KEBERADAAN DAN STATUS CENDAWAN MIKORIZA
ARBUSKULA (CMA) PADA BEBERAPA VEGETASI DI
TANAH BERKAPUR
(Studi Kasus Di Desa Ria-Ria, Kecamatan Sipoholon, Kab. Tapanuli utara, Prop. Sumatera Utara)
SKRIPSI
ERWIN SAHAT L. GAOL 021202031/ Budidaya Hutan
DEPARTEMEN KEHUTANAN FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
Judul penelitian : Keberadaan dan Status Cendawan Mikoriza Arbuskula (CMA) Pada Beberapa Vegetasi di Tanah Berkapur
(Studi Kasus di Desa Ria-Ria, Kecamatan Sipoholon, Kab. Tapanuli Utara)
Nama : ERWIN SAHAT L. GAOL
NIM : 021202031
Departemen : KEHUTANAN Program Studi : BUDIDAYA HUTAN
Disetujui Oleh Komisi Pembimbing
Dr. Delvian, SP. MP Dr. Ir. Hamidah Hanum, M.Si Ketua Anggota
Mengetahui
Ketua Departemen Kehutanan
ABSTRACT
The purpose of this research was to know existence and status of Vesicular Arbuskular Mycorrhiza at some crop in chalky ground. This research has been done at Ria-ria Village, Sipoholon District, Tapanuli Utara Region and Soil Biology Laboratorium of Agriculture Faculty of North Sumatera University. This research was using soil separating method to searched spore and root coloring method to find out root colonization. The result of this research are 12 type of spore. Highest spore there at seagegrass crop (42) and lowest at vegetable crop (22). The spore to be dominated by Glomus. The percentage colonization of observed was between 58,654 - 85,19 %
ABSTRAK
KATA PENGANTAR
Puji dan syukur penulis ucapkan kehadirat Tuhan Yang Maha Kuasa yang telah memberikan berkat beserta kesempatan pada penulis, sehingga penulis dapat menyelesaikan hasil penelitian ini dengan baik. Penelitian ini berjudul “Status Dan Keberadaan Cendawan Mikoriza Arbuskula (CMA) Pada Beberapa Vegetasi di Tanah Berkapur (Studi Kasus di Desa Ria-ria Kecamatan Sipoholon, Kabupaten Tapanuli Utara, Prop. Sumatera Utara)”.
Penulis mengucapkan terima kasih yang sebesar-besarnya kepada Bapak Dr. Delvian, SP.MP, dan Ibu Dr. Ir. Hamidah Hanum, M.Si selaku Dosen Pembimbing yang telah banyak memberikan arahan dan bimbingan kepada penulis dalam menyelesaikan penelitian ini. Penulis juga tidak lupa mengucapkan terima kasih kepada rekan-rekan yang telah banyak membantu. Terutama penulis mengucapkan terima kasih kepada kedua orangtua dan seluruh keluarga yang telah memberikan dukungan.
Semoga penelitian ini dapat bermanfaat bagi masyarakat dan dunia ilmu pengetahuan. Akhir kata penulis mengucapkan terima kasih.
Medan, Juni 2008
DAFTAR ISI
Kegunaan Penelitian ... 3
TINJAUAN PUSTAKA Pengertian Dan Pembagian Mikoriza ... 4
Struktur Umum Cendawan Mikoriza Arbuskula ... 6
Distribusi Dan Ekologi Cendawan Mikoriza Arbuskula ... 8
Faktor yang Mempengaruhi Keberadaan CMA ...11
METODOLOGI PENELITIAN Tempat dan Waktu ...17
Bahan dan Alat ...17
Metode Penelitian ...18
Pelaksanaan Penelitian ...18
Ekstraksi Spora Dan Identifikasi CMA ...18
Kolonisasi CMA Pada Akar Tanaman Sampel ...20
DAFTAR TABEL
DAFTAR GAMBAR
Halaman 1. Penampang Longitudinal Akar Yang Terinfeksi CMA ...7 2. Tipe-Tipe Spora CMA Yang Ditemukan Di Tanah Berkapur
Di Desa Ria-Ria, Kabupaten Tapanuli Utara ...24 3. Grafik Kepadatan Spora CMA Yang Ditemukan
Di Tanah Berkapur Di Desa Ria-Ria, Kabupaten Tapanuli Utara ...27 4. Tipe-Tipe Infeksi CMA Pada Akar Tanaman
ABSTRACT
The purpose of this research was to know existence and status of Vesicular Arbuskular Mycorrhiza at some crop in chalky ground. This research has been done at Ria-ria Village, Sipoholon District, Tapanuli Utara Region and Soil Biology Laboratorium of Agriculture Faculty of North Sumatera University. This research was using soil separating method to searched spore and root coloring method to find out root colonization. The result of this research are 12 type of spore. Highest spore there at seagegrass crop (42) and lowest at vegetable crop (22). The spore to be dominated by Glomus. The percentage colonization of observed was between 58,654 - 85,19 %
ABSTRAK
PENDAHULUAN
Latar belakang
Salah satu cendawan pembentuk mikoriza yang sekarang sangat popular
adalah cendawan mikoriza arbuskula (CMA). Cendawan ini bersifat kosmopolitan
artinya cendawan ini tersebar dan dapat ditemukan hampir pada sebagian besar
tanah atau ekosistem dan kondisi iklim mulai dari padang pasir sampai ke
antartika, dan pada umumnya tidak mempunyai inang yang spesifik. Namun
tingkat populasi dan komposisi jenis sangat bervariasi dan dipengaruhi oleh
karakteristik tanaman dan sejumlah faktor lingkungan seperti suhu, pH,
kelembaban tanah, kandungan fosfor dan nitrogen. Suhu terbaik untuk
perkembangan CMA adalah pada suhu 30°C, tetapi untuk kolonisasi miselia yang
terbaik adalah pada suhu 28°C -34°C (Suhardi, 1989).
Meskipun CMA tersebar hampir merata di seluruh permukaan bumi,
namun keberadaannya ditentukan oleh berbagai faktor lingkungan dan tanah.
Distribusi dan kemelimpahan CMA berhubungan erat dengan kandungan hara dan
ketersediaan air tanah, ketinggian tempat, temperatur, dan beberapa sifat kimia
tanah antara lain pH tanah. Keberadaan dari mikroflora tanah, organisme tanah
lain juga dapat berpengaruh.
Cendawan mikoriza arbuskula banyak tersebar di lahan-lahan pertanian
maupun kehutanan pada berbagai ekosistem. Melalui kolonisasi akar, mikoriza
membantu tanaman untuk menyerap berbagai unsur hara yang immobil di dalam
tanah terutama unsur P, di samping memberi manfaat potensial lainnya.
yang tersedia di lapangan. Propagul ini dapat berupa spora, akar terinfeksi, dan
hifa. Pada kondisi alam yang mencekam seperti kekeringan misalnya, makaspora
ini merupakan satu-satunya yang dapat bertahan hidup, maka jumlah spora per
satuan bobot atau volume tanah kering merupakan gambaran besarnya propagul
CMA di lapangan. Dengan kemampuan CMA ini hidup pada berbagai kondisi
tanah seperti kondisi alam yang mencekam ini maka CMA banyak digunakan
dalam bidang kehutanan dan juga pertanian.
Dalam kesempatan ini penelitian dilakukan pada tanah berkapur yang
terdapat di daerah Ria-ria, Sipoholon dimana secara administratif terletak di
wilayah kecamatan Sipoholon dan Tarutung, kabupaten Tapanuli Utara. Tanah
kapur ini dibentuk di atas bahan induk yang mengandung proporsi CaCO3 yang
tinggi (batu kapur, kapur, dan beberapa bahan lain) serta endapan glasial
cenderung mempunyai nilai pH sekitar 7 atau lebih, walaupun selama
pembentukan tanah ion-ion Ca 2+ akan tercuci, yang akibatnya secara bertahap
akan menurunkan pH (Fitter dan Hay, 1991).
Tanah berkapur memiliki sifat-sifat yang khas yang membedakannya
dengan jenis tanah lain, yang akan mempengaruhi keberadaan dan kelimpahan
CMA pada lokasi penelitian. Sifat-sifat tersebut antara lain sebaran temperatur
suhu tanah berkisar 21-29°C, pH normal < 7,5, anomali pH > 7,5, kandungan CO2
dalam tanah berkisar antara 0,07-4,5 %, warna tanah yang terang, tanaman yang
tumbuh pada tanah ini kadang-kadang kekurangan besi, mangan, seng, tembaga,
dan boron (Fitter dan Hay, 1991).
Sifat dari tanah berkapur antara lain memiliki kandungan kapur CaO yang
kimia dan biologi tanah yaitu untuk menaikkan kadar Ca dan pH tanah, sehingga
reaksi tanah mengarah ke netral. Dengan naiknya pH tanah dan tersedianya
beberapa hara yang dibutuhkan biologi tanah menyebabkan jasad hidup ini lebih
mudah memperoleh energi dan materi dalam jumlah yang banyak. Sejalan dengan
hal itu, populasi dan aktivitas mereka pun meningkat dengan penambahan kapur
(Hakim dkk, 1986).
Dengan melihat uraian di atas maka dilakukan penelitian pada tanah
berkapur yang bertujuan untuk mendapatkan informasi mengenai status dan
keberadaan CMA pada berbagai jenis tanaman yang tumbuh pada tanah berkapur.
Melalui penelitian ini diharapkan dapat ditemukan dan diketahui jenis CMA yang
dominan pada tanah berkapur.
Tujuan Penelitian
Untuk mengetahui keberadaan dan status CMA pada beberapa jenis
vegetasi pada tanah berkapur di Desa Ria-Ria Kecamatan Sipoholon, Kabupaten
Tapanuli utara.
Kegunaan Penelitian
Memberikan informasi bagi pihak-pihak yang membutuhkan mengenai
status dan keberadaan CMA di tanah berkapur di Desa Ria-Ria Kecamatan
TINJAUAN PUSTAKA
Pengertian dan Pembagian Mikoriza
Kata mikoriza terdiri dari dua kata yang berasal dari bahasa Yunani, yaitu
Mykes (cendawan) dan Rhiza (akar). Kata mikoriza pertama kali dikemukakan
oleh Frank (1885) yang melihat cendawan di dalam sel akar tumbuhan yang
ditelitinya di Jawa antara lain akar jati. Mikoriza secara harfiah berarti cendawan
akar. Dalam konteks ini merupakan kandungan simbiotik dan mutualistik
menguntungkan antara cendawan non patogen dengan sel-sel akar yang hidup,
terutama sel epidermis dan korteks.Frank membedakannya menjadi dua golongan
yakni ekto dan endomikoriza. Kebanyakan simbiosis yang tersebar luas diantara
tanaman adalah mikoriza yang meliputi berbagai macam cendawan yang
menempati akar dan akar penyerap (Fakuara, 1988; Suhardi, 1989).
Cendawan mikoriza arbuskula termasuk golongan endomikoriza
merupakan cendawan yang bersimbiosis dengan akar tanaman. Cendawan ini
membentuk vesikula dan arbuskula di dalam korteks tanaman. Maka cendawan ini
disebut juga dengan cendawan mikoriza vesikula arbuskula. Vesikula merupakan
ujung hifa berbentuk bulat, berfungsi sebagai organ penyimpanan, sedangkan
arbuskula merupakan hifa yang berperan dalam transfer hara dari tanah ke
tanaman. Berdasarkan struktur tumbuh dan cara infeksinya pada sistem perakaran
inang (host), mikoriza dikelompokkan ke dalam 2 golongan besar yaitu
ektomikoriza dan endomikoriza (Setiadi, 2000).
Cendawan ini memperoleh senyawa organik (terutama gula) dari tanaman,
air dapat berlangsung lebih baik. Bagian sistem perakaran tanaman yang terinfeksi
adalah bagian akar yang masih muda. Cendawan mikoriza ini mengharapkan
karbohidrat dari tanaman, maka untuk tanaman yang ternaungi (sehingga hasil
fotosintesisnya rendah) akan sedikit mikoriza yang terbentuk pada sistem
perakarannya. Juga tanaman yang tumbuh pada tanah-tanah yang subur, tidak
dijumpai banyak mikoriza pada sistem perakarannya (Haryantini dan Santoso,
2003).
Cendawan mikoriza arbuskula adalah salah satu tipe cendawan mikoriza
dan termasuk kedalam golongan endomikoriza. Cendawan mikoriza arbuskula
termasuk kedalam kelas Zygomycetes, dengan ordo Glomales. Terdiri dari dua sub
ordo yaitu Gigasporineae dan Glomineae. Gigasporineae dengan famili
Gigasporaceae mempunyai dua genus yaitu Gigaspora dan Scutelospora.
Glomineae mempunyai empat famili, yaitu Glomaceae dengan genus Glomus,
famili Acaulospora dengan genus Acaulospora dan Entrophospora,
Paraglomaceae dengan genus Paraglomus, dan Archaeosporaceae dengan genus
Archaeospora. Terdapat sekitar 150 jenis (spesies) spora cendawan CMA yang
telah dideskripsi (INVAM, 2004).
Cendawan mikoriza arbuskula tergolong dalam kelompok khusus dari
populasi mikoriza yang sangat banyak mengkolonisasi rizosfer, yaitu di dalam
akar, permukaan akar, dan di daerah sekitar akar. Hifa eksternal yang
berhubungan dengan tanah dan struktur infeksi seperti arbuskula di dalam akar
menjamin adanya perluasan penyerapan unsur-unsur hara dari tanah dan
peningkatan transfer hara (khususnya P) ke tumbuhan, sedangkan cendawan
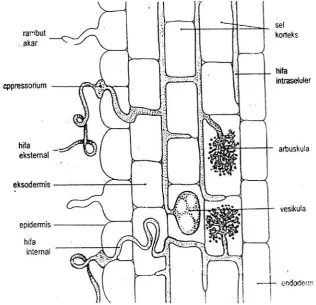
Struktur Umum Cendawan Mikoriza Arbuskula
Endomikoriza dicirikan oleh hifa yang intraseluler, yaitu hifa yang
menembus kedalam sel-sel korteks dan dari sel yang satu ke sel yang lain. Di
antara sel-sel terdapat hifa yang membelit atau struktur hifa yang
bercabang-cabang yang disebut arbuskula dan pembengkakan yang terbentuk pada hifa yang
mengandung minyak yang disebut vesikula. Vesikula berfungsi sebagai tempat
penyimpanan yang diameternya lebih kecil dari 1mm, sedangkan arbuskula
tersebut berperan dalam transfer hara dari tanah ketanaman untuk membebaskan
karbon (C) dan posphor (P) dan fungsinya yang lebih utama adalah menyediakan
unsur tersebut menjadi tersedia yang dapat dimanfaatkan oleh tanaman. Walaupun
seolah-olah cendawan ini melakukan penetrasi langsung kedalam sitosol sel
korteks membentuk struktur vesikula-arbuskula, tetapi sesungguhnya terdapat
pembatas antara vesikula dan arbuskula dengan sitosol sel korteks yakni suatu
membran plasma (Fitter dan Hay, 1991; Lakitan, 2000).
Cendawan mikoriza arbuskula dapat dibedakan dari ektendomikoriza. Hal
ini dapat kita perhatikan bahwa sistem perakaran yang terinfeksi tidak membesar,
cendawannya membentuk struktur lapisan hifa tipis dan tidak merata pada
permukaan akar, hifa menyerang ke dalam individu sel jaringan korteks, pada
umumnya ditemukan struktur khusus berbentuk oval yang disebut vesikel
Ciri utama arbuskular mikoriza adalah terdapatnya arbuskula di dalam
korteks akar. Awalnya cendawan tumbuh di antara sel-sel korteks, tetapi
kemudian segera menembus dinding sel inang dan berkembang di dalam sel.
Istilah umum bagi semua tipe mikoriza yang cendawannya berkembang di dalam
sel korteks adalah endomikoriza, sebagaimana perkembangan cendawan, sel
membran inang membungkus cendawan (Brundrett dkk, 1996).
Struktur umum dari CMA dapat dilihat pada Gambar 1. dibawah ini:
Gambar 1. Penampang Longitudinal Akar yang Terinfeksi CMA (Sumber : Brundett et al. 1996)
Menurut Abbot dan Robson (1984), infektivitas mikoriza adalah sebagai
daya atau kemampuan cendawan mikoriza untuk menginfeksi dan mengkoloni
akar tanaman. Dalam pengertian itu dicakup perkembangan dan aktivitas
juga diacu sebagai ukuran suksesi dari cendawan dalam berinteraksi dengan
tanaman dan kondisi tanah. Infektivitas dalam hal ini dinyatakan sebagai proporsi
akar tanaman yang terinfeksi (Giovanetti dan Mosse, 1980).
Peranan Cendawan Mikoriza Arbuskula
Secara umum peranan mikoriza terhadap pertumbuhan tanaman adalah :
adanya mikoriza sangat penting bagi ketersediaan unsur hara dan pertumbuhan
tanaman dimana hal ini terjadi melalui pembentukan hifa pada permukaan akar
yang berfungsi sebagai perpanjangan akar terutama di daerah yang kondisinya
sangat miskin unsur hara, pH rendah, dan kurang air. Peranan yang lain yaitu
melindungi tanaman inang terhadap cendawan patogen akar dengan cara
mengeluarkan zat antibiotik, menghasilkan hormon tumbuh auksin, sitokinin,
giberelin, dan vitamin yang dapat merangsang pertumbuhan tanaman inang. Zat
pengatur tumbuh ini sangat diperlukan dalam proses pembelahan sel, memacu
pertumbuhan serta mencegah atau memperlambat proses penuaan daun sehingga
menambah fungsi akar sebagai penyerap unsur hara dan air. Manfaat cendawan
ini akan secara nyata terlihat jika kondisi tanahnya miskin hara atau kering;
sedangkan pada kondisi tanah yang subur, peran cendawan mikoriza ini tidak
akan begitu nyata (Setiadi, 2001; Lakitan, 2000).
Keuntungan dari keberadaan mikoriza yang telah banyak diketahui adalah
meningkatkan serapan fosfat oleh tanaman walaupun sesungguhnya serapan
unsur-unsur hara yang lain dan air juga ikut meningkat. Keuntungan tersebut
berdifusi secara lambat ke permukaan akar, tetapi dibutuhkan dalam jumlah besar
oleh tanaman, misalnya fosfat, ammonium, kalium, dan nitrat ( Lakitan, 2000).
Angkutan nutrien merupakan sentral mutualisme mikoriza, dimana nutrien
diangkut CMA ke tumbuhan dan C organic dari tumbuhan ke cendawan. Ada dua
tipe hifa eksternal pada CMA yaitu absorber hypae (hifa-hifa besar) dan runner
hypae (hifa-hifa dengan diameter yang lebih besar, absorber hypae merupakan
kunci dalam absorpsi nutrien sedangkan runner hypae merupakan jalur utama
translokasi di dalam miselium eksternal) (Smith dan Read, 1997 dalam
Ekamawanti, 1998).
Perbaikan hasil tanaman dengan isolasi CMA juga berkaitan dengan
perbaikan mikoriza itu sendiri. Meski pada beberapa kasus isolasi cendawan ini
meningkatkan hasil tanaman tapi gagal memperbaiki status mikoriza. Pada saat
yang sama pada beberapa kasus isolasi gagal memperbaiki hasil tanaman,
walaupun mampu meningkatkan kondisi mikoriza (Kehri and Chandra, 1990).
Peranan mikoriza tersebut secara spesifik dalam membantu pertumbuhan
tanaman antara lain membantu memperbaiki nutrisi tanaman dengan
meningkatkan serapan hara terutama fosfor, sebagai pelindung hayati dengan
melindungi akar dari patogen akar, mencegah tanaman terhindar dari keracunan
logam berat, serta membantu meningkatkan resistensi tanaman terhadap
kekeringan (Muin, 2004; Hanafiah dkk, 2003).
Secara fisik pada tanah yang dikatakan subur terdapat sejumlah besar
agregat, baik makro ataupun mikro yang stabil. Hifa eksternal CMA yang
berkembang kedalam tanah dapat mengikat partikel-partikel tanah dan
banyak dibandingkan tanaman yang bermikoriza. Pembentukan agregat tanah
yang stabil dengan CMA merupakan faktor penting dalam meningkatkan
kesuburan fisik tanah (Baon, 1998).
Distribusi dan Ekologi Cendawan Mikoriza Arbuskula
Cendawan mikoriza arbuskula merupakan cendawan tanah yang
digolongkan kedalam famili Endogonaceae, cendawan ini dapat bersimbiosis
dengan akar banyak jenis tanaman. Dan ciri paling menonjol dari tipe CMA
dibanding jenis lainnya adalah kemampuannya bersimbiosis dengan hampir 90 %
jenis tanaman. CMA pada umumnya dapat ditemukan pada sebagian besar spesies
tanaman tingkat tinggi yang tumbuh pada berbagai habitat dengan berbagai
macam iklim. Sebaran spesies CMA bervariasi menurut iklim, lingkungan dan
tipe penggunaan lahan (Setiadi, 2001).
Lingkungan dan faktor biotik diketahui memiliki pengaruh terhadap
pembentukan CMA dan derajat infeksi dari sel korteks inang. Perbedaan waktu
yang diperlukan untuk infeksi tersebut disebabkan oleh beberapa faktor antara
lain: kerapatan akar, rata-rata pertumbuhan akar, jumlah spora / unit volume
tanah, persentase perkecambahan spora dan rata-rata pertumbuhan hifa. Interaksi
antara faktor-faktor biotik memiliki efek yang signifikan dalam merespon
pertumbuhan tanaman yang diinokulasi. Faktor lingkungan berpengaruh terhadap
pembentukan CMA dalam hal suplai dan keseimbangan hara, kelembaban dan pH
tanah (Rao, 1994).
Peningkatan persentase infeksi CMA akibat inokulasi dapat dihubungkan
eksudat atau senyawa khas yang dihasilkan dan dikeluarkan oleh akar tanaman
yang menyebabkan perkembangan CMA terangsang. Peningkatan persentase
infeksi akibat inokulasi ini sangat dipengaruhi oleh rhizobium (Fakuara, 1988).
Perbedaan lokasi dan rhizosfer menyebabkan perbedaan keanekaragaman
spesies dan populasi CMA. Tanah yang didominasi oleh fraksi lempung (clay)
merupakan kondisi yang diduga sesuai untuk perkembangan spora Glomus, dan
tanah berpasir genus Gigaspora ditemukan dalam jumlah tinggi. Pada tanah
berpasir, pori-pori tanah terbentuk lebih besar dibanding tanah lempung dan
keadaan ini diduga sesuai untuk perkembangan spora Gigaspora yang berukuran
lebih besar daripada spora Glomus (Baon, 1998).
Suksesi CMA umumnya dipengaruhi oleh umur tanaman, infeksi yang
tertinggi terutama pada akar tanaman muda, dan faktor lain yang
mempengaruhinya adalah faktor lingkungan seperti cahaya, kesuburan tanah
(unsur P dan N tersedia), kadar air, drainase tanah dan pH tanah. Di samping itu
CMA dapat berkembang dengan baik pada tanah yang mempunyai kandungan P
lebih rendah dan aerasi tanah yang lebih baik (Husien dkk., 2000).
Penyebaran dari CMA dapat terjadi melalui berbagai cara, penyebaran
aktif micelya melalui tanah. Penyebaran mikoriza melalui inokulasi agak
berkurang pada tanah yang sudah bermikoriza, tetapi meningkat pada tanah yang
tidak bermikoriza. Dengan kata lain, CMA baru akan tersebar pada tanah-tanah
yang kandungan mikorizanya rendah. Sedangkan penyebaran pasif dapat
dilakukan oleh beberapa binatang dan juga oleh angin (Suhardi, 1989; Setiadi,
Faktor-Faktor Yang Mempengaruhi Perkembangan dan Penyebaran CMA
Menurut Suhardi (1989), faktor-faktor yang mempengaruhi perkembangan
dan penyebaran CMA adalah sebagai berikut :
A. Pengaruh dari Mikro Flora Tanah
Perkecambahan dari spora CMA invitro sulit untuk diduga. Pada media
agar terdapat faktor yang menghambat perkecambahan dan menyebabkan
dormansi. Daniels dan Trappe (1980) menemukan bahwa ternyata spora dari
Glomus epigaeum gagal berkecambah pada tanah yang disterilkan dengan
autoclaf, dipanaskan dengan uap, diberi radiasi sinar gamma sebaliknya
perkecambahan yang tinggi terdapat pada tanah-tanah yang non steril.
Faktor-faktor penghambat yang hilang setelah sterilisasi ini justru merangsang
perkecambahan pada tanah non steril.
B. Pengaruh dari Faktor Lingkungan
1. Potensial Air Dari Tanah
Percobaan-percobaan telah dilakukan pada tanah-tanah dengan
berbeda-beda kadar airnya. Daniels dan Trappe (1980) menggunakan Glomus epigaeum
dikecambahkan pada debu geluh (silt loam) pada berbagai kandungan air. Glomus
epigaeum ternyata berkecambah paling baik pada kandungan air di antara
kapasitas lapang dan kandungan air jenuh.
Perkecambahan Gigaspora spp. akan menurun pada kondisi potensial air
(-31 bar) yang rendah juga ini mempengaruhi pertumbuhan hifanya. Pada -10 bar
Mikoriza akan berkembang baik bila tidak ada hambatan aerase. Oleh
karena itu mikoriza akan dapat berkembang lebih baik pada tanah berpasir
dibandingkan tanah berliat atau gambut (Islami dan Wani, 1995).
2. Kandungan Nutrien Di Dalam Tanah
Perkecambahan dari spora tidak begitu dipengaruhi oleh kesuburan tanah.
Penambahan nitrogen dan potasium ternyata dapat merangsang atau menghambat
perkecambahan. Penambahan fosfor dapat meningkatkan pertumbuhan/
perkecambahan tetapi penambahan nitrogen dan potasium kurang ada
pengaruhnya.
Bahan organik dapat mengurangi kemampuan berkecambah dari spora
baik pada medium agar maupun pada media tanah tidak steril, kelihatannya
dengan jalan mempengaruhi langsung terhadap perkecambahan spora. Pengaruh
tidak langsung yaitu dengan menggiatkan kegiatan mikrobia tanah untuk
kemudian menghambat perkecambahan spora.
Ketersediaan hara terutama nitrogen dan fosfat yang rendah akan
mendorong pertumbuhan mikoriza. Akan tetapi kandungan hara yang terlalu
rendah atau tinggi menghambat pertumbuhan mikoriza. Perkecambahan spora
tidak hanya bergantung pada spesies dari CMA tetapi juga kandungan nutrien di
dalam tanah ( Islami dan Wani, 1995).
3.pH Tanah
Tidak sama dengan jasad renik lainnya cendawan pada umumnya lebih
jenis spesies cendawan terhadap pH tanah berbeda-beda, karena pH tanah
mempengaruhi perkecambahan, perkembangan dan peran mikoriza terhadap
pertumbuhan tanaman (Suhardi, 1989).
Untuk perkecambahan spora, pH optimum dapat berbeda-beda untuk
masing-masing spesies CMA dan untuk lingkungan yang berbeda pula. Derajat
keasaman (pH) optimum untuk perkecambahan spora tidak hanya tergantung pada
spesies cendawan tetapi juga kandungan nutrien di dalam tanah. Derajat keasaman
berpengaruh langsung terhadap aktivitas enzim yang berperan dalam
perkecambahan spora (Abbot dan Robson, 1984).
Derajat keasaman optimum untuk perkecambahan spora berbeda-beda
untuk masing-masing spesies CMA dan untuk lingkungan yang berbeda-beda.
Misalnya untuk Glomus mosseae biasanya pada tanah alkali dapat berkecambah
dengan baik pada air atau pada pH 6 sampai 9. Sedangkan spora dari Gigaspora
coralloidea dan G. Heterogama dari jenis yang lebih asam dapat berkecambah
dengan baik pada pH 4 sampai 6, Glomus epigaeum perkecambahannya lebih baik
pada pH 6 sampai dengan 8 (Abbot dan Robson, 1984).
4. Suhu Udara
Baik suhu maupun sinar menunjukkan pengaruh nyata terhadap koloni dan
perkembangan spora CMA di dalam rumah kaca. Semakin tinggi suhu semakin
besar terbentuknya kolonisasi dan meningkatnya produksi spora. Suhu terbaik
untuk perkembangan CMA adalah pada suhu 30°C, tetapi untuk kolonisasi
5. Kesuburan Tanah
Sudah merupakan kesepakatan bahwa kolonisasi akar yang maksimum
akan dicapai pada tanah yang kurang subur kondisinya. Fakuara (1988)
menyatakan bahwa tingginya persentase infeksi pada tanah-tanah yang tidak subur
mungkin sebagian besar dipengaruhi oleh terbatasnya pertumbuhan akar dan
mungkin total infeksi di dalam tanah-tanah subur lebih banyak dimana persentase
panjang akar yang diinfeksi lebih rendah. Jumlah spora dapat dihubungkan
dengan infeksi akar, pada umumnya pada waktu spora membentuk miselium di
sekeliling akar yang menghambat perkembangan miselium bagian luar atau
pertumbuhan akar dihambat oleh miskinnya suplai unsur hara. Spora lebih banyak
pada tingkat fosfat sedang daripada tingkat fosfat rendah, jika kekurangan fosfat
dapat membatasi pertumbuhan spora, dan mempengaruhi keseluruhannya.
Pembentukan spora umumnya berkurang dengan perlahan-lahan atau
pertumbuhan akar berhenti dan pada musim ini spora dapat dihitung untuk
mengetahui tingkat infeksi CMA.
6. Tanaman Inang
Ada tidaknya tanaman inang yang sesuai ternyata berpengaruh terhadap
ada atau tidaknya koloni CMA dan produksi sporanya. Cendawan mikoriza
arbuskula sebenarnya bukan merupakan jenis cendawan yang sangat spesifik pada
tanaman inang tertentu. Mungkin satu spesies dapat lebih efisien membentuk
asosiasi pada tanaman tertentu, tetapi sebenarnya cendawan CMA dapat
membentuk koloni pada hampir setiap tanaman inang. Pada penelitian lebih lanjut
pada jenis tanaman inang yang terbatas saja serta kondisi lingkungan yang khusus,
tetapi ini hanya merupakan kekecualian saja. Cendawan mikoriza arbuskula ini
diketahui berasosiasi dengan berbagai macam tanaman angiospermae, baik dikotil
maupun monokotil, tanaman tahunan atau semusim dan tanaman-tanaman lokal
(Fitter dan Hay, 1991; Lakitan 2000).
C. Pengaruh dari Organisme Tanah yang Lain
Spora dapat mensuplai kebutuhan nutrien organisme di dalam tanah.
Binatang di dalam tanah seperti semut atau colembola, dan cacing dapat memakan
cendawan atau spora dari cendawan CMA. Nematoda mungkin juga dapat
memparasiti spora namun penelitian ini belum banyak dilakukan. Bakteri juga
dapat masuk ke dalam spora, biasanya mereka mula-mula hanya berada pada
permukaan dari spora dan kemudian menghasilkan enzim penghancur dinding sel
untuk ke dalam protoplasma dari cendawan. Cendawan arbuskula mungkin
menjadi kurang aktif apabila disitu terdapat juga colembola (springtail). Adanya
serangga yang hidup di dalam tanah juga akan mengurangi populasi CMA di
METODOLOGI PENELITIAN
Tempat dan Waktu Penelitian
Penelitian ini dilakukan di laboratorium Biologi Tanah Fakultas Pertanian
Universitas Sumatera Utara. Pengambilan sampel akar tanaman dan tanah
dilakukan di desa Ria-ria, Kecamatan Sipoholon, Kabupaten Tapanuli Utara.
Secara geografis terletak pada koordinat antara 98° 54’ 00’’ - 99° 01’ 30” BT dan
1° 56’ 30” – 2° 06’ 00” LS. Penelitian ini dilaksanakan mulai bulan Januari
sampai dengan April 2007.
Bahan Dan Alat
Bahan Penelitian
Bahan yang digunakan untuk prosedur teknik tuang saring dan teknik
sentrifugasi adalah sampel tanah, glukosa 60%, larutan pewarna Melzer’s reagent,
dan larutan PVLG. Sedangkan untuk kolonisasi CMA bahan yang digunakan
adalah KOH 2,5%, HCL 2%, larutan staining (Trypan blue) 0,05%, larutan
destaining (lacto glycerol), kutex (nail polish).
Alat Penelitian
Alat yang digunakan dalam penelitian ini adalah: kamera, kompas, tali,
meteran, sekop, kantong plastik, kertas label, cangkul, parang, saringan test sieve
710µ m; 215 µ m; 50 µ m, tabung sentrifuge, pipet tetes, cawan Petri, mikroskop
binokuler dan mikroskop compound, pinset spora, tabung reaksi, kaca preparat,
Metode Penelitian
Tehnik pengambilan sampel tanah dan tanaman dilakukan dengan
membuat petak pengamatan yang berukuran ± 20 m x 20 m (sesuai dengan
keadaan di lapangan). Dalam penelitian ini ada tiga jenis vegetasi yang akan
diambil contoh tanah dan tanamannya yaitu vegetasi alang-alang, umbi-umbian
dan sayur-sayuran. Setiap satu jenis vegetasi dibuat tiga petak pengamatan dimana
petak pengamatan ini bebas diletakkan dimana-mana dengan asumsi bahwa
vegetasinya sama. Sehingga jumlah petak pengamatan yang akan dibuat
berjumlah 9. Pada masing-masing petak diambil contoh tanah dari zona rhizosfer
pada kedalaman 0-20 cm dan contoh tanaman sebanyak tiga tanaman dari setiap
petak. Pengambilan contoh tanah dilakukan secara komposit dengan membuat
lima titik atau pembuatan titik secara zig-zag pada setiap petak dan diambil
contoh tanah dari tiap titik kira-kira 1 kg. Sehingga dalam satu petak ada sebanyak
5 kg tanah kemudian tanah yang dari 5 titik tadi dicampur dan diambil 1 kg untuk
dijadikan sebagai sampel tanah yang akan diteliti.
Pelaksanaan Penelitian
Ekstraksi Spora dan Identifikasi Cendawan Mikoriza Arbuskula.
Ekstraksi spora CMA dilakukan untuk memisahkan spora CMA dari
sampel tanah sehingga dapat dilakukan identifikasi guna mengetahui jumlah dan
genus spora CMA yang terdapat pada setiap petak contoh. Teknik yang digunakan
dalam mengekstraksi spora CMA adalah teknik tuang-saring dan dilanjutkan
Prosedur teknik tuang saring dan teknik sentrifugasi secara lengkap adalah
sebagai berikut :
1. 10 gram tanah sampel dituangkan dalam gelas piala, ditambahkan air 500
ml dan diaduk, dibiarkan selama 30 menit agar partikel-partikel besar
mengendap.
2. Campuran tanah sampel dengan air tersebut disaring dalam satu set
saringan dengan ukuran 710µ m; 215 µ m; 50 µ m secara berurutan dari atas
kebawah. Partikel yang tertahan dalam saringan yang paling bawah
tersebut dicuci dengan air mengalir, dan dipindahkan ke dalam tabung
sentrifuge.
3. Ke dalam tabung sentrifuge ditambahkan larutan glukosa 60%.
4. Tabung sentrifuge ditutup rapat dan disentrifuge dengan kecepatan 2500
rpm selama 5 menit, sehingga spora cendawan akan mengapung dan tanah
akan mengendap dibawah.
5. Cairan yang bening dalam tabung sentrifuge dituang ke dalam saringan
50µ m, dan dicuci dengan air mengalir dan dipindahkan ke cawan petri dan
kemudian diperiksa di bawah mikroskop binokuler.
6. Selanjutnya spora-spora yang diperoleh dipisahkan ke dalam gelas arloji
dengan menggunakan pinset spora.
7. Identifikasi jenis spora dilakukan melalui pengamatan preparat spora
Kolonisasi CMA pada Akar Tanaman Sampel
Kolonisasi akar ditandai dengan adanya hifa, vesikula dan arbuskula atau
salah satu dari ketiganya. Setiap bidang pandang (field of view) mikroskop yang
menunjukkan tanda kolonisasi diberi symbol (+) dan yang tidak diberi tanda (-).
Pengamatan kolonisasi CMA pada akar tanaman sampel dapat dilakukan melalui
teknik pewarnaan akar (staining akar), karena karakteristik anatomi yang
mencirikan ada tidaknya infeksi CMA tidak dapat dilihat secara langsung. Metode
yang digunakan dalam pewarnaan akar sampel adalah metode pewarnaan dari
Kormanik dan Mc.Graw (1982) dalam Delvian (2003), yang secara lengkap
adalah sebagai berikut:
1. Dipilih akar-akar segar dan dicuci dengan air mengalir sampai bersih,
sampel akar direndam dalam larutan KOH 10 % selama 12 jam.
2. Larutan KOH kemudian dibuang dan akar dicuci pada air mengalir selama
5-10 menit.
3. Sampel akar direndam dalam larutan HCL 2 % selama 24 jam dan pada
proses ini akar akan berwarna pucat atau putih. Larutan HCL 2 %
kemudian dibuang dengan mengalirkannya secara perlahan-lahan.
4. Selanjutnya akar sampel direndam dalam larutan staining (Trypan blue
0.05 %) selama 24 jam untuk proses pewarnaan akar.
5. Larutan trypan blue 0.05 % kemudian dibuang dan diganti dengan larutan
lacto glycerol untuk proses destaining atau pengurangan warna.
6. Secara acak diambil potong-potongan akar yang telah diwarnai dengan
panjang ± 1 cm sebanyak 10 potongan akar dan disusun pada kaca
7. Diletakkan kaca penutup (cover glass) diatas potongan akar kenudian
dengan menggunakan ujung lidi yang tumpul ditekan potongan akar secara
perlahan-lahan sehingga potongan akar menjadi lembaran tipis.
8. Sehingga kegiatan pengamatan siap dilakukan.
9. Perhitungan persentase kolonisasi akar menggunakan metode panjang
slide dari Giovanetti dan Mosse (1980) dalam Delvian (2003). Secara acak
diambil potongan-potongan akar yang telah diwarnai dengan panjang ± 1
cm sebanyak 10 potongan akar dan disusun pada preparat slide. Persentase
kolonisasi akar dihitung dengan menggunakan rumus:
% Kolonisasi =
Bidang pandang bertanda (+) = Setiap bidang pandang yang menunjukkan adanya
kolonisasi yang ditandai dengan adanya hifa, vesikula ataupun arbuskula.
Bidang pandang bertanda (-) = Setiap bidang pandang yang tidak menunjukkan
adanya kolonisasi.
Penentuan status mikoriza didasarkan pada tabel menurut Setiadi et al, (1992)
HASIL DAN PEMBAHASAN
Hasil
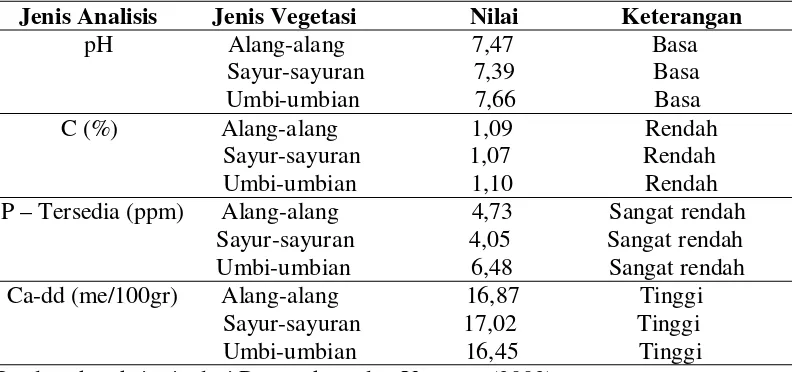
Hasil analisis tanah yang dilakukan pada setiap jenis vegetasi dapat kita
lihat pada Tabel 1.
Tabel 1. Hasil analisis tanah pada setiap jenis vegetasi
Jenis Analisis Jenis Vegetasi Nilai Keterangan Berdasarkan kriteria dari Rosmarkam dan Yuwono (2002).
Dari hasil analisis tanah tersebut dapat kita lihat bahwa untuk pH dari
ketiga jenis vegetasi tersebut tergolong tinggi (basa) karena tanah tersebut
mengandung kadar kapur (Ca) yang tinggi dimana sifat tanah berkapur adalah
memiliki pH 7 atau lebih. Sedangkan untuk C-organik memiliki nilai yang rendah
sehingga dapat kita katakan bahwa tanah ini kurang subur, begitu juga dengan
P-tersedia juga memiliki nilai sangat rendah dimana nilai dari ketiga vegetasi masih
dalam satu harkat.
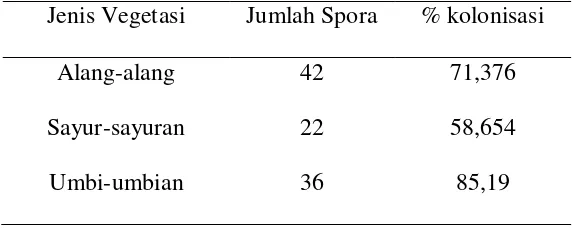
Hasil pengamatan spora dan persen kolonisasi akar dapat dilihat pada
Tabel 2. Hasil pengamatan spora dan persen kolonisasi akar
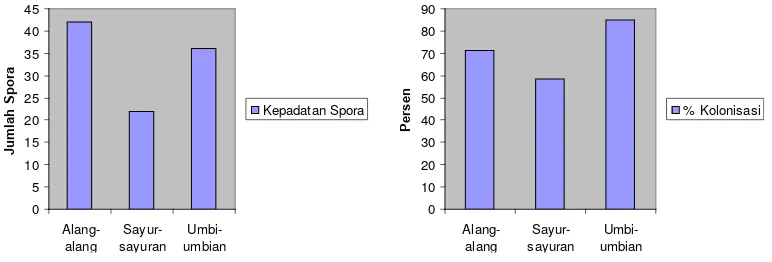
Jenis Vegetasi Jumlah Spora % kolonisasi
Alang-alang 42 71,376
Sayur-sayuran 22 58,654
Umbi-umbian 36 85,19
Dari data spora di atas dapat diketahui bahwa jumlah spora tertinggi
terdapat pada vegetasi alang-alang dan yang terendah pada vegetasi
sayur-sayuran. Untuk persen kolonisasi yang tertinggi terdapat pada vegetasi
umbi-umbian dan yang terendah terdapat pada vegetasi sayur-sayuran. Gambar spora
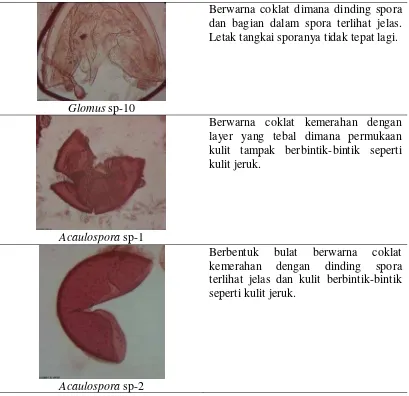
Tipe-tipe spora dari CMA yang ditemukan pada lokasi penelitian
ditampilkan pada Gambar 2.
Jenis Spora CMA Karakteristik Umum
Glomus sp-1
Berwarna merah kecoklatan dengan layer yang terlihat jelas dengan permukaan kulit yang berbintik lengkap dengan tangkai spora.
Glomus sp-2
Berbentuk bulat, berwarna coklat tua dengan dinding spora yang cukup tebal dimana di bagian dalam terdapat bulatan yang berwarna hitam.
Glomus sp-3
Berwarna merah tua sampai kecoklatan dengan dinding spora yang jelas dimana struktur spora tampak nyata bagian-bagiannya dan disertai dengan tangkai spora.
Glomus sp-4
Glomus sp-5
Berbentuk bulat dengan warna merah tua lengkap dengan tangkai spora. Dinding spora terlihat jelas.
Glomus sp-6
Berwarna merah tua kecoklatan dengan selaput berwarna hitam yang hampir menutupi seluruh permukaan spora.
Glomus sp-7
Berwarna merah kecoklatan dengan layer yang terlihat jelas dengan permukaan kulit yang berbintik lengkap dengan tangkai spora.
Glomus sp-8
Berwarna merah kecoklatan dengan layer yang terlihat jelas lengkap dengan tangkai. Sekilas tampak seperti Glomus sp-7 namun permukaan kulit lebih halus dan transparan.
Glomus sp-9
Glomus sp-10
Berwarna coklat dimana dinding spora dan bagian dalam spora terlihat jelas. Letak tangkai sporanya tidak tepat lagi.
Acaulospora sp-1
Berwarna coklat kemerahan dengan layer yang tebal dimana permukaan kulit tampak berbintik-bintik seperti kulit jeruk.
Acaulospora sp-2
Berbentuk bulat berwarna coklat kemerahan dengan dinding spora terlihat jelas dan kulit berbintik-bintik seperti kulit jeruk.
Gambar 2. Tipe-tipe Spora CMA yang Ditemukan Di Tanah Berkapur di Desa Ria-Ria Kabupaten Tapanuli Utara.
Dari hasil penyaringan spora di dapat 12 jenis spora yang didominasi oleh
genus Glomus yaitu Glomus sp-1 sampai Glomus sp-10. Genus lain yaitu
Acaulospora yang terdiri dari 2 jenis spora yaitu Acaulospora sp-1 dan
Hubungan antara nilai kepadatan spora CMA (Jumlah Spora / 10 gr tanah)
dengan persen kolonisasi akar dapat dilihat pada Gambar 3.
0
Gambar 3. Grafik Kepadatan Spora CMA (Jumlah Spora / 10 gr Tanah) dan % kolonisasi akar
Gambar 3. menunjukkan bahwa tidak selalu jumlah spora yang tinggi
memiliki persentase kolonisasi yang tinggi pula. Meskipun terdapat beberapa data
yang menunjukkan hubungan positif antara jumlah spora dan persentase
kolonisasi tetapi tetap dijumpai kolonisasi yang rendah pada lokasi yang memiliki
jumlah spora yang banyak dan sebaliknya. Jumlah spora tertinggi terdapat pada
vegetasi alang-alang dan yang terendah terdapat vegetasi sayur-sayuran.
Dari hasil penelitian diketahui bahwa masing-masing vegetasi
menunjukkan simbiosis dengan CMA. Persen kolonisasi yang tertinggi terdapat
pada vegetasi umbi-umbian dan yang terendah terdapat pada vegetasi
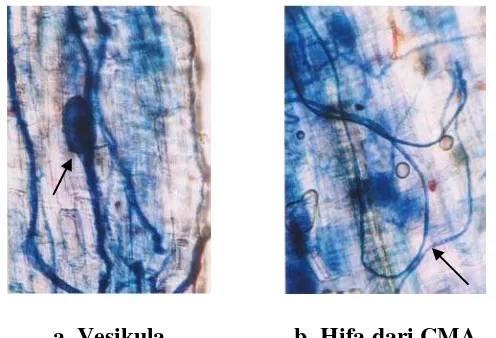
sayur-sayuran. Infeksi CMA pada akar tanaman dapat diidentifikasi melalui adanya hifa,
vesikula, dan arbuskula yang terdapat secara bersamaan atau salah satu dari
ketiganya. Infeksi CMA pada akar tanaman yang diobservasi dari lokasi penelitian
a. Vesikula b. Hifa dari CMA
Pembahasan
Kepadatan Spora Cendawan Mikoriza Arbuskula (CMA)
Tanah berkapur di desa Ria-ria, Kabupaten Tapanuli Utara yang dijadikan
sebagai areal penelitian memiliki spora CMA yang cukup tinggi. Dari hasil
penelitian diperoleh data bahwa vegetasi alang-alang memiliki jumlah rata-rata
spora 42, vegetasi sayur-sayuran 22 dan untuk vegetasi umbi-umbian memiliki
jumlah spora sebanyak 36. Dari data tersebut dapat dilihat bahwa jumlah spora
yang terbanyak terdapat pada vegetasi alang-alang dengan jumlah spora 42
sedangkan jumlah spora yang paling sedikit terdapat pada vegetasi sayur-sayuran
sebesar 22.
Perbedaan jumlah spora yang terdapat pada masing-masing vegetasi
tersebut mungkin disebabkan karena banyak faktor. Faktor abiotik misalnya
ketersediaan hara yang terdapat di dalam tanah dan penggunaan pupuk. Pada
vegetasi alang-alang tidak ada pemupukan, tidak seperti pada vegetasi
sayur-sayuran dan umbi-umbian dimana dilakukan pemupukan atau penambahan
konsentrasi hara ke dalam tanah. Kandungan hara khususnya P dan N dalam tanah
mempengaruhi perkembangan CMA. Oleh karena itu pada kedua vegetasi ini
jumlah dan peran mikoriza tidak begitu besar karena hara yang tersedia dalam
tanah sudah mencukupi. Lain halnya pada vegetasi alang-alang dimana
ketersediaan haranya bisa dibilang rendah sehingga peran mikoriza lebih besar di
dalam tanah sehingga jumlahnya lebih banyak. Hal ini dapat dilihat dari hasil
analisis tanah dimana pada vegetasi sayur-sayuran dan umbi-umbian memiliki C
organik yang lebih tinggi daripada vegetasi alang-alang meskipun nilai dari ketiga
pendapat Suhardi (1989) yang menyatakan bahwa pada kondisi tanah yang subur
dimana tingkat pengolahan tanah yang tinggi perkecambahan dari spora agak
terhambat sehingga tidak banyak dijumpai mikoriza baik spora maupun hifanya.
Faktor lain yang menyebabkan hal tersebut adalah terjadinya pengolahan
tanah seperti pembalikan tanah pada waktu sebelum penanaman. Pada waktu
pembalikan tanah mungkin mikoriza terangkat ke atas dan merusak jaringan hifa
dalam tanah sehingga banyak mikoriza yang mati yang menyebabkan jumlah
spora pada vegetasi sayur-sayuran dan umbi-umbian lebih sedikit dibanding
vegetasi alang-alang. Penggunaan pestisida juga dapat mempengaruhi jumlah
spora dan kolonisasi dari CMA. Pada tanaman sayur-sayuran misalnya tingkat
keseringan penggunaan pestisida yang tinggi menyebabkan terganggunya
kolonisasi dan produksi spora CMA sehingga jumlah spora dan kolonisasinya
lebih rendah daripada vegetasi alang-alang dan umbi-umbian..
Tanah berkapur termasuk tanah yang kurang subur atau memiliki harkat
yang rendah, hal ini dapat dilihat dari nilai C-organik dari ketiga jenis vegetasi
memiliki nilai antara 1,07 sampai dengan 1,10 %. Hal ini sesuai dengan pendapat
Rosmarkam dan Yuwono (2002) yang menyatakan bahwa suatu tanah itu
memiliki % C-organik antara 0,60-1,25 termasuk kedalam harkat rendah.
Jika dilihat dari hasil analisis tanah bahwa pH, C-organik, P tersedia, dan
Ca dari masing-masing vegetasi tersebut masih termasuk dalam satu harkat
(tingkatan). Oleh karena itu perbedaan jumlah spora yang ditemukan di lapangan
mungkin saja di pengaruhi oleh jenis inangnya. Dimana jumlah spora yang paling
tinggi di temukan pada vegetasi alang-alang, berarti vegetasi alang-alang lebih
Jenis tanaman dan kerapatan akar sangat berpengaruh terhadap penyebaran CMA.
Suhardi (1989) menyatakan bahwa walaupun sudah diketahui bahwa CMA dapat
berasosiasi dengan beraneka jenis tanaman inang namun diketahui bahwa mereka
memilih tanaman inang yang satu lebih disukai dibanding dengan tanaman inang
yang lain. Belum ada laporan tertentu bahwa spora tertentu hanya bisa berasosiasi
dengan tanaman inang tertentu.
Berdasarkan hasil yang diperoleh, diketahui bahwa secara umum tipe
spora yang ditemukan didominasi oleh jenis Glomus spp., meskipun ada juga
dijumpai jenis lain seperti Acaulospora spp. namun jumlahnya hanya 2 spora saja.
Hal ini menunjukkan bahwa tipe Glomus mempunyai daerah sebaran yang paling
luas dibanding dengan genus-genus yang lain dan Glomus merupakan spesies
yang paling toleran terhadap lingkungan maksudnya bahwa Glomus ini bisa
bertahan dan bahkan memperbanyak diri pada berbagai kondisi lingkungan mulai
dari yang baik sampai ekstrim. Misalnya pada pH tanah bahwa Glomus
berkecambah baik pada pH antara 6-8 (Abbot dan Robson, 1984). Oleh karena itu
Glomus disini dapat berkecambah dengan baik mulai dari pH asam sampai pH
basa. Hasil analisis tanah menunjukkan pH dari setiap vegetasi mulai 7,39 sampai
7,66. Nilai pH optimum untuk perkecambahan bergantung pada kemampuan
adaptasi dari CMA terhadap lingkungan, misalnya terhadap suhu optimum dan
juga tergantung pada jenis CMA. Pada umumnya Glomus tidak mempunyai inang
yang spesifik artinya cendawan ini mempunyai kisaran inang yang luas dan
merupakan salah satu karakteristik yang khas dari CMA. Oleh karena itu Glomus
Persentase Kolonisasi Akar (%)
Dari hasil pengamatan dan perhitungan persentase kolonisasi yang
dilakukan, dapat dikatakan bahwa tanah berkapur yang dijadikan sebagai areal
penelitian memiliki vegetasi yang secara umum membentuk asosiasi yang cukup
tinggi dengan CMA, dengan nilai kolonisasi yang sangat bervariasi dengan antara
58,654 % - 85,19 % termasuk dalam kelas kategori sangat tinggi (Setiadi et al.,
1992).
Dari tiga jenis vegetasi yang diobservasi, diketahui bahwa ketiga jenis
vegetasi tersebut (alang-alang, sayur-sayuran, dan umbi-umbian) membentuk
simbiosis dengan CMA. Kisaran infeksi tertinggi adalah pada vegetasi
umbi-umbian yaitu 85,19 % termasuk dalam kelas sangat tinggi, dan terendah pada
vegetasi sayur-sayuran dengan nilai 58,654 % termasuk dalam kelas tinggi. Hasil
ini menunjukkan bahwa jenis umbi-umbian merupakan tanaman dengan
persentase infeksi yang tergolong tinggi dibanding yang lain. Hal ini diduga
berkaitan dengan tipe akar dari vegetasi yang menjadi inangnya. Tipe akar yang
tumbuh pendek pada tanaman umbi-umbian (dalam hal ini ubi jalar) lebih mudah
untuk diinfeksi oleh CMA jika dibandingkan dengan tipe akar dari sayur-sayuran
dan alang-alang. Hal ini sesuai dengan pendapat Conway dan Joseph (1984) yang
menyatakan bahwa dari beberapa pengamatan jenis tanaman yang sistem
perakarannya tanpa akar-akar halus dan kurang rambut akarnya ternyata lebih
sering terkena infeksi dan lebih tergantung terhadap adanya mikoriza untuk
pertumbuhannya yang normal.
Dari hasil perhitungan derajat infeksi dan persentase kolonisasi akar dari
tersebut memiliki persentase kolonisasi akar yang bervariasi, dimana dalam
penelitian ini kemampuan tanaman berasosiasi dengan CMA dapat dikatakan
tinggi. Penentuan status mikoriza ini didasarkan pada tabel nilai persen kolonisasi
menurut Setiadi et al, (1992). Infeksi akar ini diukur untuk mengetahui efektifitas
simbiosis akar dengan CMA, karena semakin besar tingkat infeksi maka
diperkirakan akan semakin besar pula tingkat penyerapan unsur hara.
Peningkatan jumlah spora tidak selalu berhubungan terhadap peningkatan
kolonisasi akar. Hal ini dapat kita lihat secara jelas pada jumlah spora yang tinggi
pada vegetasi alang-alang dengan rata-rata jumlah spora 42 / 10 gr tanah tetapi
hanya memiliki persentase kolonisasi akar sebesar 71,376 %. Dan sebaliknya pada
vegetasi umbi-umbian memiliki rata-rata spora sebanyak 36 / 10 gr tanah dan
memiliki persentase kolonisasi akar sebesar 85,19 %. Hal ini sejalan dengan
penelitian Hayman (1970) dalam Suhardi (1989) yang menyatakan bahwa tidak
ada hubungan erat antara kolonisasi akar dengan produksi spora, sehingga hal
tersebut tidak bisa dijadikan sebagai suatu patokan bahwa jumlah spora
berpengaruh terhadap persen kolonisasi. Jumlah spora yang tinggi tidak selalu
menunjukkan adanya kolonisasi akar yang tinggi pula, karena jumlah spora yang
ada atau ditemukan tidak bisa menunjukkan secara langsung jumlah kolonisasi
yang terbentuk pada tanaman.
Hal ini disebabkan karena kemampuan dari CMA itu untuk menginfeksi
berbeda-beda untuk setiap jenis tanaman inang. Misalnya vegetasi umbi-umbian
lebih mudah diinfeksi oleh CMA dibanding vegetasi alang-alang. Selain itu bisa
disebabkan oleh keadaan dari spora itu sendiri apakah dorman atau tidak. Keadaan
ketersediaan air cukup maka spora cenderung akan berkecambah dan jika kering
maka spora akan dorman dan spora muda akan mempertahankan diri. Spora yang
terdapat pada vegetasi alang-alang mungkin saja dalam keadaan dorman karena
pada vegetasi alang-alangnya kondisi tanahnya sangat kering dan gersang
sehingga persen kolonisasinya lebih rendah dibanding vegetasi umbi-umbian.
Sedangkan pada vegetasi sayur-sayuran dan umbi-umbian ketersediaan airnya
cukup ini dapat dilihat dari kondisi tanah yang gembur dan lebih lembab daripada
tanah pada vegetasi alang-alang.
CMA ini akan berkolonisasi dengan baik jika tanah yang menjadi tempat
tumbuh inangnya memiliki kandungan P yang rendah. Jika kandungan P tanah
rendah maka persen kolonisasi tinggi dan sebaliknya. Hal ini disebabkan karena
pada kondisi P tanah rendah maka P tanaman juga rendah sehingga kandungan
fosfolipid di akar akan menurun. Hal ini menyebabkan permeabilitas akar akan
tinggi sehingga mudah diinfeksi oleh mikoriza. CMA ini memiliki suatu enzim
yang disebut enzim phosphatase yang mampu melepaskan P dari ikatan-ikatan
spesifik sehingga tersedia bagi tanaman. Oleh karena itu jika unsur P dalam tanah
tinggi dapat menghambat kolonisasi dan produksi spora CMA. Hal ini sesuai
dengan pendapat Husien dkk (2000) yang menyatakan bahwa kesuburan tanah
(unsur P dan N tersedia), kadar air, drainase tanah dan pH tanah berpengaruh
terhadap perkembangan CMA. Di samping itu CMA dapat berkembang dengan
baik pada tanah yang mempunyai kandungan P lebih rendah dan aerasi tanah yang
KESIMPULAN DAN SARAN
Kesimpulan
1. Persentase kolonisasi akar pada vegetasi alang-alang dan sayur-sayuran
termasuk tinggi dan sangat tinggi pada vegetasi umbi-umbian.
2. Jumlah spora pada vegetasi alang-alang lebih tinggi dibandingkan dengan
vegetasi sayur-sayuran dan umbi-umbian.
3. Glomus spp. merupakan tipe spora yang mendominasi pada tanah berkapur
di Desa Ria-Ria Kecamatan Sipoholon, Kabupaten Tapanuli Utara.
Saran
Lahan yang dijadikan sebagai lokasi penelitian ini cocok dikembangkan
sebagai lahan pertanian maupun kehutanan karena dari hasil penelitian diketahui
DAFTAR PUSTAKA
Abbott, L.K., dan Robson. 1994. An Ecological New of The Formation of VA Mycorrhizas. Plant and Soil.
Baon, J. B. 1998. Peranan Mikoriza VA pada Kopi dan Kakao. Makalah disampaikan dalam Workshop Aplikasi Cendawan Mikoriza Arbuskula Pada Tanaman Pertanian, Perkebunan dan Kehutanan. 5-10 Oktober 1998. Bogor.
Brundrett, M., N. Bougher, B. Dell, T. Grave and N. Malajezuk. 1996. Working with Mycorrhiza in Forestry and Agriculture. Australian Centre for International Agricultural Research (ACIAR). Carbera.
Daniels, B.A., dan Trappe, J.M. 1980. Factors Affecting Spore Germination of Vesicular-Arbuskular Mycorrhizal Fungus, Glomus Epigeus. Mycology.
Delvian, 2003. Keanekaragaman dan Potensi Pemanfaatan Cendawan Mikoriza Arbuskular di Hutan Pantai. Disertasi Doktor IPB. Bogor
Ekamawanti, A. H. 1998. Mekanisme Angkutan Nutrien dalam Simbiosis Mikoriza Arbuskula. Workshop Aplikasi Cendawan Mikoriza Arbuskula pada Tanaman Pertanian, Perkebunan dan Kehutanan. 21-23 April 2000. Bogor
Fakuara, Y.M. 1988. Mikoriza, Teori dan Kegunaan dalam Praktek. Institut Pertanian Bogor. Bogor.
Fitter, A.H., dan R.K.M. Hay. 1991. Fisiologi Lingkungan Tanaman. Gadjah Mada University Press. Yogyakarta.
Giovanetti, M. dan Mosse, B. 1980. An Evaluation of Technique for Measuring Vesicular-Arbuskular Mycorrhizal Infection In Roots.
Hanafiah, K., I. Anas, A. Napoleon, N. Ghoffar. 2003. Biologi Tanah. Ekologi dan Makrobiologi Tanah. PT. RajaGrafindo Persada. Jakarta.
Haryantini, B.A. dan Santoso.M. 2003. Pertumbuhan dan Hasil Cabai Merah (Capcicum annum) Pada Andisol Yang Diberi Mikoriza, Pupuk Fosfor dan ZPT
Ramah Lingkungan dalam Meningkatkan Produktivitas Lahan di Bidang Kehutanan, Perkebunan dan Pertanian di Era Milenium Baru. 21-23 April 2000. Bogor.
INVAM, 2004. From Versus Function, With A Focus On Levels Of Taxonomic Resolution (http// INVAM. Caf. MVU. Edu/ Fungi/ Taxonomy/ Concepts/ Scutsw. JPG). [07 Agustus 2006]
Islami, T. dan H.U. Wani. 1995. Hubungan Tanah, Air dan Tanaman. IKIP Semarang Press. Semarang.
Kehri, H.B and Chandra, S. 1990. Relative Efficacy of VAM Isolates for Green Gram Under Water Stress Condition. Proceedings of the National Conference on Mycorrhiza. 14-16 February 1990. Haryana Agricultural.
Lakitan, B. 2000. Dasar-Dasar Fisiologi Tumbuhan. PT. RajaGrafindo Persada. Jakarta.
Muin, A. 2004. Penggunaan Mikoriza Untuk Menunjang Pembangunan Hutan pada Lahan Kritis atau Marginal. Studi Ilmu Pengetahuan Kehutanan. Bogor [02 Agustus 2006].
Rao, N.S dan Subha, 1994. Mikroorganisme tanah dan pertumbuhan tanaman. Edisi Kedua. Penerbit Universitas Indonesia.
Rosmarkam, A. dan Yuwono, N. W. 2002. Ilmu Kesuburan Tanah. Penerbit Kanisius. Yogyakarta.
Setiadi, Y. 1998. Fungsi Mikoriza Arbuskular dan Prospeknya Sebagai Pupuk Biologis. Makalah disampaikan dalam Woarkshop Aplikasi Cendawan Mikoriza Arbuskula Pada Tanaman Pertanian, Perkebunan dan Kehutanan. 5-10 Oktober 1998. Bogor.
Setiadi, Y . 2001. Peran Mikoriza Arbuskula dalam Rehabilitasi Lahan Kritis Di Indonesia. Makalah Disampaikan dalam Seminar penggunaan CMA dalam Sistem Pertanian Organik dan Rehabilitasi Lahan Kritis. 23 April 2001. Bandung.
Smith, S.E. dan Read, D.J. 1997. Mycorrhizal Symbiosis. Second Edition. Academic Press. Harcourt Brace & Company Publisher. London.
RIWAYAT HIDUP
Penulis dilahirkan di Tarutung pada tanggal 16 Maret 1984 dari Ayah
Halomoan Lumban gaol dan Ibu Saida Silaban. Penulis merupakan anak keempat
dari enam bersaudara.
Penulis menyelesaikan pendidikan Sekolah Dasar di SDN No. 173146
Tarutung, pendidikan SMP di SMP swasta Santa Maria Tarutung. Selanjutnya
pendidikan tingkat atas di SMU Negeri 1 Tarutung pada tahun 2002. Pada tahun
yang sama penulis lulus pada Program Studi Budidaya Hutan Departemen
Kehutanan Fakultas Pertanian Universitas Sumatera Utara melalui jalur SPMB.
Dalam menyelesaikan pendidikan sarjana, penulis pernah mengikuti
Praktik Umum Kehutanan di hutan pegunungan Lau kawar Kabupaten Karo dan
hutan mangrove Bandar Khalifah Kabupaten Serdang Bedagai pada tahun 2004.
Penulis melakukan Praktik Kerja Lapang di PT. Toba Pulp Lestari Tbk. selama
dua bulan pada tahun 2006 dan mengerjakan penelitian yang berjudul
“Keberadaan dan Status Cendawan Mikoriza Arbuskula (CMA) Pada Beberapa
Vegetasi di Tanah Berkapur” studi kasus di Desa Ria-ria, Kecamatan Sipoholon,