, sedangkan Oryza glaberrima dibudidayakan di Afrika.
Berdasarkan aromanya, padi dikelompokkan atas padi aromatik dan nonaromatik. Varietas padi aromatik diantaranya yaitu Sintanur, Gilirang, Pulu Mandoti, Pare Bau, Gunung Perak, Pinjan, Celebes, Pandan Wangi, Pare Kembang, Rojo Lele, Cianjur, Mentik Wangi, dan Situ Patenggang. Padi aromatik merupakan bagian kecil yang istimewa dari kelompok padi karena memiliki mutu beras yang baik, yaitu beraroma dan tekstur nasi yang pulen, sangat disukai konsumen dan mempunyai harga yang tinggi (Bradbury et al. 2005b; Lang dan Buu 2008). Namun padi aromatik memiliki memiliki karakter agronomi yang kurang tahan terhadap hama dan penyakit, memerlukan kondisi lahan budidaya tertentu, waktu panen lama, dan produktivitas yang kurang tinggi (±4 ton/ha).
Padi Pandan Wangi (Gambar 1a) yang terkenal sejak tahun 1973 memiliki aroma wangi menyerupai pandan yang merupakan karakter paling menarik dari padi ini yang banyak diminati di pasaran dunia. Pandan Wangi mengandung kadar amilosa 7 – 20% dan amilopektin 80 – 93% yang menyebabkan padi varietas ini bertekstur pulen. Selain itu cocok tumbuh di suhu 25 – 30 °C dengan ketinggian 500 – 800 meter dari permukaan laut. Padi Mentik Wangi memiliki ciri bulat dan tahan rontok (Gambar 1b). Usia tanamnya mencapai 150 – 160 hari dan tingginya mencapai 150 cm. Kelemahan varietas ini diantaranya tingkat produktivitas rendah serta tidak tahan terhadap hama dan penyakit.
Varietas nonaromatik contohnya adalah Ciherang, IR64, Nipponbare, T309, Fatmawati, Situ Bagendit, Andel Rojo, Andel Lombok (beras merah) untuk tepung, dan
Andel Lombok (beras merah) untuk konsumsi langsung. Ciherang merupakan kelompok padi nonaromatik varietas unggul hasil beberapa kali persilangan, yaitu IR18349-53-1-3-1-3/IR19661-131-3-1//IR119661-131-3-1///IR64 ////IR64 (Bhattacharjee 2002). Padi Ciherang memiliki karakteristik umur tanamannya cukup singkat yaitu 116 hingga 125 hari, bentuk tanaman tegak, tingginya mencapai 107 sampai 115 cm, menghasilkan anakan produktif 14 sampai 17 batang, warna kaki hijau, warna batang hijau, warna daun hijau, muka daun kasar pada sebelah bawah, posisi daun tegak, bentuk gabah panjang ramping, warna gabah kuning bersih, tekstur nasi pulen, kadar amilosa 23%, bobot 1000 per butir 27 hingga 28 g, rata-rata produksi 5 - 8,5 ton/ha, tahan terhadap bakteri hawar daun (HDB) strain III dan IV, tahan terhadap wereng coklat biotipe 2 dan 3 (Gambar 1c). Padi Ciherang dilepas oleh menteri pertanian pada tahun 2000 dengan anjuran cocok ditanam pada musim hujan dan kemarau dengan ketinggian di bawah 500 m di atas permukaan laut (Hermanto 2006).
Introgresi karakter aroma dari padi varietas Pandan Wangi atau Mentik Wangi ke varietas Ciherang diharapkan dapat menghasilkan varietas dengan karakter agronomi yang hampir sama dengan karakter tanaman pemulih Ciherang (tahan hama dan penyakit, dapat dibudidayakan di berbagai lokasi di Indonesia, waktu panen yang tidak lama, dan produktivitasnya tinggi) ditambah dengan karakter aroma dari donor Pandan Wangi atau Mentik Wangi.
[a] [b]
[c]
Aroma Padi
Aroma padi disebabkan oleh senyawa kimia yang mudah menguap. Hasil penelitian menunjukkan terdapat lebih dari 114 senyawa terdapat pada padi aromatik. Namun, senyawa utama yang menyebabkan aroma wangi pada padi adalah 2-asetil-1-pirolin (2AP) (Itani 2004). Akumulasi dari 2AP dalam genotip padi aromatik dapat disebabkan oleh adanya mutasi delesi pada ekson 7 di kromosom nomor 8 yang mengakibatkan kodon stop sehingga menyebabkan hilangnya aktivitas enzim betain aldehida dehidrogenase (BADH2) ketika prolina mensintesis asam amino glutamat. Enzim BADH2 memainkan peranan kunci dalam jalur konversi ke arah glutamat. Penghambatan lintasan ini akan meningkatkan ketersediaan prolina untuk sintesis 2AP (Bradbury et al. 2005a,b). Berbeda dengan padi nonaromatik, pada kromosom nomor 8 tidak terjadi delesi ekson 7 sehingga prolina lebih mengarah ke pembentukan asam amino glutamat dan pembentukan 2AP lebih sedikit.
Padi aromatik mengandung senyawa 2AP lebih tinggi (0,04–0,07 ppm) dibandingkan padi nonaromatik (0.004–0.006 ppm) (Adijono et al. 1993). Hal yang sama dikemukakan oleh Buttery et al. (1983) bahwa kandungan senyawa 2AP padi aromatik mencapai 0.04–0.09 ppm, sepuluh kali jauh lebih tinggi dibanding padi nonaromatik yang hanya 0.004–0.006 ppm. Senyawa 2AP sebenarnya terdeteksi di semua bagian tanaman padi, kecuali di akar (Lorieux et al.
1996).
Biosintesis jalur reaksi yang rinci tentang kompleks ini belum sepenuhnya dijelaskan (Bradbury et al. 2005a). Namun, telah ditemukan bahwa prekursor dan sumber nitrogen 2AP pada varietas aromatik Thai Hom Mali adalah senyawa prolina (Yoshihashi et al. 2002). Diduga jalur pembentukan 2AP seperti pada gambar 2. Jalur pembentukan 2AP dimulai dari pemecahan prolina menjadi putresin kemudian membentuk senyawa gama aminobutiraldehid (GABald), substrat dari enzim BADH2. Jika enzim BADH2 aktif, maka enzim ini dapat mengubah GABald menjadi asam gama-aminobutirat (GABA). Tetapi jika enzim BADH2 tidak aktif, GABald mengalami asetilasi (penambahan gugus asetil) membentuk 2AP. Putresin akan ditemukan dalam jumlah tinggi pada jaringan yang tumbuh aktif membelah. Putresin dipecah menjadi GABald oleh diamina oksidase (DAO) selama proses pembentukan
lignin dan dinding sel, setelah sebagian besar pembelahan sel telah terjadi. Oleh karena itu, pembentukan GABald cenderung terjadi di jaringan muda yang secara aktif membelah dan dinding sel menjadi kaku.
Ujung daun padi mengandung 2AP dalam jumlah yang lebih tinggi daripada di pangkal daun, sementara daun muda lebih beraroma daripada daun tua (Lorieux et al. 1996). Penurunan jumlah 2AP dalam jaringan yang lebih tua kemungkinan besar disebabkan oleh sifat volatil kimia. Pada tanaman, prolina berfungsi sebagai zat pelindung terhadap resiko kerusakan daun ketika tanaman terpapar oleh cekaman kekeringan atau terdapatnya kandungan garam yang tinggi di dalam tanah (Heldt 2005).
Aroma padi aromatik tidak hanya tercium pada saat pemasakan nasi. Seringkali aroma dapat tercium saat tanaman padi berbunga di lahan (Mittal et al. 1995).
HO O
HN
HO O
NH2
NH2
H2N
NH2 H2N
H O
N (Z)
N (E)
O
H2N
OH O
HO OH
O O
NH2 Prolina
Ornitin
Putresin GABald
Pirolin
2-asetil-1-pirolin
GABA
Asam glutamat BADH2 aktif
BADH2 inaktif
Mutasi g tanaman pa oleh peristi evolusi dala secara gene gen badh2
mewakili su dan padi Ba
badh2 iden dari subke KDM105 d dan Azucen delesi 8 ba (Nipponbare (kotak mera mendukung mutasi badh
mengalami m menghasilka (Bourgis et a
Studi ge (Ahn et al. gen fgr (f
karakter ar aromatik tur Hasil penel lebih memp karakter aro resesif tungg karakter aro seperti Jasm merupakan BADH2. Ha gen yang m polimorfism genotip tan kromosom n dalam men terdapat pad padi nonarom
Gambar 3 P
ba
(B
Gen Aro
gen badh2 pa di di berbaga iwa domestik am suatu pop etik. Studi p pada 81 ubkelompok i asmati menunj ntik pada var elompok yan dari indica, Ba na dari tropi asa, sedangka e dan 93-11) ah pada Gamb
teori tentan
h2 pada padi mutasi dan do an polimorfi
al. 2008). enetik awal
1992) berhas (fragance) y roma pada v
runan Jasmine litian Bradbu persempit wi oma dan meng
gal yang berta oma pada v mine dan Ba
gen yang asil penelitian mengkode enzi me di daera naman. Gen nomor 8 yang nyebabkan ar
da spesies pa matik (Bradbu
erbandingan
adh2 pada be Bourgis et al.
oma
ada berbagai ai negara dise kasi atau ter pulasi yang te perbandingan varietas pad indica, japoni jukkan bahwa rietas-varietas ng berbeda, asmati dari G ical japonica an padi nona ) tidak terjad bar 2). Hal in ng asal tungg
budidaya As omestikasi yan isme yang b
kelompok T il melokalisir yang mengen
varietas Della e) pada kromo ury et al. (2
ilayah genom gidentifikasi s anggung jawa varietas-varieta asmati. Gen
menyandi menunjukkan im BADH2 m ah pengkode
badh2 disan g bertanggung roma. Gen adi aromatik m ury et al. 2005
sekuen DN erbagai variet 2008). varietas ebabkan rjadinya erisolasi sekuen di yang ica, aus, a mutasi s aroma , yaitu Grup V, terjadi aromatik di delesi i sangat gal dari sia yang ng akan berbeda Tanksley r sebuah ndalikan a (padi osom 8. 2005a,b) m untuk satu gen ab untuk as padi tersebut enzim n bahwa memiliki e pada ndi oleh g jawab badh2 maupun 5 a,b). NA alel tas padi Metode Per
Metode p juga site-dire
2009)sering
assisted bac backcrossing
metode ini, diminimalisas diinginkan y induk. Pemin nonaromatik varietas aro dilakukan den Padi nonarom aromatik se disilangbalikk nonaromatik. diulangi lima Silang balik yang dapat satu atau beb tanaman don Namun, diper kali agar d diinginkan d introgresi ne diinginkan (R Gen bad
selalu dalam gen ini han resesif dan dideteksi den digunakan m untuk melaca padi aromati Wangi pada generasi silan molekuler PC harus selalu silang balik, s menjadi dua 2007).
Persilanga persilangan y penggabunga atau selektif gen tunggal, puluhan tahu aman (bukan kepastian per dapat dikara pengerjaanny dari persilan hanya dapat atau genus d (Nasution 200 silangan Ter Crossing persilangan te ected crossing digunakan de
ckcrossing a (Mackill et
pemasukkan si dan ha yang terintrog ndahan gen pa untuk men omatik pad ngan metode matik disilan lanjutnya in kan dengan Proses sila a kali sampa merupakan digunakan un berapa gen ya nor ke tanam
rlukan persilan diperoleh intr dan sekecil m egatif fragme Reyes 2000).
dh2 pada gen keadaan het nya terekspre
keberadaan ngan uji arom marka moleku ak keberadaan
ik Pandan W a individu ng balik. Tan CR, penyerbuk
dilakukan pa sehingga jang
kali lebih la
an terarah m yang memili an sifat tanam dengan ketep memiliki sta un, menghasi n organisme rubahan gene akterisasi den ya 2 hingga 3 ngan terarah digunakan d dan melibatk 02).
arah (Site-dir
g)
erarah atau d
g (Hami Seno engan nama m
atau PCR-as al. 2007). M gen donor anya sifat gresi pada tan adi aromatik k nghasilkan tu da penelitian persilangan te ngkan dengan ndividu turun n tetuanya ang balik ini
ai generasi B teknik persi ntuk memind ang diinginka man pemulih ngan balik beb rogresi sifat mungkin mem en liar yang
nerasi silang erozigot, sem esi dalam ke nnya tidak ma. Oleh karen ular berbasis n alel gen badh
Wangi dan M turunan di npa bantuan a
kan sendiri (s
ada setiap ge gka waktu pen ama (Mackill
merupakan m iki kelebihan man bersifat sp
patan terhada abilitas gen s ilkan produk modifikasi) d etiknya mudah
ngan baik, 3 tahun. Kelem
adalah meto dalam satu s an organisme
rected
disebut o et al.
marker ssisted Melalui dapat yang naman ke padi urunan n ini erarah. n padi nannya dari i akan BC5F1. langan dahkan an dari (host). berapa yang mbawa tidak balik mentara eadaan dapat na itu, s PCR
h2 dari Mentik setiap analisis selfing) enerasi nelitian
Pengemb
Berdasar 2005b; Lan diintrogresi menginaktiv Umumnya i beberapa me rekayasa ge demikian m produk tana retensi tanam sulit dipredi pemuliaan k acak (Mack lain yang transgenik gen badh2 p tersebut dar metode pe
crossing) homozigot r Pada me diminimalis diinginkan pemulih da karakter yan Pembentuka menghasilka mendekati pemulih ma sehingga h dalam hal i tanaman pem diperlukan progeni ge karakter ar
Gambar 4 D te (H
angan Variet
rkan studi lite ng dan Buu pada padi n vasi gen badh
inaktivasi dap etode salah sa enetik (Wanc metode tersebu aman transge man pemulih iksi atau diko konvensional kill et al. 2007
tidak men adalah melal padi nonarom ri varietas aro ersilangan te untuk men resesif (Gamb
etode ini int asi, sehingga yang terintro an dapat dipe ng baik pada p an populasi h an turunan de hampir 100% aksimal) (Ma hanya karakt ini aromatik y mulih. Pembe
untuk introg nnya bersifa romatik), sed
Diagram m erarah (
site-Hami Seno et
tas Padi Arom
ratur (Bradbu 2008), aroma nonaromatik
h2 pada padi t pat dilakukan
atunya dengan chana 2004). ut akan mengh enik. Di lain
atau introgres ontrol jika di
melalui pers 7). Metode a nghasilkan t lui pengganti matik dengan a
omatik, mengg erarah (site-d
ndapatkan ar 3). trogresi dono
hanya karakt ogresi pada t ertahankan k padi host (Cih hingga BC5F engan karakte % (retensi t ackill et al. ter diinginka yang terintrog
ntukan BC5F gresi karakte at resesif (m dangkan untuk
metode pers
-directed cr al. 2009).
matik
ury et al. a dapat dengan tersebut. dengan n teknik Namun hasilkan n pihak, si donor lakukan silangan alternatif tanaman ian alel alel gen gunakan directed turunan or dapat ter yang tanaman karakter-herang). F1 akan er yang tanaman 2007), an saja gresi ke 2 hanya er yang misalnya
k yang
silangan
rossing)
dominan (mis hingga BC5
directed cros
introgresi k spesifik, men karakter reko stabilitas gen
et al. 2007 melalui pers dengan analis bahwa proge (backcross) aromatik.
Me
Sejumlah digunakan u menyeleksi p keterbatasan sampel. Peng satu metode padi aromat digunakan un namun kema karena indera terjadi kerus sebagai akiba keras (Bradbu dikembangka melakukan BADH2 mela 4-aminobutira dengan meng panjang gelom ini memiliki waktu banyak banyak.
Metode k melibatkan j dipanaskan menggunakan I2KI. Namun, kerusakan pa (Sood dan Si mengidentifik kromatografi memerlukan besar dan me (Lorieux et a
marka gen untuk mengid memiliki ke membedakan nonaromatik, homozigot ha 2009). salnya toleran 5F1. Persilan
ssing) memili karakter bers
nghasilkan pro ombinasi tera n hingga pulu ) . Introgre silangan tera sis molekuler eni yang aka telah menga etode Analisi metode pa untuk memba padi aromatik saat mempro gecapan indivi asli untuk m tik. Sekelom ntuk mendete ampuannya m a pengecap m sakan fisik at dari pengu ury et al. 2005 an oleh Srivon
pengukuran alui pemberia aldehid dan ggunakan spe
mbang 340 n sensitivitas te k, dan memer
kimia, melip jaringan daun
dalam ai n larutan pe , metode ini d
da bagian ron diq 1978). M kasi 2AP ad gas. Nam jaringan sam emakan waktu
al. 1996). Me adalah tekni dentifikasi gen elebihan yai n antara maupun asil persilanga nsi genangan) ngan terarah iki kelebihan sifat selektif oduk nontrans arah, dan me uhan tahun (M
si karakter arah perlu d r untuk mema an disilangba andung gen
is Aroma
anca indera antu petani k. Namun, te oses sejumlah idu merupakan menyeleksi k mpok panel
eksi variasi a menjadi berk menjadi jenuh (lecet) pada unyahan beras
5b). Metode l ng et al. (2008 aktivitas an sejumlah su
hasilnya ektrofotometer nm. Namun, m erbatas, meme
rlukan sampe
puti uji bau n atau butir ir atau d ereaksi KOH apat mengakib ngga nasal di h etode lainnya dalah menggu mun, metod mpel dalam j u yang cukup etode PCR be k yang digu n aroma. Meto itu murah, padi aro heterozigot an (Hami Seno
BAHAN DAN METODE
Bahan dan Alat
Bahan yang digunakan untuk isolasi DNA padi meliputi daun padi BC2F1 CM dan daun padi BC2F1 CP, etanol 70%, etanol 95%, fenol kloroform isoamilalkohol (PCI), isopropanol, larutan buffer Tris-EDTA (TE), natrium klorida (NaCl), setiltrimetil amonium bromida (CTAB), polivinil pirolidon (PVP), etilendiamin tetraasetat (EDTA), dan merkaptoetanol. Bahan untuk menguji hasil isolasi DNA dengan PCR adalah bufer PCR 10×, MgCl2 50 mM, dNTP mix 10 mM (dGTP, dATP, dTTP, dCTP), marka external sense primer (ESP), internal fragrant antisense primer (IFAP), internal nonfragrant sense primer (INSP), external antisense primer (EAP), marka RM223 (forward dan
reverse), Taq polymerase (FastStart), DNA 50 ng/μL, dan MQ H2O. Bahan untuk elektroforesis meliputi loading dye, bufer Tris HCl-EDTA 1× (TAE), agarosa, DNA hasil isolasi atau hasil PCR, DNA standar (1 kb ladder), etidium bromida (EtBr), dan akuades.
Alat yang digunakan dalam penelitian antara lain gunting, microfuge, autopipet, neraca analitik, autoklaf, ruang asam, mesin PCR PTC-100 (MJ Research, Inc.), vorteks, gelas piala, magnetic stirer, spektrofotometer (SmartSpec™ Plus Spectrophotometer, Biorad), pH meter, alat dokumentasi gel (chemidoc EQ gel system), elektroforesis, sentrifus (Backman rotor 12), mortar, tip pipet mikro, pipet mikro, inkubator, neraca analitik, labu Erlenmeyer, tabung mikro, inkubator bergoyang, kuvet, kertas aluminium,
stopwatch, penyedot vakum, bak plastik, ember, penangas air, microwave, stirrer plate, kertas minyak, dan cawan petri.
Metode Penelitian
Pembentukan Populasi BC2F1 CM dan
BC3F1 CP (Soedyanto et al. 1978)
Materi yang digunakan adalah tetua pemulih Ciherang dan tanaman BC2F1 CM serta BC3F1 CP. Masing-masing materi tanaman dikecambahkan dalam cawan petri. Kecambah-kecambah pertumbuhannya dipindahkan ke dalam bak pembibitan. Setelah itu, dipindahkan ke dalam ember selama ± 3 – 4 minggu. Sebelumnya, setiap tanaman dilakukan penomoran untuk mempermudah proses seleksi. Setelah masing-masing varietas padi berbunga secara bersamaan, maka dilakukan silang balik
(backcross). Sehari sebelum disilangbalikkan, dilakukan kastrasi atau emaskulasi pada bunga-bunga tanaman tetua betina agar putik menjadi masak sempurna saat penyerbukan sehingga tingkat keberhasilannya lebih tinggi. Kastrasi adalah membuang bagian tanaman yang tidak diperlukan. Setiap bunga memiliki enam benang sari dan dua kepala putik yang menyerupai rambut. Keduanya tidak boleh rusak atau pun cacat.
Bunga pada malai yang akan dikastrasi dijarangkan hingga tinggal 15-50 bunga. Sepertiga bagian bunga dipotong miring menggunakan gunting kemudian benang sari diambil dengan vacuum pump. Bunga yang sudah bersih dari benang sari ditutup dengan kertas minyak agar tidak terserbuki oleh tepung sari yang tidak dikehendaki. Bunga jantan diambil kemudian disimpan dalam bak plastik. Setelah kepala sari membuka, segera dilakukan penyerbukan. Bunga betina yang sudah dikastrasi dibuka tutupnya kemudian bunga jantan diletakkan di atasnya. Proses ini dilakukan dengan menyalakan semua lampu dalam ruangan untuk meningkatkan suhu hingga 32°C dan kelembapan udara hingga 80% sehingga dapat mempercepat pemasakan tepung sari.
Isolasi DNA Padi (Doyle dan Doyle 1987)
Senyawa DNA diisolasi dari daun muda tanaman padi yang berumur ± 3 minggu. Pemecahan sel dilakukan dengan cara penggerusan dalam mortar. Sebanyak 1000 µL (2 × 500 μL) bufer ekstraksi CTAB dimasukkan ke dalam mortar dan dicampur hingga homogen. Suspensi diinkubasi di dalam penangas air selama 15 menit (setiap 5 menit dikocok). Pemurnian DNA dari pengotor dihilangkan dengan penambahan 500 µL fenol kloroform isoamilalkohol (PCI) ke dalam tabung dan dikocok hingga merata. Suspensi selanjutnya disentrifugasi dengan kecepatan 12000 rpm selama 5 menit.
Kuantifikasi DNA dengan
Spektrofotometer (Sambrook et al. 1989)
Hasil isolasi DNA selajutnya dianalisis dengan spektrofotometeri untuk melihat konsentrasi dan kemurnian DNA. Sebanyak 2 µL DNA ditambahkan dengan 398 µL akuades dalam kuvet. Pengukuran konsentrasi sampel dilakukan pada panjang gelombang 260 nm. Kemurnian DNA sampel diukur pada perbandingan panjang gelombang 260/280nm. Sampel DNA yang murni mempunyai rasio 1.8 hingga 2.0. Apabila rasionya kurang dari 1.8 maka sampel DNA masih mengandung kontaminan protein, dan untuk menghilangkannya ditambahkan proteinase. Apabila rasionya lebih dari 2.0 maka sampel DNA masih mengandung kontaminan RNA, dan untuk menghilangkannya ditambahkan ribonuklease. Tahap selanjutnya DNA diencerkan dengan konsentrasi akhir 50 ng/ µL untuk proses amplifikasi PCR.
Seleksi PCR dengan Marka Bradbury
(Bradbury et al. 2005b)
Hasil isolasi DNA tanaman BC2F1 CM yang telah disamakan konsentrasinya selanjutnya diamplifikasi dengan mesin PCR. Campuran reaksi untuk PCR terdiri atas 2 µL bufer PCR 10×, 1.2 µL MgCl2 50 mM, 0.4 µL dNTP mix 50 mM, 1 µL untuk masing-masing marka external sense primer (ESP), internal fragrant antisense primer (IFAP), internal non-fragrant sense primer (INSP), dan
external antisense primer (EAP), 0.16 µL Taq polymerase, 2 µL DNA 50 ng/ µL, dan ddH2O. Kemudian diamplifikasi dengan mesin PCR sebanyak 30 siklus pada kondisi denaturasi awal 940C selama 2 menit, denaturasi 940C selama 30 detik, penempelan marka 550C selama 30 detik, perpanjangan marka 720C selama 45 detik, dan perpanjangan marka akhir 720C selama 5 menit.
Amplifikasi DNA dengan Marka RM223 (Lang dan Buu 2008)
Hasil isolasi DNA tanaman BC3F1 CP yang telah disamakan konsentrasinya selanjutnya diamplifikasi dengan mesin PCR. Campuran reaksi untuk PCR terdiri atas 2 µL bufer PCR 10×, 1.2 µL MgCl2 50 mM, 0.4 µL dNTP mix 50 mM, 1 µL masing-masing marka RM223 0.8 μL (forward dan reverse), 0.16 µL Taq polymerase, 2 µL DNA 50 ng/ µL, dan MQ H2O. Kemudian diamplifikasi dengan mesin PCR sebanyak 35 siklus pada kondisi denaturasi awal 94°C selama 5 menit,
denaturasi 94°C selama 1 menit, penempelan marka 55°C selama 30 detik, perpanjangan marka 72°C selama 1 menit, dan perpanjangan marka akhir 72°C selama 5 menit.
Elektroforesis DNA
Produk PCR selanjutnya dilihat dengan elekroforesis. Sebanyak 10 µL produk PCR ditambahkan dengan 2 µL loading dye dan dicampur sempurna serta dimasukkan ke dalam sumur gel agarosa (1.5% untuk elektroforesis dengan marka Bradbury dan 3% untuk elektroforesis dengan marka RM223). Marka 1 kb ladder disertakan untuk melihat ukuran DNA. Tahap selanjutnya sampel DNA dialiri arus dengan tegangan listrik 80 volt selama 35 menit. Gel agarosa diwarnai dengan larutan etidium bromida (10 mg/L) selama 10 menit, kemudian dihilangkan pewarnaannya dengan air selama 5 menit. Gel agarosa selanjutnya divisualisasi dengan chemidoc gel system.
HASIL DAN PEMBAHASAN
Pembentukan Populasi BC2F1 CM dan BC3F1 CP
Persilangan tanaman padi merupakan penggabungan karakter melalui pertemuan tepung sari dengan kepala putik yang menghasilkan embrio dan kemudian embrio berkembang menjadi benih. Menurut Welsh (1981), kombinasi karakter dari kedua tetua pada individu generasi pertama terjadi secara acak, sehingga dapat menghasilkan kombinasi karakter yang lebih menguntungkan dari kedua tetuanya. Komposisi gen yang diperoleh dari hasil persilangan padi BC3F1 CP yaitu 93.75%:6.25%, sedangkan padi BC2F1 CM adalah 87.5%:12.5%, artinya gen dari tetua Ciherang bersegregasi lebih banyak dalam tanaman yang disilangbalikkan. Individu BC2F1 maupun BC3F1 yang dihasilkan bersifat heterozigot dan akan mengalami pemisahan pada generasi-generasi berikutnya.
dipindahkan menyesuaik yang baru. selama kura tidak stres Tanaman k ember berisi Persilang pembungaan betina b penghitunga hal kunci d ini, tetua C seminggu (backcross) balik ditana silang balik tanaman B dapat dilihat
Gambar 5 P p C (b b d ta em ta b (p p
n ke dalam ba kan diri dengan
Masa penyes ang lebih 2 m dengan per kemudian dip i tanah hingga gan dilakuk n antara tetu bersamaan.
an jarak waktu alam persilan Ciherang ditan
sebelum tan ditanam, s am, dan semin k ditanam. Pe C2F1 CM m t pada gambar
embentukan adi BC2F1 C CP; (a) benih b) disemai da
enih sudah ipindahkan k anah; (e) dip mber berisi anaman; (g)
erbunga (bunt penyerbukan)
ersilangan.
ak berisi tana n kondisi ling suaian ini di minggu agar t rubahan lingk pindahkan ke
a masa panen. kan ketika ua jantan da
Pengaturan u tanam pun m ngan. Pada pe
nam tiga kal naman silang
aat tanaman nggu setelah t embentukan p maupun BC3 r 5.
populasi t CM maupun
yang telah d alam cawan p
h tumbuh; ke dalam bak pindahkan ke
tanah; (f) p malai yang ting); (h) disi ; (i) benih
ah untuk gkungan lakukan tanaman kungan. e dalam masa an tetua dan menjadi enelitian li yaitu: g balik silang tanaman populasi 3F1 CP tanaman BC3F1 dijemur; petri; (c) ; (d) k berisi e dalam populasi g telah langkan h hasil Ketelitian sangat dibutu Jika kastrasi maka akan penyerbukan hal itu terjad
directed cros
Kastrasi ada tanaman yan dilakukan pa tanaman BC2 hal ini, bagi benang sari memiliki ena putik yang boleh rusak. setelah kastr matang dan k lebih tinggi. kemudian dit tidak terjad diharapkan (G Penyerbuk mengambil te yaitu tanam diletakkan d digerakkan a putik. Bij dilakukannya persilangan melengkung, (Gambar 5i). Pengujian K Uji kuanti konsentrasi D digunakan u DNA. Konse gelombang pengenceran dilihat pada p 260/280 nm. rasio kemurn hingga 2.0. bawah angk sampel DNA berupa protei angka 2.0 m masih terdapa
Hasil p menunjukkan murni. Hal kemurnianny 2.2 baik itu sa maupun BC3 disebabkan k kurang steril, ataupun
n khusus d uhkan terutam
tidak dilaku n menyeba sendiri pada di maka persil
ssing) dinyata alah pembuan ng tidak dib ada bunga b 2F1 CM dan ian yang tida dan tepung am benang sa menyerupai Penyerbukan asi, sehingga keberhasilan p Bunga yan tutupi dengan di persilang Gambar 5h). kan dilakuk epung sari dar man Ciheran di atas bun agar tepung s
ji terbentuk a penyerbuk akan me ramping, Kuantitatif da itatif digunak DNA, sedan untuk meng ntrasi DNA d
260 nm 200 kali, sed perbandingan
Menurut Sam nian DNA ya Apabila rasio ka 1.8 meng
A masih ter in, sedangkan mengindikasik at kontaminan pengukuran n bahwa sam
ini dapat a yang berad ampel DNA ta 3F1 CP. Kura karena proses
, sehingga m RNA (da
dalam persi ma ketika ka ukan dengan abkan terj
bunga betina langan terarah akan tidak be ngan bagian-b
butuhkan. K betina, yaitu
BC3F1 CP. D ak diperlukan
sari. Setiap ari serta dua rambut dan n dilakukan a putik benar penyilangan m ng telah dik kertas minya gan yang
kan dengan ri bunga tetua ng. Bunga nga betina
ari jatuh ke 2 minggu kan. Biji empunyai b
dan tanpa
an Kualitatif
kan untuk men gkan uji ku getahui kem diukur pada p
dengan dangkan kem
panjang gelom mbrook et al. (
ang baik adal o berada bera gindikasikan
rdapat konta rasio berada kan bahwa s n berupa RNA yang dip mpel DNA k
dilihat dari da pada kisara
anaman BC2F ang murninya
isolasi DNA enyebabkan p ari lingku langan astrasi. benar adinya a. Jika h (
mengkontaminasi DNA sampel. Namun hal ini masih bisa ditolerir karena konsentrasi protein maupun RNA sangatlah kecil yaitu berkisar 5 – 10 μg/mL, sehingga tidak mengganggu analisa selanjutnya.
Konsentrasi DNA yang diperoleh berada pada kisaran 1000 – 5000 μg/mL baik itu sampel DNA tanaman BC2F1 CM maupun BC3F1 CP. Konsentrasi DNA yang beragam tersebut kemudian diseragamkan menjadi 50 μg/mL dengan cara pengenceran menggunakan air destilata. Tujuan penyeragaman konsentrasi DNA agar hasil amplifikasi pada sampel yang satu dengan yang lainnya tidak berbeda jauh, sehingga ketika dielektroforesis akan mempunyai ketebalan pita yang sama.
Seleksi Tanaman Padi BC2F1 CM dengan Menggunakan Marka Bradbury
Tidak semua bunga yang disilangkan menghasilkan benih BC2F1 yang mengandung gen heterozigot badh2, sehingga perlu dilakukan seleksi tanaman BC2F1 CM. Seleksi tanaman BC2F1 CM tidak dapat diuji secara langsung melalui uji aroma biasa untuk melihat apakah gen badh2 dari tanaman aromatik telah bergabung dengan alel gen padi nonaromatik. Oleh karena itu, digunakan marka molekuler berbasis PCR. Analisis PCR dalam penelitian ini menggunakan marka berdasarkan penelitian yang telah dilakukan sebelumnya oleh Padmadi (2009).
DNA tanaman padi BC2F1 CM hasil persilangan Ciherang dengan Mentik Wangi diamplifikasi dengan menggunakan marka Bradbury. Berdasarkan penelitian sebelumnya (Padmadi 2009), marka Bradbury tidak sepenuhnya dapat membedakan padi aromatik dan nonaromatik pada beberapa varietas lokal Indonesia. Varietas padi aromatik yang mempunyai pola pita DNA yang berbeda dengan padi varietas nonaromatik adalah Mentik Wangi dan Gunung Perak. Hal tersebut kemungkinan disebabkan oleh perbedaan delesi pada kromosom 8. Selain itu, penggunaan marka RM 223 tidak dapat membedakan Mentik Wangi dengan varietas nonaromatik lainnya. Oleh karena itu, marka Bradbury digunakan untuk menyeleksi tanaman BC2F1 CM. Keuntungan menggunakan marka ini yaitu mampu membedakan gen yang homozigot dominan (pada varietas nonaromatik dalam hal ini Ciherang), homozigot resesif (pada varietas aromatik dalam hal ini Mentik Wangi), dan heterozigot (pada varietas hasil persilangan). Penelitian sebelumnya Bradbury et al. (2005)
mengatakan bahwa gen aroma (badh2) merupakan gen yang bersifat resesif. Tanaman hasil persilangan disinyalir memiliki gen dengan pola heterozigot yang mana pada tanaman hasil persilangan telah mengandung pita alel dari kedua tetua. Gen aroma yang bersifat resesif ini tidak terkspresi secara utuh sehingga untuk mengekpresikan karakter aroma ini perlu dilakukan selfing pada generasi BC5F1.
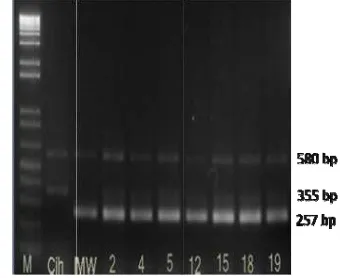
Marka Bradbury terdiri dari empat marka, yaitu ESP (External Sense Primer), EAP (External Antisense Primer), INSP (Internal Nonfragrant Sense Primer), dan IFAP (Internal Fragrant Antisense Primer). Prinsip kerja dari keempat marka tersebut adalah dua marka EAP dan ESP akan menempel pada varietas padi aromatik dan nonaromatik. Kedua marka tersebut merupakan kontrol positif yang akan menghasilkan pita DNA berukuran 580 bp. Pasangan marka ESP dan IFAP akan menempel pada varietas aromatik dan akan menghasilkan pita DNA berukuran 257 bp. Pasangan marka EAP dan INSP akan menempel pada varietas nonaromatik dan menghasilkan pita DNA berukuran 355bp (Gambar 6).
Hasil amplifikasi menunjukkan terdapat tujuh tanaman yang positif memiliki gen heterozigot badh2 dari keseluruhan dua puluh empat tanaman. Pola gen heterozigot badh2
memiliki dua buah pita yang merupakan gabungan dari kedua tetua tanaman padi Ciherang 355 bp dan Mentik Wangi berukuran 257 bp, hal ini mendukung pernyataan Bradbury et al. (2005) dan penelitian sebelumnya (Padmadi 2009; Praptiwi 2010; Sugihartati 2010). Hasil amplifikasi PCR ditampakkan dengan gel agarosa 1,5%. Penggunaan konsentrasi gel agarosa tersebut berdasarkan ukuran produk PCR yang dihasilkan. Menurut Sambrook et al. (1989), penggunaan konsentrasi tersebut untuk ukuran fragmen DNA 250 bp – 12 kbp.
Pola gen he hasil amplif elektroforeg elektroforeg ukuran prod Wangi lebih karena adan padi Menti bahwa gen mengalami dalam suatu genetik. Gen mengalami fertil.
Gambar 7 H p m la W C 1 Seleksi P
Marka a menyeleksi Ciherang d marka RM Berdasarkan 2009), mar secara jela Ciherang de polimorfism Namun, ke pola amplifi Pandan Wa karena itu, menyeleksi menggunaka membedaka (dominan), heterozigot. Marka DNA Cihera Wangi 140
eterozigot ini t fikasi yang d gram (Gam gram dapat
duk hasil amp h kecil dibandi nya delesi ba ik Wangi. I
badh2 padi mutasi akib u populasi ya
n badh2 pada mutasi atau m
Hasil amplifik adi BC2F1 marka Bradbur
adder; Ciher Wangi (MW) CM nomor 2,
9.
Padi BC3F1 C Marka RM
aromatik yan tanaman dengan Pand M223 (Lang
n penelitian se rka RM223 s pola amp engan Pandan me pada kro
tika digunak ikasi antara p angi tidak dap
marka RM2 padi BC3F an marka an homo
homozigot a
RM223 dap ang dengan uk
bp, dan BC3
terlihat dari p ditampakkan mbar 7).
diketahui plifikasi padi
ingkan padi C asa pada gen Ini mengindi Mentik Wang at peristiwa ang terisolasi a padi Ciheran masih dalam k
kasi DNA t CM mengg ry. Marker (M rang (Cih); ); Tanaman 4, 5, 12, 15,
CP Mengguna M223 ng digunakan hasil pers dan Wangi dan Buu ebelumnya (P dapat memb lifikasi antar Wangi akibat omosom nom kan marka B padi Ciherang
pat dibedakan 23 digunakan F1 CP. Keun ini yaitu zigot no aroma (resesi
pat mengamp kuran 160 bp, 3F1 CP meng
pola pita melalui Dari bahwa Mentik Ciherang n badh2
ikasikan gi telah evolusi i secara ng tidak keadaan tanaman gunakan M) 1 kb Mentik BC2F1 18, dan akan n untuk silangan adalah 2008). Padmadi bedakan ra padi t adanya mor 8. Bradbury dengan n. Oleh n untuk ntungan mampu onaroma if), dan plifikasi Pandan gasilkan
dua pita den Penelitian se telah membu membedakan Ciherang. Pen mengamplifik nonaromatik DNA antara (Lang dan Bu
Elektrofor puluh tiga menunjukkan tanaman padi gen heterozig seperti yang elektroforesis pita yang m tetua padi (Gambar 9). H produk ampl besar daripad adanya delesi Adanya dele Pandan Wan tersebut telah yang terjadi penelitian ju amplifikasi menunjukkan mengandung
Gambar 8 Pro RM
Gambar 9 Ha pad ma lad Wa nom 21 ngan ukuran ebelumnya (P uktikan mark n padi Pan nggunaan ma kasi padi
dengan varia 120 bp – 1 uu 2008).
regram yang tanaman p n bahwa s
i BC3F1 CP p got badh2. Po terlihat pad s gel agarosa merupakan kom
Ciherang da Hasil penelitia
ifikasi dari p da Pandan Wan
i basa pada p esi basa pada ngi mengindik h mengalami m
pada padi M uga menunju pada tanam n sifat h
gen dari kedu
oses amplifik M223 (Lang d
asil amplifika di BC3F1 arka RM223. dder; Ciheran angi (PW); T mor 1, 3, 7, 9 .
120–160bp
140 dan 16 admadi 2009 ka RM223 m ndan Wangi arka RM223 m
aromatik asi panjang fr
60 bp (Gamb
diperoleh da padi BC3F1 ebanyak sem positif menga ola gen heter da pola pita memiliki dua mbinasi dari an Pandan W
an diperoleh u padi Ciherang angi disebabka padi Pandan W
a gen badh2
kasikan bahw mutasi sama h Mentik Wangi. ukkan bahwa man BC3F1 heterozigot ua tetua. kasi dengan dan Buu 2008)
asi DNA tan CP menggu
Marker (M) ng (Cih); P Tanaman BC3
, 12, 14, 15, 1 60 bp. 9) juga mampu i dan mampu dan agmen mbar 8) ari dua 1 CP mbilan andung rozigot a hasil a buah kedua Wangi ukuran g lebih an oleh Wangi.
2 padi wa gen halnya . Hasil a pola 1 CP yaitu marka ). naman unakan ) 1 kb Pandan
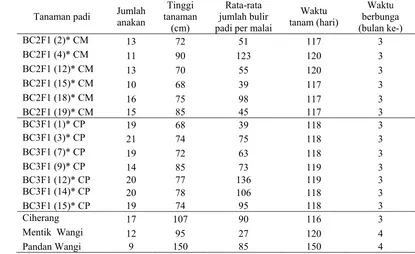
Hasil menggunaka konsentrasi basa yang tidak akan konsentrasi Pengam BC2 Pengama terhadap tan gen heteroz tanaman y anakan, tin butir padi waktu berb dilakukan ( keragaman k pemulih terh Pengama menunjukka anakan) dan (39 – 123 Ciherang (1 malai) (He tanam BC2 waktu berbu mengikuti waktu tanam (Hermanto tanaman BC belum men cm). Karak
Tabel 1 Has
Tanaman BC2F1 (2)* BC2F1 (4)* BC2F1 (12) BC2F1 (15) BC2F1 (18) BC2F1 (19) BC3F1 (1)* BC3F1 (3)* BC3F1 (7)* BC3F1 (9)* BC3F1 (12) BC3F1 (14) BC3F1 (15) Ciherang Mentik Wan Pandan Wan Ket.: (*) = n
PCR ditam an gel agaros
tersebut berd sangat kecil
terlihat berbe gel agarosa di
matan Karakt 2F1 CM dan
atan karakter naman yang p zigot badh2.
ang diamati nggi tanaman
per malai, w bunga. Hasil
(Tabel 1) me karakter dari t hadap tanaman atan tanaman an jumlah a n jumlah gab butir) telah 4 – 17 anakan rmanto 2006 2F1 CM (117 unganya (bula karakter Cih m dan berbung
2006). S C2F1 CM (6 ngikuti karakt kter ini masi
sil pengamatan
n padi Ju an CM CM * CM * CM * CM * CM CP CP CP CP * CP * CP * CP ngi ngi nomor sampel
mpakkan d sa 3%. Pengg dasarkan perb yaitu 20 bas eda jika digu i bawah 3%.
ter Tanaman BC3F1 CP tanaman dila positif menga Variabel ka meliputi j n, rata-rata j waktu tanam
pengamatan enunjukkan a tetua donor m n hasil persila n padi BC2F
anakan (11 bah padi per mengikuti ka n dan 90 gaba 6). Adapun 7 – 120 hari an ke-3) juga herang (116 ga pada bulan Sementara,
8 – 90 cm) ter Ciherang ih mengikuti
n karakter tana
umlah nakan
Tin tana (c
13 7
11 9
13 7
10 6
16 7
15 8
19 6
21 7
19 7
14 8
20 7
20 7
19 7
17 1
12 9
9 1 tanaman dengan gunaan bedaan sa dan unakan n Padi akukan andung arakter umlah umlah m, dan yang adanya maupun angan. 1 CM – 16 malai arakter ah per waktu i) dan a telah hari n ke-3) tinggi masih (107 tetua aman padi nggi aman cm) R jum padi 72 90 70 68 75 85 68 74 72 85 77 78 74 07 95 50 donornya memiliki ti (Hermanto BC2F1 CM Gabah bulat gemuk hasil penga sudah men berbulu, be gabah kunin ada yang b sudah meng bulat, dan ta
Gambar 10
Rata-rata mlah bulir
i per malai 51 123 55 39 98 45 39 75 63 73 136 106 95 90 27 85 yaitu Ment inggi tanama 2006). Se M telah mengik padi Pandan k, berbulu, dan amatan, ada b gikuti karakt entuk, panjan ng bersih). Se berbulu (Gamb gikuti karakte ahan rontok). Gabah pa (panah me BC3F1 CP Waktu tanam (hari) 117 120 120 117 117 117 118 118 118 119 119 118 118 116 120 150 tik Wangi an berkisar 9 ecara keselu kuti Ciherang. n Wangi berb
n tahan rontok bentuk gabah ter Ciherang ng ramping, ebagian lain, bar 10) akan r Ciherang (g
adi yang b erah) pada tan
Pengamatan tanaman BC3F1 CP diketahui bahwa waktu tanam BC3F1 CP (118 – 119 hari) dan waktu berbunganya (bulan ke-3) telah mengikuti karakter Ciherang (116 hari waktu tanam dan berbunga pada bulan ke-3) (Hermanto 2006). Pengamatan yang dilakukan terhadap jumlah anakan BC3F1 CP (14 – 21 anakan) dan jumlah gabah per malai (39 – 136 bulir) juga telah mengikuti karakter Ciherang (14 – 17 anakan dan 90 gabah per malai) (Hermanto 2006). Sementara, tinggi tanaman BC3F1 CP (68 – 85 cm) masih belum mengikuti karakter Ciherang (107 cm). Namun, secara keseluruhan karakter BC3F1 CP telah mengikuti karakter Ciherang.
SIMPULAN DAN SARAN
Simpulan
Pembentukan tanaman padi BC3F1 CP dan BC2F1 CM berhasil dilakukan. Keberhasilan ini dapat dilihat dari pita heterozigot menggunakan marka spesifik Bradbury untuk BC2F1 CM dan RM223 untuk BC3F1 CP. Marka Bradbury dan RM223 dapat melacak introgresi badh2
termutasi dari padi Mentik Wangi dan Pandan Wangi ke padi Ciherang. Kedua marka tersebut juga dapat menentukan status gen (fertil atau termutasi) dan alel (homozigot atau heterozigot) badh2. Tanaman BC2F1 CM dan BC3F1 CP secara umum telah mengikuti karakter tetua Ciherang.
Saran
Perlu dilakukan penelitian lebih lanjut hingga dihasilkan tanaman generasi BC5F2 untuk mendapatkan benih Ciherang nontransgenik aromatik. Penentuan urutan basa nukleotida pada padi varietas aromatik dan nonaromatik lokal lainnya juga perlu dilakukan untuk menentukan marka aromatik yang lebih spesifik untuk padi-padi tersebut.
DAFTAR PUSTAKA
Adijono P, Bambang K, Allidawati, Suwarno. 1993. Pemuliaan padi aromatik dan ketan. Dalam: Mahyudin Syam, Hermanto, A. Musadad dan Sunihaardi (eds.). Kinerja Penelitian Tanaman Pangan. Pusat Penelitian Tanaman Pangan. Bogor. hal 422-428.
Ahn SN, Bollish CN, Tanksley SD. 1992. RFLP tagging of a gene for aroma in rice. Theor Appl Genet 84: 825-828.
Berner DK, Hoff BJ. 1986. Inheritance of scent in American long grain rice. J Crop Sci 26: 876-878.
Bhattacharjee et al. 2002. Basmati rice: a review. International. J Food Sci and Tech 37: 1-12.
Bourgis et al. 2008. Characterization of the major fragrance gene from an aromatic japonica rice and analysis of its diversity in Asian cultivated rice. Theor Appl Genet 117: 353-358.
Bradbury L. 2007. Identification of the gene responsible for fragrance in rice characterisation of the enzyme transcribed from this gene and its homolog [tesis]. Australia: Faculty of Science and Management, School of Environmental.
Bradbury LM, Fitgerald TL, Henry RJ, Jin Q, Waters DLE. 2005a. The gene for fragrance in rice. J Plant Biotech 3: 363-370.
Bradbury LMT, Henry RJ, Jin Q, Reinke RF, Waters DLE. 2005b. A perfect marker for fragrance genotyping in rice. J Mol Breed 16: 279-283. Buttery RG, Ling LC, Juliano BO,
Turnbaugh JG. 1983. Cooked rice aroma and 2-acetyl-1-pyroline in rice. J Agric Food Chem 31: 823-826.
Doyle JJ, JL Doyle. 1990. Isolation of plant DNA from fresh tissue. Focus 12: 13-15.
Gasser CS, Farley RT. 1989. Genetically engineered plants for crop improvement. Science 244: 1293-1299.
Heldt W. 2005. Plant Biochemistry. German: Elvesier All Right Reserved.
Hermanto. 2006. Padi Ciherang makin populer. Warta Penelitian dan Pengembangan Pertanian 28:14-15.
[terhubung berkala]. http://crifc@indo.net.id/journal/warta
[10 Jan 2010].
Itani et al. 2004. Variation of 2-acetyl-1-pyrroline concentration in aromatic rice grains collected in the same region in Japan and factors affecting its concentration. Plant Production Science 7:178-183.
Krisnamurthi B. 2006. Produksi padi nasional naik minimum sama dengan kenaikan penduduk 1,5 %. Sinar Tani. [14 Maret 2006]
Krishnan HB, Okita TW. 1986. Structure relationship among the rice glutelin polypeptides. Pant Physiol 88: 649-655.
Lang NT, Buu BC. 2008. Development of PCR based markers for aroma (fgr) gene in rice (Oryza sativa L.).
Omonrice 16: 16-23.
Lorieux M, Petrov M, Huang N, Guiderdoni E, Ghesquiere A. 1996. Aroma in rice: genetic analysis of a quantitative trait. Theor App Genet
93: 1145-1151.
Mackill et al. 2007. Marker assisted selection for submergence tolerance in rice. J Mol Plant Breed 5: 207-208.
Mikkelsen SR, Corton E. 2004.
Bioanalytical Chemistry. New Jersey: John Wiley & Sons.
Mittal, UK, Preet K, Singh D, Shukla KK, Saini RG. 1995. Variability of aroma in some land races and cultivar of scented rice. J Crop Improv 22: 109-122.
Nasution MA. 2002. Biologi molekuler dan ketahanan pangan nasional. [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Nurcahyo. 2009. Padi dan segala permasalahannya: Rekayasa genetika padi tidak berguna dibanding praktik pengembangan padi tradisional. Warta Pertanian 1 – 5 [terhubung berkala].
http://indonesiaindonesia.com/wart a [20 Mei 2009].
Padmadi B. 2009. Identifikasi sifat aroma tanaman padi menggunakan marka berbasis gen aromatik [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Praptiwi D. 2010. Pembentukan dan seleksi F1 padi Ciherang-Pandan Wangi dan Fatmawati-Mentik Wangi menggunakan marka aromatik [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Reyes MH. 2000. A model for marker based selection in gene introgression breeding program. J Crop Sci 40: 91–98.
Sambrook J, Fritsch EF, Maniatis T. 1989.
Molecular Cloning 3rd edition. New York: Cold Spring Harbor Laboratory Pr.
Soedyanto et al. 1978. Bercocok Tanam
Jilid II. Jakarta: Yasaguna.
Sood BC, Sidiq EA. 1978. A rapid technique for scent determination in rice Indian. J Genet Plant Breed
38: 268-271.
Srivong P, Wangsomnuk P, Pongdontri P. 2008. Characterization of a fragrant gene enzymatic activity of betaine aldehyde dehydrogenase in aromatic and non aromatic Thai
rice cultivar. J KKU Sci
36:290-301.
Sugihartati. 2010. Aplikasi marka aromatik Bradbury dan RM223 untuk identifikasi hasil persilangan Ciherang-Mentik Wangi dan Ciherang-Pandan Wangi [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Wanchana S et al. 2004. Enhancing 2 acetyl-1-pyroline synthesis in rice leaves by RNAi-mediated suppression of Os2Ap convert non-aromatic to non-aromatic rice (Oryza sativa L.). Proceedings of the 1.sup.st International Conference on Rice for the future, p. 105.
Xu K, Mackill DJ. 1996. A major locus for submergence tolerance mapped on rice chromosome 9. Mol Bree 2: 219–224.
Yoshihashi T, Huong NTT, Inatomi H. 2002. Precursors of
2-acetyl-1-pyrroline, a potent flavour compound of an aromatic rice variety. J Agric Food Chem 50: 2001–2004.
Lampiran 3 Komposisi bufer ekstraksi DNA
Larutan Stok Jumlah untuk 500
mL Konsentrasi akhir Tris-HCl 1 M (pH = 8,5)
NaCl 5 M EDTA 0.25 M CTAB
PVP
Merkaptoetanol
50 mL 140 mL 40 mL 10 g 10 g 1 mL
Lampiran 4 Pengukuran konsentrasi, kemurnian, dan contoh perhitungan pengenceran DNA
•
Pada tanaman BC2F1 CM
Sampel A260 [ Konsetrasi ] A280 A260/A280
1 0.262 2624.9424 0.149 1.7568
2 0.560 5596.7666 0.359 1.5580
3 0.357 3571.5569 0.203 1.7579
4 0.319 3188.4675 0.233 1.3701
5 0.276 2764.0200 0.144 1.9223
6 0.413 4126.1475 0.280 1.4746
7 0.207 2074.6455 0.130 1.6006
8 0.271 2707.8533 0.173 1.5626
9 0.531 5312.3774 0.335 1.5853
10 0.401 4014.7375 0.219 1.8373
11 0.370 3695.0454 0.194 1.9057
12 0.384 3837.4502 0.222 1.7287
13 0.354 3536.4558 0.190 1.8601
14 0.343 3432.8232 0.226 1.5158
15 0.291 2914.5796 0.199 1.4673
16 0.284 2844.5715 0.212 1.3440
17 0.190 1904.0212 0.098 1.9446
18 0.283 2829.3538 0.157 1.8023
19 0.208 2080.3240 0.110 1.8840
20 0.521 5207.7222 0.339 1.5354
21 0.491 4912.7627 0.262 1.8777
22 0.367 3668.7546 0.205 1.7883
23 0.274 2742.4585 0.148 1.8583
24 0.408 4084.4839 0.284 1.4371
25 0.403 4027.1741 0.253 1.5889
26 0.312 3117.1870 0.161 1.9356
27 0.153 1532.9608 0.084 1.8232
28 0.329 3290.8687 0.186 1.7657
29 0.376 3761.4783 0.270 1.3926
30 0.456 4563.3438 0.294 1.5500
31 0.451 4505.0259 0.304 1.4835
32 0.270 2698.0168 0.201 1.3422
Cih 0.165 1648.0126 0.087 1.8893
MW 0.025 253.0215 0.010 2.4586
Contoh perhitungan pengenceran DNA pada sampel nomor 5: M1 = konsentrasi DNA; V1 = volume DNA yang akan diambil M2 = konsentrasi akhir (50 μg/mL); V2 = volume akhir (500 μL) M1 × V1 = M2 × V2
2764.0200 μg/mL × V1 = 50 μg/ml × 500 μL 25000 μL μg/mL
(Lanjutan lampiran 4)
•
Pada tanaman BC3F1 CP
Sampel A260 [ Konsentrasi ] A280 A260/A280
1 0.266 2656.9900 0.132 2.0134
2 0.287 2868.5237 0.149 1.9240
3 0.270 2698.4805 0.149 1.8148
4 0.295 2946.1194 0.143 2.0641
5 0.234 2336.9387 0.105 2.2238
6 0.416 4156.6206 0.209 1.9911
7 0.337 3366.4985 0.160 2.1088
8 0.330 3304.3716 0.165 1.9920
9 0.188 1882.8676 0.089 2.1117
10 0.126 1262.8243 0.062 2.0291
11 0.379 3787.0259 0.189 2.0076
12 0.155 1551.8027 0.076 2.0392
13 0.408 4084.5293 0.202 2.0220
14 0.531 5305.7427 0.264 2.0098
15 0.272 2723.9312 0.136 2.0099
16 0.321 3207.1067 0.160 2.0038
17 0.281 2810.1077 0.144 1.9544
18 0.196 1957.1079 0.090 2.1711
19 0.551 5512.1055 0.275 2.0017
20 0.401 4010.3457 0.216 1.8574
21 0.195 1946.4240 0.095 2.0444
22 0.358 3576.1089 0.183 1.9501
23 0.255 2552.3354 0.126 2.0326
Cih 0.165 1648.0126 0.087 1.8893
PW 0.093 929.4803 0.049 1.8801
Contoh perhitungan pengenceran DNA pada sampel nomor 3: Nomor sampel 3
M1 : konsentrasi DNA; V1 : volume DNA yang akan diambil M2 : konsentrasi akhir (50 μg/mL); V2 : volume akhir (500 μL) M1 × V1 = M2 × V2
2698.4805 μg/mL × V1 = 50 μg/ml × 500 μL 2698.4805 μg/mL × V1 = 25000 μLμg/mL 25000 μL μg/mL
V1 = 2698.4805 μg/mL = 9.26 μL = 9.3 μL
Lampiran 5
•
Co
Ket: (M) sampel tana
•
Co
Ket: (M) tanaman BC
5 Contoh has
ontoh hasil s
= marker; (C aman BC2F1
ontoh hasil s
= marker; ( C3F1 CP; [*
*
*
sil elektrofor
seleksi BC2
C) = tetua Ci 1 CM; [*] =
seleksi BC3
C) = tetua C *] = tanaman
* * *
*
resis produk
2F1 CM
iherang; (MW tanaman yan
3F1 CP
Ciherang; (P n yang menga
* *
PCR
W) = tetua M ng mengandu
P) = tetua Pa andung gen h
*
* *
Mentik Wang ung gen hete
andan Wangi heterozigot b
* *
* * *
gi; (1-24) = n rozigot badh
i; (1-23) = n badh2.
*
* *
nomor h2.
Lampiran 6 Sekuens marka Bradbury dan RM 223
Marka Bradbury (Bradbury et al. 2005) EAP : 5’-AGTGCTTTACAAAGTCCCG-3’ ESP : 5’-TTGTTTGGAGCTTGCTGATG-3’ INSP : 5’-CTGGTAAAAAGATTATGGCTTCA-3’ IFAP : 5’-CATAGGAGCAGCTGAAATATATACC-3’
Marka RM223 (Lang dan Buu 2008)
forward : 5’-GAGTGAGCTTGGGCTTGGGCTGAAAC-3’
ABSTRAK
TAUFIQ. Seleksi Menggunakan PCR Berdasarkan Marka Gen badh2 pada
Pembentukan BC2F1 Ciherang/Mentik Wangi dan BC3F1 Ciherang/Pandan
Wangi. Dibimbing oleh DJAROT SASONGKO HAMISENO dan TRI JOKO
SANTOSO.
PENDAHULUAN
Berdasarkan aromanya, padi dapat dibedakan menjadi padi aromatik dan nonaromatik. Umumnya padi nonaromatik lebih tahan terhadap penyakit dan stres, tidak terlalu selektif dalam pemilihan lokasi dan kondisi lahan, penanaman dan pemeliharaan lebih mudah, waktu tanam lebih singkat, dan produktivitasnya lebih tinggi. Namun demikian, rasa dan aroma yang merupakan satu-satunya kelebihan padi aromatik mempunyai aspek komersial yang tinggi. Selain nilai jualnya yang lebih tinggi, permintaan pasar nasional maupun internasional akan padi aromatik sangatlah tinggi dan meningkat terus, terutama pada masyarakat dengan taraf ekonomi yang baik.
Krisnamurthi (2006) menyebutkan bahwa setiap peningkatan pendapatan per kapita Rp 1.000,- menyebabkan kenaikan permintaan beras aromatik sebesar 3.570 kg, sedangkan setiap kenaikan jumlah penduduk 1.000 orang menyebabkan kenaikan permintaan beras nonaromatik sebesar 2.140 kg. Oleh karena itu, dalam rangka ketahanan pangan akan sangat prospektif jika aroma dapat ditambahkan pada padi nonaromatik dengan tanpa merusak kelebihan-kelebihan padi tetua pemulih (host) seperti yang telah disebutkan. Hal ini akan membuat petani mendapatkan produk beras aromatik dengan kemudahan, waktu, resiko dan produktivitas seperti menanam padi nonaromatik.
Hasil penelitian mendapatkan aroma padi disebabkan karena mutasi pada gen badh2
(Bradbury et al. 2005a,b; Borguis et al. 2008). Oleh karena itu karakter aroma dapat diintrogresi (dimasukkan) pada padi nonaromatik melalui inaktivasi gen badh2nya. Inaktivasi dapat dilakukan dengan berbagai metode rekayasa genetik (Wanchana et al. 2004, Vanavichit et al. 2008). Namun metode-metode ini akan menghasilkan produk varietas tanaman transgenik yang pemasarannya terhambat dengan regulasi GMO (Genetically Modified Organisms) yang ketat. Di lain pihak, persilangan acak (random crossing) akan menghasilkan turunan dengan karakter yang sulit diramalkan. Aromatisasi nontransgenik padi nonaromatik dapat dilakukan melalui penggantian alel gen badh2
padi nonaromatik dengan alel gen terkait dari varietas aromatik, menggunakan metode persilangan terarah (site-directed crossing) untuk mendapatkan turunan homozigot resesif (Hami Seno et al. 2009).
Penelitian ini merupakan bagian dari usaha pengembangan varietas aromatik baru nontransgenik yang karakter agronominya sebaik padi nonaromatik (BC5F2), menggunakan host Ciherang dan donor aroma Mentik Wangi atau Pandan Wangi. Tanaman BC1F1 Ciherang/Mentik Wangi (CM) dan BC2F1 Ciherang/Pandan Wangi (CP) yang diperoleh dari penelitian sebelumnya (Sugihartati 2009) disilangbalikkan (backcross) lebih lanjut dengan Ciherang sehingga menghasilkan BC2F1 CM dan BC3F1 CP. Seleksi dilakukan mengunakan PCR berbantuan marka Bradbury (Bradbury et al. 2005b) untuk CM dan RM223 (Lang dan Buu 2008) untuk CP. Pemilihan Ciherang karena merupakan varietas pengembangan lokal yang akrab dengan petani maupun konsumen, tahan hama dan penyakit, kondisi lahan budidaya tidak memerlukan prasyarat yang ketat sehingga budidaya dapat dilakukan di berbagai lokasi di Indonesia dengan produktivitas yang tinggi (hampir 8 ton/ha) (Krisnamurthi 2006).
Mentik Wangi dan Pandan Wangi juga merupakan varietas lokal yang juga telah akrab dengan petani dan konsumen serta aroma digemari masyarakat Indonesia, bahkan Internasional. Namun demikian, kurang tahan hama dan penyakit, lokasi dan kondisi lahan budidaya spesifik, waktu panen lebih lama, dan produktivitas kurang tinggi (hampir 4 ton/ha) (Krisnamurthi 2006). Diharapkan jika varietas aromatik tersebut disilangkan dengan varietas Ciherang, disilangbalikkan lima kali, kemudian diselfing, seperti pada metode persilangan terarah (site-directed crossing) (Hami Seno et al. 2009), akan diperoleh varietas Ciherang aromatik (BC5F2). Varietas baru dengan karakter agronomi sebaik Ciherang seperti yang telah disebutkan, serta beraroma seperti Mentik Wangi dan Pandan Wangi. Karakter positif dari varietas baru tersebut diharapkan dapat meningkatkan ekonomi petani dan sekaligus devisa negara jika diekspor, serta lebih menggairahkan minat pertanian, baik petani maupun industri.
dengan resiko kegagalan panen, kemudahan tanam, produktivitasnya seperti menanam padi Ciherang sehingga berimplikasi padi aromatik menjadi murah.
TINJAUAN PUSTAKA
Deskripsi Tanaman Padi
Tanaman padi (Oryza sativa L.) diklasifikasikan dalam divisi Spermatophyta, subdivisi Angiospermae, kelas Monocotyledone, ordo Poales/Glumiflorae, famili Graminae, genus Oryza, dan spesies
Oryza sativa (Siregar 1981). Padi yang termasuk ke dalam genus Oryza, Oryza sativa
L. merupakan tanaman padi yang banyak dibudidayakan di Asia, sedangkan Oryza glaberrima dibudidayakan di Afrika.
Berdasarkan aromanya, padi dikelompokkan atas padi aromatik dan nonaromatik. Varietas padi aromatik diantaranya yaitu Sintanur, Gilirang, Pulu Mandoti, Pare Bau, Gunung Perak, Pinjan, Celebes, Pandan Wangi, Pare Kembang, Rojo Lele, Cianjur, Mentik Wangi, dan Situ Patenggang. Padi aromatik merupakan bagian kecil yang istimewa dari kelompok padi karena memiliki mutu beras yang baik, yaitu beraroma dan tekstur nasi yang pulen, sangat disukai konsumen dan mempunyai harga yang tinggi (Bradbury et al. 2005b; Lang dan Buu 2008). Namun padi aromatik memiliki memiliki karakter agronomi yang kurang tahan terhadap hama dan penyakit, memerlukan kondisi lahan budidaya tertentu, waktu panen lama, dan produktivitas yang kurang tinggi (±4 ton/ha).
Padi Pandan Wangi (Gambar 1a) yang terkenal sejak tahun 1973 memiliki aroma wangi menyerupai pandan yang merupakan karakter paling menarik dari padi ini yang banyak diminati di pasaran dunia. Pandan Wangi mengandung kadar amilosa 7 – 20% dan amilopektin 80 – 93% yang menyebabkan padi varietas ini bertekstur pulen. Selain itu cocok tumbuh di suhu 25 – 30 °C dengan ketinggian 500 – 800 meter dari permukaan laut. Padi Mentik Wangi memiliki ciri bulat dan tahan rontok (Gambar 1b). Usia tanamnya mencapai 150 – 160 hari dan tingginya mencapai 150 cm. Kelemahan varietas ini diantaranya tingkat produktivitas rendah serta tidak tahan terhadap hama dan penyakit.
Varietas nonaromatik contohnya adalah Ciherang, IR64, Nipponbare, T309, Fatmawati, Situ Bagendit, Andel Rojo, Andel Lombok (beras merah) untuk tepung, dan
Andel Lombok (beras merah) untuk konsumsi langsung. Ciherang merupakan kelompok padi nonaromatik varietas unggul hasil beberapa kali persilangan, yaitu IR18349-53-1-3-1-3/IR19661-131-3-1//IR119661-131-3-1///IR64 ////IR64 (Bhattacharjee 2002). Padi Ciherang memiliki karakteristik umur tanamannya cukup singkat yaitu 116 hingga 125 hari, bentuk tanaman tegak, tingginya mencapai 107 sampai 115 cm, menghasilkan anakan produktif 14 sampai 17 batang, warna kaki hijau, warna batang hijau, warna daun hijau, muka daun kasar pada sebelah bawah, posisi daun tegak, bentuk gabah panjang ramping, warna gabah kuning bersih, tekstur nasi pulen, kadar amilosa 23%, bobot 1000 per butir 27 hingga 28 g, rata-rata produksi 5 - 8,5 ton/ha, tahan terhadap bakteri hawar daun (HDB) strain III dan IV, tahan terhadap wereng coklat biotipe 2 dan 3 (Gambar 1c). Padi Ciherang dilepas oleh menteri pertanian pada tahun 2000 dengan anjuran cocok ditanam pada musim hujan dan kemarau dengan ketinggian di bawah 500 m di atas permukaan laut (Hermanto 2006).
Introgresi karakter aroma dari padi varietas Pandan Wangi atau Mentik Wangi ke varietas Ciherang diharapkan dapat menghasilkan varietas dengan karakter agronomi yang hampir sama dengan karakter tanaman pemulih Ciherang (tahan hama dan penyakit, dapat dibudidayakan di berbagai lokasi di Indonesia, waktu panen yang tidak lama, dan produktivitasnya tinggi) ditambah dengan karakter aroma dari donor Pandan Wangi atau Mentik Wangi.
[a] [b]
[image:34.595.330.502.510.739.2][c]
Aroma Padi
Aroma padi disebabkan oleh senyawa kimia yang mudah menguap. Hasil penelitian menunjukkan terdapat lebih dari 114 senyawa terdapat pada padi aromatik. Namun, senyawa utama yang menyebabkan aroma wangi pada padi adalah 2-asetil-1-pirolin (2AP) (Itani 2004). Akumulasi dari 2AP dalam genotip padi aromatik dapat disebabkan oleh adanya mutasi delesi pada ekson 7 di kromosom nomor 8 yang mengakibatkan kodon stop sehingga menyebabkan hilangnya aktivitas enzim betain aldehida dehidrogenase (BADH2) ketika prolina mensintesis asam amino glutamat. Enzim BADH2 memainkan peranan kunci dalam jalur konversi ke arah glutamat. Penghambatan lintasan ini akan meningkatkan ketersediaan prolina untuk sintesis 2AP (Bradbury et al. 2005a,b). Berbeda dengan padi nonaromatik, pada kromosom nomor 8 tidak terjadi delesi ekson 7 sehingga prolina lebih mengarah ke pembentukan asam amino glutamat dan pembentukan 2AP lebih sedikit.
Padi aromatik mengandung senyawa 2AP lebih tinggi (0,04–0,07 ppm) dibandingkan padi nonaromatik (0.004–0.006 ppm) (Adijono et al. 1993). Hal yang sama dikemukakan oleh Buttery et al. (1983) bahwa kandungan senyawa 2AP padi aromatik mencapai 0.04–0.09 ppm, sepuluh kali jauh lebih tinggi dibanding padi nonaromatik yang hanya 0.004–0.006 ppm. Senyawa 2AP sebenarnya terdeteksi di semua bagian tanaman padi, kecuali di akar (Lorieux et al.
1996).
Biosintesis jalur reaksi yang rinci tentang kompleks ini belum sepenuhnya dijelaskan (Bradbury et al. 2005a). Namun, telah ditemukan bahwa prekursor dan sumber nitrogen 2AP pada varietas aromatik Thai Hom Mali adalah senyawa prolina (Yoshihashi et al. 2002). Diduga jalur pembentukan 2AP seperti pada gambar 2. Jalur pembentukan 2AP dimulai dari pemecahan prolina menjadi putresin kemudian membentuk senyawa gama aminobutiraldehid (GABald), substrat dari enzim BADH2. Jika enzim BADH2 aktif, maka enzim ini dapat mengubah GABald menjadi asam gama-aminobutirat (GABA). Tetapi jika enzim BADH2 tidak aktif, GABald mengalami asetilasi (penambahan gugus asetil) membentuk 2AP. Putresin akan ditemukan dalam jumlah tinggi pada jaringan yang tumbuh aktif membelah. Putresin dipecah menjadi GABald oleh diamina oksidase (DAO) selama proses pembentukan
lignin dan dinding sel, setelah sebagian besar pembelahan sel telah terjadi. Oleh karena itu, pembentukan GABald cenderung terjadi di jaringan muda yang secara aktif membelah dan dinding sel menjadi kaku.
Ujung daun padi mengandung 2AP dalam jumlah yang lebih tinggi daripada di pangkal daun, sementara daun muda lebih beraroma daripada daun tua (Lorieux et al. 1996). Penurunan jumlah 2AP dalam jaringan yang lebih tua kemungkinan besar disebabkan oleh sifat volatil kimia. Pada tanaman, prolina berfungsi sebagai zat pelindung terhadap resiko kerusakan daun ketika tanaman terpapar oleh cekaman kekeringan atau terdapatnya kandungan garam yang tinggi di dalam tanah (Heldt 2005).
Aroma padi aromatik tidak hanya tercium pada saat pemasakan nasi. Se