KEDELAI INDIA DAN ARGENTINA
PADA AYAM PEDAGING
HANY WIDJAJA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis berjudul Evaluasi Kinerja Enzim pada Bungkil Kacang Kedelai India dan Argentina pada Ayam Pedaging adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Maret 2013
Hany Widjaja
D152090051
___________________________________
dan Argentina pada Ayam Pedaging. Dibimbing oleh NAHROWI dan DESIANTO BUDI UTOMO.
Penelitian telah dilakukan dengan menggunakan ayam pedaging jantan dari strain komersial untuk mengevaluasi efek dari enzim pada penampilan produksi ayam, kecernaan asam amino, kecernaan serat kasar, kandungan air ekskreta dan nilai ekonomis pada ransum yang menggunakan bungkil kacang kedelai (BKK) India dan Argentina.
Penelitian menggunakan 8 perlakuan dengan 10 ulangan dan berjumlah 5 ekor setiap ulangan. Penlitian menggunakan dua ransum dasar yang berdasarkan pada jagung dan kacang kedelai diformulasikan dengan BKK India atau Argentina dengan level metabolisme energi dan asam amino yang sama, yakni 1% lisina tercerna, 0.38% metionina tercerna dan 0.75% metionina dan sisteina tercerna. Ransum dalam penelitian ini tidak menggunakan bahan baku hewani untuk memaksimalkan penggunaan BKK. Bahan baku utama lainnya yang diikutsertakan dalam formulasi pakan adalah jagung, minyak kelapa sawit, garam, vitamin dan mineral. Masing-masing ransum dasar (basal diet) dibagi menjadi 4 bagian, bagian pertama diberikan kepada ayam sebagai kontrol, kedua ditambahkan enzim proteinase, ketiga ditambahkan enzim karbohidrasedan bagian terakhir ditambahkan kombinasi enzim proteinase dan karbohidrase. Enzim karbohidrase yang dipergunakan pada penelitian ini adalah - galaktosidase dengan aktifitas enzim 2500 Ga IAU/g dan mannanase dengan aktifitas enzim 5000 MIAU/g. Enzim proteinase mempunyai aktifitas enzim sebesar 75,000 PROT units/g. Dosis untuk enzim proteinase dan karbohidrase masing-masing sebesar 500 g/ T and 200 g/T. Ayam diberi ransum tunggal dari umur 1 hingga 32 hari. Penelitian menggunakan rancangan acak lengkap faktorial (RAL Faktorial). Hasil dianalisis dengan ANOVA dan dilanjutkan dengan uji Duncan.
Hasil penelitian menunjukkan bahwa kecernaan asam amino esensial dan bukan esensial ileum tidak dipengaruhi oleh BKK dan tidak berbeda nyata (P > 0.05) diantara perlakuan enzim yang diberikan.
Kecernaan serat kasar tidak dipengaruhi oleh BKK dan tidak berbeda nyata (P > 0.05) diantara perlakuan yang diberikan. Secara numerik, kecernaan serat kasar pada BKK India lebih tinggi dari pada BKK Argentina dan pemberian enzim khususnya kombinasi enzim proteinase dan karbohidrase memberikan peningkatan kecernaan serat kasar tertinggi.
Kandungan air ekskreta dan konsumsi air minum diantara ayam yang diberi ransum dengan BKK India tidak berbeda nyata (P > 0.05) dengan BKK Argentina. Penambahan enzim juga tidak memberikan perbedaan nyata diantara perlakuan yang diberikan.
Penambahan enzim proteinase dan karbohidrase tunggal tidak dapat menunjukkan perbedaan yang nyata pada penampilan produksi ayam. Hal ini dapat disebabkan karena fungsi dari masing-masing enzim yang tidak optimal. Kombinasi kedua enzim memberikan perbedaan nyata, karena keduanya mempunyai mekanisme kerja sinergis. Enzim karbohidrase akan mendegradasi polisakarida bukan pati dan menurunkan tegangan permukaan sehingga dapat membuka akses bagi enzim proteinase dan enzim dalam tubuh ayam (endogenous) untuk menyempurnakan proses pencernaan.
Kesimpulan dari penelitian ini adalah bungkil kacang kedelai India menunjukkan penampilan produksi yang lebih rendah dari bungkil kacang kedelai Argentina. Kombinasi enzim proteinase dan karbohidrase menunjukkan efek sinergis. Keduanya dapat memperbaiki penampilan produksi dan meningkatkan nilai ekonomis pakan khususnya pada bungkil kacang kedelai India.
and India with and without enzyme supplementation in broiler. Supervised by NAHROWI and DESIANTO BUDI UTOMO.
A study was conducted with male chicks of a commercial broiler strain to evaluate the effects of commercial enzyme preparation on chick performance and digestibilities of amino acid and fiber in diets containing India and Argentina soybean meal.
Two corn-soy based diets were formulated to contain either soybean meal (SBM) India or SBM Argentina with same levels of metablizable energy (ME) and digestibile amino acids. Digestible lysine 1.0%, digestible methionine 0.38% and digestible methionine and cysteine 0.75% . No animal protein was used so as to allow for maximum usage SBM. Other major raw materials included in formulation were corn, crude palm oil (CPO), salt, vitamins and minerals. Each basal diet was split into 4 equal portions, one portion was fed as control and the other portions were supplemented with commercial single enzyme protease, carbohydrase or combination enzymes protease + carbohydrase. The carbohydrase enzyme used in these studies were galactosidase with 2500 Ga IAU/g enzyme activity and mannanase of 5000 MIAU/g enzyme activity. The protease enzyme activity was 75,000 PROT units/g. Dosages for single carbohydrase and protease enzyme were 500 g/ T and 200 g/T, respectively and dosage for combination enzyme was 700 g/T.
Birds were fed this single test feed from day 1 to day 32. Data collections were done for 32 days with 8 treatments, 10 replicates and 5 birds per replicate. Research using factorial completely randomized design. Results were analyzed by ANOVA followed by Duncan test
The result showed that performance on 32 days, body weight and FCR of birds fed India SBM diets were significantly different (P < 0.05) from birds fed Arg SBM diets. Supplementation of enzymes produced a significantly different (P < 0.05) results between single enzyme (carbohydrase or protease) and combination enzymes (carbohydrase and protease) for both body weight and feed conversion ratio (FCR); consequently combination enzymes improved the performance.
Ileal amino acid digestibility was not significantly affected by dietary treatment. Body weight and FCR showed positive correlation with amino acid digestibility, body weight positively correlated with Methionine (R2=0.406), FCR positively correlated with Methionine (R2=0.625) and Glycine (R2=0.412).
Fiber digestibility was not significantly affected by dietary treatment. Fiber digestibility of India SBM diets was numerically higher than Argentine SBM diets. Supplementation of combination carbohydrase and protease showed the best improvement in dietary fiber digestibility.
failed to show significant improvement in bird performance, this is possibly due to improper function of the enzymes. Since single protease may not easily access to some of the soybean proteins that are entrapped in the cell wall matrix without prior breakdown of the cell wall structure by carbohydrase. Results from this study indicated supplementation of single protease or carbohydrase to SBM based diets is hardly proven effective or beneficial. However, results from this study showed positive response of broiler performance, especially body weight gain and FCR, and fiber digestibility for birds fed SBM diets supplemented with combination enzymes in comparison with single enzyme. Combination of protease and carbohydrase possiblly have synergistic mode of action in terms of degrading NSP and entrapped protein in the cell wall metrix, which resulted in improvement in bird performance and numerically higher digestibility of fiber.
© Hak Cipta milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
PADA AYAM PEDAGING
HANY WIDJAJA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Nutrisi dan Pakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Evaluasi Kinerja Enzim pada Bungkil Kacang Kedelai India dan Argentina pada Ayam Pedaging
Nama : Hany Widjaja
NIM : D152090051
Disetujui oleh
Komisi Pembimbing
Prof Dr Ir Nahrowi, MSc Dr Desianto B. Utomo, PhD Ketua Anggota
Diketahui oleh
Ketua Program Studi Ilmu Nutrisi dan Pakan
Dr Ir Dwierra Evvyernie A, MS, MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Puji syukur penulis panjatkan kepada Tuhan Yesus Kristus dan Bunda Maria karena dengan kasih karuniaNya penulis dapat menyelesaikan tesis yang merupakan salah satu tugas akhir untuk memperoleh gelar Magíster Sains dalam program Pascasarjana Ilmu Nutrisi dan Pakan Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Prof Dr Ir Nahrowi, MSc selaku ketua komisi pembimbing yang selalu memberikan semangat, pengarahan dan kesempatan untuk mengembangkan penelitian dengan penuh pengertian dan bijaksana.
Ucapan terima kasih kepada Dr Desianto B. Utomo, PhD selaku anggota komisi pembimbing yang selalu mengingatkan penulis untuk menyelesaikan sekolah, dan membuka pintu untuk penulis saat membutuhkan bimbingan hingga memperjuangkan fasilitas pengujian.
Ucapan terima kasih kepada Dr Ir Sumiati, MSc sebagai penguji luar komisi dan Dr Ir Dwierra Evvyernie A, MS, MSc sebagai ketua program studi Ilmu Nutrisi dan Pakan, yang berkenan memberikan evaluasi dan masukkan untuk isi tesis ini pada ujian akhir.
Ucapan terima kasih disampaikan pula kepada Dr Vinai Rakphongpairoj yang memberikan kesempatan untuk menimba ilmu kembali, dan meluangkan waktu untuk membimbing dalam menyelesaikan tugas akhir.
Kata-kata terima kasih ini tidak akan pernah cukup untuk Dr Kenny R. Hazen yang membimbing penulis dari persiapan penelitian hingga akhirnya, dengan penuh ketulusan dan kesabaran, mengajar, mengarahkan, membimbing, memberikan inspirasi dan semangat baru.
Terima kasih kepada Dr Shaoyan Li yang memberikan kesempatan kepada penulis untuk bersekolah meskipun menyita waktu kerja, memberikan bimbingan dengan detail pada penulisan tugas akhir ini.
Penulis juga menyampaikan ucapan terima kasih kepada team Feed Technology CPI yakni Ibu Sri, Bapak Bagus, Mbak Ina, Mas Andi, Bapak Maryanto, Maureen, Wenny, Akbari, Bapak Ratiman, Mbak Noni, Mbak Elis dan Rizqi. Team dari Laboratorium yakni Ibu Binarti, Ibu Nursiah, Bapak Irwan dan Ibu Lela. Team dari RnD yakni Bapak Johnny Wang dan Bapak Faisal. Teman-teman seangkatan Sahera, Eva, Franky dan staff Pascasarjana Bapak Supriyadi dan Ibu Ade, serta teman-teman yang tidak dapat disebutkan satu persatu, yang berkenan membantu dengan tulus hati. memberikan kekuatan baru saat penulis jatuh. Untuk pengorbanan yang tidak pernah dapat terbalas.
Ucapan terima kasih untuk Ie Giem dan sahabat penulis Ingrid - Tay yang selalu berkenan untuk tunduk berdoa untuk penulis. Senantiasa ikut “ujian” saat penulis ujian.
DAFTAR PUSTAKA 60
1 Anti nutrisi pada bungkil kacang kedelai 3
2 Karbohidrat pada bungkil kacang kedelai 6
3 Komposisi dan kandungan nutrien ransum 22
4 Analisa bungkil kacang kedelai India dan Argentina 28
5 Berat badan pada periode starter 0-21 hari 30
6 FCR pada periode starter 0-21 hari 31
7 Konsumsi pakan pada periode starter 0-21 hari 31
8 Berat badan pada periode finisher 22-32 hari 32
9 FCR pada periode finisher 22-32 hari 32
10 Konsumsi pakan pada periode finisher 22-32 hari 33
11 Berat badan pada saat panen umur 32 hari 33
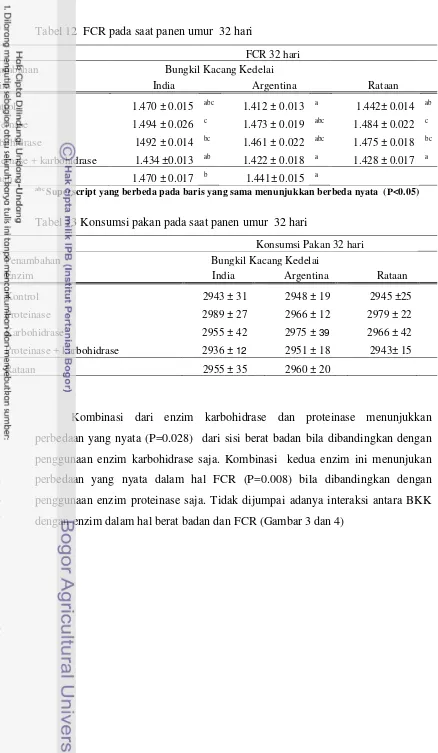
12 FCR pada saat panen umur 32 hari 34
13 Konsumsi pakan saat panen umur 32 hari 34
14 Kandungan air ekskreta 37
15 Konsumsi air minum 38
16 R2 dari korelasi kecernaan asam amino ileum dengan berat
badan dan FCR 39
17 Kecernaan asam amino esensial ileum (1) 40
18 Kecernaan asam amino esensial ileum (2) 41
19 Kecernaan asam amino buksn esensial ileum 42
20 R2 dari korelasi kecernaan serat kasar dengan berat badan dan
FCR 43
21 Kecernaan serat kasar 44
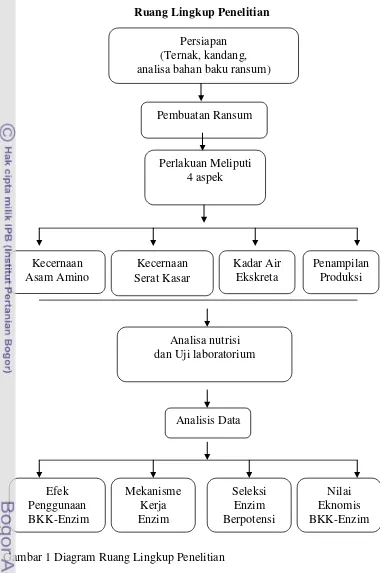
1 Ruang lingkup penelitian 19
2 Bungkil kacang kedelai India dan Argentina 29
3 Berat badan pada umur 32 hari 35
4 FCR pada umur 32 hari 35
5 Perubahan bedah bangkai pada pankreas ayam yang menggunakan
bungkil kacang kedelai India 46
6 Perubahan bedah bangkai pada pankreas ayam yang menggunakan
bungkil kacang kedelai Argentina 46
7 Perubahan bedah bangkai pada usus halus ayam yang menggunakan
bungkil kacang kedelai India 47
8 Perubahan bedah bangkai pada usus halus ayam yang menggunakan
bungkil kacang kedelai Argentina 47
9 Protein yang terperangkap dalam matriks serat kasar dinding sel 53
10 Proses pencernaan serat kasar 54
11 Strukur polisakarida pada bungkil kacang kedelai 56
12 Mekanisme kerja sinergis 58
1 Penampilan produksi pada umur 7 hari hingga 32 hari 64
2. Kurva penampilan produksi 1-32 hari 69
1
Latar Belakang
Kualitas Bungkil Kacang Kedelai dari Berbagai Negara
Bungkil kacang kedelai adalah salah satu bahan baku sumber protein
berkualitas yang dipergunakan pada pakan unggas. Bungkil kacang kedelai
umumnya berasal dari 5 negara pengekspor yakni Argentina yang merupakan
pengekspor terbesar sebanyak 48 % diikuti oleh Brazil 24 %, Amerika Serikat
15 %, Paraguay 2% dan India 7% (Swick 2012).
Tiap negara mempunyai perioda panen yang berbeda beda. Musim panen
kedelai di India umumnya terjadi pada bulan November hingga April. Sepanjang
perioda ini, bungkil kacang kedelai India mempunyai harga yang lebih rendah dari
bungkil kedelai yang lainnya. Selisih harga bungkil kedelai India dengan
Argentina berkisar 10 hingga 45 U$/MT. Kondisi ini menyebabkan ahli nutrisi
mempuyai peluang untuk menggunakan bungkil kacang kedelai India untuk
membuat pakan unggas dengan harga yang kompetitip di lapangan.
Akan tetapi, menggantikan bungkil kacang kedelai asal Argentina, Amerika
atau Brazil dengan bungkil kacang kedelai India dalam ransum unggas
membutuhkan perhitungan yang masak. Hal ini disebabkan oleh kualitas nutrisi
dari bungkil kacang kedelai tidak selalu seragam. Variasi ini dapat disebabkan
oleh varietas, genotip, anti nutrisi dan metoda pembuatan (Douglas & Parson
2000).
Menurut Lee (2001), bungkil kacang kedelai India mempunyai tingkat serat
kasar dan urease yang lebih tinggi dari bungkil kacang kedelai Brazil tetapi
sebaliknya mempunyai tingkat asam amino dan lemak yang lebih rendah.
Pendapat ini kemudian diperkuat oleh penelitian Thakur dan Hurburgh (2005)
yang melakukan pengamatan di Universitas IOWA State. Keduanya menyatakan
bahwa meskipun kandungan protein kasar dari bungkil kacang kedelai Argentina
lebih rendah dari bungkil kacang kedelai India, akan tetapi 5 asam amino utama
yakni lisina, treonina, metionina, sisteina dan triptofan lebih tinggi dari pada
Liu dan Zhu (2011) mengumpulkan bungkil kacang kedelai dari Argentina,
Amerika Serikat dan India dan menentukan kandungan nutrisi ketiganya. Bungkil
kacang kedelai India menunjukan kualitas yang paling rendah. Kandungan serat
kasar, abu dan urease dari bungkil kacang kedelai India lebih tinggi, sedangkan tingkat asam aminonya lebih rendah dari bungkil kacang kedelai Amerika Serikat
dan Argentina.
Ketersediaan dan pemanfaatan nutrisi dari bungkil kacang kedelai dapat
dipengaruhi oleh beberapa faktor anti nutrisi. Anti nutrisi ini dapat mempengaruhi
tingkat asupan pakan dan metabolisme nutrisi (Sinova et al. 2008). Kandungan anti nutrisi seperti proteinase inhibitor, khususnya tripsin inhibitor akan menekan tingkat kecernaan dari protein dan ketersediaan asam amino. Di sisi lain, anti
nutrisi karbohidrat seperti oligosakarida dan polisakarida bukan pati juga tidak
dapat dicerna oleh unggas. Kehadiran dari fraksi serat kasar ini dapat memberikan
dampak negatip pada tingkat ketersediaan asam amino dan energi (Swick 2007).
Di dunia saat ini, terdapat tiga metoda yang dipergunakan untuk
meningkatkan ketersediaan nutrisi dari bungkil kacang kedelai. Ketiga metoda
tersebut adalah manipulasi genetik, modifikasi proses pembuatan dan modifikasi
komposisi ransum dengan menambahkan enzim (Graham dan Partridge 2003).
Penggunaan enzim pada ransum yang menggunakan bungkil kacang kedelai pada
unggas memberikan perbaikan ketersediaan nutrisi dan penampilan produksi
unggas dengan tingkatan yang berbeda-beda.
Komposisi Kimia dari Bungkil Kacang Kedelai
Kedelai terdiri dari 8% kulit ari atau hull, 90% kotiledon dan 2% bahan
kecambah atau germ. Kedelai bukan hanya kaya akan minyak tetapi kaya akan protein dan karbohidrat. Dehulled fullfat soybean meal mengandung 20% minyak, 40% protein, 35 % karbohidrat dan 5% mineral dalam bahan keringnya (Choct
2010). Pada saat minyak diekstrak, sisanya disebut bungkil dan mengandung 48%
protein, 35-40% karbohidrat , 7-10% air, 5-6% mineral dan kurang dari 1%
Faktor Anti Nutrisi
Sebagaimana telah disebutkan diatas, faktor anti nutrisi diduga sebagai
salah satu pemicu perbedaan kualitas dari bungkil kacang kedelai karena
mempengaruhi ketersediaan nutrisi dan proses metabolisme (Swick 2007; Choct
et al 2010). Faktor-faktor anti nutrisi yang terdapat pada bungkil kacang kedelai adalah proteinase inhibitor, allergins, lektin, phytoesterogens, lipoxygenase,
saponin, phytin, oligosakarida dan polisakarida bukan pati (Tabel 1). Tabel 1 Anti nutrisi pada bungkil kacang kedelai
Faktor Anti Nutrisi Efek fisiologis Catatan
Proteinase Inhibitors
Lektin Aglutinasi sel darah merah Labil panas
Phytoesterogens Efek esterogenik (jumlah kecil) Tahan panas
Lipoxygenase Catalyzes oxidation asam lemak Labil panas Saponin Pahit, iritasi mukosa (jumlah kecil) Tahan panas
Phytin Interferensi dengan kecernaan mineral Tahan panas Oligosakarida dan
polisakarida bukan pati
Penampilan produksi, ekskreta lengket
dan diare Tahan panas
Sumber: Swick (2007)
Proteinase inhibitor yang terdapat dalam bungkil kacang kedelai berupa tripsin inhibitor. Anti nutrisi ini akan menyebabkan pembengkakan kelenjar pembentuk enzim pada pankreas, mengikat dan menginaktifkan enzim pankreas
yakni tripsin dan kimotripsin. Selanjutnya kerusakan ini akan menurunkan tingkat kecernaan protein. Proteinase inhibitor ini dapat diinaktifkan dengan
proses pemanasan dalam pembuatan bungkil kacang kedelai.
Allergins dalam bungkil kacang kedelai berupa protein glisinina dan konglisinina yang bersifat immunoreactive dan menyebabkan reaksi hipersensitip pada anak babi. Anak babi akan mempunyai titer antibodi yang tinggi terhadap
kedelai dan vili ususnya akan terkikis. Protein ini lebih stabil pada pemanasan bila
Lektin adalah glikoprotein yang dapat mengikat permukaan sel melalui
oligosakarida spesifik atau glycipeptida dan mempunyai daya ikat yang tinggi
pada epitel usus halus. Lektin dapat merubah struktur dari epitel usus dan tahan
pada proteolisis usus. Perubahan ini dapat mempengaruhi aktifitas vili usus dan
bahkan menyebabkan luka tukak pada usus sehingga meningkatkan kehilangan
nitrogen endogenus akibat dari kerusakan enterosit usus. Lektin dapat
dihancurkan oleh proses pemanasan pada pembuatan bungkil kacang kedelai
(Douglas et al. 1999). Lektin atau hemagglutinins adalah protein yang dapat menggumpalkan sel darah merah bila disuntikan dalam dosis tinggi (Swick 2007).
Phytoesterogens adalah senyawa isoflavone yang mempunyai efek esterogen. Senyawa ini tidak rusak dalam pemanasan tetapi terdapat dalam
konsentrasi yang jauh dibawah ambang untuk dapat memberikan efek negatif
pada unggas.
Lypoxygenase adalah enzim yang dapat melakukan proses oksidasi katalisa pada asam lemak. Enzim ini terpisah dari lubang minyak pada kedelai tetapi
dilepaskan dengan penggerusan kedelai. Lypoxygenase dapat dirusak oleh pemanasan pada saat pembuatan bungkil kacang kedelai.
Saponins adalah glycocides dengan deterjen kuat yang dapat memberikan rasa pahit dan mengiritasi membran mukosa. Saponins dapat menyebabkan ekskresi kolesterol dan menekan pertumbuhan. Senyawa ini tidak dirusak oleh
panas tetapi konsentrasinya dalam kacang kedelai jauh lebih rendah dari tingkat
yang dapat memberikan dampak negatip pada unggas.
Phytin adalah kompleks mineral dari ester hexaphosphate inositol. Senyawa ini dapat menggangu kecernaan mineral dan selanjutnya mengganggu proses
metabolisme pada unggas. Ikatan fosfor dengan phytin tidak banyak ditemukan pada kedelai.
Oligosakarida termasuk dalam kelompok gula-gula pada karbohidrat yang
mempunyai tingkat kecernaan terbatas. Berdasarkan bahan kering, bungkil kacang
kedelai dapat mengandung 6,2% sukrosa, 1,4 % raffinose dan 5,2% stachyose. Sukrosa dapat dicerna menjadi fruktosa dan glukosa, akan tetapi raffinose dan
stachyose sulit dicerna karena mempunyai ikatan -galaktosidase. Bila raffinose
halnya dengan nilai metabolisme energi bungkil kacang kedelai. Gula-gula
raffinose dan stachyose ini tidak dirusak oleh pemanasan dan dapat dikurangi dengan proses ekstraksi (Swick 2007). Anti nutrisi tersebut diatas dapat dilihat
pada Tabel 1.
Polisakarida bukan pati merupakan bagian dari karbohidrat pada kacang
kedelai yang bersifat anti nutrisi pada unggas. Dari 35-45 % karbohidrat yang
dapat dijumpai pada bungkil kacang kedelai 1% diantaranya adalah pati, 10%
berikutnya adalah gula bebas dan 20-30% sisanya adalah polisakarida bukan pati.
Komposisi dari karbohidrat bungkil kacang kedelai ini bervariasi, dipengaruhi
oleh lokasi penanaman, genotip dan proses pembuatan bungkil kacang kedelai
(Chot et al. 2010). Polisakarida bukan pati ini terutama terdiri dari kelompok Selulosa, hemiselulosa danpektin(Tabel 2).
Empat belas persen dari polisakarida bukan pati merupakan polisakarida
bukan pati yang dapat larut dan 16% sisanya yang tidak dapat larut. Polisakarida
bukan pati yang tidak dapat larut, tidak dapat dicerna sama sekali, sedangkan
yang larut masih dapat dicerna sebagian. Hal ini disebabkan karena unggas tidak
mempunyai endogenus enzim yang mampu mencerna polisakarida bukan pati dan
beberapa oligosakarida. Kecernaan dari senyawa-senyawa ini juga dipengaruhi
oleh species hewan, umur, struktur kimia, jumlahnya dalam ransum, bahan kimia
seperti asam yang terdapat pada tembolok unggas atau degradasi mikroba di usus
buntu (Chot et al. 2010).
Polisakarida bukan pati terutama terdapat di dalam dinding sel dari kacang
kedelai. Dinding sel ini dibentuk dari struktur selulosa-xyloglucan, structur pektin dan struktur protein. Benang-benang selulosa berikatan dengan polimer dari
xyloglycan. Struktur selulosa–xyloglucan lalu berikatan dengan sturtur pektin (Fischer 2006). Substansi pektin bersifat mengikat polisakarida lain pada matriks
dinding sel, bahkan mengikat 10 % dari protein bahan kering yang berada
diantara matriks dinding sel (Tahir et al. 2008).
Struktur selulosa-xyloglucan mempunyai selulosa (1,4)--D-glucan)
sebagai komponen utama. Rantai selulosa berikatan dengan hemiselulosa yang
Tabel 2 Karbohidrat pada bungkil kacang kedelai dalam g/kg bahan kering.
Polisakarida bukan pati 2.0-3.0 2.75 2.27
Selulosa 0.8 0.62 1.68 1.03 2.00
Sumber : *Choct 2011; **Graham dan Bedford 2007; ***Swick 2007
Struktur berikutnya adalah struktur pektin yang pada bungkil kacang
kedelai terdiri dari polisakarida pektik. Struktur pektin ini tersusun oleh
xylogalacturonan dengan rangka dasar yang mengandung (1,4)-a-D-galakturonik.
Xylose berikatan dengan asam galakturonat. Xylogalacturonan diduga berikatan dengan rhamnogalakturonan.
Rhamnogalakturonan (RG) adalah struktur lain yang terdapat pada pektik
I dan II. RG I mengandung rantai panjang dari (1,4)- galakturonosil dan
(1,2)- rhamnose. Disamping itu RG I juga mengandung arabinofuranosil-,
galactopyranosil-, dan residu fucopyranosil. RGII hanya terdiri dari rantai
galakturonan.
Pada bungkil kacang kedelai juga dijumpai rantai arabinan, galaktan dan
arabinogalaktan yang berikatan pada rantai dasar rhamnose. Struktur-struktur ini
nampak seperti rambut-rambut. Arabinogalaktan dibagi kedalam 2 type yaitu type
I dan type II. Pektin umumnya mengandung type I. Type I mempunyai rantai
dasar (1,4)- galactans dengan percabangan (1,5)- arabinose atau rantai
galaktosa yang berikatan dengan (1,6) pada struktur galaktan. Perbandingan
arabinose dan galaktosa dalam arabinogalaktan dan jumlah rantai percabangan
dalam arabinose bervariasi tergantung pada species dari kedelai (Fischer 2006)
Tahir et al. (2008) menyatakan bahwa asam galakturonat adalah komponen utama dari pektin dan muncul sebagai polimer asam rhamno galakturonat atau
poli- asam galakturonat. Bungkil kacang kedelai mengandung
rhamnogalakturonan dengan struktur dasar 1,4- asam galakturonat dan
1,2- rhamnose, dengan sejumlah rantai samping yang terutama terdiri arabinose
dan galaktosa.
Dinding sel kacang kedelai juga mengandung Beta-mannan yang disebut
sebagai β-galactomannan yang merupakan polisakarida dengan pengulangan unit
mannose dengan galaktosa atau glukosa atau keduanya, yang ditemukan
berlekatan dengan rantai dasar β-mannan. Kelarutan dari β-mannan meningkat dengan bertambahnya molekul galaktosa pada rantai dasar mannan. β-mannan ini akan mempengaruhi penampilan dari unggas, karena mempengaruhi penyerapan
glukosa dan air. β-mannan juga mempengaruhi produksi insulin, glukagon dan
insulin-like growth factor (Hsiao et al. 2006).
Metoda untuk Mengurangi dan Mengeliminasi Faktor Anti Nutrisi
Metoda untuk mengurangi dan mengeliminasi faktor-faktor anti nutrisi saat
ini terdiri dari proses bioteknologi, modifikasi proses produksi dan modifikasi
komposisi ransum (Graham & Partridge 2003). Proses bioteknologi yang
sehingga bungkil kacang kedelai yang baru mempunyai tingkat stachyose yang rendah, kandungan lemak dan methionin yang lebih tinggi dari sebelumnya (Nab
& Boorman 2002).
Metoda berikutnya adalah modifikasi proses produksi. Dalam proses
pembuatan bungkil kacang kedelai, kacang kedelai mentah akan melalui proses
ekstraksi minyak dengan metoda steam conditioned, flaked dan pencucian dengan menggunakan hexane untuk melepaskan minyak. Hasil dari proses ini berupa
deffated flakes yang kemudian dimasukkan ke dalam desolventizer-toaster untuk melalui proses pemanasan agar dapat melepaskan residu hexane dan merusak anti
nutrisi peka terhadap panas seperti tripsin inhibitor. Proses eliminasi senyawa anti nutrisi dapat dilanjutkan dengan menggunakan vacuum loop, low heat flash desolventizer. Pada alat ini, bungkil kacang kedelai akan melewati cairan acid aqueous ethanol extractor untuk kemudian mengeluarkan sukrosa, raffinose, stachyose, phytoesterogens, mendenaturasi antigenik protein, tripsin inhibitor dan
conglisinina. Produk yang dihasilkan diberi nama konsentrat protein kedelai atau
soy protein concentrate (Swick 2007).
Metoda terakhir adalah dengan menambahkan enzim pada formula pakan.
Metoda ini akan dibahas lebih lengkap pada sub bab berikut.
Enzim
Enzim adalah katalis organik. Dalam kimia klasik, katalis adalah substansi
yang mempengaruhi kecepatan reaksi kimia yang karakternya tidak berubah pada
akhir reaksi. Mayoritas enzim adalah kompleks dari protein dengan berat molekul
yang besar. Sebagian dari enzim membutuhkan kofaktor organik (koenzim) untuk
bekerja efisien dan kofaktor metalik untuk berikatan dengan enzim dengan ikatan
kovalen atau berikatan dengan substrat utama (Mc Donald et al. 2002).
Di dalam aktifitasnya, enzim akan membentuk kompleks enzim-substrate.
Kompleks ini kemudian akan dipecah dan menghasilkan produk baru beserta
enzimnya, sebagaimana rumus berikut:
E + S ES E + P
Kompleks yang dibentuk oleh substrate dan sisi aktif pada permukaan enzim terjadi dengan ikatan ionik, ikatan hidrogen, kovalen kelompok sulphydryl atau
antara sisi aktif dari kelompok substrat dengan sisi aktif enzim. Model sepesifisitas ini dikenal dengan nama model lock and key. Spesifisitas enzim dikatakan absolut bila aktifitasnya hanya terbatas pada satu substrat saja. Akan
tetapi enzim yang dapat mengkatalisa lebih dari satu kelompok substrat sehingga
dapat disebut spesifisitas relatif. Tripsin dan pepsin mengkatalisa ikatan peptida
yang hancur. Sedangkan kimotripsin mengkatalisa hydrolytic cleavage pada ikatan peptida di bagian residu karboxyl yang dibentuk dari asam amino aromatic
(Mc Donald et al. 2002).
Efektifitas dari kerja enzim dapat dipengaruhi oleh beberapa faktor seperti
misalnya konsentrasi substrat, konsentrasi enzim, inhibitors dan faktor lingkungan
seperti suhu dan pH. Dalam suatu reaksi apa bila enzim berlebih dan
konsentrasinya tetap, maka peningkatan konsentrasi substrat akan meningkatkan
kecepatan reaksi. Hal ini disebabkan oleh peningkatan penggunaan pusat
keaktifan (active centres) oleh enzim. Jika konsentrasi substrat terus ditingkatkan, penggunaan pusat keaktifan menjadi maksimal dan kecepatan reaksi akan
menurun. Dampak negatif yang mungkin terjadi adalah ikatan yang tidak
sempurna antara enzim dan substrat akibat dari kompetisi pusat keaktifan. Efek dari konsentrasi substrat pada kecepatan reaksi katalisa enzim dapat dianalisa
dengan Michaelis-Menten constant, Km (Mc Donald et al. 2002).
Di sisi lain, apa bila dalam suatu reaksi, substrat ada dalam jumlah yang
berlebih, peningkatan konsentrasi enzim akan memberikan respon linier dengan
kecepatan reaksi yang meningkatkan pusat keaktifan untuk pembentukan
kompleks enzim-substrate. Bila konsentrasi enzim terus ditingkatkan akan
menyebabkan faktor pembatas seperti ketersediaan koenzim menurun.
Substansi inhibitors pada aktifitas enzym dapat digolongkan pada 2 kelompok yakni reversible inhibition dan non-reversible inhibition. Reversible inhibition
meliputi non-covalent bonding pada inhibitor enzim seperti misalnya pada sintesa pembentukan asam folat dari PABA dengan sulfanilamid.. Irreversible inhibition
meliputi covalent bonding pada sisi aktif dan tidak dapat dikembalikan dengan kelebihan substrate atau dengan dialisis. Sisi aktif akan ditutup dan proses katalisa
diinaktifkan. Contoh yang paling populer adalah racun organofosfor pada
Faktor lingkungan pertama yang mempengaruhi aktifitas enzim adalah pH.
Konsentrasi ion hidrogen mempunyai pengaruh yang penting pada efisiensi kerja
enzim. Banyak enzim yang aktif pada pH 6-7. Enzim ekstraselular dapat
menunjukkan aktifitas maksimum di dalam asam atau basa, tetapi selang aktual
dari setiap individual enzim hanya bekerja pada 2.5-3.0 unit. Diluar dari kisaran
ini aktifitas akan menurun dengan cepat. Penurunan efisiensi ini diakibatkan
karena derajat ionisasi dari substrat dan enzim, dimana ikatan diantara pusat
keaktidan bersifat elektrostatik. Selanjutnya pada kondisi yang terlalu asam atau
terlalu basa, akan terjadi proses denaturasi dan potensi enzim akan hilang (Mc
Donald et al. 2002).
Reaksi enzim sangat sensitif terhadap perubahan temperatur. Efisiensi
dari reaksi katalisa enzim meningkat dengan peningkatan suhu. Secara garis besar,
kecepatan reaksi akan meningat mengikuti van t’Hoff rule dengan faktor 2-3 untuk setiap peningkatan 10 derajat Celcius (Bisswanger 2008). Bila
temperatur meningkat terus, proses denaturasi protein akan terjadi. Hal ini
disebabkan karena pada saat panas meningkat, terjadi pergerakan molekul yang
akan menyebabkan hilangnya pusat keaktifan pada sisi aktif permukaan enzim.
Diatas 50 derajat Celcius, proses kerusakan ini akan meningkat dengan cepat.
Banyak enzim yang akan rusak pada pemanasan 100 derajat Celcius. Aktifitas
enzim terhadap perubahan temperatur dapat digambarkan dengan kurva lonceng.
Akan tetapi peningkatan dan penurunan aktifitas berbeda dengan pH, hal ini
disebabkan karena pada titik tertinggi bukanlah merupakan temperatur optimum,
akan tetapi merupakan temperatur maksimum, titik dimana mulai terjadi
denaturasi protein. Di lain pihak, denaturasi protein tidak hanya dipengaruhi oleh
temperatur pemanasan, akan tetapi oleh jangka waktu pemanasan. Enzim dari
mikroorganisma yang dapat beradaptasi pada kondisi dingin akan dapat tetap
efektif pada suhu dingin mendekati nol, dan sebaliknya, enzim dari
mikroorganisma yang hidup pada suhu tinggi dapat tetap optimal pada suhu 100
derajat Celcius (Mc Donald et al. 2002).
Di lapangan, enzim banyak dipergunakan untuk membuat least cost feed
atau formula pakan yang ekonomis. Pendekatan formula ini dapat dilakukan
meningkatkan nilai nutrisi terhadap enzim berdasarkan pada nilai matriks bahan
baku atau meningkatkan nilai metabolisme energi dari bahan baku tertentu
(Francesh M, Geraert PA. 2009). Pendekatan ini memerlukan akurasi pengetahuan
efektifitas enzim terhadap bahan baku, karena masing masing enzim mempunyai
target substrat, mekanis kerja dan interaksi yang berbeda-beda. Faktor-faktor
yang dapat mempengaruhi efektifitas enzim termasuk spesifisitas enzim,
konsentrasi substrat, dosis enzim, interaksi enzim-enzim, kualitas dan type bahan
baku, level nutrisi dalam pakan, dan umur dari unggas (Francesh & Geraert 2009)
.
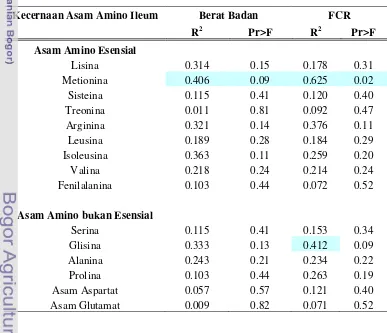
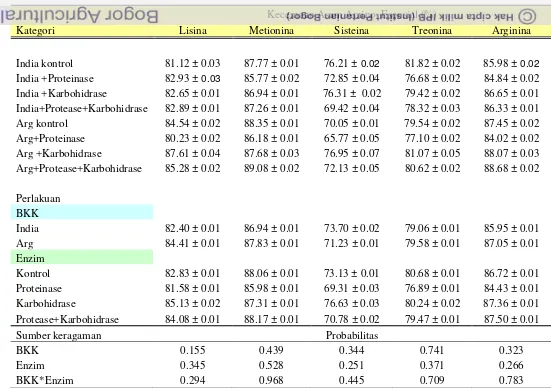
Kecernaan Asam Amino
Selain dipengaruhi oleh negara asal bungkil kacang kedelai, kecernaan
asam amino dapat dipengaruhi oleh proses pembuatan, penyimpanan, penanganan
dan penambahan enzim (Swick 2012).
Proses pembuatan yang terlalu masak atau over toasting, akan menyebabkan nilai bioavailability lisina yang rendah akibat dari pembentukan
Amadori dan produk reaksi Maillard. Senyawa ini debentuk saat kelompok free epsilon amino dalam lisina bereaksi dengan reducing sugars selama proses pemanasan sehingga membentuk polymers yang tidak dapat dicerna. Pengujian
unbound lysine atau lisina yang tidak terikat menggunakan o-methul isourea dapat dipergunakan untuk menguji protein tidak tercerna ini (Swick 2012).
Penyimpanan pada suhu lebih dari 30 derajat Celcius dan kelembaban
84 %RH selama lebih dari 7 bulan akan merubah struktur dan ekstraksi glisinina ,
sebagai major globulin storage proteins pada kedelai. Perubahan karakter glisinina akan menurunkan kelompok free sulfhydryl dan meningkatkan kandungan glikoprotein, dan menurunkan daya cerna protein (Swick 2012).
Penyimpanan juga mempengaruhi kecernaan asam amino. Meskipun total
asam amino tidak berubah akan tetapi nilai kecernaan lisina yang diprediksi
dengan NIRS menurun pada rataaan 0.019% per bulan dan FCR dari ayam yang
mengkonsumsi bungkil kacang kedelai akan lebih besar. Peningkatan FCR
sebanyak 5 poin dari 1.47 menjadi 1.52 setelah penyimpanan pada kondisi hangat
Anti nutrisi pada bungkil kacang kedelai dapat mengganggu kecernaan
protein dan meningkatkan ekskresi dari endogenous protein (Feng et al. 2007). Tripsin inhibitor dan lektin adalah anti nutrisi yang dapat mengurangi kecernaan protein dan menyebabkan hypertrophy pankreas. Tripsin inhibitor yang dapat dijumpai pada kedelai mentah ini dapat berinterferensi dengan fungsi tripsin dan
kimotripsin sehingga mengurangi total aktifitas dari enzim proteolitik. Di sisi lain,
tripsin inhibitor dapat merubah morfologi dari usus yang nomal. Perubahan pada usus yang pernah terjadi adalah atropi villi usus, peningkatan mitosis sel kripta
usus, hiperplasia kripta bahkan mendukung terjadinya kasus kegagalan
penyerapan atau malabsorption syndrome.
Kecernaan asam amino dapat meningkat setelah penambahan enzim
karbohidrase dan proteinase, akibat dari protein yang berada diantara matriks
dinding sel terlepas. Substansi pektik bersifat mengikat polisakarida pada matriks
dinding sel, bahkan mengikat 10 % dari protein bahan kering yang berada
diantara matriks dinding sel (Tahir et al. 2008).
Kelompok enzim hemiselulasedanenzimpektinase (rhamnogalakturonase
dan poli-galakturonase) dapat membelah polisakarida pektik pada ikatan antara
asam galakturonat dan rhamnose sehingga membuka peluang untuk nutrisi yang
terperangkap seperti protein untuk mengalamai proses hidrolisa. Pemecahan
matriks kompleks diding sel oleh multi-karbohidrase juga akan meningkatan
peluang untuk kontak enzim proteinase pada protein yang terperangkap pada
matriks dinding sel (Tahir et al. 2008).
Kecernaan Oligosakarida dan Polisakarida bukan Pati
Kecernaan oligosakarida dalam bungkil kacang kedelai dipengaruhi oleh
variasi level raffinose dan stachyose. Level keduanya dipengaruhi oleh komposisi karbohidrat yang berbeda-beda menurut lokasi geografis, proses panen, dan
proses produksi bungkil. Oleh karena itu tidak heran jika sukrosa mempunyai
level 3-8%, raffinose 0.1-1.5% dan stachyose 1-6% (Choct 2010).
Kecernaan dari oligosakarida bungkil kacang kedelai juga sangat rendah.
Beberapa peneliti menyatakan prebiotik dari oligosakarida meningkatkan proses
fermentasi dari mikroflora usus sedangkan peneliti lainnya menyatakan, kehadiran
oligosakarida akan meningkatkan retensi cairan atau fluid retention, mengganggu tekanan osmotik, meningkatkan tegangan permukaan, produksi hidrogen dan
diare. Kecernaan oligosakarida sangat tergantung pada populasi mikroflora usus,
sumber, type dan konsentrasi (Choct 2010).
Kecernaan polisakarida bukan pati tergantung pada level polisakarida
bukan pati dapat larut dan tidak dapat larut serta daya cerna dari ayam itu sendiri.
Polisakarida bukan pati yang larut dapat meningkatkan tegangan permukaan isi
usus dan merubah fisiologi dan ekosistem usus. Hal ini mungkin berhubungan
dengan lambatnya gerakan pencernaan atau digesta passage rate. Digesta yang bergerak lambat dengan tekanan oksigen yang sangat rendah dapat menggangu
proses pencernaan dan menjadi lingkungan yang baik bagi kehidupan Clostridium
yang menyebabkan necrotic enterisis. Polisakarida bukan pati khususnya metil
selulosa dan pektin dapat membentuk larutan kental yang dapat menghambat
pencernaan protein pada terminal ileum disamping itu dapat meningkatkan kehilangan endogenus asam amino akibat sekresi usus yang lebih tinggi dalam
usaha untuk meningkatkan kecernaan (Choct 2010).
Kecernaan dari polisakarida bukan pati sangat tergantung pada spesies
hewan, umur, kelarutan, struktur kimia dan jumlahnya dalam pakan, kapasitas
mikroflora dalam usus dan jangka waktu (transit time) isi usus berada di dalam usus besar. Kecernaan polisakarida bukan pati pada unggas sangat kecil. Pada
ayam jantan nilai kecernaan polisakarida bukan pati dari pakan yang mengandung
6.9% polisakarida bukan pati hanyalah berkisar 13% (Choct 2010).
Proses pencernaan polisakarida bukan pati pada unggas diawali dengan
proses pencernaan asam pada tembolok, lalu diiikuti dengan fermentasi mikroba
pada bagian akhir usus halus dan seluruh bagian usus besar. Pencernaan fisik pada
unggas terjadi sangat sedikit, yakni pada saat material serat masuk ke dalam
ampela untuk dihaluskan menjadi partikel yang lebih halus. Proses ini kemudian
dilanjutkan dengan proses fermentasi oleh mikroflora usus pada usus buntu.
Produk dari pencernaan dan fermentasi ini adalah asam lemak rantai pendek yang
menyatakan bahwa total kontribusi asam lemak terbang dapat mencapai 42 J per
hari yang tidak dipengaruhi oleh sumber polisakarida bukan pati manapun dan
dapat dimanfaatkan untuk metabolisme energy, kira-kira sebesar 2-3% dari energi
metabolis yang dimakan. Bila polisakarida bukan pati dihidrolisa, maka anti
nutrisi dapat dicerna, difermentasi dan diserap sehingga mengurangi dampak
negatif.
Metoda Penambahan Enzim pada Bungkil Kacang Kedelai
Penelitian dengan menggunakan enzim untuk menekan anti nutrisi dan
meningkatkan nilai nutrisi bungkil kacang kedelai umumnya menggunakan
dua kelompok enzim, yakni enzim proteinase dan enzim karbohidrase.
Enzim proteinase adalah enzim yang mengkonduksi proses proteolisis
dengan menghidrolisa ikatan peptida pada rantai asam amino di dalam rangkaian
polipeptida. Enzim proteinase dapat diklasifikasikan berdasarkan keaktifannya
yakni pada kondisi asam, basa atau netral. Disamping itu dapat pula
dikelompokan berdasarkan karakter dari sisi aktif proses katalisa yakni
serina proteinase, sisteina (thiol) preoteinase, aspartikproteinase dan
metalloproteinases. Perlekatan dari proteinase pada substrat tergantung pada struktur dari sisi katalisa dan asam amino utama dari aktifitasnya. Serina
proteinase adalah kelompok enzim yang memotong ikatan peptida protein. Asam
amino pada sisi aktif dari enzim ini selalu serine. Ada 3 anggota dari serine
proteinase yang umum dikenal yakni kimotripsin, tripsin dan elastase. Ketiga enzim ini pada unggas disintesa oleh sel-sel acinar pankreas, disekresi ke dalam usus halus dan bertanggung jawab untuk mengkatalisa proses hidrolisa dari ikatan
peptida. Ketiga enzim ini mempunyai lokasi target rantai polipeptida yang
berbeda. Kimotripsin, memotong rantai peptida pada residu bulky hydrophobic amino acid, termasuk fenilalanina, triptofan dan tirosina. Tripsin memotong ikatan peptida pada residu positively-charged amino acid, yang muncul sebagai residu asam aspartat dan kemudian berinteraksi dengan arginina dan lisina pada
kacang kedelai bertujuan untuk meningkatkan hidrolisa protein dan mendegradasi
residu faktor anti nutrisi yakni tripsin inhibitor (Yu et al. 2006). Berdasarkan publikasi yang ada, pemberian enzim proteinase pada ransum unggas yang
menggunakan jagung dan bungkil kacang kedelai dapat memberikan peningkatan
berat badan sebesar 3.6 % lebih baik dari kontrol (Beal et al. 1998; Graham dan Partridge 2003). Sedangkan penggabungan proteinase dengan enzim karbohidrase
dapat meningkatkan berat badan sebesar 0.5 – 10.9 % lebih baik dari kontrol dan
perbaikan konversi pakan sebesar 0.78 – 8.70% dibandingkan dengan kontrol
(Cowieson dan Adeola 2005).
Enzim eksogenus proteinase dapat membantu melepaskan peptide dari
protein pakan dan bekerja sama dengan enzim endogenus pankreas yang terdapat
di dalam usus. Target utama dari eksogenus proteinase adalah faktor-faktor anti
nutrisi seperti residu tripsin inhibitor, lektin dan antigenik protein (Waldroup et al. 2006; Coon 2010).
Enzim karbohidrase adalah kelompok enzim yang umumnya dipergunakan
untuk ransum dengan bahan baku biji-bijian. Saat ini terdapat banyak jenis enzim
diantaranya: (1) enzym -glukanase yang menghidrolisa -glukans menjadi
oligosakarida dan glukosa, (2) amilase yang mendegradasi pati menjadi
maltoligosakarida dan dekstrin untuk kemudian menjadi D-glukosa, (3) selulase
yang menghidrolisa selulosa menjadi D-glukosa, (4) xylanase yang menghidrolisa
arabinoxylan menjadi D-xylose (5) -galaktosidase dan -mannanase yang menghidrolisa galaktomannan menjadi D-manosa dan D-galaktosa.
Berdasarkan penelitian pada struktur dinding sel, bungkil kacang kedelai
mempunyai struktur polisakarida yang sangat kompleks. Oleh karena itu,
penggunaan enzim karbohidrase pada bungkil kacang kedelai bertujuan untuk
menghidrolisa strukur polisakarida dan oligosakarida pada dinding sel untuk
melepaskan nutrien dan meningkatkan penyerapan (Yu et al. 2006). Tegangan isi usus akan menurun setelah struktur polisakarida bukan pati terhidrolisa sehingga
memungkinkan kontak antara isi usus dengan endogenus enzim, mengembalikan
Fischer (2006) menyatakan bahwa untuk kesempurnaan proses degradasi
dinding sel dari kedelai diperlukan gabungan aktifitas dari beberapa enzim. Pada
dasarnya enzim-enzim tersebut dapat digolongkan menjadi tiga kelompok enzim
yaitu pektinase, hemiselulase dan sellulase. Kelompok enzim cellulolytic yang diperlukan untuk mendegradasi selulosa adalah endo-(1,4)--D-glukanase
(selulase), exo-(1,4)--D-glukanase (cellobiohydro-lase), -D-glukosidase (sellobiase).
Fischer (2006) menyatakan beberapa beberapa kelompok enzim
hemiselulase diperlukan untuk mendegradasi stuktur hemiselulosa yang sesuai.
Kelompok hemiselulase yang utama adalah endo--(1,4)-D-xylanases,
endo-- (1,4)-D-mannanases, dan endo--(1,3)-(1,4)-glukanases. Kelompok hemiselulase yang berikutnya adalah enzim yang mendegradasi galaktan dan
arabinan yakni endo--(1,5)-Larabinanases,endo--(1,4)- dan -
-(1,3)/(1,6)-D-galaktanases, dimana keduanya disebut sebagai enzim pendegradasi hemiselulase dan pektin. Selanjutnya dinyatakan pula bahwa untuk menyempurnakan aktifitas
enzim ini diperlukan penggunaan enzim kelompok glikosidase yakni
-xylosidase, -galaktosidase, dan - arabinofuranosidases (Fischer, 2006).
Enzim -galaktosidase akan menghidrolisa oligosakarida raffinose dan
stachyose menjadi asam lemak rantai pendek seperti asetat, propionat dan butirat yang selanjutnya dapat dipergunakan sebagai sumber energi (Choct 2010).
Berdasarkan publikasi yang ada, pemberian enzim karbohidrase pada ransum
yang menggunakan jagung dan bungkil kacang kedelai dapat memberikan
peningkatan berat badan sebesar 2.22 - 9.30 % lebih baik dari kontrol.
Sedangkan dari sisi konversi pakan, akan dijumpai peningkatan sebesar
0.88 – 11.04% lebih baik dari kontrol (West et al. 2007; Tahir et al. 2008)
Enzyme xylanase dan β-glukanase yang diberikan pada ransum ayam pedaging akan meningkatkan penampilan, kecernaan, merubah morfologi usus,
ukuran organ cerna dan level VFA (Wang et al. 2005).Level polisakarida bukan pati yang tinggi dapat meningkatkan tegangan permukaan dan menghambat
kontak antara isi usus dengan enzim. Saluran pencernaan beradaptasi dengan
kondisi ini dengan melakukan kompensasi seperti merubah morfologi saluran
Polisakarida bukan pati dengan ukuran besar juga sangat sulit masuk ke dalam
usus belakang karena tegangan permukanan yang tinggi memperlambat aliran isi
usus. Di lain pihak, mikroba pada usus buntu tidak dapat memfermentasi molekul
polisakarida bukan pati dengan ukuran yang besar. Penambahan enzim akan
mendegradasi molekul polisakarida dan oligosakarida menjadi lebih kecil
sehingga tegangan permukaan isi usus menurun, kecepatan aliran usus normal.
Karbohidrat dengan berat molekul yang lebih rendah dapat difermentasi oleh
mikroba di dalam usus buntu sehingga lebih banyak VFA yang dapat dihasilkan.
Kelompok enzim hemiselulase dan pektinase (rhamnogalakturonase dan
poli-galakturonase) akan membelah polisakarida pektik, ikatan antara asam galakturonat dan rhamnosa sehingga membuka peluang untuk nutrisi yang
terperangkap agar dapat dihidrolisa (Tahir et al. 2008). Xylanase dan mannanase dari hemiselulase diharapkan memecah xylan dan mannan menjadi xylose dan mannosa. Pemecahan matriks kompleks diding sel oleh multi-karbohidrase juga akan meningkatan peluang untuk kontak enzim proteinase pada protein yang
terperangkap pada matriks dinding sel.
Penambahan enzim proteinase dan karbohidrase secara individual dan
kombinasi pada ransum yang menggunakan bungkil kacang kedelai akan
meningkatkan kecernaan dari protein kasar dan meningkatkan kecernaan dari
polisakarida bukan pati. Enzim karbohidrase yang menunjukan efek sinergisme
dengan enzim proteinase. Pada penambahan enzim proteinase dan karbohidrase,
total protein tercerna meningkat 2.9%, meskipun efek ini dinyatakan tidak sama
untuk setiap individu asam amino. Perbedaan ini tergantung dari sumber asam
amino dan asam amino yang merupakan substrat dari endogenus enzim.
Penambahan enzim dalam pakan selanjutnya akan meningkatkan keseluruhan
kecernaan dan mengurangi kehilangan asam amino endogenus. Di lain pihak
peningkatan kecernaan ini akan meningkatkan efisiensi energi dari pencernaan
dan menyediakan lebih banyak energi untuk pertumbuhan (Zanella et al. 1999) . Menurut Baker et al. (1996) dalam Zanella et al. (1999), ransum yang menggunakan jagung dan bungkil kacang kedelai mempunyai keterbatasan asam
Penambahan enzim akan meningkatkan treonina sebesar 3% dan valinasebanyak
2.3 % dan dapat meningkatkan keseimbangan asam amino dalam ransum.
Tujuan
Kajian mengenai bungkil kacang kedelai India masih sangat terbatas dan
kajian mengenai penambahan enzim dan dampaknya terhadap tingkat kecernaan
nutrisi bungkil kacang kedelai India belum pernah dipublikasikan. Di lain pihak
penggunaan ransum yang mengandung bungkil kacang kedelai pada ayam
pedaging dilapangan menunjukkan gangguan penampilan produksi. Oleh sebab
itu penelitian ini difokuskan pada BKK India.
Penelitian ini mempunyai 5 tujuan sebagaimana berikut :
1. Mengetahui efek dari penggunaan BKK India dan Argentina pada
penampilan produksi ayam pedaging.
2. Mengetahui efek dari penambahan enzim pada tingkat kecernaan asam
amino dan serat kasar.
3. Mekanisme kerja dari enzim di dalam fisiologi usus dan interaksi diantara
enzim.
4. Menyeleksi enzim yang berpotensi untuk meningkatkan kecernaan asam
amino dan serat kasar bungkil kacang kedelai.
5. Menentukan nilai ekonomis dari penggunaan bungkil kacang kedelai
India dan Argentina dengan dan tanpa enzim
Manfaat yang didapat dari hasil penelitian ini adalah diperoleh enzim yang
dapat meningkatkan kecernaan dan penampilan produksi dari ransum yang
menggunakan bungkil kadang kedelai. Enzim ini dapat membantu praktisi
peternakan menyusun ransum unggas yang ekonomis dengan menggunakan
bungkil kacang kedelai India di bulan November hingga April saat harga bungkil
METODE PENELITIAN
Waktu dan Tempat
Penelitian ini berlangsung selama bulan September hingga bulan
Nopember 2011. Penelitian dilaksanakan di kandang penelitian Cikupa dan
Laboratorium pabrik pakan Balaraja PT Charoen Pokphand Indonesia.
Bahan dan Alat Ternak
Penelitian ini menggunakan anak ayam broiler strain Ross berumur satu
hari dan berjenis kelamin jantan sebanyak 400 ekor. Anak ayam ini berasal dari
penetasan komersial yang telah divaksinasi dengan vaksin NDIBD killed dan
NDIB live di penetasan pada umur satu hari. Anak ayam kemudian dibagi ke
dalam 8 perlakuan secara acak. Masing masing perlakuan menggunakan 10
ulangan dan masing masing terdiri dari 5 ekor ayam.
Kandang
Kandang yang dipergunakan adalah kandang batere sebanyak 80 buah
dengan kapasitas 5 ekor ayam. Masing-masing batere dilengkapi dengan lampu
pemanas 60 watt, 1 tempat pakan dan 1 tempat air minum. Kandang batere juga
dilengkapi dengan tempat penampungan ekskreta.
Ransum
Jenis ransum yang dipergunakan adalah ransum ayam pedaging, yang
dipergunakan dari umur 1 hari hingga panen di umur 32 hari. Pemberian pakan
dan air minum dilakukan dengan terus menerus dan tidak dibatasi.
Bahan baku yang dipilih hanya 4 macam, yakni jagung, bungkil kacang
kedelai India atau Argentina, minyak kelapa sawit dan premiks. Komposisi
ransum dibuat dengan tingkat metabolisme energi yang sama yaitu 2900 kcal dan
tingkat kecernaan asam amino yang sama 1% lisina tercerna, 0.38% metionina
tercerna dan 0.75% metionina dan sisteina tercerna yang dapat dilihat pada
Tabel 3.
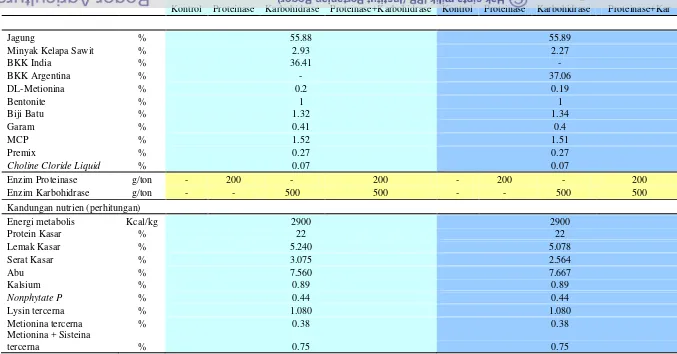
dipakai diproduksi dari Aspergillus niger dan mengandung 2500 - galaktosidase unit dan 5000 - mannanase 5000 unit. Dosis
penggunaannya adalah 500 g/ton pakan. Enzim proteinase yang dipergunakan
diproduksi dari Bacillus licheniformi danmengandung min 75.000 PROT. Dosis penggunaannya adalah 200 g/ton pakan. Aktifitas enzim untuk protease diukur
dengan PROT unit, dimana 1 unit didefeinisikan sebagai jumlah enzim yang dapat
melepaskan 1 µmol p-nitroaniline dari 1 µM substrat Suc-Ala-Ala-Pro-Phe-p -nitroaniline setiap 1 menit pada pH 9.0 dan suhu 37°C.
Perlakuan
Dua jenis kacang kedelai India dan Argentina dipergunakan dalam
penelitian ini dengan ditambahkan enzim-enzim berupa enzim proteinase dan
karbohidrase.
Perlakuan jenis kacang kedelai, yaitu :
A1 : Kacang kedelai India
A2 : Kacang kedelai Argentina
Perlakuan enzim, yaitu :
B1 : tanpa enzim
B2 : dengan enzim proteinase
B3: dengan enzim karbohidrase
Tabel 3 Komposisi dan kandungan nutrien ransum
Komponen Satuan Ransum BKK India Ransum BKK Argentina
Kontrol Proteinase Karbohidrase Proteinase+Karbohidrase Kontrol Proteinase Karbohidrase Proteinase+Kar
Energi metabolis Kcal/kg 2900 2900
Prosedur Penelitian
Aspek penelitian meliputi empat hal:
Kecernaan Asam Amino Ileum
Penelitian ini bertujuan untuk menentukan dampak penambahan enzim
pada kecernaan asam amino di dalam bagian usus halus ileum. Total perlakuan
yang dipergunakan ada 8, dengan 5 ulangan. Setiap ulangan terdiri dari 5 ekor
ayam. Ayam dipelihara hingga 32 hari. Pada umur 32 hari, ayam dipotong, dan
isi usus yang berada diantara Meckel’s diverticulum dengan perbatasan ileum-cecal-colon dikeluarkan dengan menggunakan air destilasi. Selanjutnya isi usus ditampung pada gelas specimen berukuran 50 ml. Setiap sampel merupakan
gabungan dari 5 ekor ayam.
Sampel isi usus langsung dibekukan dengan suhu -15 derajat Celcius, lalu dilanjutkan dengan proses kering beku (freeze dried), kemudian dihaluskan untuk melewati ukuran saringan 0,5 mm dan disimpan untuk analisa laboratorium
selanjutnya.
Peubah yang di amati berjumlah 6 buah:
Pakan: bahan kering, asam amino, penanda (marker)
Ayam : isi usus halus bagian ileum meliputi bahan kering, asam amino, penanda
Kecernaan asam amino ditentukan dengan rumus (Gracia et al 2007): Kecernaan AA ileum= (AA/AIA)p – (AA/AIA)i
(AA/AIA)p Keterangan:
(AA/AIA)p = rasio dari AA terhadap penanda AIA dalam pakan
(AA/AIA)i = rasio dari AA terhadap penanda AIA dalam isi usus halus
bagian ileum
Peningkatan Kecernaan Serat Kasar
Penelitian ini bertujuan untuk menentukan dampak penambahan enzim
pada kecernaan serat kasar dengan mengamati selisih jumlah serat kasar yang
terdapat di dalam usus halus dengan jumlah serat kasar yang terdapat pada
Total perlakuan yang dipergunakan ada 8, dengan 5 ulangan. Setiap
ulangan terdiri dari 5 ekor ayam. Ayam akan dipelihara hingga 32 hari.
Penampungan ekskreta dilakukan 3 hari berturut-turut sebelum ayam dimatikan.
Pada umur 32 hari, ayam dipotong, dan isi usus yang berada diantara Meckel’s
diverticulum dengan perbatasan ileal-cecal-colon dikeluarkan dengan menggunakan air destilasi. Isi usus ditampung pada gelas specimen berukuran 50
ml. Setiap sampel merupakan gabungan dari 5 ekor ayam.
Sampel isi usus langsung dibekukan dengan suhu -15 derajat Celcius, lalu dilanjutkan dengan proses kering beku, kemudian dihaluskan untuk melewati
ukuran saringan 0,5 mm dan disimpan untuk analisa laboratorium selanjutnya.
Peubah yang dianalisa sebanyak 9 buah:
Pakan : bahan kering, serat kasar, penanda (marker)
Ayam : a. isi usus halus bagian ileum meliputi bahan kering, serat kasar, penanda
b. ekskreta rataan 3 hari berturut-turut sebelum pembedahan meliputi
Perbedaan kecernaan serat kasar pada ileaum dan ekskreta
= kecernaan serat kasar ekskreta – ileum
Penelitian ini bertujuan untuk mengamati manfaat penggunaan enzim pada
perubahan kandungan air yang terdapat dalam ekskreta. Total perlakuan yang
Ayam dipelihara hingga 32 hari. Pada umur 29, 30 dan 31 hari, ekskreta dikoleksi
dan kemudian dikeringkan dengan menggunakan oven pada suhu 105 derajat
Celcius hingga mencapai berat konstan.
Peubah yang dianalisa sebanyak 2 buah
Ayam: ekskreta 3 hari berturut turut meliputiberat basah dan berat kering oven
dari ekskreta
Kadar air ditentukan dengan rumus:
Kadar air = Ba-Bk x 100% Ba
Keterangan:
Ba = berat awal sampel (g)
Bk = berat kering oven sampel (g)
Penampilan Produksi
Penelitian ini bertujuan untuk mengamati manfaat penggunaan enzim pada
penampilan produksi ayam. Total perlakuan yang dipergunakan ada 8, dengan 10
ulangan. Setiap ulangan terdiri dari 5 ekor ayam. Ayam dipelihara hingga 32
hari. Setiap minggu hingga panen, berat badan ayam dan jumlah asupan pakan
ditimbang pula. Jumlah ayam mati perhari dicatat.
Peubah yang diamati sebanyak 4 buah meliputi:
Pakan: jumlah pakan terkonsumsi mingguan dan saat panen.
Ayam: jumlah ayam mati dan berat badan mingguan dan saat panen. Untuk
selanjutnya dihitung nilai FCR.
Rancangan Percobaan dan Analisis Data
Rancangan percobaan yang digunakan dalam penelitian ini adalah Rancangan
Acak Lengkap (RAL) faktorial 2x4 dengan 8 ulangan (Steel &Torrie 1991).
Faktor A adalah perlakuan jenis kacang kedelai, yaitu :
A1 : Kacang kedelai India
Faktor B adalah perlakuan enzim, yaitu :
B1 : tanpa enzim
B2 : dengan enzim proteinase
B3: dengan enzim karbohidrase
B4: dengan enzim proteinase dan karbohidrase
Adapun model matematik rancangan tersebut adalah:
Yijk= µ + αi +βj + (αβ)ij +εijk
Yijk adalah nilai pengamatan pada faktor A (jenis kacang kedelai) taraf ke
i, faktor B (perlakuan enzim) taraf ke-j dan ulangan ke k. (µ, αi, βj) adalah
komponen aditif dari rataan, pengaruh utama faktor A dan pengaruh utama faktor
B. (αβ)ij merupakan komponen interaksi dari faktor A dan faktor B dan (εij) adalah
pengaruh acak yang menyebar normal (0,σε2). Data yang diperoleh dianalisis
dengan menggunakan sidik ragam (ANOVA), apabila berbeda nyata maka akan
HASIL PENELITIAN
Analisa Bungkil Kacang Kedelai
Hasil analisa kimiawi menunjukkan bahwa bungkil kacang kedelai (BKK)
India mempunyai kandungan protein, serat kasar dan abu yang lebih tinggi akan
tetapi mempunyai kandungan lemak yang lebih rendah bila dibandingkan dengan
BKK Argentina. Persentasi asam amino terhadap protein kasar untuk mayoritas
asam amino pada BKK India lebih rendah dari pada BKK Argentina, terutama
untuk lisina, metionina, cystein, treonina, valina dan triptofan. BKK India mempunyai kandungan urease dan daya ikat air (water holding capacity) yang lebih tinggi daripadaBKK Argentina (Tabel 4 ).
BKK India mempunyai kandungan lemak yang lebih rendah dari BKK
Argentina, sehingga kandungan energi metabolis (ME) dari BKK India lebih
rendah dari pada BKK Argentina. Nilai ME dari BKK India adalah sebesar 2315
kcal/kg sedangkan BKK Argentina sebesar 2395 kcal/kg.
Hasil dari uji mikroskopi menunjukkan bahwa warna dari pecahan bungkil
dan kulit ari (flatted kernel dan hulls) dari BKK India pucat. BKK India mempunyai lebih banyak kulit ari, batang, pasir atau silika dan benda asing bila
Tabel 4 Analisa bungkil kacang kedelai India dan Argentina
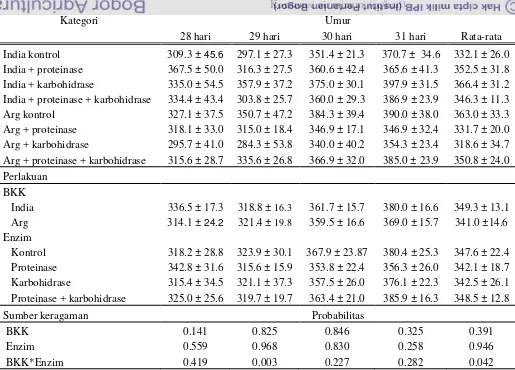
Penampilan Produksi Ayam
Penampilan produksi ayam pada perioda starter 1-21 hari
Pada perioda starter umur 0-21 hari, ayam yang diberi ransum dengan
BKK India menunjukkan berat badan yang nyata lebih rendah ( P<0.05) bila
dibandingkan dengan ayam yang diberikan ransum dengan BKK Argentina. FCR
dari ayam yang diberi ransum BKK India nyata lebih tinggi (P<0.05) bila
dibandingkan dengan ayam yang diberikan ransum dengan BKK Argentina.
Konsumsi pakan dari ayam yang diberi ransum dengan BKK Argentina tidak
berbeda nyata (P>0.05) bila dibandingkan dengan ayam yang diberikan ransum
dengan BKK Argentina. Pemberian enzim tidak menunjukkan perbedaan yang
nyata diantara ayam yang diberi ransum dengan BKK India dan Argentina dalam
hal berat badan, konsumsi pakan dan FCR. Tidak dijumpai adanya interaksi antara
BKK India dan enzim baik untuk berat badan, konsumsi pakan maupun FCR
sebagaimana disajikan pada Tabel 5, 6 dan 7.
Tabel 5 Berat badan pada periode starter 0-21 hari
Berat Badan Periode Starter
Penambahan Bungkil Kacang Kedelai
Enzim India (g) Argentina (g) Rataan (g)
Kontrol 1042.6 ± 17.9 ab 1060.7 ± 14.1 ab 1051.7 ± 15.7 a
Proteinase 1052.2 ± 9.7 ab 1064.2 ± 16.0 ab 1057.5 ± 12.1 a
Karbohidrase 1030.5 ± 9.4 a 1078.0 ± 7.7 b 1057.2 ± 7.9 a
Proteinase + karbohidrase 1046.2 ± 9.8 ab 1077.8 ± 13.02 b 1062.0 ± 10.8 a
Rataan 1043.9 ± 11.5 a 1070.6 ± 15.9 b
Tabel 6 FCR pada periode starter 0-21 hari
abc Superscript yang berbeda pada baris yang sama menunjukkan berbeda nyata (P<0.05)
Tabel 7 Konsumsi pakan pada periode starter 0-21 hari
Konsumsi Pakan Periode Starter
Penambahan Bungkil Kacang Kedelai
Penampilan produksi ayam pada perioda finisher 22-32 hari
Pada perioda finisher umur 22-32 hari, ayam yang diberi ransum dengan
BKK India menunjukkan berat badan yang tidak berbeda nyata ( P>0.05) bila
dibandingkan dengan ayam yang diberikan ransum dengan BKK Argentina. FCR
dari ayam yang diberi ransum BKK India nyata lebih tinggi (P<0.05) bila
dibandingkan dengan ayam yang diberikan ransum dengan BKK Argentina.
Konsumsi pakan dari ayam yang diberi ransum dengan BKK Argentina tidak
berbeda nyata (P>0.05) bila dibandingkan dengan ayam yang diberikan ransum
dengan BKK Argentina. Pemberian enzim tidak menunjukkan perbedaan yang
nyata diantara ayam yang diberi ransum dengan BKK India dan Argentina dalam
hal berat badan. FCR dari ayam yang diberi ransum dengan menggunakan enzim
kombinasi proteinase dan karbohidrase nyata lebih rendah ( P<0.05) bila