KETAHANAN TERHADAP HAWAR DAUN BAKTERI DAN
KERAGAAN PRODUKTIVITAS PADI SAWAH YANG
DITANAM TUNGGAL DAN CAMPURAN
TRI HASTINI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
DITANAM TUNGGAL DAN CAMPURAN
TRI HASTINI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa Tesis Ketahanan terhadap Hawar Daun Bakteri dan Keragaan Produktivitas Padi Sawah yang Ditanam Tunggal dan Campuran adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Desember 2012
TRI HASTINI. Bacterial Leaf Blight Resistance and Productivity Performance of Rice on Single and Mixture Planting. Supervised by HAJRIAL ASWIDINNOOR, DESTA WIRNAS and ABDJAD ASIH NAWANGSIH.
The core of Indonesian agricultural development policy is to meet the demand of rice as staple food. The improved varieties which have been used still faced constraints such as abiotic and biotic stress. One of the important biotic stress is Bacterial Leaf Blight disease caused by Xanthomonas oryzae pv.oryzae. According to the previous research, mixture varieties planting could reduce disease intensity and increase the yield. The aim of this research was to find out the genotype that resistance to BLB, yield stability, genetic variability and other characters stability on single and mixture planting. The experiment used 17 genotypes as treatments which consisted of 7 cultivars and 10 cultivar mixtures with different resistance to BLB background. The trial was designed in Randomized Complete Block Design and replicated three times. The trial was conducted during two seasons in Majalengka and Pacitan. The characters observed were Bacterial Leaf Blight spreading, plant height, number of panicle per hill, number of well-filled grain per panicle, number of unfilled grain per panicle, number of total grain per panicle, 1000 grains weight, well-filled grain percentage, yield per ha and sink size. The disease spreading data was used to analyze the first appearance of disease, disease incidence, disease severity and Area Under Disease Progress Curve (AUDPC). The yield data was used to analyze yield stability and genetic parameters, while agronomic characters data was used to analyze the difference between two seasons and between single and mixture genotype using t test. According to the result of analysis, there was no difference between single and mixture genotype on the first appearance of disease, disease incidence, disease severity and AUDPC. The result of combine and stability analysis showed that genotype, season and their interactions influenced the yield; and the stable, unstable and specific single and mixture genotype could be determined. According to t test to other characters, there was no difference between single and mixture genotype. Based on the result of the analysis, the effect of mixture to reduce the Bacterial Leaf Blight disease and to increase the yield has not been known yet.
RINGKASAN
TRI HASTINI. Ketahanan terhadap Hawar Daun Bakteri dan Keragaan Produktivitas Padi Sawah yang Ditanam Tunggal dan Campuran. Dibimbing oleh
HAJRIAL ASWIDINNOOR, DESTA WIRNAS dan ABDJAD ASIH
NAWANGSIH.
Permintaan padi sebagai makanan pokok penduduk Indonesia sangat
tinggi. Pemenuhan kecukupan pangan pokok menjadi prioritas dalam kebijakan
pembangunan pertanian di Indonesia. Salah satu cara untuk memenuhi kecukupan
pangan tersebut dengan penanaman varietas unggul. Penanaman varietas unggul
seringkali mengalami kendala berupa cekaman abiotik dan cekaman biotik. Salah
satu cekaman biotik pada tanaman padi saat ini adalah penyakit Hawar Daun
Bakteri (HDB) yang disebabkan oleh bakteri Xanthomonas oryzae pv. oryzae (Xoo). HDB dapat mengakibatkan kerugian hingga 50% bahkan puso. Selama ini penanggulangan HDB terutama dilakukan dengan cara penanaman varietas yang
tahan, namun ketahanan tersebut mudah patah di lapangan karena bakteri Xoo sangat mudah bermutasi membentuk strain baru.
Penggunaan genotipe campuran dalam menurunkan laju perkembangan
penyakit telah dicoba pada beberapa tanaman pangan seperti gandum, barley dan
padi. Penanaman genotipe campuran tersebut mampu menurunkan keparahan
penyakit dan meningkatkan hasil. Penanaman genotipe campuran pada tanaman
padi diharapkan dapat menjadi alternatif dalam mengendalikan penyakit HDB.
Hasil penelitian pada empat lingkungan diperoleh informasi tidak terdapat
perbedaan kemunculan pertama penyakit, keparahan penyakit dan Area Under Disease Progress Curve (AUDPC) antara genotipe tunggal dan campuran. Perbedaan terjadi antar musim tanam, dimana pada musim hujan terlihat
keparahan penyakit lebih tinggi.
Analisis gabungan dan analisis stabilitas model Additive Main Effect and Multiplicative Interaction (AMMI) memberikan informasi bahwa semua sumber keragaman berpengaruh terhadap hasil kecuali ulangan/lokasi*musim. Hasil
analisis AMMI menunjukan terdapat genotipe tunggal dan campuran yang
tergolong stabil, tidak stabil dan spesifik. Produktivitas pada genotipe campuran
Analisis terhadap karakter pengamatan yang lain memberikan informasi
yang sama. Tidak terdapat perbedaan antara genotipe tunggal dan campuran pada
karakter tinggi tanaman, jumlah malai per rumpun, jumlah gabah isi per malai,
jumlah gabah hampa per malai, jumlah gabah total per malai, persentase gabah isi,
hasil per ha dan ukuran sink. Berdasar uji t, perbedaan hanya terlihat antara musim kemarau dan musim hujan.
Berdasar analisis terhadap penyakit, stabilitas hasil serta karakter
pengamatan yang lain dapat diketahui tidak ada pola yang spesifik ada genotipe
tunggal maupun campuran. Hal tersebut mengindikasikan bahwa pencampuran
tidak mempengaruhi intensitas penyakit, stabilitas hasil dan nilai pengamatan
karakter lainnya.
©Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
NIM : A253100121
Disetujui
Komisi Pembimbing
Dr. Ir. Hajrial Aswidinnoor, M.Sc. Ketua
Dr. Desta Wirnas, S.P., M.Si. Anggota
Dr. Ir. Abdjad Asih Nawangsih, M.Si. Anggota
Mengetahui
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dekan Sekolah Pascasarjana
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, M.Sc.Agr.
Puji dan syukur penulis panjatkan ke hadirat Alloh SWT atas segala
karuniaNya sehingga karya ilmiah ini berhasil diselesaikan. Tema penelitian yang
dipilih adalah ketahanan terhadap Hawar Daun Bakteri dan keragaan produktivitas
padi sawah yang ditanam secara tunggal dan campuran. Penelitian dilaksanakan
sejak bulan Mei 2011 dan berakhir bulan Maret 2012. Judul karya ilmiah ini
adalah Ketahanan terhadap Hawar Daun Bakteri dan Keragaan Produktivitas Padi
Sawah yang Ditanam Tunggal dan Campuran.
Terima kasih penulis ucapkan kepada Dr. Ir. Hajrial Aswidinnoor, M.Sc.
dan Dr. Desta Wirnas, S.P., M.Si., serta Dr. Ir. Abdjad Asih Nawangsih, M.Si.,
yang telah banyak memberikan saran. Terima kasih kepada Dr. Reflinur Basyirin
dan Tinche yang telah meluangkan waktu untuk mengoreksi penulisan abstract. Ungkapan terima kasih juga penulis sampaikan kepada Badan Penelitian dan
Pengembangan Pertanian yang telah memberikan dana dan kesempatan studi,
program I-HERE B.2c yang membantu pendanaan dalam penelitian, rekan-rekan
teknisi dan penyuluh pertanian di lapangan yang membantu jalannya penelitian
dan petani yang menyediakan lahan untuk percobaan serta rekan-rekan
seperjuangan PBT 2010. Tidak lupa terima kasih kepada suami, orang tua,
anak-anak, Ibu Uki yang telah mengasuh anak-anak serta seluruh keluarga atas
pengertian, dukungan moral dan doanya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Desember 2012
RIWAYAT HIDUP
Penulis dilahirkan di Pacitan pada tanggal 17 Desember 1971 dari ayah
Danoeri dan ibu Rusmiatun. Penulis merupakan anak ketiga dari tiga bersaudara.
Pada tahun 2004 penulis menikah dengan Mujiaman Manik dan dikaruniai dua
putra yaitu Dzaky Muhammad Dzakir Damanik dan Hilmi Muhammad Faruq
Damanik.
Tahun 1992 penulis terdaftar sebagai mahasiswa pada Program Studi
Pemuliaan Tanaman Fakultas Pertanian Universitas Padjadjaran Bandung dan
lulus pada tahun 1997. Tahun 2010 penulis mendapatkan kesempatan melanjutkan
pendidikan pascasarjana dari Badan Penelitian dan Pengembangan Pertanian
Kementerian Pertanian pada Program Studi Pemuliaan dan Bioteknologi Tanaman
Institut Pertanian Bogor. Penulis bekerja sebagai peneliti pada Balai Pengkajian
Teknologi Pertanian Jawa Barat sejak tahun 2002, dengan bidang kajian
Halaman
DAFTAR TABEL... vii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... ix
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 3
Hipotesis... 3
TINJAUAN PUSTAKA Tanaman Padi... 5
Penyakit HDB dan Pengaruhnya terhadap Hasil Padi Sawah... 6
Mekanisme Pertahanan Tanaman Padi terhadap HDB ... 8
Peran Pencampuran Varietas dalam Menekan Perkembangan Penyakit ... 9
Stabilitas Produktivitas dan Keragaman Varietas Campuran ... 11
PENGARUH PENCAMPURAN GENOTIPE PADI SAWAH TERHADAP PERKEMBANGAN HAWAR DAUN BAKTERI Abstrak ... 15
Abstract ... 16
Pendahuluan ... 17
Bahan dan Metode... 19
Hasil dan Pembahasan... 22
Kesimpulan ... 29
Daftar Pustaka ... 29
PENGARUH PENCAMPURAN TERHADAP BEBERAPA KARAKTER AGRONOMI PADI SAWAH Abstrak ... 33
Abstract ... 34
Pendahuluan ... 35
Bahan dan Metode... 36
Hasil dan Pembahasan... 38
Kesimpulan ... 44
ii STABILITAS DAN NILAI PARAMETER GENETIK PRODUKTIVITAS PADI SAWAH GENOTIPE TUNGGAL DAN CAMPURAN
Abstrak ... 47
Abstract ... 48
Pendahuluan ... 49
Bahan dan Metode... 51
Hasil dan Pembahasan... 55
Kesimpulan ... 60
Daftar Pustaka ... 61
PEMBAHASAN UMUM ... 63
KESIMPULAN DAN SARAN... 67
DAFTAR PUSTAKA ... 69
Halaman
1 Materi yang digunakan dalam percobaan ... 20
2 Skala keparahan penyakit... 22
3 Kemunculan pertama penyakit di Majalengka dan Pacitan pada MK II dan MH (hss)... 23
4 Total nilai AUDPC di Majalengka dan Pacitan pada MK II dan MH ... 27
5 Materi yang digunakan dalam percobaan ... 36
6 Ringkasan hasil anova gabungan pada genotipe tunggal ... 38
7 Ringkasan hasil anova gabungan pada genotipe campuran ... 39
8 Rata-rata pengamatan genotipe tunggal MK II dan MH di Majalengka dan hasil ujit... 40
9 Rata-rata pengamatan genotipe tunggal MK II dan MH di Pacitan dan hasil ujit... 40
10 Rata-rata pengamatan genotipe campuran MK II dan MH di Majalengka dan hasil ujit... 41
11 Rata-rata pengamatan genotipe campuran MK II dan MH di Pacitan dan hasil ujit... 41
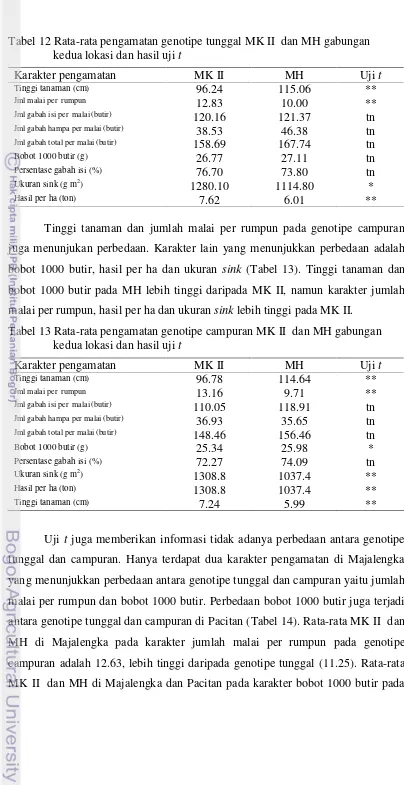
12 Rata-rata pengamatan genotipe tunggal MK II dan MH gabungan kedua lokasi dan hasil ujit... 42
13 Rata-rata pengamatan genotipe campuran MK II dan MH gabungan kedua lokasi dan hasil ujit... 42
14 Perbandingan genotipe tunggal dan campuran di Majalengka dan Pacitan berdasarkan ujit... 43
15 Materi genetik yang digunakan dalam percobaan... 52
16 Anova gabungan untuk musim dan lokasi ... 54
17 Rentang, rata-rata hasil, koefisien keragaman (KK) genotipe tunggal dan campuran padi sawah di dua musim dua lokasi ... 55
18 Produktivitas (t/ha) 17 genotipe di Majalengka dan Pacitan pada MK II dan MH ... 56
DAFTAR GAMBAR
Halaman
1 Mekanisme kekebalan jenis ETI tanaman padi terhadapXoo... 9
2 Kejadian penyakit di Majalengka... 24
3 Keparahan penyakit di Majalengka... 25
4 Kejadian penyakit di Pacitan... 26
5 Keparahan penyakit Pacitan ... 27
6 Boxplot produktivitas (t ha-1) berdasarkan lingkungan pengujian ... 56
Halaman
1 Deskripsi varietas Inpari 6 Jete ... 77
2 Deskripsi varietas Inpari 13 ... 78
3 Deskripsi varietas Ciherang ... 79
4 Deskripsi varietas Maros... 80
5 Deskripsi varietas Cigeulis... 81
6 Tata letak percobaan pada MK II di Majalengka... 82
7 Tata letak percobaan pada MK II di Pacitan ... 83
8 Tata letak percobaan MH di Majalengka ... 84
9 Tata letak percobaan MH di Pacitan ... 85
10 Contoh analisis persen kejadian penyakit (PKP) HDB... 86
11 Contoh analisis keparahan penyakit HDB ... 87
12 Contoh analisis AUDPC ... 88
13 Analisis ragam model AMMI dan nilai RMSPD... 89
14 Analisis ragam gabungan karakter produktivitas padi sawah pada MK II dan MH di Majalengka dan Pacitan ... 90
15 Nilai ragam, heritabilitas dan koefisien keragaman genetik karakter produktivitas padi sawah pada MK II dan MH di Majalengka dan Pacitan 91 16 Analisis ragam beberapa karakter genotipe tunggal ... 92
17 Analisis ragam beberapa karakter genotipe campuran... 94
18 Hasil ujit karakter penyakit HDB... 91
19 Hasil ujit karakter agronomi lainnya... 97
20 Hasil analisis tanah pada MK II di Majalengka ... 99
21 Hasil analisis tanah pada MK II di Pacitan ... 100
22 Hasil analisis tanah MH di Majalengka ... 101
23 Hasil analisis tanah MH di Pacitan ... 102
PENDAHULUAN
Latar Belakang
Beras merupakan makanan pokok penduduk Indonesia. Permintaan beras
setiap tahun semakin meningkat karena laju pertambahan penduduk yang
diperkirakan mencapai 1,43% pada kurun waktu 2010-2015 (Statistik Indonesia
2011). Saat ini konsumsi beras Indonesia tergolong tinggi di dunia yaitu sebesar
139 kg per kapita per tahun (BPS 2009). Kecukupan pangan terutama beras
dengan harga yang terjangkau telah menjadi tujuan yang utama kebijakan
pembangunan pertanian (Swastikaet al. 2007).
Peningkatan produksi padi dapat dilakukan melalui penanaman varietas
unggul berdaya hasil tinggi. Indonesia telah berhasil meningkatkan produksi dari
20.2 juta ton pada tahun 1971 menjadi 54 juta ton pada tahun 2006 melalui
peningkatan produktivitas padi, peningkatan luas panen dan interaksi keduanya
dengan kontribusi masing-masing sebesar 56.1%, 26.3% dan 17.5% (Sembiring
2007). Upaya lain untuk meningkatkan produksi adalah peningkatan sumberdaya
lahan, irigasi, intensitas tanam dan teknologi usaha tani (Swastika et al. 2007). Penanaman suatu varietas unggul berdaya hasil tinggi secara terus menerus akan
mengalami penurunan produktivitas apabila tidak disertai karakter ketahanan
terhadap cekaman biotik atau abiotik. Mekanisme penurunan produktivitas padi
dan ketahanan terhadap serangan penyakit utama disebabkan oleh terjadinya
degradasi genetik apabila suatu varietas ditanam secara terus-menerus (Suprapto
& Widyantoro 2005). Salah satu penyakit yang sangat penting dalam penurunan
produktivitas padi adalah Hawar Daun Bakteri (HDB).
Menurut IRRI (2009) penyakit HDB dapat menyebabkan kehilangan hasil
panen sebesar 6– 60%. Secara umum di dunia, infeksi HDB dapat mengakibatkan penurunan hasil hingga 50% (Ou 1985). Bahkan pada serangan HDB yang parah
dapat berakibat puso (Kadir et al. 2009). Penyakit HDB umum terjadi di negara tropik dan subtropik di lahan beririgasi maupun tadah hujan. HDB menyerang
tanaman pada fase vegetatif maupun generatif. Tanaman yang terinfeksi akan
mengalami kerusakan daun sehingga menyebabkan fotosintesis terganggu
HDB berkembang dengan baik pada suhu 25oC – 35oC, kondisi angin kencang, curah hujan dan kelembaban tinggi (Ou 1985). Selain itu, pemberian
pupuk nitrogen yang tinggi juga memicu berkembangnya HDB. Suprihanto et al. (2008) melaporkan bahwa dosis pupuk Nitrogen memberikan pengaruh nyata
terhadap keparahan beberapa penyakit padi. Bahkan pada musim kemarau (MK),
dosis pupuk Nitrogen berpengaruh nyata terhadap keparahan penyakit busuk
batang, hawar pelepah, hawar daun jingga, HDB danfalse smut. Pemberian pupuk Nitrogen dengan dosis yang semakin tinggi menunjukkan keparahan penyakit
busuk batang, hawar pelepah, hawar daun jingga dan HDB semakin tinggi.
Pemupukan dengan dosis Nitrogen tinggi (135 kg/ha) menyebabkan keparahan
paling tinggi dibandingkan dosis Nitrogen yang lebih rendah. Menurut Sudiret al. (2001) ketahanan genetik suatu varietas akan memunculkan reaksi ketahanan
terhadap serangan penyakit sehingga setiap varietas akan menunjukkan reaksi
yang berbeda terhadap serangan penyakit.
Keragaman genetik merupakan salah satu cara yang dapat mencegah
kerapuhan genetik tanaman yang ditanam secara monokultur. Di Propinsi Yunan
China, penanaman tanaman padi secara campuran menunjukkan hasil 89% lebih
tinggi dan penurunan serangan penyakit blas sampai 94% dibandingkan padi yang
ditanam secara monokultur (Zhu et al. 2001). Penggunaan fungisida berkurang dan penanaman varietas campuran merupakan pendekatan yang efektif untuk
menahan serangan penyakit terutama untuk wilayah penanaman yang luas
sehingga dapat menjaga kestabilan hasil panen (Zhuet al. 2000).
Varietas campuran didefinisikan sebagai pencampuran beberapa varietas
yang dibuat menjadi satu genotipe tanpa ada usaha pemuliaan untuk menciptakan
keseragaman fenotipiknya (Samonte 2008). Percampuran dua atau lebih varietas
pada tanaman menyerbuk sendiri merupakan usaha untuk memperbaiki populasi
yang bertujuan mendapatkan varietas unggul yang heterogen homozigot. Varietas
yang dicampur dipilih yang mempunyai potensi hasil tinggi, stabil dan
mempunyai ketahanan terhadap cekaman biotik atau abiotik. Percampuran untuk
memperoleh varietas yang lebih tahan harus tetap memperhatikan keseragaman
dari segi mutu, karakter agronomi, umur panen, bentuk, ukuran dan sebagainya
3
tunggal yang dipilih sebagai komponen percampuran harus mempunyai kontribusi
terhadap perbaikan sifat genetik yang dituju tanpa mengurangi keseragaman
fenotipik (Samonte 2008). Lebih jauh pemilihan komponen dalam percampuran
varietas umumnya berdasarkan ketahanan terhadap patotipe yang berbeda-beda
pada jenis patogen yang sama (Smithson & Lenne 1996).
Penanaman genotipe campuran tanaman padi dengan berbagai latar
belakang tingkat ketahanan HDB diharapkan dapat menurunkan laju mutasi
Xanthomonas oryzae pv oryzae (Xoo) sehingga kecepatan untuk memecahkan resistensi tanaman terhambat namun potensi hasil tetap tinggi dan stabil selama
beberapa musim tanam. Oleh karena itu perlu dilakukan penelitian mengenai
penanaman secara tunggal dan campuran pada tanaman padi untuk mengetahui
ketahanan genotipe terhadap penyakit HDB dan stabilitas hasilnya.
Tujuan Penelitian
1. Mendapatkan informasi tentang ketahanan padi sawah terhadap HDB pada
genotipe tunggal dan campuran.
2. Mendapatkan informasi tentang hasil dan keragaan karakter agronomi lainnya
antar musim pada genotipe tunggal dan campuran.
3. Mendapatkan informasi tentang stabilitas hasil dan keragaman genetik pada
genotipe tunggal dan campuran.
Hipotesis
1. Genotipe campuran lebih tahan terhadap HDB daripada genotipe tunggal.
2. Hasil dan keragaan karakter agronomi lainnya asal benih dari genotipe
campuran lebih baik daripada asal benih genotipe tunggal
3. Hasil dan stabilitas hasil genotipe tunggal dan campuran tidak berbeda dan
Padi merupakan satu-satunya jenis serealia yang dapat beradaptasi pada
tempat tumbuh yang kering maupun tergenang. Tanaman padi tersebar luas di
lima benua sehingga menjadi makanan pokok sebagian besar penduduk dunia
(IRRI 2009).
Menurut sejarahnya padi berasal dari China, namun ada juga yang
mengatakan dari India. Sejarawan yang menyatakan padi berasal dari China
berdasarkan sastra China yang dibuat pada 5000 tahun sebelum Masehi bahwa
padi sudah dibudidayakan pada masa kekaisaran Shen Mung. Padi mulai
menyebar ke seluruh dunia karena dibawa oleh bangsa India. Bangsa India
menyebarkan padi ke bagian selatan Spanyol melalui negara-negara Arab dan
kemudian berkembang luas di bagian Selatan Perancis, Italia dan masuk
negara-negara Balkan. Selain itu, dari India padi juga menyebar ke wilayah Asia bagian
timur seperti Jepang, Filipina dan kepulauan di lautan Pasifik. Padi juga tersebar
ke Malaysia dan akhirnya masuk ke Indonesia tahun 1500 sebelum Masehi
(Silitonga 2004).
Padi dengan nama ilmiah Oryza sativa L. merupakan anggota dari kingdom Plantae, Divisi Spermatophyta, Sub divisi Angiospermae, Kelas
Monocotyledoneae, Ordo Poales dan Famili Poaceae. Padi termasuk tumbuhan
setahun dalam bentuk rumpun, tinggi 80 – 130 cm, batang berbuku-buku yang pada setiap bukunya terdapat satu daun yang secara keseluruhan tersusun dalam
dua baris. Pelepah daun yang sebelah bawah menutupi pelepah daun yang di
atasnya sehingga terbentuklah batang semu yang menyelimuti buku-bukunya.
Bunga majemuk di ujung batang berbentuk malai dengan spikelet biseksual
berjumlah 50 – 500. Buahnya kariopsis dalam bentuk butiran. Berdasarkan habitat, tanaman padi dibedakan menjadi padi gogo, padi sawah dan padi rawa.
Sedangkan berdasarkan geografis, tanaman padi dibedakan ke dalam tiga sub
spesies yaitu indica,japonicadan javanica. Padi indica terutama di wilayah Asia Tenggara, japonica di Jepang, Korea Selatan, China Utara, New South Wales, Eropa bagian Selatan, California dan negara-negara Amerika Selatan, sedangkan
6
Penyakit HDB dan Pengaruhnya Terhadap Hasil Padi Sawah
Penyakit HDB diketahui pertama kali di Asia Timur dan sejak
diperkenalkan varietas berdaya hasil tinggi di Asia Selatan dan Asia Tenggara
pada 1960, HDB mulai menyebar luas. Sejak saat itu, HDB menjadi penyebab
utama menurunnya produksi padi pada lahan sawah irigasi, lahan sawah tadah
hujan dan lahan rawa (IRRI 1989). Selain itu, HDB juga ditemukan di Australia
menyerang padi liar Oryza rufipogon dan O. australiensis. Saat ini HDB menyebar luas dari benua Asia sampai Afrika dan Amerika. Distribusi
penyebarannya mulai dari 20o LS di Quensland Australia sampai 58o LU di
Heilang Jiang China dan serangan lebih tinggi pada musim hujan pada dataran
rendah daripada musim kemarau pada dataran tinggi (Mew 1989).
Menurut Mew (1989) HDB adalah penyakit vaskular, infeksinya bersifat
sistemik dan patogen masuk ke dalam tanaman inang melalui luka atau lubang
alami seperti stoma. Terkadang patogen masuk ke dalam xylem dan menyebar ke
seluruh tubuh tanaman. Patogen ini menyebabkan dua gejala tergantung fase
tanaman diserang. Apabila menyerang pada saat vegetatif pada fase persemaian
sampai menjelang fase terbentuknya anakan, gejalanya disebut kresek dan bersifat
lebih merusak. Seluruh daun tanaman menjadi pucat, kuning dan menggulung.
Penyakit kresek pertama kali ditemukan di Indonesia dan sangat umum
menyerang di daerah tropik. Apabila serangan terjadi pada saat fase anakan
maksimum sampai biji pada malai menguning disebut hawar daun. Gejala
serangannya terlihat mulai dari pinggir daun dan meluas sampai pelepah.
Selanjutnya jerami terlihat memutih, keabu-abuan atau kekuningan dalam 1-2
minggu. Ooze dari bakteri dapat diamati pada keadaan panas dan lembab.
Genus Xanthomonas pada tanaman dapat menyebabkan nekrosis, gummosis dan penyakit vaskular atau parenkhim pada daun, batang dan buah
pada 124 tumbuhan monokotil dan 268 tumbuhan dikotil. Genus Xanthomonas berwarna kuning, bentuk koloni tipis, merupakan bakteri gram negatif yang
patogenik, bergerak dengan satu flagel yang tedapat pada satu kutubnya atau tidak
Pengklasifikasian strain atau pathovar Xanthomonas berdasarkan pathogenesitasnya terhadap tanaman inang. Pengujian tingkat pathogenesitas
bakteri dilakukan menggunakan uji serologi. Dengan pendekatan serologi dapat
diketahui arsitektur molekuler sel bakteri serta hubungan antar bakteri. Sel bakteri
terdiri dari lapisan tipis, kapsul, flagela, pili, dinding sel dan membran sitoplasma
dimana terdapat antibodi yang dapat digunakan untuk mengelompokkan
jenis-jenis bakteri (Quimio 1989).
Bakteri menyerang tanaman inang diawali dengan proses pergerakan
bakteri, kontak antara bakteri dengan tanaman inang, penetrasi bakteri ke dalam
tanaman inang dan proliferasi bakteri di dalam tanaman inang. Pergerakan bakteri
secara kemotaksis dipicu oleh zat yang dikeluarkan oleh tanaman. Bakteri
kemudian mulai masuk ke tanaman inang dengan proses penetrasi melalui
stomata, hidatoda atau nektar serta luka pada tanaman. Masuknya organisme
patogen ke dalam inang dan kemudian terjadi hubungan parasitik baik permanen
atau tidak di antara keduanya disebut infeksi. Faktor lingkungan yang sangat
berperan dalam proses infeksi adalah kelembaban relatif, unsur hara, suhu dan
cahaya (Huang & Cleene 1989; Yudiarti 2007).
Pengaruh penyakit HDB terhadap pertumbuhan dan hasil padi sawah
berbeda-beda tergantung dari varietas dan stadia tumbuh tanaman. Penelitian
Sudir (2008) menunjukkan bahwa varietas yang tidak mempunyai gen ketahanan
terhadap HDB lebih mudah terserang dan populasi bakteri Xanthomonas oryzae
pv.oryzae (Xoo) lebih besar. Populasi bakteri Xoo pada varietas yang mempunyai gen ketahanan terhadap HDB lebih kecil bahkan tidak ada. Stadia tumbuh
tanaman juga berpengaruh nyata terhadap populasi bakteri Xoo. Populasi Xoo tertinggi terlihat pada stadia pengisian bulir dan terendah pada stadia anakan awal.
Populasi Xoo yang tinggi pada stadia pengisian bulir merupakan akumulasi dari stadia-stadia sebelumnya. Pada stadia pemasakan populasi Xoo mulai menurun karena kondisi tanaman yang sudah tua tidak mampu lagi menyediakan nutrisi
bagi perkembangan bakteri.
Menurut Ou (1985) penyakit HDB dapat menyebabkan kehilangan hasil
hingga 60%. Penyakit HDB juga dapat menurunkan mutu beras karena pengisian
8
mutu beras tersebut disebabkan karbohidrat/energi yang digunakan untuk tumbuh
dan berkembangnya tanaman yang dihasilkan oleh daun tidak maksimal. Daun
yang mengandung klorofil berfungsi untuk menangkap sinar matahari (Makarim
& Suhartatik 2009), maka apabila terjadi kerusakan pada daun karena penyakit
HDB, maka kandungan klorofil akan menurun sehingga fotosintat yang dihasilkan
juga menurun.
Mekanisme Pertahanan Tanaman Padi Terhadap HDB
Terdapat tiga jenis kekebalan tanaman terhadap patogen. Kekebalan
tersebut adalah PAMP-triggered imunity (PTI), effector-triggered imunity (ETI) dan systemic aqcuired resistance (SAR). PTI merupakan kekebalan yang dipicu oleh asosiasi molekuler patogen/mikrobia, sedangkan ETI adalah jenis kekebalan
yang dipicu oleh molekul efektor. Kekebalan yang mampu bertahan lama terhadap
berbagai jenis patogen adalah SAR, dimana dalam mekanismenya memerlukan
Asam Salisilat (SA) (Chen & Ronald 2011). Asam Jasmonik (JA), Etylene (ET)
dan SA merupakan fitohormon penting dalam mekanisme pertahanan tanaman
terhadap serangan patogen (Fujita et al 2006). Pada padi JA dan SA ditemukan pada bagian akar dan tajuk dengan konsentrasi yang bervariasi antar genotipe
yang berbeda (Youet al 2011).
Tanaman padi memiliki respon tingkat ketahanan terhadap bakteri Xoo yang berbeda-beda. Umumnya tingkat ketahanan tanaman inang dikelompokkan
menjadi dua yaitu ketahanan yang bersifat kualitatif dan kuantitatif. Ketahanan
kualitatif atau ketahanan vertikal dikendalikan oleh gen mayor yang monogenik
sedangkan ketahanan kuantitatif atau ketahanan horizontal dikendalikan oleh gen
minor yang multigenik. Hingga saat ini telah teridentifikasi 21 gen ketahanan
yang kualitatif (Gnanamanickamet al 1999).
Sumber : TRENDS in Plant Science
Gambar 1 Mekanisme kekebalan jenis ETI tanaman padi terhadapXoo
Apabila terjadi serangan patogen, XA21 kinase terlepas dari XB24, selanjutnya XA21 mengaktifkan mekanisme pertahanan. Respon selanjutnya melibatkan
mitogen-activated protein kinase 5 (MAPK5), MAPK12 dan XB3 yang sangat
diperlukan dalam aktivitasXA21(Chen & Ronald 2011).
Peran Pencampuran Genotipe Dalam Menekan Perkembangan Penyakit
Penelitian tentang penggunaan genotipe campuran telah dilakukan antara
lain untuk menekan penyakit blas pada padi di China (Zhu et al 2000; Zhu et al 2001), penyakit karat pada tanaman gandum (Biabani 2009), penyakit karat pada
10
gandum (Ninget al. 2012). Smithson & Lenne (1996) melaporkan bahwa varietas campuran telah banyak digunakan di pertanian subsisten maupun pertanian
modern. Pemilihan varietas untuk percampuran tidak hanya bertujuan untuk
menjaga stabilitas hasil dan menekan serangan penyakit namun juga untuk
kesesuaian jenis tanah dan musim tanam.
Penggunaan genotipe campuran merupakan pendekatan secara ekologi
untuk menekan serangan penyakit. Percobaan varietas campuran padi ketan yang
peka terhadap blas dan padiindica yang relatif tahan blas yang dilakukan dengan menanam satu baris padi ketan pada setiap empat baris padi indica mampu mengurangi kejadian dan keparahan penyakit blas. Kejadian penyakit blas pada
padi ketan yang ditanam secara campuran berkurang 94-99% apabila
dibandingkan dengan yang ditanam secara monokultur, dan pada padi hibrida
1-25%. Berkurangnya kejadian penyakit blas tersebut berpengaruh nyata terhadap
hasil. Hasil yang diperoleh dari percampuran varietas Xianyou 63 dan
Huangkenuo sebesar 10533,1 kg atau lebih tinggi 825,8 kg dibandingkan Xianyou
63 yang ditanam secara monokultur, hasil Xianyou 63 yang dicampur Zinuo
10461,7 kg atau lebih tinggi 833,9 kg, sedangkan hasil percampuran varietas
Xianyou 22 dan Huangkenuo serta Xianyou 22 dengan Zinuo masing-masing
sebesar 9996 kg dan 9993 kg atau lebih tinggi 916,8 kg dan 852,5 kg (Zhu et al 2001).
Hasil penelitian Biabani (2009) pada tanaman gandum memperlihatkan
hasil yang lebih tinggi pada pertanaman campuran dibanding pertanaman
monokultur. Penggunaan materi yang mempunyai toleransi lebih luas terhadap
keadaan lingkungan dan arsitektur kanopi diduga sebagai faktor utama yang
mendukung hasil yang tinggi. Arsitektur kanopi yang beragam memperbesar
intersepsi cahaya matahari ke dalam pertanaman sehingga memperbesar produksi
bahan kering.
Keragaman mendukung produktivitas dan stabilitasnya, dengan demikian
keragaman akan mendukung fungsi ekosistem sehingga kelestarian organisme
tetap terjaga (Phillips & Wolfe 2004). Pertanaman varietas campuran akan
mempertahankan kestabilan hasil meskipun pada kondisi lingkungan yang
keragaman biologi dan mempunyai potensi untuk mengendalikan hama dan
penyakit. Kelemahan penggunaan genotipe campuran adalah kesulitan
diaplikasikan dalam skala kecil dan kesulitan dalam melakukan mekanisasi
pertanian pada saat penanaman, pemupukan, penyiangan dan panen (Biabani
2009). Pada saat ini, pertanaman monokultur mendominasi pertanian modern.
Pertanaman monokultur adalah pertanaman yang menggunakan satu spesies dan
satu varietas sehingga keragaman genetik dalam populasi pertanaman tersebut
sangat terbatas. Alasan penggunaan pertanaman monokultur adalah kemudahan
dalam penanaman, panen dan prosesing serta hasil yang diperoleh akan seragam
kualitasnya. Pertanaman monokultur lebih banyak menyerap input produksi
berupa pemupukan dan pengendalian hama, penyakit dan gulma. Pertanaman
campuran dengan penggunaan varietas yang beragam dalam ekosistem
bermanfaat untuk mendaur ulang unsur hara, mengendalikan iklim mikro,
mengatur proses hidrologi secara lokal, menekan organisme yang tidak diinginkan
dan detoksifikasi bahan kimia berbahaya. Altieri (1999) dikutip Phillips dan Wolfe (2004) berpendapat bahwa keragaman genetik dengan penggunaan varietas
campuran atau varietas multilini dapat mengatasi masalah yang terjadi pada
pertanaman monokultur.
Stabilitas Produktivitas dan Keragaman Genotipe Campuran
Penelitian mengenai hasil dan stabilitas genotipe campuran telah dilakukan
sejak lama. Harlan dan Martini telah melakukan studi ini pada tahun 1938 pada
tanaman barley (Wortmann et al. 1996). Allard (1961) melaporkan bahwa hasil lima bean lebih stabil jika ditanam dalam populasi yang heterogen dibandingkan populasi yang homogen. Lebih lanjut Ayeh (1988) melaporkan bahwa interaksi
genotipe x lingkungan lebih kecil pada genotipe campuran bean dibandingkan genotipe tunggal. Meskipun tidak berbeda nyata dengan genotipe tunggal, namun
genotipe campuran lebih stabil dan rata hasilnya lebih tinggi daripada
rata-rata hasil komponen penyusunnya. Penelitian ini juga menunjukkan hubungan
yang linier antara diversitas dan stabilitas.
Berdasarkan penelitian Wortmann (1996), tidak terdapat indikasi bahwa
12
campuran menunjukkan hasil lebih tinggi dibandingkan rata-rata genotipe
penyusunnya. Slope garis regresi genotipe campuran >1 karena pada dua lokasi
pengujian genotipe tersebut menunjukkan hasil yang sangat baik. Analisis
stabilitas menggunakan metode AMMI dan Principal Coordinate Model pada faba bean menunjukkan bahwa genotipe yang stabil dalam resistensi terhadap penyakit tidak seiring dengan kestabilan hasil (Flores 1996).
Keragaman dapat diartikan sebagai perbedaan-perbedaan yang terdapat
dalam suatu populasi. Keragaman dalam populasi tanaman dapat dibedakan
menjadi keragaman fenotipik dan keragaman genotipik. Keragaman fenotipik
adalah keragaman yang dapat diukur atau dilihat langsung untuk karakter-karakter
tertentu, sedangkan keragaman genotipik adalah keragaman yang tidak dapat
dilihat atau diukur secara langsung namun dapat diduga melalui analisis data-data
statistik. Suatu populasi yang memiliki keragaman fenotipik yang luas belum
tentu mempunyai keragaman genotipik yang luas karena penampilan fenotipik
dipengaruhi oleh lingkungan (Hermiati 2001).
Hasil merupakan karakter yang bersifat kuantitatif yang dikendalikan oleh
banyak gen dan sangat dipengaruhi oleh lingkungan (Hermiati 2001). Cukup sulit
menentukan seberapa jauh kontribusi faktor genetik sebagai akibat aksi gen dan
seberapa jauh kontribusi lingkungan. Bagaimanapun juga, hasil yang tampak
(fenotipe) merupakan hasil dari genetik dan lingkungan, P = G + E (Roy 2000;
Syukuret al. 2009).
Keragaman yang diwariskan adalah keragaman genetik, sedangkan
keragaman lingkungan tidak diwariskan. Keragaman lingkungan harus dibuat
sekecil mungkin supaya pendugaan yang diperoleh terhadap aksi gen lebih
mendekati kebenaran. Beberapa sumber dari keragaman lingkungan adalah E1
(atau Ew), E2 dan Eb. E1 atau Ew merupakan keragaman lingkungan antara
individu dalam satu famili. Keragaman ini muncul karena keragaman sampel yang
diambil, kesalahan pengukuran dan keragaman pertumbuhan tanaman. E2 adalah
keragaman nilai rata-rata famili dan Eb adalah keragaman yang muncul karena
pengaruh petak percobaan (Roy 2000).
Keragaman dalam suatu populasi pemuliaan sangat penting artinya dalam
besar. Beberapa cara untuk menciptakan keragaman antara lain melalui
persilangan (hibridisasi), introduksi, mutasi dan transformasi genetik. Selain itu
penggunaan varietas campuran juga mampu untuk meningkatkan keragaman
genetik.
Koefisien keragaman dinyatakan sebagai standar deviasi dibagi dengan
rata-rata pengamatan (Roy 2000). Koefisien keragaman genetik (KKG)
menyatakan sempit atau luasnya keragaman genetik dari populasi yang sedang
dipelajari. Nilai KKG relatif rendah mengindikasikan sempitnya keragaman
genetik dari suatu populasi dan sebaliknya (Tampake & Luntungan 2002)
Heritabilitas adalah rasio proporsi ragam genotipik terhadap ragam
fenotipiknya. Heritabilitas menyatakan seberapa besar genotipe berperan terhadap
PENGARUH PENCAMPURAN GENOTIPE PADI SAWAH TERHADAP PERKEMBANGAN HAWAR DAUN BAKTERI
Abstrak
Percobaan dilakukan untuk membuktikan bahwa pencampuran genotipe dapat meningkatkan ketahanan terhadap Hawar Daun Bakteri. Percobaan menggunakan 17 genotipe padi sawah sebagai perlakuan, terdiri atas genotipe tunggal dan genotipe campuran dengan berbagai latar belakang ketahanan terhadap Hawar Daun Bakteri. Percobaan disusun Rancangan Acak Kelompok dengan tiga ulangan dan dilaksanakan di dua lokasi yaitu di Kabupaten Majalengka dan Pacitan selama dua musim tanam berturut-turut. Musim tanam pertama bertepatan dengan musim kemarau II 2011 (MK II) dan musim tanam kedua bertepatan dengan musim hujan 2011/2012 (MH). Luas petak percobaan 6.84 m2 atau petak bersih 5.78 m2. Pengamatan penyakit dimulai pada masa primordia dan berakhir saat menjelang panen dengan interval waktu ± 14 hari. Pengamatan luas area luka dilakukan terhadap 15 rumpun sampel dari setiap plot dan kemudian diskoring sesuai ketentuan SES IRRI. Data hasil pengamatan dianalisis menggunakan uji t untuk mengetahui perbedaan kejadian penyakit, keparahan penyakit dan Area Under Disease Progress Curve (AUDPC) pada genotipe tunggal dan campuran serta perbedaan antara MK II dan MH. Kemunculan pertama penyakit dianalisis secara deskriptif. Hasil uji t menunjukkan tidak terdapat perbedaan antara genotipe tunggal dan campuran dalam kejadian penyakit, keparahan penyakit dan AUDPC di Majalengka MK II, Majalengka MH dan Pacitan MK II. Perbedaan hanya terlihat di Pacitan MH dimana genotipe tunggal mempunyai nilai kejadian penyakit, keparahan penyakit dan AUDPC lebih tinggi daripada genotipe campuran. Hal tersebut menunjukkan adanya pengaruh pencampuran dalam menekan perkembangan penyakit Hawar Daun Bakteri di Pacitan pada MH.
Abstract
Seventeen rice genotypes used as treatments to prove that Bacterial Leaf Blight (BLB) resistance increased in mixture. The 17 genotypes have different levels of resistance againts BLB. The trial was designed in Randomized Complete Block Design with three replications and conducted at two locations those are Majalengka and Pacitan during second dry season and wet season. The plot area was 6.84 m2 or net plot was 5.78 m2. The data was gathered from primordia stage until harvest date to 15 sample hills per plot then scored based on International Rice Research Institute Standard Evaluation System for Rice (IRRI SES). Data were recorded for the first disease appearance, disease incidence, disease severity and Area Under Disease Progress Curve (AUDPC) analysis. Differences between cultivar vs cultivar mixtures and between dry season vs wet season were analyzed by t test. No difference in the first appearance of BLB between cultivar and cultivar mixtures during dry season and wet season in Majalengka and Pacitan. It was also no difference between dry season and wet season in cultivar and cultivar mixtures in Pacitan. Disease incidence showed that there was no difference between cultivar and cultivar mixtures during dry season and wet season in Majalengka and during wet season in Pacitan. Between dry season and wet season, there was disease incidence difference in cultivar and cultivar mixtures in Pacitan and cultivar in Majalengka. There were also no disease severity difference between cultivar and cultivar mixtures during dry season and wet season in Majalengka and during dry season in Pacitan. While between dry season and wet season, the difference occured in cultivar in Majalengka and Pacitan, and also in cultivar mixtures in Pacitan. The difference in AUDPC between cultivar and cultivar mixtures only occurred during wet season in Pacitan.
17
Pendahuluan
Salah satu penyakit utama pada komoditas padi sawah adalah Hawar Daun
Bakteri (HDB) yang disebabkan oleh bakteri Xanthomonas oryzae pv oryzae (Xoo). Xoo mempunyai banyak strain dan Indonesia merupakan negara yang mempunyai strain Xoo terbanyak dibandingkan negara-negara di Asia lainnya berdasarkan analisis kluster (Adhikari et al. 1995). Sejak tahun 1950 telah ditemukan 12 strainXoo di Indonesia (Wahyudiet al. 2011).
Penyakit HDB menyebar secara luas di dunia seperti Indonesia (Suryadiet al. 2006), India (Rajarajeswari & Muralidharan 2006) dan China (Qi 2009) serta wilayah lain dan mengakibatkan kehilangan hasil sampai 50% (Elinget al. 1997). Kejadian dan intensitas penyakit HDB sangat dipengaruhi oleh faktor lingkungan
antara lain sinar matahari, curah hujan, suhu, kelembaban udara, dosis Nitrogen
tinggi yang dapat meningkatkan laju asimilasi CO2, dan kondisi angin yang
kencang (Ou 1985; Horino et al. 1982). Apabila konsep segitiga penyakit terpenuhi, HDB berkembang sangat pesat bahkan bisa mengakibatkan puso (Kadir
et al. 2009). Elingset al. (1997) melaporkan bahwa kehilangan hasil merupakan korelasi antara tanaman inang x patogen x lingkungan.
Serangan bakteri Xoo dimulai dari ujung daun (bagian hidatoda) dan menjalar ke tepi daun hingga akhirnya merusak seluruh daun sehingga terlihat
seperti kering (Mew et al. 1993). Dalam jaringan daun, Xoo bermultiplikasi dan masuk ke dalam jaringan xylem sambil terus bermultiplikasi, meluas dan
memblokir transportasi air dan mengurangi laju transpirasi (Horino & Kaku
1989). Keadaan daun yang demikian mengakibatkan berkurangnya laju
fotosintesis dan diduga hal ini menjadi penyebab utama dalam mekanisme
kerusakan tanaman (Elings et al. 1997). Dengan demikian tanaman menjadi tidak mampu menghasilkan asimilat untuk proses pertumbuhan dan perkembangannya.
Hasil penelitian Philip & Devadath (1981) membuktikan adanya korelasi positif
antara luas area luka pada daun dengan laju fotosintesis.
Beberapa metode telah dilakukan untuk mengatasi serangan HDB, antara
lain peramalan penyakit (Yang 2010), pengendalian secara biologi misalnya
merupakan merupakan cara yang murah dan efisien (Josi & Nayak 2010) serta
ramah lingkungan, namun satu gen ketahanan hanya efektif untuk satu patogen
spesifik (Adhikari et al. 1995). Beberapa tahun terakhir upaya pemuliaan berupa pemanfaatan gen ketahanan terhadap Xoo yang berasal dari varietas padi liar juga telah dilakukan. Pemanfaatan gen ketahanan tersebut harus memperhatikan
interaksi inang-patogen (Qi 2009). Gen tersebut diintroduksikan ke dalam varietas
komersial untuk memberi ketahanan terhadap Xoo strain tertentu. Zhou et al. (2011) memanfaatkan gen ketahanan Xa23 dari Oryza rufipogon ke dalam padi hibrida Minghui63, YR293 dan Y1671 menggunakan pendekatanMarker Assisted Backcross (MAB) dan berhasil memperbaiki ketahanannya terhadap HDB. Namun genotipe dengan satu gen ketahanan akan mudah patah di lapangan karena
patogen akan terus berevolusi untuk mematahkan ketahanan tersebut. Misalnya
genotipe padi dengan gen ketahanan Xa4 saja yang sudah ditanam luas di India, Philipina dan Indonesia menjadi rentan terhadap HDB. Genotipe pada dengan gen
Xa21 yang sebelumnya dikenal mempunyai ketahanan yang luas juga telah patah ketahanannya di India, Korea, Nepal dan China (Qi 2009).
Strategi berbasis gen untuk mengurangi tekanan seleksi patogen dapat
dilakukan dengan cara rotasi varietas yang mempunyai gen ketahanan berbeda,
piramidanisasi gen, penanaman varietas multilini dan varietas campuran (Samonte
2008). Piramidanisasi gen adalah upaya untuk mengurangi intensitas penyakit
dengan cara mengumpulkan beberapa gen ketahanan ke dalam satu varietas.
Meskipun piramidanisasi gen terbukti paling efektif untuk mengatasi Xoo (Samonte 2008), namun cara ini relatif sulit dan tidak aplikatif di lapangan. Tidak
banyak dilaporkan keberhasilan piramidanisasi gen ketahanan terhadap Xoo, namun contoh genotipe yang telah berhasil diintroduksi dengan beberapa gen
ketahanan adalah galur PR106 yang diintroduksi tiga gen ketahanan terhadapXoo yaituxa5, xa12danXa21(Singhet al. 2001) dan galur padi yang diintroduksi gen Xa4, xa5, xa13 dan Xa21 (Huang et al. 1997). Introduksi tersebut menggunakan Marker Assisted Selection (MAS).
Piramidanisasi gen dalam populasi tanaman dengan menggunakan varietas
campuran merupakan alternatif pengendalian HDB yang murah, aplikatif dan
19
Samonte 2008). Banyak dilaporkan bahwa penggunaan varietas campuran mampu
menurunkan intensitas penyakit (Mundt & Leonard 1986; Mahmood et al. 1991; Finckh & Mundt 1992; Mundt 2002; Ning et al. 2012). Penggunaan varietas campuran mampu menurunkan intensitas penyakit diduga karena inokulum
menjadi lemah, percampuran merupakan hambatan fisik bagi patogen dan
percampuran akan menginduksi resistensi pada genotipe yang rentan (Ning et al. 2012). Garret & Mundt (1999) mempostulatkan bahwa semakin luas diversitas
ketahanan yang dimiliki oleh tanaman inang, akan menghambat kemampuan
adaptasi patogen terhadap gen ketahanan. Ohtsuki & Sasaki (2006) menyatakan
bahwa dengan mencampur genotipe yang rentan dan tahan akan mendorong strain
patogen berkompetisi untuk mencari inang yang kompatibel.
Percobaan ini dilakukan dengan tujuan untuk membuktikan bahwa
penanaman genotipe campuran dengan berbagai latar belakang ketahanan
terhadap HDB mampu menunda kemunculan pertama HDB, mengurangi
kejadian, intensitas dan AUDPC penyakit HDB.
Bahan Dan Metode
1. Materi percobaan
Perlakuan yang digunakan dalam percobaan adalah 17 genotipe padi
sawah berupa genotipe tunggal, campuran dua genotipe dan campuran tiga
genotipe dengan latar belakang ketahanan terhadap HDB beragam (Tabel 1).
Materi percobaan berasal dari Balai Besar Penelitian Tanaman Padi (BB Padi) dan
Institut Pertanian Bogor (IPB).
2. Pelaksanaan Percobaan
Rancangan percobaan yang digunakan adalah Rancangan Acak Kelompok
(RAK) dengan 17 perlakuan dan 3 ulangan. Percobaan dilakukan di lahan petani
di Kabupaten Majalengka Jawa Barat dan di Kabupaten Pacitan Jawa Timur
selama dua musim tanam berturut-turut. Generasi pertama ditanam bertepatan
dengan Musim Kemarau II 2011 (MK II) bulan Juni - September 2011 dan
generasi kedua ditanam bertepatan dengan Musim Hujan 2011/2012 (MH) bulan
Luas petak percobaan 6.84 m2 (petak bersih 5.78 m2). Benih disemai dan
pindah tanam bibit dilakukan pada 14 hari setelah semai (hss). Penanaman 2 bibit
per lubang tanam dengan jarak tanam 20 cm x 20 cm. Pupuk diberikan sesuai
kebutuhan tanaman berdasarkan hasil analisis tanah menggunakan Perangkat Uji
Tanah Sawah (PUTS). Pengujian sampel tanah dilakukan sebelum pengolahan
tanah. Perawatan lain yaitu penyiangan, pengaturan air dan pengendalian hama.
Pengamatan penyakit dimulai pada saat tanaman memasuki masa primordia
(Djatmiko & Fatichin 2009) sampai dengan menjelang panen dengan interval
14 hari sekali terhadap 15 rumpun sampel per plot. Luas area luka (%) setiap
rumpun sampel dicatat dan diskoring sesuai standar SES IRRI (1996) yaitu :
Skala 1 : 1– 5% Skala 3 : 6– 12% Skala 5 : 13– 25% Skala 7 : 26– 50% Skala 9 : 51– 100%
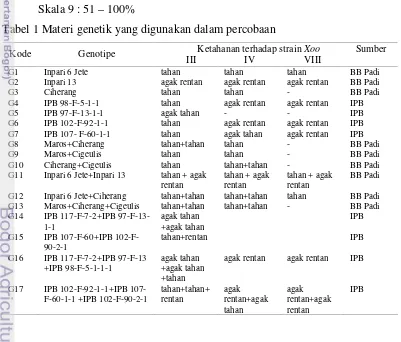
Tabel 1 Materi genetik yang digunakan dalam percobaan
Ketahanan terhadap strainXoo Sumber
Kode Genotipe
III IV VIII
G1 Inpari 6 Jete tahan tahan tahan BB Padi
G2 Inpari 13 agak rentan agak rentan agak rentan BB Padi
G3 Ciherang tahan tahan - BB Padi
G4 IPB 98-F-5-1-1 tahan agak rentan agak rentan IPB
G5 IPB 97-F-13-1-1 agak tahan - - IPB
G6 IPB 102-F-92-1-1 tahan agak rentan agak rentan IPB G7 IPB 107- F-60-1-1 tahan agak tahan agak rentan IPB
G8 Maros+Ciherang tahan+tahan tahan - BB Padi
G9 Maros+Cigeulis tahan tahan - BB Padi
G10 Ciherang+Cigeulis tahan tahan+tahan - BB Padi
G11 Inpari 6 Jete+Inpari 13 tahan + agak rentan
97-F-13-21
3. Analisis data
Data pengamatan penyakit di Majalengka dan Pacitan pada MK II dan MH
dianalisis menggunakan Microsoft Excel 2003 dan SAS 9.2. Analisis dilakukan
terhadap kemunculan pertama penyakit, kejadian penyakit, keparahan penyakit
danArea Under Disease Progress Curve (AUDPC). Untuk mengetahui perbedaan antara genotipe tunggal dan campuran dilakukan uji t pada taraf alpha 5%.
Kemunculan pertama penyakit ditentukan berdasarkan waktu mulai terlihatnya
gejala penyakit secara fisik. Kejadian penyakit merupakan jumlah penyakit dari
unit sampel yang dinyatakan sebagai persentase dari jumlah total unit sampel
yang diamati. Sedangkan keparahan penyakit adalah proporsi antara area yang
terinfeksi pada unit sampel dan total area unit sampel dan dinyatakan dalam
persen (Narayanasamy 2002)
Kejadian penyakit dihitung menggunakan formula (Amadioha 2000):
% n : jumlah rumpun yang terinfeksi N : jumlah rumpun yang diamati
Analisis keparahan penyakit dihitung menggunakan rumus Townsend dan
Heuberger (1943)dalamRekanovicet al. (2007):
%
I : keparahan serangan penyakit (%)
n : jumlah rumpun yang terkena serangan pada skala tertentu V : nilai skala serangan
Tabel 2 Skala keparahan penyakit (IRRI, 1996)
Skala Gejala luas daun terinfeksi (%) Tingkat ketahanan Kelas ketahanan
1 1-5 Tahan T
3 6-12 Agak tahan AT
5 13-25 Agak rentan AR
7 26-50 Rentan R
9 51-100 Sangat rentan SR
Kurva perkembangan penyakit atau AUDPC merupakan luasan total area di
bawah kurva perkembangan penyakit dihitung menurut formula (Narayanasamy
2002):
si: intensitas penyakit pada pengamatan ke-i
d : interval waktu antara dua pengamatan
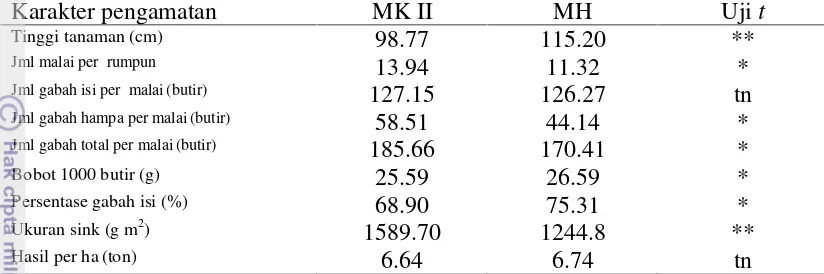
Hasil Dan Pembahasan
Kemunculan Pertama Penyakit HDB
Kemunculan penyakit pada MK II pada genotipe tunggal di Majalengka
pada umur 61 hari setelah semai (hss), demikian pula pada genotipe campuran.
Pada MK II di Pacitan, rata-rata kemunculan pertama penyakit HDB pada
genotipe tunggal lebih awal daripada genotipe campuran yaitu pada 61,57 hss dan
genotipe campuran pada 60.7 hss. Pada Tabel 3 terlihat bahwa pada MH
kemunculan pertama penyakit HDB pada genotipe tunggal di Majalengka berkisar
64 – 92 hss dan pada genotipe campuran berkisar 64 – 112 hss, sedangkan di Pacitan kisaran kemunculan pertama penyakit HDB pada genotipe tunggal adalah
77– 112 hss dan pada genotipe campuran 108– 112 hss.
Genotipe yang mempunyai kemunculan pertama paling lambat (67 hss) di
Pacitan MK II adalah IPB 98-F-5-1-1 dan genotipe campuran IPB
107-F-60-1-1+IPB 102-F-90-2-1. Pada MH di Majalengka, genotipe campuran IPB
23
lain (112 hss), sedangkan di Pacitan terdapat lima genotipe yang tidak terserang
HDB yaitu IPB 102-F-92-1-1, campuran Maros+Ciherang, Ciherang+Cigeulis,
Inpari 6 Jete+Ciherang dan IPB 117-F-7-2+IPB 97-F-13-1-1.
Munculnya gejala awal penyakit HDB terjadi pada saat massa bakteri
sudah cukup terakumulasi untuk menginfeksi tanaman (Mazolla et al. 1994). Perkembangan massa bakteri ditentukan oleh faktor lingkungan yang mendukung,
salah satunya suhu yang hangat. Perbedaan waktu kemunculan pertama penyakit
HDB pada MK II dan MH diduga lebih disebabkan faktor suhu. Suhu pada MK II
yang bertepatan dengan MK II lebih tinggi daripada MH. Kondisi yang hangat
memicu pertumbuhan bakteri sehingga penyakit muncul lebih awal. Pada MH
suhu relatif lebih rendah daripada MK II karena ketersediaan air cukup sepanjang
musim serta adanya curah hujan. Suhu yang relatif rendah membuat bakteri
kurang berkembang sehingga gejala muncul lebih lambat.
Tabel 3 Kemunculan pertama penyakit di Majalengka dan Pacitan pada MK II dan MH (hss)
Genotipe MK II MH
Majalengka Pacitan Majalengka Pacitan
G1 Inpari 6 Jete 61 60 64 101
G2 Inpari 13 61 60 78 96
G3 Ciherang 61 60 64 96
G4 IPB 98-F-5-1-1 61 67 92 112
G5 IPB 97-F-13-1-1 61 64 64 112
G6 IPB 102-F-92-1-1 61 60 78
-G7 IPB 107- F-60-1-1 61 60 69 77
G8 Maros+Ciherang 61 60 64
-G9 Maros+Cigeulis 61 60 64 112
G10 Ciherang+Cigeulis 61 60 64
-G11 Inpari 6 Jete+Inpari 13 61 60 73 112
G12 Inpari 6 Jete+Ciherang 61 60 69
-G13 Maros+Ciherang+Cigeulis 61 60 64 112
G14 IPB 117-F-7-2+IPB
97-F-13-Kejadian, keparahan dan AUDPC Penyakit HDB
Majalengka MK II
Uji t menunjukkan tidak terdapat perbedaan antara genotipe tunggal dan
campuran. Pengamatan kejadian penyakit pertama dan kedua tidak ada perbedaan
genotipe mengalami kejadian penyakit 100% pada pengamatan ketiga dan
keempat.
Analisis terhadap keparahan penyakit pada pengamatan pertama sampai
pengamatan keempat tidak terlihat adanya perbedaan antara genotipe tunggal dan
campuran, demikian pula untuk nilai AUDPC (p>0.05).
0
Gambar 2 Kejadian penyakit di Majalengka
Majalengka MH
Pengamatan kejadian penyakit pertama menunjukkan terdapatnya
perbedaan antara genotipe tunggal dan campuran, dimana genotipe campuran
terlihat lebih tinggi serangannya (p=0.018). Pada pengamatan selanjutnya, tidak
terlihat perbedaan antara genotipe tunggal dan campuran (p>0.05).
Keparahan penyakit pada pengamatan pertama memperlihatkan bahwa
genotipe campuran lebih parah daripada genotipe tunggal (p=0.0075). Pada
pengamatan kedua keparahan penyakit antara genotipe tunggal dan campuran
tidak terdapat perbedaan, namun pada pengamatan ketiga genotipe campuran
25
Gambar 3 Keparahan penyakit di Majalengka
Keparahan genotipe tunggal dan campuran tidak berbeda lagi pada
pengamatan keempat. Analisis terhadap nilai AUDPC memperlihatkan tidak
terdapat perbedaan antara genotipe tunggal dan campuran (p>0.05).
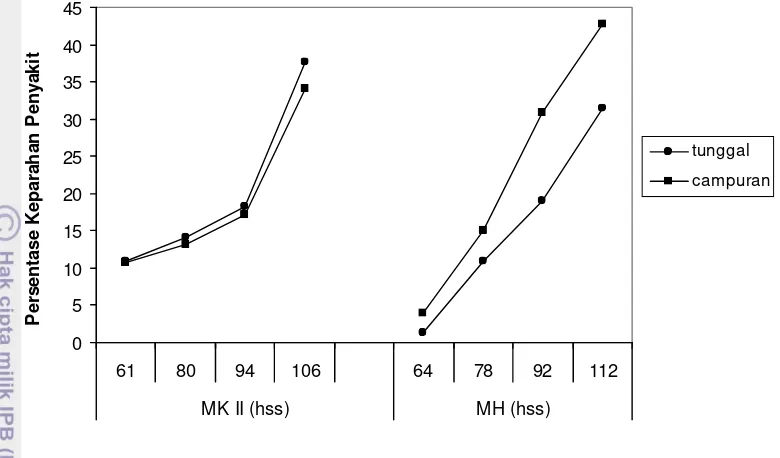
Pacitan MK II
Berdasarkan ujit, kejadian penyakit antara genotipe tunggal dan campuran terlihat pada pengamatan kedua, dimana genotipe campuran mempunyai kejadian
penyakit lebih tinggi (p=0.0035). Pengamatan pertama terhadap kejadian penyakit
tidak terlihat adanya perbedaan antara genotipe tunggal dan campuran, demikian
juga pada pengamatan ketiga, keempat dan kelima (p>0.05).
Keparahan penyakit pada pengamatan pertama belum ada perbedaan
antara genotipe tunggal dan campuran. Pada pengamatan kedua, genotipe
campuran lebih parah daripada genotipe tunggal (p=0.0032), namun pada
pengamatan ketiga keparahan penyakit antara genotipe tunggal dan campuran
tidak terdapat perbedaan. Perbedaan terlihat kembali pada pengamatan keempat,
namun pada pengamatan keempat genotipe tunggal lebih parah daripada genotipe
campuran (p=0.0147). Pada pengamatan kelima, antara genotipe tunggal dan
0
Gambar 4 Kejadian penyakit di Pacitan
Pengamatan pertama ke pengamatan kedua menunjukkan bahwa genotipe
campuran mempunyai nilai AUDPC lebih tinggi (p=0.0119). Nilai AUDPC pada
pengamatan-pengamatan selanjutnya tidak terdapat perbedaan antara genotipe
tunggal dan campuran (p>0.05).
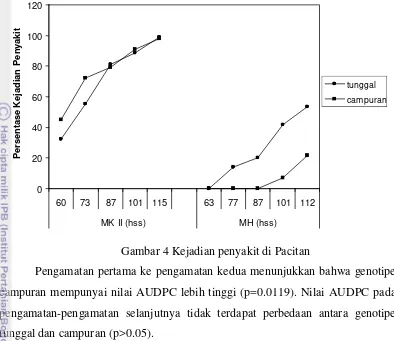
Pacitan MH
Pengamatan pertama belum menunjukkan adanya gejala penyakit sehingga
skor seluruh genotipe adalah nol. Gejala mulai tampak pada pengamatan kedua
dan pada pengamatan kedua ini tidak terdapat perbedaan antara genotipe tunggal
dan campuran. Perbedaan antara genotipe tunggal dan campuran terlihat pada
pengamatan ketiga, keempat dan kelima dimana kejadian penyakit pada genotipe
tunggal lebih tinggi daripada genotipe campuran. Nilai p untuk pengamatan
27
Gambar 5 Keparahan penyakit di Pacitan
Belum terlihat adanya perbedaan keparahan penyakit antara genotipe
tunggal dan campuran pada pengamatan pertama dan kedua. Pada pengamatan
ketiga sampai pengamatan kelima, genotipe tunggal terserang lebih parah
dibandingkan genotipe campuran (nilai p berturut-turut 0.0306, 0.0037 dan
0.007).
Tabel 4 Total nilai AUDPC di Majalengka dan Pacitan pada MK II dan MH
Genotipe Majalengka Pacitan
MK II MH MK II MH
G1 Inpari 6 Jete 5.48 6.04 6.90 0.53
G2 Inpari 13 8.01 6.49 14.46 2.48
G3 Ciherang 8.91 2.51 4.52 1.58
G4 IPB 98-F-5-1-1 9.01 0.62 10.53 0.14
G5 IPB 97-F-13-1-1 10.46 3.49 9.72 0.38
G6 IPB 102-F-92-1-1 6.55 6.36 2.53 0.00
G7 IPB 107- F-60-1-1 7.51 18.55 9.73 5.48
G8 Maros+Ciherang 6.26 6.18 7.40 0.00
G9 Maros+Cigeulis 10.04 5.23 5.51 0.11
G10 Ciherang+Cigeulis 6.55 17.29 5.71 0.00
G11 Inpari 6 Jete+Inpari 13 7.19 9.46 10.25 0.34
G12 Inpari 6 Jete+Ciherang 7.43 9.05 6.33 0.00
G13 Maros+Ciherang+Cigeulis 6.93 9.95 6.76 0.12
G14 IPB 117-F-7-2+IPB 97-F-13-1-1 7.58 4.16 6.13 0.00
G15 IPB 107-F-60+IPB 102-F-90-2-1 7.51 19.63 9.79 1.29 G16 IPB 117-F-7-2+IPB 97-F-13
+IPB 98-F-5-1-1-1 8.89 0.00 8.37 0.05
G17 IPB 102-F-92-1-1+IPB 107-F-60-1-1 +IPB
Analisis nilai AUDPC pada pengamatan pertama ke pengamatan kedua
serta pengamatan kedua ke pengamatan ketiga tidak menunjukkan perbedaan
antara genotipe tunggal dan campuran (p>0.05). Perbedaan AUDPC terlihat pada
pengamatan ketiga ke pengamatan keempat dan pengamatan keempat ke
pengamatan kelima dengan nilai p berturut-turut 0.0128 dan 0.0069.
Perkembangan penyakit HDB di lapangan sangat ditentukan oleh faktor
patogen, lingkungan serta tanaman inang. Benih padi yang ditanam dapat
berfungsi sebagai agen pembawa Xoo sebagai penyebab HDB. Perbedaan yang terjadi pada keempat lingkungan pengujian diduga disebabkan oleh perbedaan
jumlah inokulum awalXoo yang terbawa benih. Menurut Walcott (2003), patogen yang terbawa benih tidak terlihat gejalanya sehingga tidak mungkin dapat
dideteksi secara visual. Populasi patogen terbawa benih umumnya tidak terlalu
besar dan tidak merata jumlahnya pada setiap benih. Keadaan yang demikian
menyebabkan bervariasinya kejadian penyakit, keparahan penyakit dan AUDPC
penyakit HDB pada genotipe yang diuji.
Secara umum, tidak adanya perbedaan antara genotipe tunggal dan
campuran di Majalengka dan di Pacitan pada MK II diduga karena patogen yang
terbawa benih mempunyai kemampuan untuk menyerang tanaman inangnya
didukung oleh faktor lingkungan terutama suhu. Suhu yang hangat pada MK II
sangat mendukung perkembangan bakteri Xoo, sehingga tersedia inokulum yang cukup untuk menyerang tanaman. Seluruh genotipe baik tunggal maupun
campuran dengan berbagai latar belakang ketahanan terhadap Xoo terserang seluruhnya dengan tingkat serangan yang bervariasi. Gejala serangan yang terlihat
pada genotipe tunggal dan campuran tidak memperlihatkan pola yang spesifik.
Pada MH Xoo lebih lambat berkembang karena suhu lebih rendah. Pada MH di Majalengka, tidak ada perbedaan antara genotipe tunggal dan campuran
dalam hal kejadian penyakit, keparahan penyakit dan AUDPC pada akhir
pengamatan. Perbedaan kejadian penyakit, keparahan penyakit dan AUDPC
antara genotipe tunggal dan campuran terlihat di Pacitan, dimana genotipe tunggal
mempunyai nilai yang lebih tinggi. Dapat dikatakan bahwa di Pacitan pada MH,
pencampuran genotipe dapat menekan perkembangan kejadian, keparahan dan
29
Ketahanan yang muncul di Pacitan pada MH adalah ketahanan horizontal.
Genotipe campuran yang mempunyai lebih banyak gen ketahanan dalam
populasinya memperlihatkan gejala yang lebih sedikit. Strain Xoo yang menyerang pun tidak spesifik. Genotipe IPB 107-F-60-1-1 mempunyai sifat tahan
terhadap Xoo strain III, agak tahan strain IV namun agak rentan terhadap strain VIII. Genotipe campuran Ciherang+Cigeulis mempunyai sifat tahan terhadap
strain III dan IV, sedangkan genotipe IPB 107-F-60-1-1+IPB 102-F-90-2-1
mempunyai sifat tahan dan agak rentan terhadap strain III. Meskipun demikian
tidak dapat dikatakan bahwa strain Xoo yang menyerang adalah strain VIII, karena terdapat genotipe lain dengan AUDPC jauh lebih rendah padahal bersifat
agak rentan atau tidak mempunyai ketahanan terhadap strain VIII. Dengan
demikian pola AUDPC tidak berkaitan dengan sifat ketahanan genotipe terhadap
strain tertentu.
Kesimpulan
Hasil analisis uji t untuk membandingkan genotipe tunggal dan campuran pada setiap lokasi pada MK II dan MH diketahui bahwa tidak terdapat perbedaan
kejadian penyakit, keparahan penyakit dan AUDPC di Majalengka MK II,
Majalengka MH dan Pacitan MK II. Perbedaan hanya terlihat di Pacitan pada MH
dimana genotipe tunggal mempunyai kejadian penyakit, keparahan penyakit dan
AUDPC lebih tinggi daripada genotipe campuran. Dapat dikatakan bahwa
pencampuran dapat mengurangi perkembangan penyakit HDB di Pacitan pada
MH.
Daftar Pustaka
Adhikari TB et al. 1995. Genetic diversity ofXanthomonas oryzae pv. oryzae in Asia.Appl. Environ. Microbiol. 61 : 966– 971.
Amadioha AC. 2000. Controlling rice blast in vitro and in vivo with extracts of Azadirachtaindica.Crop Protection 19:287–290.
Djatmiko AH, Fatichin. 2009. Ketahanan dua puluh satu varietas padi terhadap penyakit hawar daun bakteri. J. HPT Tropika 9: 168-173
Finckh MR, Mundt CC. 1992. Stripe rust, yield, and plant competition in wheat cultivar mixtures. Phytopathol 82 : 905– 913.
Garret KA, Mundt CC. 1999. Epidemiology in mixed host population. Phytopathol 89 : 984– 990.
Horino O, Kaku H. 1989. Defense mechanism of rice againts bacterial blight caused by Xanthomonas campestris pv. oryzae di dalam: Banta S.J, Cervantes E, Mew T.W (Editor), Bacterial Blight of Rice, Proc. Of the Int. Workshop on Bacterial Blight of Rice, IRRI, Los Banos, 135– 152.
Horino O, Mew TW, Yamada T. 1982. The effect of temperature on the development of bacterial leaf blight on rice.Ann. Phytopathol. Soc. Jpn 48 : 72– 75
Huanget al. 1997. Pyramiding of bacterial blight resistance genes in rice : marker assisted selection using RFLP and PCR.Theor. Appl. Genet. 95 : 313– 320. [IRRI] International Rice Research Institute.1996.Standard Evaluation System for
Rice. Ed ke-4. INGER Genetic Resources Center.
Ji GH, Wei LF, He YQ, Wu YP, Bai XH. 2008. Biological control of rice bacterial blight byLysobacter antibioticus strain 13-1.Biological Control 45 : 288– 296.
Josi RK, Nayak S. 2010. Gene pyramiding-A broad spectrum technique for developing durable stress resistance in crops.Biotechnol. Mol. Biol. Rev. 5 : 51-60.
Kadir TS, Suryadi Y, Sudir, Machmud M. 2009. Penyakit bakteri padi dan cara pengendaliannya. Inovasi Teknologi Padi, BB Padi.
Lopez CG, Mundt CC. 2000. Using mixing ability analysis from two way cultivar mixtures to predict the performance of cultivar in complex mixtures. Field Crop Research 68 : 121-132
Mahmood T, Marshall D, McDaniel ME. 1991. Effects of winter wheat cultivar mixtures on leaf severity and grain yield. Phytopathol 81 : 471– 474.
Mazzola M, Leach JE, Nelson R, White FF. 1994. Analysis of the interaction between Xanthomonas oryzae pv. oryzae and the rice cultivars IR24 and IRBB21. Phytopathology 84 : 392– 397.
Mew TW, Alvarez AM, Leach JE, Swings J. 1993. Focus on bacterial blight of rice.Plant dis 77 : 5– 12
Mundt CC, Leonard KJ. 1986 Analysis of factors affecting disease increase and spread in mixtures of immune and susceptible plants in computer simulated epidemics.Phytopathol 76:832– 840
Mundt CC. 2002. Performance of wheat cultivars and cultivar mixtures in the presence of Chepalosporium stripe.Crop Protection 21 : 93– 99
31
Ning I, et al. 2012. The effect of wheat mixtures on the powdery mildew disease and some yield components. Journal of Integratif Agriculture 11 : 611 – 620.
Ohtsuki A, Sasaki A. 2006. Epidemiology and disease control under gene-for-gene plant-pathogen interaction.Journal of Theoritical Biology 238 : 780 – 794.
Ou SH. 1985.Rice Disease. Ed ke-2. Commonwealth Mycological Institute. Philip R, Devadath S. 1981. Studies on the physiology of bacterial leaf blight
infected tolerant and susceptible rice cultivar.Phytopathol 101 : 65– 71. Qi Z. 2009. Genetics and improvement of bacterial blight resistance of hybrid rice
in China.Rice Science 16 : 83– 92.
Rajarajeswari NVL, Muralidharan K. 2006. Assesment of farm yield and district production loss from bacterial leaf blight epidemics in rice.Crop Protection 25 : 244– 252.
Rekanovic E, S Milijasevic, B Todorovic, I Potocnik. 2007. Possibilities of biological and chemical control of verticillium wilt in pepper. Phytoparasitica 35:436–441.
Samonte O. 2008. Rice Multilines and Mixtures. Texas Rice. Agrilife Research Texas A&M System VIII : 7.
Sharma RC, Dubin HJ . 1996. Effect of wheat cultivar mixtures on spot blotch (Bipolaris sorokiniana) and grain yield.Field Crop Research 48 : 95 -101. Singh S, et al. 2001. Pyramiding three bacterial blight resistance genes (xa5, xa12
and Xa21) using marker assisted selection into indica rice cultivar PR 106. Theor. Appl. Genet. 102 : 1011– 1015.
Suryadi Y, Kadir TS, Machmud M. 2006. Deteksi Xanthomonas oryzae pv oryzae, penyebab hawar daun bakteri pada tanaman padi. Penelitian Pertanian Tanaman Pangan 25
Wahyudi AT, Meliah S, Nawangsih AA. 2011. Xanthomonas oryzae pv oryzae bakteri penyebab hawar daun pada padi: isolasi, karakterisasi, dan telaah mutagenesis dengan transposon.Makara Sains 15 : 89– 96.
Walcott R.R. 2003. Detection of seedborne pathogens. Horttechnology 13 : 40 -47
Yang CM. 2010. Assesment of the severity of bacterial leaf blight in rice using canopy hypercpestral reflectance.Precision Agric 11 : 61-81