MIKROENKAPSULASI BIOMASA Porphyridium cruentum
HASANAH
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR
BOGOR
RINGKASAN
HASANAH. C34060357. Mikroenkapsulasi Biomasa Porphyridium cruentum.
Dibimbing oleh IRIANI SETYANINGSIH dan PIPIH SUPTIJAH.
Mikroalga adalah mikroorganisme bersel satu yang umumnya memiliki pertumbuhan secara autotrof, menggunakan CO2 sebagai sumber karbon, dan
memerlukan cahaya untuk proses fotosintesis. Beberapa jenis mikroalga memiliki komponen aktif yang dapat dimanfaatkan dalam bidang industri pangan, kosmetik, pharmaceutical dan neutraceutical. Komponen aktif mikroalga antara lain fenol, terpenoid, sterol, flavonoid dan polisakarida. Mikroalga juga mengandung pigmen, tokoferol, EPA dan DHA. Salah satu mikroalga yang mempunyai komponen aktif tersebut adalah Porphyridium cruentum. Porphyridium cruentum memiliki komponen aktif dan pigmen (phycobilliprotein, karotenoid, klorofil). Komponen aktif dan pigmen yang terkandung pada mikroalga bersifat tidak stabil dan sensitif terhadap lingkungan. Salah satu teknologi yang dapat digunakan untuk melindungi kandungan zat padat, cair, atau gas dari kondisi yang tidak dapat dikontrol adalah mikroenkapsulasi.
Tujuan penelitian adalah mendapatkan kurva pertumbuhan Porphyridium cruentum, menentukan umur panen untuk menghasilkan pigmen fikoeritrin tertinggi, mempelajari pengaruh mikroenkapsulasi terhadap komposisi kimia, komponen aktif, pigmen fikoeritrin, dan aktivitas antioksidan.
Penelitian ini meliputi 3 tahapan, yaitu : (1) kultivasi dalam media Becker yang dilakukan pada suhu ruang (26-28oC), intensitas cahaya 2000-3000 lux, dan pH 7,31; (2) pemanenan biomasa; (3) mikroenkapsulasi biomasa Porphyridium cruentum menggunakan maltodekstrin. Analisis yang dilakukan antara lain perhitungan jumlah sel, analisis pigmen dengan spektrofotometer, analisis proksimat, analisis komponen aktif, analisis zat padat terlarut, dan analisis aktivitas antioksidan terhadap mikrokapsul.
MIKROENKAPSULASI BIOMASA Porphyridium cruentum
HASANAH
SKRIPSI
Sebagai salah satu syarat memperoleh gelar Sarjana Perikanan pada
Departemen Teknologi Hasil Perairan
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Penelitian : Mikroenkapsulasi Biomasa Porphyridium cruentum Nama Mahasiswa : Hasanah
NRP : C34060357
Menyetujui
Pembimbing I Pembimbing II
Dr.Ir. Iriani Setyaningsih, MS Dra. Pipih Suptijah, MBA (NIP. 19600925 198601 2 001) (NIP. 19531020 198503 2 001)
Mengetahui
Ketua Departemen Teknologi Hasil Perairan
Dr. Ir. Ruddy Suwandi, MS, M.Phil (NIP. 19580511 198503 1 002)
Pernyataan Mengenai Skripsi dan Sumber informasi
Dengan ini saya menyatakan bahwa skripsi “Mikroenkapsulasi Biomasa Porphyridium cruentum” adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun ke perguruan tinggi manapun. Sumber infomasi yang berasal dari atau kutipan dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir skripsi ini.
Bogor, Maret 2011
KATA PENGANTAR
Puji syukur kita panjatkan kepada Allah SWT atas segala rahmat dan karunia serta kelancaran selama penelitian sehingga penulis dapat menyelesaikan skripsi yang berjudul “Mikroenkapsulasi Biomasa Porphyridium cruentum”.
Penulisan skripsi ini mendapat banyak dukungan dari berbagai pihak.
Pada kesempatan kali ini penulis mengucapkan terima kasih yang sebesar -besarnya kepada pihak yang telah bersedia membantu, di antaranya adalah :
1. Dr. Ir. Iriani Setyaningsih MS dan Dra. Pipih Suptijah, MBA sebagai dosen pembimbing yang tidak henti-hentinya memberikan masukan, motivasi, dan semangat kepada penulis sehingga skripsi ini dapat diselesaikan dengan baik.
2. Desniar, SPi, MSi sebagai dosen penguji yang memberikan masukan dan saran sehingga skripsi ini dapat diselesaikan dengan baik.
3. Ir. Winarti Zahiruddin, MS sebagai dosen pembimbing akademik yang memberikan bimbingan, saran, dan motivasi kepada penulis.
4. Dr. Ir. Ruddy Suwandi, MS. M. Phil sebagai Ketua Departemen Teknologi Hasil Perairan.
5. Dr. Ir. Agoes M. Jacoeb, Dipl Biol sebagai komisi pendidikan Departemen Teknologi Hasil Perairan.
6. Orang tua, saudara (Kak Yuli, Kak Lastri dan Iyan) serta seluruh keluarga yang berada di Jambi atas segala doa dan dukungannya selama menyelesaikan studi di IPB.
7. Dosen, staf THP, Laboran (Ibu Ema, Mas Ipul, Mb Lastri, Mb Silvi, Mb Sherlyn, Mb Siti, Mb Ina) atas bantuan dan kerjasamanya.
8. Tim Mikroalga (Dwi, Dina, Ely, dan Budi), Septin, Tyas, Fatma, Ratih, Putri, Rapita, Caca, An, THP 43, together, kost Ramadhan serta kepada seluruh pihak yang tidak dapat disebutkan satu persatu. Terima kasih atas segala bantuan dan dukungannya.
DAFTAR RIWAYAT HIDUP
Penulis memiliki nama lengkap Hasanah, dilahirkan di Jambi, 8 Juni 1988. Penulis merupakan anak ketiga dari empat bersaudara dari pasangan Bapak Gunarto dan Ibu Paijah. Penulis mengawali pendidikan formal tahun 1994 di SD N 97/IV Kota Jambi dan Madrasah Ibtidaiyah, Kotabaru, Jambi. Kemudian pada tahun 2000 melanjutkan pendidikan ke SLTP N 14 Kota Jambi dan SMA N 3 Kota Jambi pada tahun 2003. Pada tahun 2006, penulis diterima di Institut Pertanian Bogor melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB). Pada tahun 2007 diterima pada Mayor Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
Selama menjadi mahasiswa, penulis aktif dalam organisasi dan kepanitian. Organisasi yang pernah diikuti penulis antara lain: Ikatan Keluarga Muslim TPB (IKMT) periode 2006/2007, Forum Scientist for Studies (FORCES) periode 2006/2007, LDK Al-Hurriyah periode 2006/2008, Bendahara HIMASILKAN tahun 2007, KAMMI periode 2007/2008, Senior Resident Asrama TPB IPB periode 2008/2010, Blue policy School (BPS) angkatan I periode 2008/2009. Penulis juga aktif sebagai asisten praktikum m.k. Mikrobiologi Hasil Perairan tahun 2010/2011 dan asisten m.k Pendidikan Agama Islam tahun 2011. Selain itu penulis juga aktif dalam kepanitiaan berbagai kegiatan mahasiswa dan penulisan karya ilmiah (PKM) serta Kompetisi Karya Tulis Mahasiswa (KKTM) di Institut Pertanian Bogor. Selama masa studinya, penulis juga mendapatkan beasiswa POM, BBM, dan Pemerintah Daerah Jambi.
DAFTAR ISI ... ix
2.1 Deskripsi dan Klasifikasi Porphyridium cruentum ... 3
2.2 Pertumbuhan Mikroalga ... 4
2.3 Pengaruh Faktor Lingkungan Porphyridium cruentum ... 5
2.4 Mikroenkapsulasi ... 8
3.3.1 Kultivasi mikroalga Porphyridium cruentum ... 24
3.3.2 Pemanenan biomasa ... 25
3.3.3 Mikroenkapsulasi dengan metode spray dryer ... 25
3.4 Prosedur Analisis ... 25
3.4.1 Perhitungan jumlah sel ... 25
3.4.2 Laju pertumbuhan Porphyridium cruentum ... 26
3.4.6 Analisis proksimat ... 28
3.4.7 Analisis aktivitas antioksidan dengan metode DPPH ... 31
3.4.8 Analisis zat padat terlarut ... 32
4 HASIL DAN PEMBAHASAN ... 33
4.1 Kultivasi dan Pertumbuhan Porphyridium cruentum ... 33
4.2 Pigmen Fikoeritrin ... 39
4.3 Mikrokapsul Porphyridium cruentum ... 41
4.3.1 Zat padat terlarut mikrokapsul Porphyridium cruentum ... 41
4.3.2 Komposisi kimia mikrokapsul Porphyridium cruentum ... 42
4.3.3 Komponen aktif Porphyridium cruentum ... 44
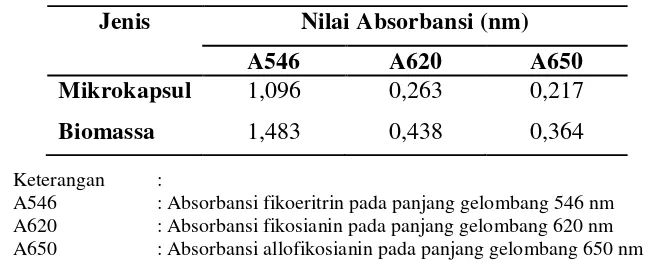
4.3.4 Pigmen fikoeritrin mikrokapsul ... 47
4.3.5 Antioksidan ... 48
5 KESIMPULAN DAN SARAN ... 50
5.1 Kesimpulan ... 50
5.2 Saran ... . 50
DAFTAR PUSTAKA... ... 51
1 Perkembangan tehnik mikroenkapsulasi di dunia ... 9
2 Dua jenis struktur utama mikrokapsul ... 9
3 Diagram alir proses mikroenkapsulasi ... 12
4 Struktur phycoerytrobillin ... 13
5 Pigmen fikoeritrin a) sel Porphyridium cruentum b) struktur ... phycobillisome dalam sel Porphyridium cruentum ... 14
6 Reaksi penangkapan radikal bebas DPPH oleh antioksidan ... 22
7 Diagram alir tahapan proses penelitian ... 24
8 Kondisi kultivasi ... 33
9 Sel Porphyridium cruentum secara a) mikroskop b) SEM ... 36
10 Kurva pertumbuhan Porphyridium cruentum ... 37
11 Kultur Porphyridium cruentum umur 0,5, 8, dan 12 hari... 39
12 Pigmen Porphyridium cruentum H-5, H-8, H-12 ... 39
13 Peak pigmen Porphyridium cruentum a) aquades b) buffer ... 40
14 Produk mikrokapsul Porphyridium cruentum ... 41
1 Laju pertumbuhan Porphyridium cruentum ... 38
2 Nilai absorbansi pigmen Porphyridium cruentum ... 40
3 Komposisi kimia mikrokapsul ... 42
4 Komponen aktif Porphyridium cruentum ... . 45
Mikroalga adalah mikroorganisme bersel satu yang umumnya memiliki pertumbuhan secara autotrof, menggunakan CO2 sebagai sumber karbon, dan
cahaya untuk fotosintesis. Mikroalga dapat menambah nilai gizi pada makanan dan mempunyai pengaruh yang positif terhadap kesehatan manusia (Spolaore et al. 2006). Biomassa mikroalga kaya nutrien antara lain asam lemak omega 3 dan 6, asam amino esensial (leusin, isoleusin, valin, dan lain-lain), dan karoten (Fuentes et al. 2000). Beberapa jenis mikroalga juga memiliki kandungan protein yang tinggi. Asam amino pada mikroalga lebih baik jika dibandingkan dengan sumber protein makanan yang lain. Mikroalga juga memiliki kandungan karbohidrat dalam bentuk pati, glukosa, gula, dan polisakarida lain. Beberapa keunggulan lain dari mikroalga tidak tergantung pada iklim dan cuaca, waktu tumbuh cepat sehingga dapat dipanen dalam waktu yang tidak terlalu lama, dapat diproduksi terus-menerus, tidak dapat menyebabkan dampak buruk bagi lingkungan, serta produksinya dapat dikendalikan sesuai dengan kebutuhan dan keinginan, serta aman bagi kesehatan (Arylza 2005).
Mikroalga memiliki komponen aktif yang dimanfaatkan dalam bidang industri pangan, kosmetik, pharmaceutical dan neutraceutical. Komponen aktif mikroalga antara lain fenol, terpenoid, sterol, flavonoid dan polisakarida. Selain itu, mikroalga juga mengandung pigmen (klorofil, phycobillinsome, karoten) tokoperol, EPA dan DHA (El-Baky et al. 2008). Komponen aktif mikroalga mempunyai aktivitas antimikroba (Abedin dan Taha 2008); antitumor dan antimikroba (Taskin et al. 2010); dan aktivitas antioksidan (Marxen et al. 2007).
padat, cair, atau gas dari kondisi yang tidak dapat dikontrol adalah mikroenkapsulasi. Mikroenkapsulasi dapat digunakan untuk melindungi komponen aktif yang terdapat dalam suatu bahan. Penelitian Ferreira et al. (2007) menunjukkan bahwa mikroenkapsulasi digunakan untuk melindungi komponen aktif berupa antioksidan dari kondisi lingkungan, sehingga antioksidan alami dapat digunakan dalam industri pangan, kosmetik, pharmaceutical, dan neutraceutical. Namun penelitian tentang mikroenkapsulasi dari Porphyridium cruentum belum dilakukan, oleh karena itu perlu dilakukan kajian mikroenkapsulasi pada biomassa Porphyridium cruentum.
1.2 Tujuan
Tujuan dari penelitian, antara lain :
1) Mendapatkan kurva pertumbuhan Porphyridium cruentum yang dikultivasi dengan media Becker.
2) Menentukan umur panen Porphyridium cruentum untuk menghasilkan pigmen fikoeritrin tertinggi.
3) Mempelajari pengaruh mikroenkapsulasi terhadap komposisi kimia, komponen aktif, pigmen fikoeritrin, dan aktivitas antioksidan.
2 TINJAUAN PUSTAKA
2.1 Deskripsi dan Klasifikasi Porphyridium cruentum
Porphyridium cruentum adalah mikroalga merah bersel satu yang termasuk kelas Rhodophyceae, hidup bebas atau berkoloni yang terikat dalam mucilago. Senyawa mucilago dieksresikan secara konstan oleh sel membentuk sebuah kapsul yang mengelilingi sel. Mucilago merupakan polisakarida sulfat yang bersifat larut dalam air (Borowitzka dan Borowitzka 1988). Klasifikasi Porphyridium cruentum (Vonshak 1988) adalah sebagai berikut :
Divisi : Rhodophyta Subkelas : Bangiophycidae Ordo : Porphyridiales Famili : Porphyridiaceae Genus : Porphyridium
Species : Porphyridium cruentum
Pemberian nama alga merah untuk Porphyridium cruentum didasarkan atas kelebihan dan dominasi dari pigmen merah r(red)-fikoeritrin dan r(red) -fikosianin yang dimilikinya. Jenis klorofil yang dimilikinya adalah klorofil a sedangkan klorofil b tidak ada dan diganti dengan klorofil d (Sharma 1986). Kumar dan Shingh (1979) menyatakan bahwa pigmen merah menutupi warna dari pigmen fotosintesis lainnya. Pigmen r-fikoeritrin, r-fikosianin, dan alllofikosianin terkandung dalam fikobillin dari alga merah (Sharma 1986). Fikobillin berperan penting dalam fotosintesis sebagai pigmen penerima cahaya terutama pada fotosistem II (PSII) dalam phycobillisome (Arylza 2005).
Komposisi biomassa Porphyridium cruentum yaitu 32,1% (w/w) karbohidrat dan 34,1% protein kasar. Kandungan mineral dalam 100 g biomassa kering : Ca (4960 mg), K (1190 mg), Na (1130 mg), Mg (629 mg), Zn (373 mg). Kandungan asam lemak terdiri dari 1,6% untuk16:0; 0,4% untuk 18:2ώ6; 1,3%, 20:4ώ6; 1,3% untuk 20:5ώ3. Biomassa mengandung pigmen Porphyridium cruentum berupa fikoeritrin dengan karakteristiknya berwarna merah. Biomassa juga mengandung tokoferol, vitamin K, dan karoten (Fuentes et al. 2000).
Struktur sel Porphyridium cruentum merupakan tipe struktur sel eukariotik. Setiap sel dikelilingi oleh dinding sel yang terdiri dari dua lapisan. Lapisan bagian luar terdiri dari bahan pectic dan lapisan bagian dalam terbuat dari cellulosic microfibrils (Sharma 1986). Biomassa kering sel Porphyridium
cruentum mengandung protein 28-39%, karbohidrat 40-57%, lipid 9-14% (Spolaore et al. 2006).
2.2 Pertumbuhan Mikroalga
Pertumbuhan mikroalga pada kultur dapat ditandai dengan bertambah besarnya ukuran sel atau bertambah banyaknya jumlah sel. Perkembangan sel dalam kultur mikroalga terdiri atas lima fase, yaitu fase lag (adaptasi), fase eksponensial (logaritmik), fase penurunan laju pertumbuhan (deklinasi), fase stasioner, dan fase kematian. Fase pertama adalah fase lag atau adaptasi dimana pada fase ini populasi yang baru ditransfer mengalami penurunan tingkat metabolisme karena fase inokulum yang tidak merata dan terjadi proses adaptasi terhadap media kultur. Fase kedua adalah fase eksponensial (logaritmik) dimana percepatan pertumbuhan dan perbandingan konsentrasi komponen biokimia menjadi konstan (Fogg 1975).
2.3 Pengaruh Faktor Lingkungan Porphyridium cruentum
Pertumbuhan Porphyridium cruentum dipengaruhi oleh faktor lingkungan meeliputi cahaya, suhu, salinitas, pH, dan nutrien yang digunakan (karbon, nitrogen, sulfur, dan fosfor).
a) Cahaya
Cahaya memiliki pengaruh yang besar terhadap komposisi kimia alga fotosintesis. Umumnya penurunan intensitas cahaya akan meningkatkan klorofil a dan pigmen yang lain (klorofil b, klorofil c, fikobilliprotein, dan karotenoid) sedangkan intensitas cahaya yang tinggi akan menurunkan klorofil a dan pigmen yang lain (Richmond 2004).
Pertumbuhan Porphyridium cruentum pada air laut juga tergantung cahaya tetapi toleransi intensitas cahayanya relatif lebih besar. Peningkatan intensitas cahaya menyebabkan pengecilan ukuran kloroplas dan meningkatkan granulasi pada kandungan sel (Borowitzka dan Borowitzka 1988).
b) Suhu
Suhu merupakan salah satu faktor yang berpengaruh terhadap komposisi biokimia alga. Suhu berpengaruh terhadap komposisi dan kandungan membran lipid. Menurunnya suhu pertumbuhan di bawah kondisi optimal meningkatkan asam lemak tak jenuh dalam sistem membran. Bertambahnya stabilitas dan fluiditas membran sel khusunya membran tilakoid (meningkatkan asam lemak tak jenuh pada membran lipid) melindungi fotosintesis dari fotoinhibisi pada suhu rendah (Richmond 2004).
Sel Porphyridium dapat tumbuh pada kisaran suhu 10-35 °C dengan suhu optimum 25 °C. Aktivitas optimum fotosintesis dari kultur Porphyridium cruentum terjadi pada suhu 25 °C (Vonshak 1988). Suhu optimum untuk pertumbuhan Porphyridium adalah 21-26 °C dan pada suhu dibawah 13 °C pertumbuhannya lambat sedangkan pada suhu diatas 31 °C pertumbuhannya terhambat (Richmond 1988).
c) Salinitas dan pH
hidup dengan mikroalga lainnya jika ditumbuhkan pada kultur terbuka. Salinitas sebesar 4,6% tidak menghambat proses pertumbuhan. Meskipun demikian, salinitas dengan kisaran 3,5-4,5% dapat memacu pertumbuhan yang optimal (Richmond 1988).
Porphyridium cruentum dapat tumbuh dengan baik pada kisaran pH 5,2-8,3. Derajat keasaman (pH) optimum untuk fotosintesis Porphyridium cruentum adalah 7,5. Pertumbuhan akan terhambat jika pH kurang dari 5 (Borowitzka dan Borowitzka 1988).
d) Nutrien
1) Sumber Karbon
Porphyridium menggunakan CO2 sebagai sumber karbon (Vonshak 1988).
Pertumbuhan menjadi lebih cepat pada kultur yang diberi cahaya dan aerasi dengan udara yang mengandung CO2. Pertumbuhan kultur pada 4400 lux dengan
aerasi mempunyai waktu pembelahan 20 jam sedangkan ketika ditambahkan udara sebesar 1% CO2 (v/v) waktu pembelahan menjadi setengahnya (Borowitzka
dan Borowitzka 1988).
2) Sumber Nitrogen
Porphyridium dapat menggunakan KNO3 dan amonium sebagai sumber
nitrogen. Sumber nitrogen diperlukan untuk pertumbuhan sel dan sintesis enzim pada pembentukan polisakarida. Kelebihan nitrogen umumnya mengurangi produksi polisakarida ekstraseluler. Jika ditumbuhkan dalam media dengan sumber nitrogen terbatas maka produksi polisakaridanya lebih tinggi (Borowitzka dan Borowitzka 1988).
Nitrogen diperlukan sel dalam jumlah besar sebagai komponen protein, asam nukleat dan komponen seluler yang lain. Nitrogen pada umumnya digunakan oleh organisme dalam bentuk nitrat (NO3-) dan amonium (NH4+).
3) Sulfur
Sulfur merupakan elemen essensial untuk autotrof dan heterotrof. Sulfur memainkan peranan penting pada struktur dan fungsi protein. L-metionin, L-cistein, glutationin dan sulfide bukan termasuk sumber sulfur bagi P.cruentum. Sulfur yang berasal dari MgSO4, Na2SO3, atau Na2S2O3 antara 5,4 sampai 27 Mm
menghasilkan pertumbuhan yang baik bagi mikroalga (Borowitzka dan Borowitzka 1988).
4) Fosfor
Fosfor merupakan makronutrien utama lainnya yang berperan penting pada proses metabolit seluler dengan membentuk banyak struktur dan komponen fungsional untuk pertumbuhan dan perkembangan mikroalga. Ketika nutrien
berlebih maka cahaya merupakan faktor pembatas pertumbuhan (Richmond 2004). Fosfor dipergunakan oleh alga dalam bentuk H2PO4- atau
HPO42-. Konsentrasi fosfor optimum dalam media bervariasi berdasarkan spesies.
Rata-rata kisaran toleransi fosfor untuk kebanyakan alga adalah 50 µg/l sampai 20 µg/l (Becker 1994).
Fosfor mengalami penurunan sama dengan nitrogen. Pada sel eukariotik dan prokariotik, kandungan klorofil a menurun ketika kandungan karbohidrat meningkat. Namun, berkebalikan dengan nitrogen, kerusakan phycobillisome sedikit ketika fosfor berkurang. Penurunan phycobillisome menyebabkan pembelahan sel dan terjadi sintesis phycobillisome baru (Richmond 2004).
Fosfor berperan sebagai elemen struktural asam nukleat, adenosine tifosfat dan fosfolipid, serta metabolisme energi terutama untuk menghasilkan ATP pada proses fotosintesis (Raynods 1994). Fosfor merupakan salah satu unsur yang
berperan dalam proses penyusunan karbohidrat dan senyawa nitrogen (Bold dan Wynne 1985). Fosfat dapat menurunkan klorofil dalam sel. Gula
terfosforilasi yang kaya energi muncul dalam proses fotosintesis (Lombardi dan Wangersky 1991).
Se) dalam air hanya terbatas untuk pertumbuhan alga. Chelator merupakan trace metal bufer.
2.4 Mikroenkapsulasi
Mikroenkapsulasi merupakan suatu tehnik dimana cairan, padatan, atau gas dilapisi dengan lapisan tipis berupa material pelindung. Lapisan tipis melindungi dari kebusukan, mengurangi penguapan komponen aktif, dan menghindari dari kondisi yang tidak diinginkan (Selim et al. 2008). Tujuan utama umum mikroenkapsulasi adalah untuk membuat bahan cairan bersifat seperti padatan. Hal ini menyebabkan beberapa sifat bahan inti menjadi berubah, misalnya sifat aliran bahan dan penangan bahan menjadi lebih mudah dalam bentuk padatan. Bahan yang memiliki higroskopis dapat dilindungi dari kelembaban lingkungan. Selain melindungi zat aktif, proses ini juga bermanfaat untuk menutupi rasa, aroma ataupun yang tidak diinginkan dari bahan aktif. Kestabilan dari bahan yang mudah menguap, sensitif terhadap cahaya, oksidasi atau panas dapat dipertahankan.Hal penting lain dalam proses mikroenkapsulasi bahan makanan adalah juga untuk mengatur pelepasan bahan aktif pada waktu yang dikehendaki. Bahan-bahan yang berhubungan dengan makanan yang dienkapsulasi meliputi asam, pewarna, enzim, mikroorganime, perasa, lemak dan minyak, vitamin dan mineral, garam, pemanis dan gas (Paramita 2010)
Gambar 1 Perkembangan tehnik mikroenkapsulasi di dunia (Gouin 2004)
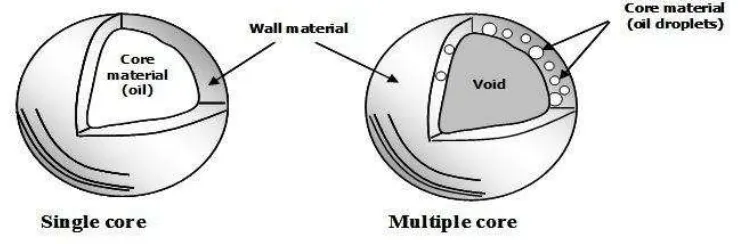
Pengelompokan kapsul berdasarkan pada ukuran partikel > 5000 μm (makro), 1,0-5000 μm (mikro) dan < 1,0 μm (nano).Mikrokapsul dapat berbentuk bola, persegi panjang ataupun tak beraturan. Dua jenis struktur utama dari mikrokapsul adalah satu inti (single core) dan banyak inti (multiple core) pada bagian dindingnya. Struktur utama dari mikrokapsul dapat dilihat pada Gambar 2.
Gambar 2 Dua jenis struktur utama mikrokapsul (Jafari et al. 2008)
Mikrokapsul dengan satu inti biasanya diproduksi dengan cara coacervation, droplet co-extrusion dan pemasukan molekul. Model ini biasanya memiliki muatan inti yang tinggi, misalnya 90% dari total berat mikrokapsul. Mikrokapsul dengan struktur banyak inti di bagian dinding umumnya diproduksi menggunakan spray drying. Bahan inti tersebar secara merata di bagian dinding dan bagian
tengah mikrokapsul biasanya berupa rongga kosong yang dihasilkan dari pemuaian selama tahap-tahap pengeringan akhir. Biasanya, struktur ini memiliki persentasi pelapis hingga 70% dari berat mikrokapsul (Paramita 2010).
Bahan di dalam mikrokapsul disebut sebagai inti, fasa internal, atau pengisi. Bahan inti dapat berupa emulsi, bahan kristalin, suspensi padatan, atapun gas. Isi dalam mikrokapsul dilepaskan dengan berbagai macam mekanisme. Pelapis dapat rusak secara mekanik, misalnya akibat dikunyah, meleleh ketika terekspos dengan panas, terlarut dalam solvent (pelarut). Perubahan pH dapat mengubah kemampuan proses penembusan bahan aktif sehingga mengendalikan pelepasan. Pelapis dari lemak (lipid) dapat terdegradasi akibat enzim lipase dan bahan aktif berdifusi ke lingkungan. Sifat fisik dan kimia dari bahan aktif (seperti kelarutan, difusivitas, tekanan uap, dan koefisien partisi) dan pelapis (seperti ketebalan, porositas dan kemampuan bereaksi) juga mempengaruhi pelepasan bahan aktif (Paramita 2010).
Bahan pelapis yang disebut juga sebagai kulit, dinding, atau membran, dapat berasal dari film-forming (pembuat lapisan tipis) polimer natural atau sintesis. Memilih pelapis harus berdasarkan pada sifat kimia maupun fisik bahan aktif, juga proses yang digunakan untuk membuat mikrokapsul. Bahan pelapis harus tidak larut dan tidak bereaksi terhadap zat aktif. Umumnya, polimer yang tidak larut dalam air digunakan untuk membuat mikrokapsul dengan bahan aktif seperti air, dan polimer yang dapat larut air digunakan untuk mikrokapsul pada bahan aktif organik. Untuk meningkatkan kualitas lapisan, lapisan dibuat beberapa lapis, memiliki sifat yang seperti plastik, cross-linking, juga ada perlakuan pada permukaannya. Ketebalan lapisan dimanipulasi untuk meningkatkan permeabilitas dan stabilitas dari mikrokapsul (Paramita 2010).
Proses mikroenkapsulasi secara umum terjadi melalui tiga tahap dalam suatu pengadukan yang kontinyu. Ketiga tahap tersebut (Nugroho 2009), yaitu :
a) Bentuk tiga fase kimia yang belum saling bercampur, yaitu fase pembawa (air), fase material inti yang akan dilapisi, dan fase penyalut.
c) Pemadatan pelapis untuk membentuk mikrokapsul yang biasanya terjadi akibat adanya panas.
Proses mikroenkapsulasi dari komponen inti terdiri dari dua tahap utama. Pertama, emulsifikasi dari materi inti dengan larutan dindingnya seperti karbohidrat dan protein. Kedua, pengeringan atau pendinginan emulsi tersebut. Secara sistematis proses mikroenkapsulasi dapat dilihat pada Gambar 3.
Mikroenkapsulasi dengan spray drying merupakan salah satu metode yang umum digunakan untuk enkapsulasi ingredien makanan karena lebih ekonomis penggunaannya dalam industri makanan. Mikroenkapsulasi digunakan untuk menghasilkan dry powder, granula atau butiran. Pada proses mikroenkapsulasi komponen aktif dikelilingi oleh matrik pelindung yang umumnya berupa polimer.
Polimer itu antara lain gum, maltodekstrin, pati, dan sebagainya (Parize et al. 2008).
Beberapa bahan yang digunakan sebagai penyalut seperti pati, gum arab, metilselulosa, gelatin, whey protein, sirup jagung, β-cyclodekstrin, maltodekstrin, disakarida, pulunan, dan sodium kasein. Mikroenkapsulasi diaplikasikan dalam proses kimia dan industri makanan untuk melindungi aroma dan oleoresin, vitamin, warna, dan enzim. Enkapsulasi warna lebih mudah untuk dilakukan dan meningkatkan stabilitas (Selim et al. 2008).
Maltodekstrin merupakan jenis karbohidrat hasil dari turunan hidrolisis sebagian pati (jagung atau kentang) dan digunakan dalam pangan sebagai bahan pengisi untuk meningkatkan tekstur dan cita rasa. Struktur maltodekstrin, yaitu (C6H12O5)n H2O merupakan produk hidrolisat pati (polimer sakarida tidak manis)
berwarna putih, dan tidak berbau sehingga dapat digunakan dalam berbagai aplikasi produk yang luas (Nugroho 2009).
Gambar 3 Diagram alir proses mikroenkapsulasi (Nugroho 2009)
2.5 Pigmen
Semua organisme fotosintesis mengandung pigmen organik yang digunakan untuk menghasilkan energi. Ada tiga kelas utama pigmen yaitu klorofil, karatenoid, dan phycobilin. Klorofil (pigmen hijau) dan karatenoid (pigmen kuning atau orange) bersifat lipofilik dan berasosiasi dengan Chl-protein komplek, sedangkan phycobilin bersifat hidrofilik (Richmond 2004).
Billin adalah tetrapyrol terbuka yang mengandung struktur sistem cincin skeleton. Billin berbentuk lebar, meskipun tidak teratur, dan berada pada kingdom tanaman dan hewan. Pada hewan, billin ada pada sebagian besar katabolit fungsional haem meskipun dalam beberapa spesies mereka memainkan peranan
Materi dinding Bahan inti
Emulsi
Rasio bahan inti dan material dinding
Proses kimia Proses mekanika
Koaservasi Kokristalisasi Inklusi molekuler Polimerisasi interfasial
Spray drying Spray chilling Ekstrusi Fluidized bed
Partikel mikro (mikrokapsul)
yang kecil, prinsipnya sebagai kulit atau eggshell pigmen. Selain itu, pada billin tumbuhan menunjukkan peranan penting sebagai penangkap cahaya dan sensor cahaya. Billin tumbuhan berbeda dari billin hewan pada umumnya karena kovalent nya menumpuk pada apoprotein membentuk konjugasi yang disebut billiprotein ( Goodwin 1976).
Alga protein (phycobilliprotein) adalah pigmen yang terjadi pada divisi Rhodophyta (alga merah), Cyanophyta (alga hijau-biru), Cryptophyta (alga cryptomonad). Mereka berfungsi sebagai penyerap cahaya pada fotosinteis. Alga billiprotein dapat diekstrak dengan air ataupun larutan garam. Alga billiprotein memiliki titik isoelektrik antara 4,3-4,9 dan akan mengkristal bila tidak berada dalam range pH tersebut (Goodwin 1976).
Pada alga merah, pigmen utamanya berupa phycobilin (phycoerythrobilin, phycocyanobilin, dan phycourobilin) yang berbentuk tetrapyrolles linear dan tidak berikatan dengan atom magnesium. Phycobillin menyerap cahaya biru-hijau, hijjau, kuning atau orange (500-650 nm) Phycobilliprotein bersifat larut air dan pigmen berikatan kovalen dengan apoprotein (Richmond 2004). Struktur phycoerythrobilin dapat dilihat pada Gambar 4.
Pigmen r-fikoeritrin, r-fikosianin dan allofikosianin, terkandung dalam fikobillin dari alga merah. Pigmen-pigmen tersebut merupakan jenis pigmen larut air. Beberapa famili Rhodophyceae tidak memperlihatkan warna merah karena fikoeritrinnya rusak disebabkan oleh cahaya atau photo destruction (Sharma 1986).
Gambar 4 Struktur phycoerythrobillin
Salah satu alga merah yang memiliki pigmen fikoeritrin adalah Porphyridium cruentum. Fikoeritrin dapat digunakan sebagai pewarna alami untuk menggantikan pewarna sintesis yang merupakan penyebab karsinogen. Fikoeritrin mempunyai potensial pasar yang luas karena permintaan pigmen yang tinggi terhadap jenis pigmen merah yang aman bagi kesehatan (Borowitzka dan Borowitzka 1988). Pigmen fikoeritrin dalam sel mikroalga dapat dilihat pada Gambar 5.
(a) (b)
Gambar 5 Pigmen fikoeritrin a) sel Porphyridium cruentum b) struktur phycobillisome dalam sel Porphyridium cruentum
(Barsanti dan Gualteri 2006)
2.6 Komponen Aktif
Komponen aktif merupakan kelompok senyawa fungsional yang terkandung dalam bahan pangan dan dapat memberikan pengaruh biologis. Sebagian besar komponen aktif adalah kelompok alkohol aromatik seperti polifenol dan komponen asam (phenolic acid). Komponen aktif tidak terbatas pada hasil metabolisme sekunder saja, tetapi juga termasuk metabolit primer yang memberikan aktivitas biologis fungsional, seperti protein dan peptida (Kannan et al. 2009).
kesehatan atau memiliki peran aktif dalam pencegahan penyakit (metabolit sekunder) (Astawan dan Kasih 2008). Metode uji fitokimia meliputi komponen karbohidrat, gula pereduksi, peptida, asam amino (metabolit primer), alkaloid, steroid, flavonoid, saponin, dan fenol hidrokuinon (metabolit sekunder) (Harborne 1987).
2.6.1 Alkaloid
Alkaloid berasal dari dua suku kata yaitu alkali yang berarti basa dan oid yang berarti mirip sehingga alkaloida adalah senyawa yang mengandung nitrogen bersifat basa dan mempunyai aktivitas farmakologis. Komponen alkaloid merupakan golongan senyawa organik yang paling banyak ditemukan di alam. Alkaloid merupakan substansi dasar yang memiliki satu atau lebih atom nitrogen yang bersifat basa dan tergabung dalam suatu sistem siklis yaitu cincin heterosiklik. Alkaloid biasanya tidak berwarna dan sebagian besar berbentuk kristal amorf atau cairan pada suhu ruang. Secara organoleptik, alkaloid akan terasa pahit di lidah. Fungsi alkaloid pada tumbuhan diperkirakan sebagai pelindung tumbuhan dari serangan hama dan penyakit, pengatur tumbuh, atau sebagai basa mineral untuk mempertahankan keseimbangan ion (Putra 2007). Alkaloid ada yang bersifat racun pada manusia, tetapi ada pula yang memiliki aktivitas fisiologis pada kesehatan manusia sehingga digunakan secara luas dalam pengobatan (Harborne 1987).
2.6.2 Steroid/triterpenoid
Triterpenoid merupakan komponen dengan kerangka karbon yang tersusun oleh 6 unit isoprene dan dibuat secara sintesis dari skualen (C30 hidrokarbon
asiklik). Triterpenoid memiliki struktur siklik yang kompleks, sebagian besar terdiri atas alkohol, aldehid, atau asam karboksilat. Triterpenoid tidak berwarna, jernih, memiliki titik lebur tinggi dan merupakan komponen aktif yang sulit dikarakterisasi. Pengujian yang telah digunakan secara luas untuk mendeteksi triterpenoid adalah dengan pereaksi Liebermann-Buchard, yang memberikan warna biru-hijau pada triterpenoid dan steroid. Triterpenoid dapat digolongkan menjadi 4 grup komponen, yaitu triterpenoid sebenarnya, steroid, saponin dan cardiac glycoside. Triterpenoid umumnya meiliki rasa yang pahit
Produk triterpenoid alami memiliki aktivitas antitumor karena memiliki kemampuan menghambat kinerja enzim topoisomerase II, dengan cara berikatan dengan sisi aktif enzim yang nantinya akan mengikat DNA dan membelahnya. Hal ini menyebabkan enzim menjadi terkunci dan tidak dapat mengikat DNA (Setzer 2008).
2.6.3 Flavonoid
Flavonoid merupakan golongan terbesar dari senyawa polifenol, karena itu larutan ekstrak yang mengandung flavonoid akan berubah warna jika diberi larutan basa atau amonia. Flavonoid mengandung sistem aromatik konjugasi dan dapat menunjukkan pita penyerapan yang kuat pada spektrum wilayah UV dan sinar tampak. Flavonoid umumnya merupakan komponen larut air (polar). Komponen ini dapat diekstrak dengan etanol 70% dan tertinggal pada lapisan aqueous, diikuti dengan pemisahan ekstrak menggunakan petroleum eter. Flavonoid pada tanaman berikatan dengan gula sebagai glikosida dan adapula yang berada dalam aglikon. Flavonoid dapat dikelompokkan menjadi 9 kelas, yaitu antosianin, proantosianidin, flavonol, flavon, glikoflavon, biflavonil, chlacone dan aurone, flavanon, serta isoflavon (Harborne 1987).
2.6.4 Saponin
Saponin merupakan glikosida yang apabila dihidrolisis secara sempurna akan menghasilkan gula dan satu fraksi non-gula yang disebut sapogenin atau genin. Gula-gula yang terdapat dalam saponin jumlah dan jenisnya bervariasi di antaranya glukosa, galaktosa, arabinosa, ramnosa, serta asam galakturonat dan glukoronat. Saponin merupakan agen aktif permukaan dengan sifat yang menyerupai sabun. Saponin larut dalam air, sedikit larut atau tidak sama sekali dalam etanol dan metanol pekat yang dingin. Kehadirannya dapat dideteksi dengan mudah karena komponen ini mampu membentuk busa (Muchtadi 1989).
Komponen saponin berperan dalam mereduksi kolestrol dan melawan kanker kolon. Saponin juga memiliki aktivitas antimikroba, merangsang sistem imun, dan mengatur tekanan darah (Astawan dan Kasih 2008).
2.6.5 Fenol hidrokuinon
Komponen fenolat merupakan struktur aromatik yang berikatan dengan satu atau lebih gugus hidroksil, beberapa mungkin digantikan dengan gugus metal atau glikosil. Komponen fenolat bersifat larut air selama komponen tersebut berikatan dengan gula membentuk glikosida, dan biasanya terdapat dalam vakuola sel. Flavonoid merupakan kelompok terbesar di antara komponen fenolat alami yang strukturnya telah diketahui, tetapi fenol monosiklik sederhana, fenilpropanoid dan fenolat quinon terdapat dalam jumlah sedikit (Harborne 1987). Pigmen quinon alami berada pada kisaran warna kuning muda hingga hitam. Quinon mengandung kromatofor dasar yang sama dengan kromatofor benzoquinon, yang terdiri dari dua grup karbonil yang berkonjungasi dengan dua ikatan rangkap karbon-karbon. Untuk tujuan identifikasi, quinon dapat dibagi menjadi empat kelompok, yaitu benzoquinon, naftaquinon, antraquinon, dan isoprenoid quinon. Tiga kelompok pertama umunya terhidrolisis dan memiliki sifat fenol, sedangkan isoprenoid quinon terdapat pada respirasi seluler (ubiquinon) dan fotosintesis (plastoquinon) (Harborne 1987).
2.6.6 Karbohidrat
Karbohidrat merupakan komponen organik kompleks yang dibentuk melalui proses fotosintesis pada tanaman, dan merupakan sumber energi utama dalam respirasi. Karbohidrat berperan dalam penyimpanan energi (pati), transportasi energi (sukrosa), serta pembangun dinding sel (selulosa). Karbohidrat berperan dalam interaksi hewan dan tumbuhan, perlindungan dari luka dan infeksi, serta detoksifikasi dari substansi asing. Karbohidrat yang memiliki berat molekul rendah, umumnya mempunyai banyak kegunaan. Komponen tersebut aktif secara optis, merupakan komponen alifatik polihidroksi, yang biasanya sangat larut air. Komponen ini sukar untuk mengkristal bahkan dalam keadaan murni sekalipun, dan biasanya diisolasi dengan mereaksikannya dengan komponen lain (Harborne 1987).
Karbohidrat umumnya dapat dikelompokkan menjadi tiga kelompok berdasarkan ukuran molekulnya, yaitu monosakarida, oligosakarida, dan polisakarida. Monosakarida merupakan suatu molekul yang dapat terdiri dari
5-6 atom C (fruktosa, glukosa), oligosakarida merupakan polimer dari 2-10 monosakarida (sukrosa), dan polisakarida merupakan polimer yang terdiri
dari 10 monomer monosakarida yang membentuk rantai lurus ataupun bercabang (Harborne 1987).
2.6.7 Gula pereduksi
Gula pereduksi merupakan kelompok gula atau karbohidrat yang mampu mereduksi senyawa pengoksidasi. Monosakarida akan segera mereduksi senyawa-senyawa pengoksida seperti ferisianida, hidrogen peroksida, atau ion kupri (Cu2+). Gula dioksidasi pada gugus karbonil dan senyawa pengoksidasi menjadi tereduksi pada reaksi ini. Sifat gula pereduksi ini dapat berguna dalam analisis gula, dengan mengukur jumlah dari senyawa pengoksidasi yang tereduksi oleh suatu larutan gula tertentu, dapat dilakukan pendugaan konsentrasi gula. Prinsip tersebut berguna dalam menganalisis kandungan gula dalam darah dan air (Lehninger 1988).
pada fruktosa (ketosa) terletak pada karbon nomor dua. Sukrosa tidak mempunyai gugus OH bebas yang reaktif karena keduanya saling terikat, sedangkan laktosa mempunyai gugus OH bebas pada atom C nomor satu pada gugus glukosanya (Winarno 2008).
Salah satu metode untuk mengukur jumlah gula pereduksi ini dapat digunakan larutan Fehling. Larutan Fehling merupakan larutan alkali tembaga (II) yang mengoksidasi aldosa menjadi aldonat, dalam prosesnya akan tereduksi menjadi tembaga (I) yang mengendap sebagai Cu2O yang berwarna merah bata.
Reagen lain yang dapat digunakan dalam mengukur jumlah gula pereduksi adalah reagen Nelson-Somogyi dan Benedict. Aldosa termasuk gula pereduksi, begitu juga ketosa. Ketosa dalam suasana alkali larutan Fehling akan terisomerasi menjadi aldosa, sehingga dalam reagen Benedict yang tidak alkali hanya komponen aldosa yang dapat terdeteksi, tetapi ketosa tidak (Fennema 1996).
2.7 Antioksidan
vitamin C, vitamin E dan betakaroten serta senyawa fenolik (Andayani et al. 2008).
Antioksidan adalah senyawa kimia yang dapat menyumbangkan satu atau lebih elektron kepada radikal bebas, sehingga radikal bebas tersebut dapat diredam. Antioksidan bereaksi dengan radikal bebas dengan cara mengurangi konsentrasi oksigen, mencegah pembentukan singlet oksigen yang reaktif, mencegah inisiasi rantai pertama dengan menangkap radikal primer seperti radikal hidroksil, mengikat katalis ion logam, mendekomposisi produk-produk primer radikal menjadi senyawa non-radikal, dan memutus rantai hidroperoksida. Antioksidan berdasarkan mekanisme kerjanya dikelompokan menjadi (Mardawati et al. 2008):
1) Antioksidan primer yaitu antioksidan yang bereaksi dengan radikal lipid berenergi tinggi untuk menghasilkan produk yang memiliki kestabilan termodinamis lebih baik. Antioksidan golongan fenol seperti isoflavon termasuk dalam antioksidan yang memiliki mekanisme ini.
2) Antioksidan sekunder yang juga dikenal dengan antioksidan pencegah (preventive antioxidant) yang dapat memperlambat reaksi inisiasi dengan cara memutus rantai (chain-breaking antioxidant) hidroperoksida. Contoh antioksidan ini yaitu dilauril thiodipropionate dan asam thiodipropionic. Antioksidan golongan ini adalah antioksidan yang berikatan dengan gugus thiol.
Mekanisme kerja antioksidan senyawa fenolik adalah sebagai berikut :
Radikal lipid antioksidan radikal antioksidan
ROO· + AH ROOH + A·
RO· + AH ROH + A·
R· + AH RH + A·
OH· + AH H2O + A·
isomer valensi. Peningkatan jumlah gugus hidroksil (alkil hidrogen pada struktur kimianya) pada posisi para atau ortho seperti pada genistein dapat meningkatkan aktivitas antioksidan isoflavon. Reaksi radikal bebas dengan komponen sel baik komponen struktural (molekul penyusun membran) maupun komponen fungsional yaitu enzim dan DNA dapat merusak sel melalui oksidasi lemak tidak jenuh dan protein sel. Kerusakan lebih lanjut pada organel sel dapat mencapai kerusakan DNA dan membran sel. Berdasarkan mekanisme tersebut, radikal bebas tentunya akan turut mempengaruhi akan timbulnya berbagai jenis penyakit degeneratif seperti aterosklerosis (pengendapan lemak yang mengeras dalam pembuluh darah arteri) (Mardawati et al. 2008).
Mikroalga mempunyai kemampuan menetralkan radikal bebas melalui mekanisme enzimatis dan non enzimatis. Karotenoid dan asam lemak termasuk kelompok non enzimatis yang mampu melindungi organisme dari kerusakan oksidasi. Tokoferol, flavonoid, alkaloid termasuk kelompok enzimatis. Mekanisme enzimatis dan non enzimatis sama-sama menurunkan radikal hanya berbeda pada kecepatan dan kecenderungan reaksinya. Antioksidan menetralkan radikal bebas dengan mendonorkan atom hidrogennya (Marxen et al. 2007).
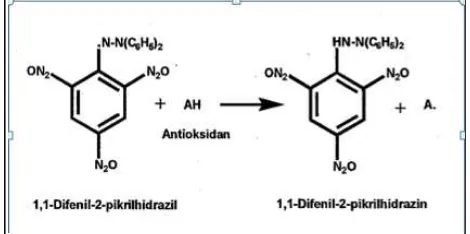
Metode uji DPPH merupakan salah satu metode yang paling banyak digunakan untuk memperkirakan efisiensi kinerja dari substansi yang berperan sebagai antioksidan (Molyneux 2004). Metode pengujian ini berdasarkan pada kemampuan substansi antioksidan tersebut dalam menetralisir radikal bebas. Radikal bebas yang digunakan adalah 1,1-diphenyl-2-picrylhydrazyl (DPPH) yang memiliki rumus molekul C18H12N5O6 dan Mr=394,33 (Vattem dan Shetty 2006).
mengandung antioksidan maka akan terjadi reaksi penangkapan hidrogen dari antioksidan oleh radikal bebas DPPH (ungu) yang kemudian berubah menjadi 1,1-difenil-2-pikrilhidrazin (kuning) (Artanti dan Hanafi 2008). Reaksi penangkapan radikal bebas DPPH oleh antioksidan dapat dilihat pada Gambar 6.
3 METODE PENELITIAN
3.1 Waktu dan Tempat
Penelitian dilaksanakan pada bulan Februari 2010-Januari 2011 di Laboratorium Bioteknologi Hasil Perairan 2, Laboratorium Mikrobiologi Hasil Perairan, dan Laboratorium Biokimia Hasil Perairan, Departemen Teknologi Hasil Perairan, Laboratorium Terpadu Fakultas Kedokteran Hewan, Laboratorium Fisika Fakultas Matematika dan Ilmu Pengetahuan, Institut Pertanian Bogor dan Pusat Studi Biofarmaka.
3.2 Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian adalah mikroalga Porphyridium cruentum yang diperoleh dari Laboratorium Marikultur Pusat
Penelitian Oseanografi-LIPI Ancol, Jakarta Utara; air laut, media Becker (NaCl, MgSO4.7H2O, MgCl2.6H2O, CaCl2.2H2O, KNO3, KH2PO4, NaHCO3, Tris HCl,
trace element, chelated iron yang dapat dilihat pada Lampiran 1, alkohol; aquades untuk ekstraksi pigmen; maltodekstrin untuk mikroenkapsulasi ; DPPH, metanol untuk uji aktivitas antioksidan, asam sulfat, pereaksi Dragendorff, pereaksi Meyer, pereaksi Wagner, kloroform, anhidra asetat, serbuk magnesium, amil alkohol, HCl, etanol 70 %, FeCl3 5%, pereaksi Molisch, asam sulfat pekat,
pereaksi benedict, pereaksi biuret, larutan ninhidrin 0,1 %, n-heksana, H2SO4
pekat, NaOH, H3BO3, selenium, indikator bromcherosol green-methyl red.
Alat yang digunakan dalam penelitian ini adalah toples kaca ukuran 3L, tabung reaksi, UV, sentrifus, lampu neon, aerator, selang, mikroskop, haemasitometer, pipet volumetrik, spektrofotometer, freeze dryer, spray dryer, baterai, timbangan, vorteks, hot plate, UV, gelas ukur, tisu, aluminium foil, kertas label, spidol, sudip, cawan, lemari es, timbangan, sudip, sentrifus dingin, mikropipet 10-100 µm, bulb, lux meter, labu Kjeldahl, oven, kertas Whatman No. 42, erlemenyer, gelas piala.
3.3 Prosedur Kerja
Gambar 7 Diagram alir tahapan proses penelitian
3.3.1 Kultivasi mikroalga Porphyridium cruentum
Penyegaran stok mikroalga perlu dilakukan sebelum dikultivasi. Penyegaran dilakukan selama tujuh hari dalam elemenyer dan media yang digunakan adalah media Becker. Volume kultur untuk penyegaran adalah 500 ml dan mikroalga yang dimasukkan sebanyak 20 % V/V. Penyegaran stok mikroalga dilakukan dalam keadaan aseptik pada suhu ruang di bawah penyinaran lampu
Biomassa basah 400 g Biomassa basah 400 g
kaca dari 100 ml, 500 ml, 2 L. Selanjutnya, dilakukan scale up 3 L untuk menentukan kurva pertumbuhan.
Selama kultivasi dilakukan pengambilan sampel setiap hari pada pagi hari sebanyak 1 ml. Selanjutnya sampel digunakan untuk menghitung kepadatan sel mikroalga.
3.3.2 Pemanenan biomassa
Pemanenan dilakukan pada umur 5 hari mewakili fase log, 8 hari mewakili fase awal stasioner, dan 12 hari mewakili fase akhir stasioner. Selanjutnya biomassa yang diperoleh diekstraksi pigmen fikoeritrin. Waktu panen yang menghasilkan pigmen fikoeritrin terbesar digunakan untuk panen pada kultivasi selanjutnya.
3.3.3 Mikroenkapsulasi dengan metode spray dryer (Dityanawarman et al. 2009)
Biomassa basah sebanyak 400 gr dicampurkan dengan maltodekstrin 15%. Kemudian ditambahkan aquades 200 ml dan dihomogenkan. Pengeringan biomassa basah dilakukan dengan spray dryer suhu inlet dan outlet diatur masing -masing pada suhu 180 ± 5 °C dan 80 ± 5°C.
3.4 Prosedur Analisis
Analisis yang dilakukan, antara lain perhitungan sel, laju pertumbuhan, analisis pigmen fikoeritrin biomassa dan mikrokapsul dengan menggunakan spektrofotometer, analisis komponen aktif, proksimat, dan aktivitas antioksidan terhadap mikrokapsul.
3.4.1 Perhitungan jumlah sel (Hadioetomo 1993)
Pertumbuhan Porphyridium cruentum diamati dengan cara mengambil sampel setiap hari menggunakan mikro pipet, kemudian dimasukkan ke dalam chamber hemasitometer, selanjutnya dihitung jumlah sel secara langsung menggunakan mikroskop. Hasil perhitungan nilainya dikonversikan ke dalam nilai logaritmik dan dibuat kurva pertumbuhan dengan jumlah sel (logaritmik) sebagai sumbu y dan waktu (hari) sebagai sumbu x. Proses perhitungan jumlah sel ini dengan metode hitung langsung sebagai berikut :
2) Tutup kaca hemasitometer diletakkan pada permukaan hemasitometer. Suspensi biakan Porphyridium cruentum hasil pengambilan sampel dikocok, kemudian diambil dengan mikropipet sekitar 20 µL. Suspensi tersebut diteteskan pada tempat menaruh sampel yang terdapat pada hemasitometer hingga suspensi Porphyridium cruentum menyebar pada ruang hitung.
3) Hemasitometer diletakkan di atas pentas mikroskop. Jumlah sel yang terdapat dalam 80 kotak kecil yang terletak dalam bagian tengah yang mempunyai luas 0,2 mm2 (5 x 16 x 0,0025 mm2) dihitung dengan mikroskop pada pembesaran 400x. Perhitungan jumlah sel dilakukan sebanyak 3 kali ulangan.
Formulasi yang dipakai dalam menghitung kepadatan sel adalah sebagai berikut :
N1 : jumlah sel dalam 80 kotak kecil (ulangan ke-1) N2 : jumlah sel dalam 80 kotak kecil (ulangan ke-2) 0,2 mm : luas hemasitometer dalam 80 kotak
0,1 mm : kedalaman cairan di bawah hemasitometer
Hasil perhitungan diplotkan pada grafik hingga diperoleh kurva pertumbuhan dengan umur kultur (hari) sebagai sumbu x dan log kepadatan sel (sel/mL) sebagai sumbu y.
3.4.2Laju pertumbuhan Porphyridium cruentum (Chrismadha et al. 1997)
Laju pertumbuhan mikroalga diperoleh dari data jumlah kepadatan sel selama kultivasi mikroalga. Rumus laju pertumbuhan mikroalga adalah sebagai berikut :
3.4.3 Analisis pigmen fikoeritrin biomassa
Metode yang digunakan dalam ekstraksi pigmen adalah berdasarkan metode dari Techernov et al (1993). Sampel sebanyak 80 mg masing-masing umur panen 5, 8, dan 12 hari dilarutkan ke dalam aquades 10 ml. Selanjutnya kocok menggunakan vortex dan disimpan di dalam refrigerator selama 24 jam. Sampel kemudian di sentrifugasi selama 30 menit 3000 rpm. Supernatan yang dihasilkan kemudian dianalisis dengan menggunakan spektrofotometer
3.4.4 Analisis pigmen fikoeritrin mikrokapsul (Dityanawarman et al. 2009)
Ekstraksi pigmen dilakukan dengan cara sebagai berikut : sebanyak 1 g produk mikrokapsul dan biomassa Porphyridium cruentum yang telah dikeringkan dilarutkan dalam aquades sebanyak 12,5 ml. Sampel kemudian dikocok menggunakan vortek sampai biomassanya larut dan disimpan pada refrigerator selama 8 hari. Suspensi sel kemudian disentrifugasi selama 30 menit 3000 rpm. Supernatan yang dihasilkan kemudian dianalisis dengan menggunakan spektrofotometer pada panjang gelombang 546 nm, 620 nm, dan 650 nm.
3.4.5 Analisis komponen aktif (Harborne 1987)
Analisis fitokimia dilakukan untuk mengetahui ada tidaknya komponen -komponen aktif yang terdapat pada Porphyridium cruentum yang memiliki
aktivitas antioksidan. Analisis fitokimia meliputi uji alkaloid, steroid/triterpenoid, flavonoid, saponin, fenol hidrokiunon, Molisch, Benedict, Biuret, dan Ninhidrin. 1)Alkaloid
Sejumlah sampel dilarutkan dalam beberapa tetes asam sulfat 2 N kemudian diuji dengan tiga pereaksi alkaloid yaitu, pereaksi Dragendorff, pereaksi Meyer, dan pereaksi Wagner. Hasil uji dinyatakan positif bila dengan pereaksi Meyer terbentuk endapan putih kekuningan, endapan coklat dengan pereaksi Wagner, dan endapan merah hingga jingga dengan pereaksi Dragendroff. 2)Steroid/triterpenoid
3) Flavonoid
Sejumlah sampel ditambahkan serbuk magnesium 0,10 mg dan 0,40 ml amil alkohol (campuran asam klorida 37 % dan etanol 95 % dengan volume yang sama) dan 4 ml alkohol kemudian campuran dikocok. Warna merah, kuning, atau jingga yang terbentuk pada lapisan amil alkohol menunjukkan adanya flavonoid. 4) Saponin (uji busa)
Saponin dapat dideteksi dengan uji busa dalam air panas. Busa yang stabil selama 30 menit dan tidak hilang pada penambhan 1 tetes HCl 2 N menunjukkan adanya saponin.
5) Fenol hidrokuinon (pereaksi FeCl3)
Sebanyak 1 gram sampel diekstrak dengan 20 ml etanol 70 %. Larutan asam sulfat pekat melalui dinding tabung. Uji positif menunjukkan adanya karbohidrat ditandai terbentuknya kompleks berwarna ungu di antara 2 lapisan cairan
7) Uji Benedict
Larutan sampel sebanyak 8 tetes dimasukkan ke dalam 5 ml pereaksi Benedict. Campuran dikocok dan dididihkan selama 5 menit. Warna hijau, kuning, atau endapan merah bata yang terbentuk menunjukkan adanya gula pereduksi.
3.4.6 Analisis proksimat
Analisis proksimat merupakan suatu analisis yang dilakukan untuk memprediksi komposisi kimia suatu bahan, termasuk di dalamnya analisis kadar air, lemak, protein, dan abu.
1) Analisis kadar air (AOAC 2005)
selanjutnya ditimbang beratnya sampai diperoleh berat cawan yang konstan (XI). Sampel ditimbang sebanyak 2-3 g (A), kemudian dimasukkan ke dalam cawan. Cawan yang berisi sampel diletakkan dalam oven selama 4-6 jam dengan suhu 105°C. Cawan tersebut dipindahkan dalam desikator dan setelah dingin ditimbang kembali sampai diperoleh berat yang konstan (X2). Kadar air dihitung dengan
menggunakan rumus :
X1 + A −X2
A X 100%
Keterangan :
X1 : berat cawan kosong setelah dikeringkan (g) X2 : berat cawan + sampel setelah dikeringkan (g) A : berat sampel (g)
2) Analisis kadar lemak (AOAC 2005)
Sampel sebesar 5 g (W1) dimasukkan ke dalam kertas saring pada kedua
ujung bungkus ditutup dengan kapas bebas lemak dan selanjutnya dimasukkan ke dalam selongsong lemak, kemudian sampel yang telah dibungkus dimasukkan ke dalam labu lemak yang sudah ditimbang berat tetapnya (W2) dan disambungkan
dengan tabung Soxhlet. Selongsong lemak dimasukkan ke dalam ruang ekstraktor tabung Soxhlet dan disiram dengan pelarut lemak (n-heksana p.a). Kemudian dilakukan refluks selama 6 jam. Pelarut lemak yang ada dalam labu lemak didestilasi hingga semua pelarut lemak menguap. Pada saat destilasi pelarut akan tertampung di ruang ekstraktor, pelarut dikeluarkan sehingga tidak kembali ke dalam labu lemak, selanjunya labu lemak dikeringkan dalam oven pada suhu 105°C, setelah itu labu didinginkan dalam desikator sampai beratnya konstan (W3). Perhitungan kadar lemak adalah sebagai berikut :
3) Analisis kadar abu (AOAC 2005)
Cawan pengabuan dikeringkan di dalam oven selama 1 jam pada suhu 105°C, kemudian didinginkan selama 15 menit di dalam desikator dan ditimbang hingga didapatkan berat yang konstan. Sampel sebanyak 5 g dimasukkan ke dalam cawan pengabuan dan dipijatkan di atas nyala api hingga tidak berasap lagi. Setelah itu, dimasukkan ke dalam tanur pengabuan dengan suhu 600°C selama 1 jam, kemudian ditimbang hingga didapatkan berat yang konstan. Kadar abu ditentukan dengan rumus :
Berat abu (g) = berat sampel dan cawan akhir (g) – berat cawan kosong (g) % kadar abu = berat abu (g)
berat sampel awal � 100%
4) Analisis kadar protein (SNI 01-2354.4-2006)
Tahap yang dilakukan untuk analisis kadar protein terdiri dari tahap destruksi, destilasi, dan titrasi.
a) Tahap destruksi
Sampel ditimbang sebanyak 0,5 g, kemudian dimasukkan ke dalam tabung Kjeldahl. Satu buah tablet Kjeldahl dimasukkan ke dalam tabung tersebut dan ditambahkan 10 ml H2SO4. Tabung yang berisi larutan tersebut dimasukkan
ke dalam alat pemanas dengan suhu 410˚C ditambahkan 10 ml air. Proses destruksi dilakukan sampai warna larutan menjadi bening.
b) Tahap destilasi
Proses destilasi terdiri dari 2 tahap yaitu :
Tahap pertama adalah tahap persiapan alat yaitu kran air dibuka dan dilakukan pengecekan alkali dan air dalam tangki, tabung, dan erlemenyer yang berisi akuades diletakkan pada tempatnya. Tombol power (pada Kjeldahl system) ditekan dan dilanjutkan dengan penekanan tombol stream dan ditunggu beberapa saat sampai air di dalam tabung mendidih. Steam dimatikan kemudian tabung Kjeldahl dan erlemenyer dikeluarkan dari alat Kjeldahl system.
yang sudah diberi asam borat. Destilasi dilakukan sampai volume larutan dalam erlemenyer mencapai 200 ml.
c) Tahap titrasi
Titrasi dilakukan dengan menggunakan HCl 0,1 N sampai warna larutan pada erlemenyer berubah warna menjadi pink, selanjutnya kadar protein dari sampel dapat diperoleh dengan perhitungan menggunakan :
Nitrogen % = HCl−ml blanko x 0,1 N HCl x 14,007
� � ℎ � 100%
kadar protein (%) = Nitrogen (%) x faktor konversi (6,25)
3.4.7 Analisis aktivitas antioksidan dengan metode DPPH (1,1-difenil-2- pikrilhidrazil) (Dityanawarman et al. 2009)
Analisis aktivitas antioksidan dengan DPPH diawali dengan menyiapkan stok BHT sebagai larutan kontrol positif dan larutan sampel produk dalam metanol p.a. Pada larutan stok BHT yang telah dibuat kemudian dilakukan pengenceran dalam metanol pro analysis dengan konsentrasi 6,25 ppm.
Pada larutan stok sampel dilakukan pengenceran dalam metanol p.a dengan konsentrasi 6,25 ppm. Pengenceran larutan BHT maupun sampel ditetapkan dalam larutan metanol p.a. Larutan DPPH yang akan digunakan dibuat dengan melarutkan kristal DPPH dalam pelarut methanol p.a dengan konsentrasi 1mM. Larutan sampel dan BHT masing-masing diambil 4,5 ml dan direaksikan dengan 500 µl larutan DPPH 1mM dalam tabung reaksi yang berbeda dan diberi label.
Masing-masing larutan kemudian diinkubasi dalam inkubator pada suhu 37°C yang sesuai dengan suhu normal tubuh manusia selama 30 menit agar DPPH dapat bereaksi. Pengukuran dilakukan dengan menggunakan spektrofotometer UV-Visisble (UV-Vis) pada panjang gelombang 517 nm.
Inhibisi (%) = [ − �] � 100% Keterangan ;
AB = absorbansi blanko
AS = absorbansi larutan standar atau sampel
3.4.8 Analisis zat padat terlarut, metode gravimetri (Purba 2003)
Satu gram bahan pengkapsul dilarutkan dalam 150 ml akuades dan disaring dengan menggunakan kertas saring Whatman No.42. Sebelum digunakan kertas saring terlebih dahulu dikeringkan dalam oven 105°C selama 30 menit dan ditimbang. Setelah penyaringan, kertas saring beserta residu dikeringkan dalam oven 105°C selama tiga jam, didinginkan dengan desikator dan ditimbang. Pengeringan dilakukan hingga diperoleh bobot yang konstan.
Kelarutan = 100 –100 − −%KAx c 100
x 100
Keterangan :
a = berat kertas saring dan residu (gram) b = berat kertas saring (gram)
4 HASIL DAN PEMBAHASAN
4.1 Kultivasi dan Pertumbuhan Porphyridium cruentum
Porphyridium cruentum merupakan mikroalga merah dari kelas rhodophyceae, bersel satu, mempunyai bentuk sel bulat dengan diameter 4 - 9 μm, hidup bebas atau berkoloni (Borowitzka dan Borowitzka 1988).
Kultivasi Porphyridium cruentum menggunakan intensitas cahaya antara 2000-3000 lux. Hal ini sesuai dengan penelitian Kusumawarni (1998) yang menunjukkan bahwa kandungan pigmen Porphyridium cruentum terbesar dengan menggunakan intensitas cahaya 2000 lux.
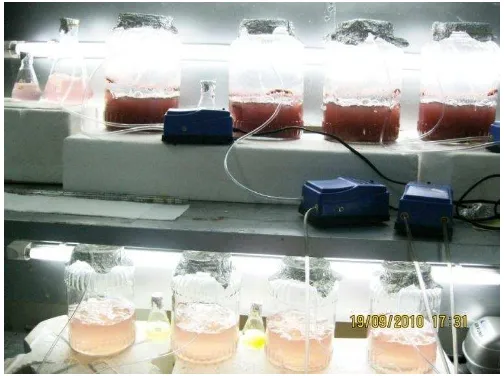
Suhu yang digunakan adalah suhu ruang sekitar 26-28 °C. Kelembaban udara sekitar 70-90 %. Suhu yang digunakan dalam penelitian ini masih berada dalam kisaran suhu pertumbuhan Porphyridium cruentum. Vonshak (1988) menyatakan bahwa sel Porphyridium dapat tumbuh pada kisaran suhu 10-35°C. Aktivitas optimum fotosintesis dari kultur Porphyridium cruentum terjadi pada suhu 25°C. Kondisi kultivasi di laboratorium dapat dilihat pada Gambar 8.
Gambar 8 Kondisi kultivasi Porphyridium cruentum
bahwa Porphyridium cruentum dapat tumbuh dengan baik pada kisaran pH 5,2-8,3. Derajat keasaman (pH) optimum untuk fotosintesis Porphyridium
cruentum yaitu 7,5. Pertumbuhan Porphyridium cruentum akan terhambat pada pH kurang dari 5.
Media kultivasi Porphyridium cruentum memiliki salinitas 6,08%. Salinitas ini dua kali lipat dengan salinitas air laut, namun Porphyridium cruentum tetap tumbuh dengan baik. Borowitzka dan Borowitzka (1988) menjelaskan bahwa Porphyridium dapat bertahan hidup pada kisaran salinitas yang cukup besar, yaitu 0,5-2 kali konsentrasi air laut (3,5%). Richmond (1988) menjelaskan bahwa salinitas media Becker Porphyridium cruentum pada kisaran 3,5-4,5% dapat memacu pertumbuhan yang optimal namun salinitas 4,6% tidak menghambat proses pertumbuhan, sedangkan pada kondisi salinitas kurang dari 3,5%, Porphyridium tidak mampu bersaing hidup dengan mikroalga lainnya, jika ditumbuhkan pada kultur terbuka.
Kultivasi Porphyridium cruentum menggunakan media Becker. Nutrien media Becker meliputi MgSO4.7H2O, MgCl2.6H2O, CaCl2.2H2O, KNO3, KH2PO4,
NaHCO3, Tris-HCl, Chelate iron, dan trace element. Ketersediaan MgSO4.7H2O
berfungsi menyediakan sulfur bagi mikroalga. Borowitzka dan Borowitzka (1988) menjelaskan sulfur memainkan peranan penting pada struktur dan fungsi protein. L-metionin, L-cistein, glutationin dan sulfida bukan termasuk sumber sulfur bagi Porphyridium cruentum. Sumber sulfur Porphyridium cruentum berasal dari MgSO4, Na2SO3, atau Na2S2O3 antara 5,4 sampai 27 Mm. Senyawa MgCl2.6H2O
berfungsi menyediakan magnesium bagi mikroalga. Becker (1994) menyatakan bahwa magnesium diperlukan oleh semua spesies mikroalga karena magnesium berperan pada fotosintesis sebagai atom pusat molekul klorofil. Magnesium juga berfungsi menyatukan ribosom menjadi unit fungsional. Kekurangan magnesium menyebabkan terhambatnya pembelahan sel sehingga sel abnormal. CaCl2.2H2O
Nutrien media Becker lainnya adalah KNO3 yang berfungsi menyediakan
nitrogen pada mikroalga. Nitrogen merupakan elemen terbesar yang penting bagi mikroalga. Borowitzka dan Borowitzka (1988) menjelaskan bahwa Porphyridium dapat menggunakan KNO3 dan amonium sebagai sumber nitrogen. Sumber
nitrogen diperlukan untuk pertumbuhan sel dan sintesis enzim pada pembentukan polisakarida. Kelebihan nitrogen umumnya mengurangi produksi polisakarida ekstraseluler. Jika ditumbuhkan dalam media dengan sumber nitrogen terbatas maka produksi polisakaridanya lebih tinggi. NaHCO3 dalam media Becker
berfungsi menyediakan karbon bagi mikroalga. Vonshak (1988) menyatakan Porphyridium menggunakan CO2 sebagai sumber karbon. Borowitzka dan
Borowitzka (1988) menjelaskan pertumbuhan menjadi lebih cepat pada kultur yang diberi cahaya dan aerasi dengan udara yang mengandung CO2. Pertumbuhan
kultur dengan aerasi mempunyai waktu pembelahan 20 jam, sedangkan ketika ditambahkan udara sebesar 1 % CO2 (v/v) waktu pembelahan menjadi
setengahnya.
Media Becker juga mengandung Tris-HCl dengan pH 7,6 yang berfungsi sebagai bufer. Barsanti dan Gualtieri (2006) menyatakan bahwa sistem bufer digunakan sebagai pengontrol pH pada media kultur. Selain itu, bufer juga mencegah terjadinya presipitasi.
Porphyridium cruentum juga menggunakan chelate iron sebagai media kulturnya. Chelate iron yang digunakan berupa Fe dan EDTA. Barsanti dan Gualtieri (2006) menjelaskan chelator berfungsi sebagai bufer trace metal untuk mengatur konsentrasi ion logam bebas. Ion logam bebas pada mikroalga bisa berfungsi sebagai nutrien ataupun toksik. Tidak semua chelate iron cocok digunakan, misalnya keberadaan Cu menyebabkan toksik sedangkan Fe dapat mencegah terjadinya presipitasi. Penambahan EDTA pada media kultur berfungsi untuk mencegah presipitasi terutama Ca dan Mg.
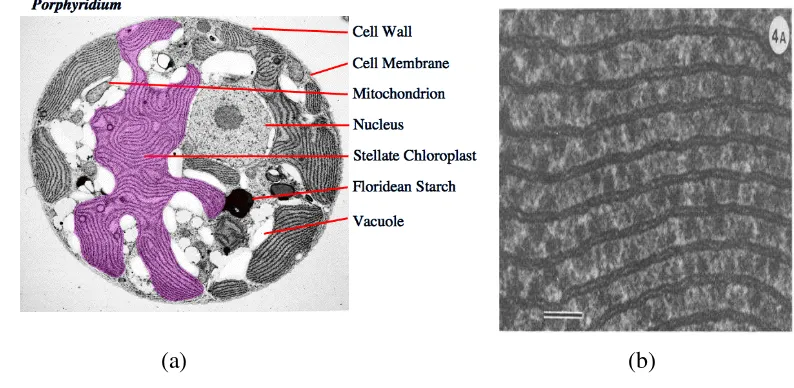
terbuat dari cellulosic microfibrils. Wanner dan Kost (1980) menyatakan bahwa sel Porphyridium cruentum terdiri dari kloroplas, vakuola, badan golgi, pati, pyrenoid, dan mitokondria. Sel Porphyridium cruentum dapat dilihat pada Gambar 9.
(a) (b)
Keterangan : C : Kloroplas G : Badan golgi L1 : Lapisan pertama L2 : Lapisan kedua M : Mitokondria P : Pyrenoid S : Pati V : Vakuola
Gambar 9 Sel Porphyridium cruentum secara a) mikroskop b) SEM (Wanner dan Kost 1980)
Pertumbuhan mikroalga pada kultur dapat ditandai dengan bertambah besarnya ukuran sel atau bertambah banyaknya jumlah sel. Penentuan hari panen pada kultivasi dilakukan berdasarkan kurva pertumbuhan yang diperoleh pada saat kultivasi. Kurva pertumbuhan Porphyridium cruentum dapat dilihat pada Gambar 10.
C
P
V G
M
S L1 L2
Gambar 10 Kurva pertumbuhan Porphyridium cruentum a) fase lag b) fase log c) fase stasioner
Porphyridium cruentum yang dikultivasi dengan media Becker memiliki fase lag (adaptasi) selama 1 hari. Fase lag terjadi sangat singkat karena inokulum yang ditambahkan berumur 7 hari dan berada pada fase eksponensial. Inokulum yang berada pada fase eksponensial akan lebih cepat beradaptasi terhadap media kultur dan mampu tumbuh serta membelah dengan cepat. Selain itu karena media inokulum (stok) dan media kultur yang digunakan sama-sama menggunakan media Becker. Prihantini et al. (2005) menyatakan bahwa salah satu faktor yang menentukan lamanya fase adaptasi adalah umur kultur yang digunakan sebagai inokulum. Fase adaptasi akan menjadi lebih singkat apabila sel-sel yang dinokulasikan berasal dari kultur yang berada dalam fase eksponensial. Inokulasi sejumlah sel mikroorganisme ke dalam media dan kondisi lingkungan yang sama seperti pada pemeliharaan kultur sebelumnya menyebabkan fase adaptasi tidak terlihat dan kultur lebih cepat memasuki fase eksponensial.
Fase eksponensial Porphyridium cruentum terjadi pada H-1 sampai H-7. Pada fase ini jumlah sel meningkat tajam karena sel telah beradaptasi dengan media yang baru sehingga metabolismenya berjalan dengan baik. Fogg (1975) menyatakan bahwa pada fase eksponensial (logaritmik) terjadi percepatan pertumbuhan dan perbandingan konsentrasi komponen biokimia menjadi konstan. Fase stasioner Porphyridium cruentum terjadi pada H-8 sampai H-12. Fase stasioner ini terjadi karena nutrien yang berada pada media kultur dan intensitas
Prihantini et al. (2007) menyatakan bahwa fase stasioner pada kultur mikroalga berkaitan dengan berkurangnya sejumlah besar nutrien dalam media dan akumulasi senyawa-senyawa beracun sisa metabolisme. Penurunan juga terjadi akibat berkurangnya intensitas cahaya yang diterima oleh sel.
Penentuan hari panen pada penelitian ini dilakukan berdasarkan fase pertumbuhan Porphyridium cruentum. Pemanenan Porphyridium cruentum dilakukan pada fase log, awal stasioner, dan akhir stasioner. Kultur fase log (H-5) mulai berwarna lebih merah jika dibandingkan kultur pada H-0. Kultur pada fase awal stasioner (H-8) berwarna merah, lebih tua jika dibandingkan kultur H-5. Perubahan kultur yang semakin berwarna merah tua disebabkan kerapatan sel yang semakin tinggi. Hal ini ditunjukkan dengan laju pertumbuhan pada Tabel 1.
Tabel 1 Laju pertumbuhan Porphyridium cruentum
Laju pertumbuhan Jumlah (pembelahan sel/hari)
Hari ke-5 0,68
Hari ke-8 1,24
Hari ke-12 0,83
0 hari 5 hari 8 hari 12 hari Gambar 11 Kultur Porphyridium cruentum umur 0, 5, 8, dan 12 hari
4.2 Pigmen Fikoeritrin
Porphyridium cruentum memiliki pigmen dominan berupa fikoeritrin. Biomassa Porphyridium cruentum diekstraksi pigmennya pada umur panen 5 hari (fase log), 8 hari (fase awal stasioner), dan 12 hari (fase akhir stasioner). Pigmen Porphyridium cruentum dapat dilihat pada Gambar 12.
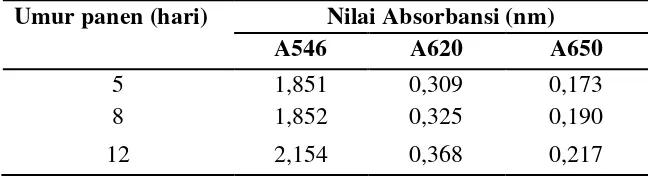
Gambar 12 Pigmen Porphyridium cruentum pada kultur H-12, H-8, dan H-5 Pigmen Porphyridium cruentum secara visual berwarna pink kemerah-merahan. Hal ini sesuai yang dinyatakan Bermejo et al. (2003) bahwa fikoeritrin murni mempunyai warna pink. Porphyridium cruentum diukur pigmennya pada panjang gelombang 546 nm, 620 nm, dan 650 nm. Nilai absorbansi Porphyridium cruentum dapat dilihat pada Tabel 2.
Absorbansi fikoeritrin, fikosianin, dan allofikosianin mengalami peningkatan warna dari kultur umur 5 hari, 8 hari, dan 12 hari. Pada kultur umur 12 hari (fase stasioner) mempunyai absorbansi fikoeritrin, fikosianin, dan allofikosianin tertinggi, yaitu sebesar 2,154 nm; 0,368 nm; 0,217 nm karena pada fase akhir stasioner sel lebih banyak menghasilkan metabolit sekunder, intensitas cahaya yang diterima sel berkurang dan ketersediaan nitrogen pada media kultur berkurang.