POTENSIAL AKSI TERSTIMULASI ARUS EKSTERNAL SERTA SINKRONISASI CHAOTIK JARINGAN SYARAF
MADA SANJAYA WS
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASINYA
Dengan ini saya menyatakan bahwa tesis Pemodelan dan Simulasi Sistem Dinamika Propagasi Potensial Aksi Terstimulasi Arus Eksternal serta Sinkronisasi Chaotik Jaringan Syaraf adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
MADA SANJAYA WS. Modeling and Simulation Dynamical Systems of Propagating Action Potentials Stimulated by External Current and Chaotic Synchronization in Neuronal Networks. Under direction of AGUS KARTONO and IRZAMAN
System of signals propagation from one neuron to another represent event of very complex electrochemical mechanism. Through model of Hodgkin-Huxley obtained that at membrane there are special channels which can only be entered by certain ion (selective permeable), in experiment which Hodgkin-Huxley proceeded that there are two main channels membrane, potassium channel (K+) and the sodium channel (Na+). That channels were hypotetized had influenced by three variables that is m, h, and n. From analysis of Fitzhugh-Nagumo models, it was obtained a phase space curve which very depend on external current stimulation. When external current was zero, the oscillation was not happened but there will be oscillation after external current was stimulated. But there was limit of maximum current where oscillation of membrane potentials was not happened anymore because the potential have stabilized asymptotically. From approach of dynamics, it also seen bifurcation process when the parameter was changed ,in this case the changes is external current and fulfill theory of Hopf bifurcation with the appearances of critical point that have “stable limit cycle” character. In this paper, it shown that there was qualitative similarity between Hodgkin-Huxley model and Fitzhugh-Nagumo two dimension models. This examination naturally comes to the study of neuron networks. The analysis of these networks uses the synchronization theory via connections between neurons and can give rise to emergent properties and self-organization.
MADA SANJAYA WS. Pemodelan dan Simulasi Sistem Dinamika Propagasi Potensial Aksi Terstimulasi Arus Eksternal serta Sinkronisasi Chaotik Jaringan Syaraf. Dibimbing oleh AGUS KARTONO dan IRZAMAN
Salah satu bidang yang menjadi awal berkembangnya biofisika adalah studi tentang fisiologi syaraf, yang pada beberapa dekade lalu telah dipahami bahwa terjadi proses dasar sistem komunikasi unik elektrokimia yang berperan penting dalam sistem syaraf kita. Otak kita dan setiap subsistem lain pada sistem syaraf terdiri dari sel yang disebut neuron (sel syaraf).
Penelitian ini bertujuan untuk mengkonstruksi model dan membuat simulasi proses propagasi sinyal potensial aksi dari sel syaraf (neuron) tunggal sampai membentuk jaringan syaraf sederhana (neuronal networks) dengan menggunakan model Hodgkin-Huxley dan model Fitzhugh-Nagumo yang telah dimodifikasi, dalam hal ini modifikasinya berupa arus eksternal yang menjadi stimulus dan membuat modifikasi model untuk beberapa neuron yang terkopel membentuk jaringan syaraf (neuronal networks) melalui teori sinkronisasi chaotik.
Sistem propagasi sinyal dari satu neuron ke neuron lainya merupakan peristiwa elektrokimia yang sangat kompleks. Melalui model Hodgkin-Huxley diperoleh bahwa pada membran terdapat saluran-saluran khusus yang hanya dapat dimasuki oleh ion tertentu, dalam eksperimen yang dilakukannya Hodgkin-Huxley menyatakan bahwa terdapat dua saluran utama membran yaitu saluran potassium (ion K+) dan saluran sodium (ion Na+). Kedua saluran tersebut dihipotesiskan dipengaruhi oleh tiga variabel yaitu m, h dan n. Variabel m merupakan probabilitas aktivasi (terbukanya) saluran sodium sedangkan h adalah probabilitas inaktivasi (tertutupnya) saluran sodium dan variabel n yang merupakan probabilitas aktivasi saluran potassium. Dari grafik hasil simulasi parameter Hodgkin-huxley diketahui bahwa m berubah secara cepat sehingga berpengaruh sebagai penyebab terjadinya depolariazation potensial aksi kemudian diikuti dengan variabel n yang menyebabkan setelah mencapai puncak, potensial aksi akan mengalami hyper-polarization dan h merupakan variabel yang paling lambat berubah memiliki peranan sebagai penyebab potensial aksi turun melebihi batas potensial istirahat. Selain itu dalam penelitian ini juga diperoleh grafik pengaruh suhu terhadap terjadinya potensial aksi, dalam penelitian diperoleh bahwa semakin besar suhu yang diberikan maka terjadinya potensial aksi semakin cepat (Depolarization) karena gerakan ion sodium semakin cepat dengan meningkatnya suhu sesuai dengan teori yang mendasarinya. Dengan memasukan data tentang akson tak bermyelin dengan a = 0.0238 cm,
1 ) . ( 4 . 35 /
1
cm
, C 1 F/cm2
m diperoleh kecepatan propagasi potensial aksi sebesar 18.80 m/s pada suhu 6.30C ketika tidak ada arus stimulus eksternal. Sedangkan ketika diberikan stimulus arus eksternal maka kecepatan propagasinya akan semakin besar karena frekuensi osilasinya bertambah besar, hasil ini terdapat kesesuaian dengan penelitian sebelumnya.
eksternal terjadi osilasi pada potensial membran yang memiliki frekuensi bergantung pada arus yang diberikan. Tetapi terdapat batas arus maksimal dimana ketika arus dinaikan tidak lagi terjadi osilasi potensial membran karena potensial sudah stabil asimtotik. Dari pendekatan teori dinamika juga terlihat terjadinya bifurkasi atau perubahan sifat titik kritis ketika parameternya diubah dalam hal ini arus eksternal dan memenuhi teori bifurkasi Hopf dengan kemunculan titik kritis bersifat ”Stable Limit Cycle”. Dalam penelitian inipun dikaji terjadinya gejala chaotik ketika arus stimulus yang diberikan berupa arus stimulus eksternal yang periodik sinusoidal.
Hasil terakhir dalam penelitian ini adalah mempelajari mekanisme komunikasi antara beberapa neuron membentuk jaringan syaraf dengan pendekatan teori sinkronisasi diperoleh bahwa dengan semakin banyaknya neuron yang berinteraksi maka terjadinya ambang sinkronisasi semakin kecil. Dari simulasi yang dibuat terlihat adanya pengaturan diri (self-organization) dari sel neuron yakni ketika jumlah neuron yang berinteraksi semakin banyak maka secara otomatis hanya dibutuhkan konstanta kopling yang kecil untuk menghasilkan sinyal yang sinkron (sesuai antara satu neuron dengan yang lain) meski sinyal dari neuron tunggal berupa potensial aksi yang bersifat chaotik.
Untuk kelanjutan penelitian diperlukan pembuktian secara eksperimen lanjutan mengenai hasil simulasi perhitungan numerik sehingga penelitian teori ini selain menjadi dasar teori juga dapat diaplikasikan secara nyata dalam bidang medis dan neuroscience. Selain itu, metode sistem dinamika dan sinkronisasi chaotik yang terdapat dalam penelitian ini dapat digunakan dalam penelitian bidang lain seperti penelitian karakteristik distribusi potensial pada jantung yang bermanfaat di bidang medis (pemodelan jantung), sistem dinamika penyebaran penyakit (epidemics), sistem dinamika pada ekologi, untuk meningkatkan kekuatan laser, untuk mensinkronisasi output sirkuit elektronik, untuk mengkontrol osilasi pada reaksi kimia, atau untuk mengkode pesan elektronik untuk keamanan komunikasi dan lannya. Sehingga, teori ini dapat dipakai dalam berbagai bidang disiplin ilmu.
(C)
Hak Cipta milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutioan tersebut tidak merugikan kepentingan yang wajar IPB
POTENSIAL AKSI TERSTIMULASI ARUS EKSTERNAL SERTA SINKRONISASI CHAOTIK JARINGAN SYARAF
MADA SANJAYA WS
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biofisika
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Aksi Terstimulasi Arus Eksternal serta Sinkronisasi Chaotik Jaringan Syaraf.
Nama : Mada Sanjaya WS NIM : G751070061
Disetujui Komisi Pembimbing
Dr. Agus Kartono Ketua
Dr. Irzaman Anggota
Diketahui
Ketua Program Studi Biofisika
Dekan Sekolah Pascasarjana
Dr. Akhiruddin Maddu Prof. Dr. Ir. Khairil A. Notodiputro, M.S
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan taufik dan hidayah-Nya, sehingga penulis dapat menyelesaikan tesis berjudul “Pemodelan dan Simulasi Sistem Dinamika Propagasi Potensial Aksi Terstimulasi Arus Eksternal serta Sinkronisasi Chaotik Jaringan Syaraf “, yang dilakukan dalam rangka tugas akhir untuk menyelesaikan program pendidikan S2 Biofisika pada Sekolah Pascasarjana, Institut Pertanian Bogor.
Pada kesempatan ini, penulis mengucapkan banyak terima kasih dan memberikan penghargaan yang sebesar-besarnya kepada Bapak Dr. Agus Kartono dan Bapak Dr. Irzaman atas segala bimbingan dan motivasinya yang diberikan kepada penulis untuk segera menyelesaikan penelitian ini serta kepada Departemen Pendidikan Nasional melalui Beasiswa Unggulan yang diterima penulis. Kepada orangtua dan seluruh sanak keluarga penulis, juga kepada “Dua Rembulan dalam HatiKu”, seluruh staff dan dosen Fisika khususnya, dan IPB pada umumnya. Serta seluruh pihak yang telah membantu penulis yang tak mungkin disebutkan satu-persatu, juga kepada semua teman-teman seperjuangan Fisika.
Penulis memahami bahwa tiada gading yang tak retak. Oleh karena itu, segala macam saran dan kritik yang membangun akan penulis terima dengan senang hati. Semoga apa yang disampaikan oleh penulis akan bermanfaat bagi ilmu pengetahuan.
Bogor, Januari 2009
RIWAYAT HIDUP
Penulis dilahirkan di Cirebon, pada tanggal 11 Oktober 1985 sebagai anak kedua dari tiga bersaudara, pasangan Waryano Sunaryo dan Eti.
Penulis menyelesaikan studinya di SMU Negeri 1 Lemah Abang,Cirebon pada tahun 2003 dan pada tahun yang sama diterima di Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis terdaftar sebagai mahasiswa Departemen Fisika, Fakultas Matematika dan Ilmu Pengetahuan Alam. Selama duduk di bangku kuliah, penulis aktif dalam berbagai kegiatan (kepanitiaan) dan organisasi intra kampus seperti menjadi anggota DKM Masjid Al Hurriyah IPB pada tahun 2003 – 2007 , Departemen Kerohanian Himpunan Mahasiswa Fisika (HiMaFi) IPB pada tahun 2003 – 2004, staff Departemen kerohanian BEM FMIPA pada tahun 2004-2005,staff Departemen Kebijakan Nasional BEM KM IPB pada tahun 2005. Penulis pernah menjadi asisten praktikum mata kuliah Fisika Dasar I dan II 2004-2006, Penulis juga aktif mengajar private di kota Bogor, menjadi staff pengajar A-Project ,Physics Challange dan Best Student Program pada tahun 2004 – 2006. Serta menjadi tutor di Bimbingan Belajar Bintang Pelajar Regional Bogor pada tahun 2006, Bimbingan Belajar Nurul Fikri pada tahun 2007 dan guru fisika SMA Dwi Warna pada tahun 2007.
DAFTAR ISI
DAFTAR TABEL iii
DAFTAR GAMBAR iv
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
1.1 Latar Belakang 1
1.2 Perumusan Masalah 3
1.3 Tujuan Penelitian 3
1.4 Manfaat Penelitian 4
1.5 Ruang Lingkup Penelitian 4
2 MODEL NEURON HODGKIN-HUXLEY 5
2.1 Potensial Membran 5
2.2 Model Sirkuit Hodgkin-Huxley 7
2.3 Pengaruh Suhu pada Model Hodgkin-Huxley 9
2.4 Propagasi Potensial Aksi 10
2.5 Hipotesis Propagasi Seragam 11
3 ANALISIS DAN SIMULASI MODEL HODGKIN-HUXLEY 14
3.1 Analisis Dinamik Model Hodgkin-Huxley 14
3.1.1 Kuat Arus Stimulus I = 0 14
3.1.2 Kuat Arus Stimulus I > 0 16
3.2 Simulasi Pengaruh Suhu pada Model Hodgkin-Huxley 17 3.3 Metode Numerik Kecepatan Propagasi Potensial Aksi 18
4 MODEL DINAMIK NEURON FITZHUGH-NAGUMO 20
4.1 Model Dinamik Neuron Fitzhugh-Nagumo 20
4.2 Teori Dasar Sistem Dinamik 21
4.2.1 Sistem Dinamik dan Deterministik 21
4.2.2 Persamaan Diferensial Orde Pertama 21
4.2.4 Konstruksi Matrik Jacobi 22
4.2.5 Vektor Eigen dan Nilai Eigen 22
4.2.6 Orbit Kestabilan 23
4.2.7 Bifurkasi Hopf 24
4.3 Penentuan Titik Kritis Model Fitzhugh-Nagumo 25
4.4 Matrik Jacobian Model Fitzhugh-Nagumo 26
4.5 Nilai Eigen dan Syarat Kestabilan Model Fitzhugh-Nagumo 26 4.6 Analisis Kestabilan Titik Kritis Model Fitzhugh-Nagumo 26
4.6.1 Kasus Arus Stimulus I = 0 27
4.6.2 Kasus Arus Stimulus I = 0.33 28
4.6.3 Kasus Arus Stimulus I = 1.25 29
4.6.4 Kasus Arus Stimulus I = 1.43 30
4.6.5 Kasus Arus Stimulus I = 1.45 31
4.6.6 Kasus Arus Stimulus I = 2 32
5 SINKRONISASI CHAOTIK NEURONAL NETWORK 34
5.1 Gejala Chaos Model Neuron Fitzhugh-Nagumo 34
5.2 Sensitivitas Terhadap Kondisi Awal 36
5.3 Kopling Neuron Model Fitzhugh-Nagumo 38
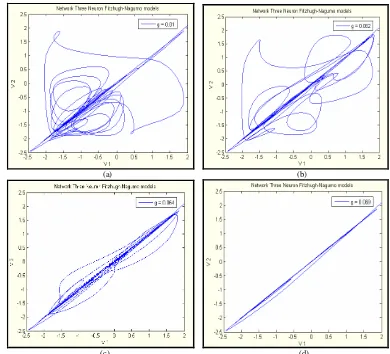
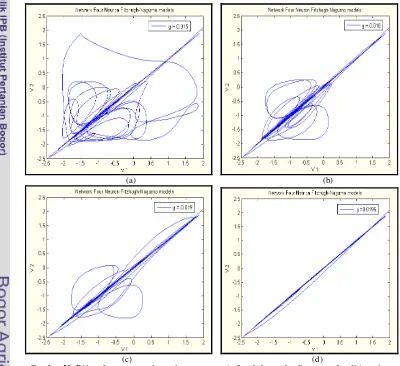
5.3.1 Model Dua Neuron Terkopel Fitzhugh-Nagumo 39 5.3.2 Model Tiga Neuron Terkopel Fitzhugh-Nagumo 41 5.3.3 Model Empat Neuron Terkopel Fitzhugh-Nagumo 42
SIMPULAN 45
DAFTAR TABEL
DAFTAR GAMBAR
Gambar 1. Struktur dasar neuron 2
Gambar 2. Model sirkuit membran Hodgkin-Huxley 7 Gambar 3. Sistem dinamika membran model Hodgkin-Huxley saat
I = 0; (a) potensial aksi saat t = 5 ms, (b) dinamika
variabel m, h dan n saat t = 5 ms 14
Gambar 4. Sistem dinamika membran model Hodgkin-Huxley saat I > 0; (a) potensial aksi (mV) saat t = 100 ms saat diberi stimulus I = 7 mA, (b) dinamika variabel m, h dan n saat t = 100 ms, I = 7 mA,(c) potensial aksi (mV) saat t = 100 ms diberi stimulus I = 15 mA, (d) dinamika variabel m, h
dan n saat t = 100 ms diberi stimulus I = 15 mA 16 Gambar 5. Sistem dinamika membran model Hodgkin-Huxley saat
I =15 mA; (a) grafik hubungan antara potensial aksi (mV)
dan variable n, (b) grafik 3D hubungan antara V, n dan t 17 Gambar 6. Sistem dinamika model Hodgkin-Huxley dengan variasi
suhu 6.30C, 18.50C, 250C, dan 320C 18 Gambar 7. Propagasi Potensial aksi pada saat K = 3x 105 19 Gambar 8. Orbit kestabilan disekitar titik kritis; (a) spiral stabil,
(b) spiral tak stabil, (c) titik saddle, (d) center, (e) titik
stabil dan (f) titik tak stabil 24
Gambar 9. Bifurkasi pada persamaan van der Pol ketika parameter
divariasikan 25
Gambar 10. Sistem dinamika membran model Fitzhugh-Nagumo saat I = 0; (a) bidang fase antara v dan w bersifat spiral stabil
(b) dinamika v, w terhadap waktu t. 27
Gambar 11. Sistem dinamika membran model Fitzhugh-Nagumo saat I = 0.33; (a) bidang fase antara v dan w bersifat stabil limit cycle, (b) dinamika v, w terhadap waktu t = 100, (c) dinamika v, w terhadap waktu t = 100 dan (d) grafik
Gambar 12. Sistem dinamika membran model Fitzhugh-Nagumo saat I = 1.25; (a) bidang fase antara v dan w bersifat spiral tak
stabil (b) dinamika v, w terhadap waktu t. 30 Gambar 13. Sistem dinamika membran model Fitzhugh-Nagumo saat
I = 1.43; (a) bidang fase antara v dan w bersifat spiral stabil (b) dinamika v, w terhadap waktu t. 31 Gambar 14. Sistem dinamika membran model Fitzhugh-Nagumo saat
I = 1.45; (a) bidang fase antara v dan w bersifat spiral stabil (b) dinamika v, w terhadap waktu t. 31 Gambar 15. Sistem dinamika membran model Fitzhugh-Nagumo saat
I = 2; (a) bidang fase antara v dan w bersifat stabil node
(b) dinamika v, w terhadap waktu t. 32
Gambar 16. Model Fitzhugh-Nagumo saat A = 0.7; (a) respon waktu saat = 0.4 (b) bidang fase saat = 0.4, (c) respon waktu saat = 0.8145 (d) bidang fase saat = 0.8145, (e) respon waktu saat = 0.93 dan (f) bidang fase saat
= 0.93 35
Gambar 17. Model Fitzhugh-Nagumo saat A = 0.7 dan = 0.945 serta memvariasikan kondisi awal potensial aksi V0 36
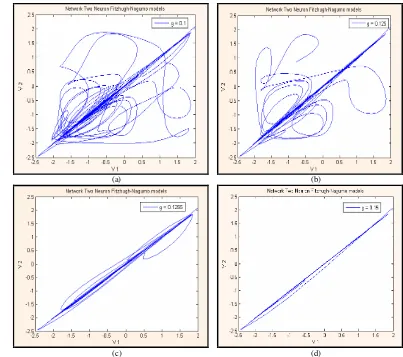
Gambar 18. Bidang fase antara v1 dan v2 dengan memvariasikan
kekuatan kopling gs saat kondisi awalnya tak sama;
(a) gs = 0.1, (b) gs = 0.125, (c) gs = 0.1255, dan
(d) gs = 0.15 40
Gambar 19. Bidang fase antara v1 dan v2 dengan memvariasikan
kekuatan kopling gs saat kondisi awalnya tak sama;
(a) gs = 0.01,(b) gs = 0.062, (c) gs = 0.064, dan
(d) gs = 0.069. 42
Gambar 20. Bidang fase antara v1 dan v2 dengan memvariasikan
kekuatan kopling gs saat kondisi awalnya tak sama;
(a) gs = 0.015, (b) gs = 0.018, (c) gs = 0.019, dan
DAFTAR LAMPIRAN
Lampiran 1. Penurunan Persamaan Kabel 49
Lampiran 2. Analisis Titik Kritis Model Dinamika Fitzhugh-Nagumo 54 Lampiran 3. Analisis Nilai Eigen Model Dinamik Fitzhugh-Nagumo 55 Lampiran 4. Sintaks Plot Grafik Dengan Software Matlab 7.01 57
PENDAHULUAN
1.1 Latar Belakang
Salah satu bidang yang menjadi awal berkembangnya biofisika adalah studi tentang fisiologi syaraf, yang pada beberapa dekade lalu telah dipahami bahwa terjadi proses dasar sistem komunikasi unik elektrokimia yang berperan penting dalam sistem syaraf kita. Otak kita dan setiap subsistem lain pada sistem syaraf terdiri dari sel yang disebut neuron (sel syaraf).
Dalam neuron terdapat axon berupa struktur panjang menyerupai tabung (Edelstein, 1988) dan diketahui bahwa propagasi sinyal syaraf berasal dari listrik yang timbul secara alami. Setelah menginisiasi bagian yang disebut axon hillock, propagasi turun melalui axon ke terminal berikutnya dengan konveksi bebas (synapses) yang dekat dengan neuron. Sinyal propagasi tersebut disebut sebagai ”potensial aksi”. Sebuah neuron memiliki bagian yang disebut dendrit yang menerima sinyal yang diterimanya dan membawanya menuju soma (badan sel).
Gambar 1. Struktur dasar neuron
Deskripsi lengkap telah dilakukan pada tahun 1952 oleh Hogkin, Huxley dan Katz dengan melakukan eksperimen pada sebuah axon berukuran besar dari squid (gurita). Setelah melakukan eksperimen, mereka membuat sebuah model membran yang analogi dengan sirkuit listrik yang memiliki kandungan fisis, seperti konduktivitas ionik yang digambarkan dengan sebuah elemen sirkuit berupa resistor. Model Hodgkin-Huxley adalah berupa empat persamaan diferensial nonlinier terkopel dan membuat hipotesis dengan mengusulkan adanya tiga variabel m, h, dan n yang mempengaruhi koduktivitas ion K+ dan ion Na+ saat melewati membran. Keempat persamaan ODE (Ordinary Differential equation) ini sulit untuk dipecahkan secara eksak, karena derajat nonliniernya yang tinggi. Tetapi dengan memanfaatkan sifat dinamika dari keempat variabel tersebut makna fisis dari eksperimen Hodgkin-Huxley dapat diteliti. Untuk membuat analisis yang lebih umum, pada tahun 1961 Fitzhugh dan Nagumo membuat sebuah model penyederhanaan dari model Hodgkin-Huxley menjadi dua persamaan diferensial nonlinier terkopel. Model yang diusulkan ini mampu menerangkan proses dasar eksitasi dan osilasi pada neuron secara kualitatif (Edelstein KL 1988; Medvedev GS, Kopell N 2001; Georgiev NV 2003).
syaraf dapat dipelajari dengan pendekatan chaos controlling (Mishra D et al 2006) dan sistem terkopel (Medvedev GS, Kopell N 2001; Igor Belykh et al 2008).
Permasalahan menarik dalam penelitian pemodelan neuron adalah adanya arus eksternal sebagai trigger yang menghasilkan potensial aksi (impuls syaraf) sebagai informasi dari satu neuron menuju neuron lainnya dalam neuronal network. Arus eksternal ini dapat berupa arus konstan, arus periodik maupun berupa medan listrik (Mishra D et al 2006; Sims BA 2008)
1.2 Perumusan Masalah
a. Bagaimanakah membangun persamaan yang dapat menjelaskan fenomena terjadinya potensial aksi ( impuls syaraf ) dan apakah fenomena tersebut merupakan sistem yang bersifat deterministik ataukah non-deterministik. b. Bagaimanakah memodelkan sistem tersebut dan apakah simulasinya
memberikan hasil yang sesuai kenyataan (eksperimen).
c. Bagaimanakah bentuk persamaan yang lebih umum yang dapat menjelaskan fenomena perambatan potensial aksi yang secara kualitatif terdapat kemiripan dengan hasil eksperimen.
d. Jika telah dibangun persamaan umum untuk sebuah sel neuron, bagaimanakah bentuk persamaan yang dapat menjelaskan interaksi antara banyak neuron membentuk jaringan syaraf.
1.3 Tujuan
a. Mempelajari model Dinamika nonlinier yang berlaku pada sistem perambatan impuls sel syaraf.
f. Membandingkan hasil analisis model Hodgkin-Huxley dengan model Fitzhugh-Nagumo.
g. Membuat model mekanisme sinkronisasi jaringan syaraf berdasarkan model Fitzhugh-Nagumo serta Memahami arti fisis dari semua pemodelan impuls syaraf.
1.4 Manfaat Penelitian
Penelitian ini menjadi dasar acuan teori biofisika tentang mekanisme kerja sel syaraf yang berawal dari transport membran menjadi sinyal potensial aksi yang dapat ditransfer dari satu sel ke sel lainnya. Selain itu, mekanisme inipun memiliki kesamaan dengan sistem sel lainya sehingga dapat digunakan sebagai dasar mempelajari mekanisme yang terjadi pada berbagai sel makhluk hidup yang bermanfaaf bagi dunia medis, fisiologi maupun bioteknologi.
Selain terkait bidang neurologi prinsip kerja pemodelan dan simulasi jaringan syaraf yang berdasar pada teori sistem dinamika, ini dapat diaplikasikan juga untuk bidang lainnya seperti sinkronisasi sinar laser dibidang sains dan teknik, propagasi gelombang seismik pada seismologi, pemodelan sistem jaringan otot jantung dalam bidang medis, dan sinkronisasi pada sirkuit pada bidang elektronika dan instrumentasi.
1.5 Ruang Lingkup
MODEL NEURON HODGKIN-HUXLEY
2.1 Potensial Membran
Saluran ionik pada membran terlihat sebagai pori makroselular. Saluran tersebut dapat melewatkan molekul memasuki membran. Terdapat saluran antara bagian dalam dan luar sel. Terdapat banyak tipe saluran ionik. Sebagai contoh, kita bisa lihat itu dapat terbuka yang bergantung pada potensial atau menseleksi molekul yang akan melewatinya. Dua diantaranya bergantung pada potensial yaitu saluran sodium dan saluran potassium (Jackson MB 2006).
Disebut saluran sodium karena khusus untuk ion sodium. Faktanya, mereka memiliki kekhususan: mereka bisa dalam keadaan aktif atau tidak aktif. Saluran potassium terbuka dan tertutup dengan delay. Saluran leak (kebocoran) selalu terbuka.
Neuron ditutupi sebuah membran yang membatasi bagian dalam sel dari bagian luar sel. Dalam sel, konsentrasi ion berbeda dari cairan yang mengelilinginya. Perbedaan konsentrasi menghasilkan potensial listrik yang berperan penting dalam Dinamika neuronal.
Ketika neuron tidak mengirim sinyal, disebut “keadaan istirahat”. Dalam kasus ini, bagian dalam neuron negative dibandingkan dengan bagian luarnya. Saat istirahat, ion potassium (K+) dapat memasuki membran dengan mudah. Juga saat istirahat, saluran sodium (Na+) mengalami kesulitan saat melewatinya. Muatan negatif molekul protein di dalam neuron tidak dapat melewati membran. Ketika beda potensial antara dalam dan luar neuron diukur, kita dapatkan potensial istirahat. Potensial istirahat membran neuron adalah sekitar -70 mV, artinya bagian dalam neuron memiliki potensial 70 mV lebih kecil dibandingkan bagian luarnya. Ketika istirahat, terdapat lebih banyak ion sodium di luar neuron dan lebih banyak ion potassium di dalam neuron (Corson N et al 2007).
arah yang sama. Keadaan equilibrium dapat diprediksi melalui persamaan Nernst, pada membran sel berbentuk;
inp out S
S S zF RT v
] [
] [ ln
(1)
Potensial istirahat memberikan informasi kepada kita apa yang terjadi ketika neuron istirahat. Potensial aksi terjadi ketika neuron mengirim informasi melewati axon menjauhi tubuh sel. Ahli syaraf menggunakan nama lain, seperti ”spike” atau ”impulse” untuk potensial aksi. Potensial aksi adalah sebuah ledakan dari aktivitas listrik yang terbentuk dengan arus depolarisasi. Artinya, sebuah stimulus menyebabkan potensial istirahat bergerak ke arah 0 mV. Ketika polarisasi naik sekitar -55 mV, neuron akan mengalami potensial aksi. Ini yang disebut batas ambang. Jika neuron tidak melewati batas ambang, tidak terjadi potensial aksi (Corson N et al 2007).
2.2 Model Sirkuit Hodgkin – Huxley
Model ini menggunakan sirkuit listrik yang memiliki kandungan fisis seperti konduktivitas ionik yang digambarkan dengan sebuah elemen sirkuit berupa resistor (Hodgkin AL & Huxley AF 1952; Marquie P 2004). Tegangan melewati membran memiliki analogi tegangan yang melewati resistor. Salah satu penggambaran pori ionik yang selektif terhadap arus ionik. Akan kembali dibahas bahwa konduktivitas pori tergantung dari tegangan.
Gambar 2. Model sirkuit membran Hodgkin-Huxley
Dalam sirkuit yang analogi dengan axon, hambatan aliran ionik melewati membran digambarkan dengan gK, gNa, dan gL. Hambatan dari gerakan ionik di
dalam axon pada suatu arah digambarkan dengan elemen longitudinal dengan hambatan persatuan panjang, bernilai tetap dan lebih tinggi dari medium dibandingkan dengan bagian luar membran (axon memiliki jari-jari yang kecil sehingga hambatannya menjadi besar). Ukuran finite dari membran diasosiasikan dengan kandungan kapasitansinya yang sebanding dengan muatannya. Kita harus ingat, bahwa bentuk axon adalah silinder, sehingga perlu adanya modifikasi dalam menganalogikan.
Terdapat sebuah hubungan
) , ( 2 ) ,
(x t aCv x t
q (2)
hanya dapat berubah dengan arus yang membawa ion melewati membran sel. Dalam kasus ini, perubahan muatan internal rata-rata dapat ditulis,
i aI dt dq 2 (3)
Arus i dapat diekspresikan menjadi penjumlahan INa, IK,dan IL (sodium,
potassium dan ion lainnya) yang berelasi dengan beda potensial yang menyebabkannya. Seperti persamaan berikut;
) (
2 a INa IK IL
dt
dq (4)
) ( ) ( ) ( L L L K K K Na Na Na v v g I v v g I v v g I (5)
Dimana vNa, vK dan vL menggambarkan potensial membran karena adanya berkontribusi dari ion Na, K, dan L. Lebih jauh, persamaaan (4) dapat ditulis kembali sebagai,
gNa v v vNa gK v v vk gL v vL Ieks
C dt
dv
) ( ) )( ( ) )( ( 1 (6)
Dengan asumsi bahwa gL konstan. Pada titik ini Hodgkin-Huxley dan Katz menganalisis listrik secara langsung dan menghitung mekanisme yang mungkin dari konduktivitas ionik yaitu gNa dan gK. Setelah membuat trial and error model, dengan kerja keras dipecahkan melalui kalkulkator mekanik. Mereka menemukan bahwa diperlukan tiga buah variabel n, m dan h pada Dinamika ion pori. Hipotesis ini secara kuantitatif mungkin berkaitan dengan penggambaran konsentrasi pada protein yang harus diberikan untuk membuka atau menutupnya sebuah pori. Bagaimanapun, persamaan ini menggunakan data experimen bukan berasal dari pemahaman dasar dari mekanisme molekular (Jackson MB 2006).
h m g
gNa Na 3 (7)
4 n g
gK K (8)
Dimana g adalah parameter konduktivitas yang konstan. Mereka mengajukan n, m dan h sensitif terhadap tegangan pada jembatan protein. Mereka membuat persamaan diferensial yang bergantung tegangan, yang digambarkan,
n v n v dt dn n n( )(1 ) ( )
(9a) m v m v dt dm m m( )(1 ) ( )
(9b) h v h v dt dh h h( )(1 ) ( )
(9c)
Dengan beberapa variabel yang diasumsikan,
1 10 / ) 40 ( ) 1 )( 40 ( 1 . 0 )
( v
m v v e
,
m(v)4e(v65)/18 (10a) 20 / ) 65 ( 07 . 0 )( v
h v e
, ( 35)/10 1
) 1 (
)
( v h v e
(10b) 1 10 / ) 55 ( ) 1 )( 55 ( 01 . 0 )
( v
n v v e
,n(v)0.125e(v65)/80(10c)
Nilai konstanta lain yang terdapat pada persamaan yaitu gNa = 120, gK= 36 dan gL = 0.3 mmho cm-2, vNa = 50 mV, vK = -77 mV dan vL = -54.4 mV. Dari persamaan diatas (6 dan 9) terdapat empat persamaan diferensial biasa yang terkopel sehingga berdimensi (v, n, m, h) yang merupakan inti persamaan Hodgkin-Huxley.
Kesuksesan dalam model ini adalah yaitu kemampuannya untuk memprediksi secara akurat hasil dari banyak pengamatan eksperimen.
2.3 Pengaruh Suhu Pada Model Hodgkin Huxley
faktor koreksi terhadap variabel dan , dan sangat efektif untuk banyak aplikasi (Fitzhugh R 1966; Kuang S 2008).
Faktor koreksi tersebut adalah berupa mengkalikan bagian kanan persamaan (9) dengan faktor suhu k,
10 / ) 3 . 6 ( 3 T k (11)
2.4 Propagasi Potensial Aksi
Untuk propagasi potensial aksi, membrannya tereksitasi di atas ambang. Di bawah kondisi ini, persamaan sebagai berikut :
v t
i t v c
i ion m
m m
m ,
(12)
dengan iion adalah arus ionik tiap unit panjang. Melalui substitusi Persamaan (12) ke Persamaan kabel (lampiran 1) berikut :
m m e i i x v r
r
2
2 1
(13)
Diperoleh persamaaan atas-ambang yang sesuai adalah. :
) , ( 1 2 2 t v i t v c x v r
r ion m
m m m e i (14)
Pada kondisi ini, untuk kasus khusus serat lingkar dengan radius a, parameter rm, cm, dan ri pada Persamaan (14) bisa diekspresikan dalam bentuk Rm, Cm, dan σi, yaitu secara berurutan, resistansi membran dikali unit luas, kapasitansi per unit luas, dan konduktivitas intraselular:
rm =
a Rm
2
cm = 2πaCm
ri =
i
a
Dengan potensial membran lagi-lagi dinyatakan dalam bentuk deviasi dari nilai potensial diamnya (resting potential), Vr. Jika diasumsikan kabel melingkar dan re = 0, didapatkan,
) , ( 2 2 2 t v I t v C x v a m ion m m m i (15)
Dengan Iion adalah arus ionik tiap unit luas membran. Dengan mengganti nilai Iion, didapatkan
) ( ) ( ) ( 2 2 2 L r m L Na r m Na K r m K m m m i E V g E V g E V g t C x a (16)
Variasi pada konduktansi gK dan gNa diberikan oleh persamaan (7) dan (8) (Hodgkin AL & Huxley AF 1952; Gulrajani RM 1998; Muratov CB 2008).
2.5 Hipotesis Propagasi “Seragam”
Untuk mencari solusi dari potensial aksi propagasi, bagaimanapun, harus dipecahkan persamaan differensial parsial (16). Pada tahun 1952, dengan hanya menggunakan kalkulator mekanik sebagai panduannya, Hodgkin dan Huxley tidak memecahkan Persamaan (16). Namun, dengan dibantu pengamatan eksperimennya, mereka mengasumsikan bahwa potensial aksi muncul dengan kecepatan konstan sepanjang akson, dan, karena sifat regeneratif dari membran, tanpa adanya penurunan atau distorsi pada bentuk gelombangnya. Propagasi tanpa penurunan dengan kecepatan konstan ini dinamakan propagasi “seragam” . Sebagai hasilnya, dapat ditulis
2 2 2 2 2 1 t x m m (17)
waktu 0 dan t. Jika dimisalkan saat t = 0, variasi ruang dari potensial aksi sepanjang akson diberikan oleh ύm(x), di bawah dua asumsi dari Hodgkin dan Huxley, dapat dengan mudah dilihat bahwa untuk tiap nilai x dan t, harus diperoleh hubungan
) ( ) ,
(x t m x t
m
(18)
Akibatnya bisa ditulis turunan berikut:
, ' m m v x m m v x " 2 2
(19)
, 'm m v t v m m
v
t
" 2 2 2
(20)dengan persamaan awal menyatakan turunan dari ύmterhadap nilai (x –t). Jelas, dari Persamaan (19) dan (20), vm memenuhi persamaan gelombang Persamaan (17). Dengan melakukan substitusi Persamaan (17) ke dalam Persamaan (16), didapatkan
m r K
Na
m r Na
L
m r L
K m m m
i g v V E g v V E g v V E
dt dv C dt v d
a
2 2 2 2 (21)
Dengan mengasumsikan karakteristik membran (Cm, gK, gNa, gL) tetap tak berubah, setiap solusi dari Persamaan (21) tetap merupakan solusi jika nilai aσi/2Cm2 konstan (=1/K). Lalu, kecepatan propagasi yang baru didapatkan dengan berbagai cara selain yang mempengaruhi membran, misalnya, yang didapatkan dengan merubah konduktivitas aksoplasmik atau dengan mengubah diameter akson, yang harus memenuhi hubungan
2 / 1
2
m i
C
Ka
(22)
ANALISIS DAN SIMULASI MODEL HODGKIN-HUXLEY
3.1 Analisis Dinamika Model Hodgkin Huxley
Persamaan Hodgkin-Huxley berisi empat persamaan ODE terkopel dengan derajat nonlinear yang tinggi dan sangat sulit untuk dicari pemecahan secara matematikanya bahkan dengan menggunakan komputasi sekalipun. Tetapi meski tidak dapat dicari solusi secara eksaknya, dari persamaan tersebut disertai dengan parameter hasil eksperimen kita dapat mengambil banyak informasi mengenai perilaku eksitasi dan osilasi yang terjadi pada membran axon sel syaraf melalui grafik Dinamika potensial aksi dan variabel m, n, dan h.
3.1.1 Kasus Arus Stimulus I = 0
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan memasukan nilai-nilai parameter (10) dapat diperoleh grafik hubungan tiga variabel m, n dan h serta grafik Dinamika potensial aksi ketika tidak ada pemberian arus (impuls) eksternal .
[image:31.612.142.492.481.659.2]
(a) (b)
Gambar 3 (a) menunjukan mekanisme potensial aksi yanng terdiri dari tiga tahapan, yang terdiri dari tahap pertama fase naik (Depolarization) yaitu peningkatan potensial yang tajam dan bernilai positif dalam gambar terlihat potensial maksimum pada 35-38 mV. tahap kedua fase jatuh (hyper-polarization) yaitu fase mengiringi fase naik yang cepat, terjadi penurunan drastis pada potensial membran yang akhirnya melampaui potensial istirahat hingga mencapai 75 mV. Fase ketiga adalah fase balik yaitu Potensial membran berangsur-angsur kembali ke nilai istirahat pada potensial sekitar 65-70 mV. Jika tidak ada arus eksternal yang mempengaruhi maka potensial membran akan konstan stabil pada kondisi istirahat yang terlihat dalam gambar 3 (a).
Sedangkan gambar 3 (b) menggambarkan Dinamika antara variabel m, h dan n. Dari grafik terlihat bahwa variabel m merupakan variabel yang meningkat dan berubah paling cepat, dimana variabel m memiliki peranan fase pertama dari potensial aksi yaitu terjadinya depolarisasi, jika m adalah peluang membukanya tiga subunit saluran ion sodium (Na+) maka meningkatnya m menyebabkan terbukanya subunit saluran sodium dan membran menjadi berlimpah ion sodium (Na+) sehingga meningkatkan potensial aksi (Depolarization). Setelah variabel m meningkat kemudia diikuti dengan meningkatnya variabel n dengan perubahan yang lebih lambat. Variabel n memiliki peranan pada fase kedua potensial aksi yakni fase jatuh (hyper-polarization) karena jika variabel n adalah peluang membukanya empat subunit saluran ion potassium maka dengan meningkatnya variabel n menyebabkan banyak ion potassium yang keluar dari membran dan potensial membran menjadi turun sedangkan penurunan variabel n berarti peluang menutupnya saluran potassium sehingga potensial membran kembali pada fase istirahat. Sedangkan variabel h merupakan variabel yang paling lambat berubah, jika variabel h merupakan peluang menutupnya saluran sodium maka dengan meningkatnya h menyebabkan saluran sodium tertutup dan penurunan tegangan berlangsung sampai melebihi batas potensial istirahat.
3.1.2 Kasus Arus Stimulus I > 0
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan memasukan nilai-nilai parameter (10) dapat diperoleh grafik hubungan tiga variabel m, n dan h serta grafik Dinamika potensial aksi ketika diberi arus (impuls) eksternal.
Pada kasus arus eksternal diberikan sebesar 7 mA maka potensial aksi akan mengalami osilasi secara periodik. Begitupun dengan Dinamika variabel m, h dan n akan berosilasi secara periodik. Jika kita naikan terus arus eksternalnya maka frekuensi osilasi dari potensial aksi akan semakin besar. Pada gambar 5 dapat dilihat grafik osilasi potensial aksi dan Dinamika variabel m, h dan n pada kasus arus eksternal yang diberikan sebesar 15 mA.
(a) (b)
[image:33.612.145.494.330.607.2]
(c) (d)
Gambar 4. Sistem Dinamika membran model Hodgkin-Huxley saat I > 0 ; (a) potensial aksi (mV) saat
t = 100 ms saat diberi stimulus I = 7 mA, (b) Dinamika variabel m, h dan n saat t = 100 ms I = 7 mA, (c) potensial aksi (mV) saat t = 100 ms diberi stimulus I = 15 mA, (d) Dinamika variabel m, h dan n saat
(a) (b)
Gambar 5. Sistem Dinamika membran model Hodgkin-Huxley saat I =15 mA ; (a) grafik hubungan antara potensial aksi (mV) dan variable n, (b) grafik 3D hubungan antara V, n dan t
Dari grafik pada gambar 4 (c) terlihat jelas bahwa frekuensi osilasi dari potensial membran akan semakin bertambah seiring dengan bertambah besarnya arus eksternal yang diberikan , hal ini dapat terlihat jika kita membandingkan gambar 4 (a) dengan gambar 4(c). konsekuensi yang sama pun berlaku pada Dinamika variabel m, h dan n. Dalam gambar 4 (a) dan 4 (c) juga terlihat bahwa
potensial maksimum sekitar 35 mV sedangkan potensial terendah sekitar – 70 mV.
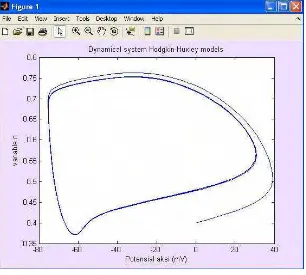
Gambar 5 (a) dan 5 (b) menggambarkan grafik ruang fase dari potensial aksi terhadap variabel n. Pada grafik terlihat ruang fase tersebut membentuk sebuah trayektori “stable limit cycle” disekitar titik yang ditunjukkan tanda panah. Titik tersebut sebenarnya merupakan titik kritis (steady state) dimana semua vektor dari kondisi awal akan mengelilinginya dan pada titik tersebut juga dimungkinkan merupakan titik tejadinya bifurkasi dari satu keadaan ke keadaan yang lain. Secara lebih lengkap titik kritis tersebut akan dibahas pada model Fitzhugh-Nagumo.
3.2 Simulasi Pengaruh Suhu pada Potensial Aksi Model Hodgkin-Huxley
Gambar 6. Sistem Dinamika model Hodgkin-Huxley dengan variasi suhu 6.30C, 18.50C, 250C, dan 320C
Dalam gambar 6. memperlihatkan semakin besar suhu yang diberikan maka terjadinya potensial aksi semakin cepat (Depolarization) karena gerakan ion sodium semakin cepat dengan meningkatnya suhu sesuai dengan teori yang mendasarinya (Fitzhugh R 1966; Kuang S 2008).
3.3 Metode Numerik Penentuan Kecepatan Propagasi Potensial Aksi
Dengan sedikit manipulasi matematika persamaan (21) yang merupakan persamaan diferensial orde dua dapat diubah menjadi persamaan diferensial biasa berode satu (Scott AC 2002; Muratov CB 2008), melalui pemisalan sebagai berikut : w dt dv (23) ) / )) ( ) ( ) ( ( (
2 4 3
2 m L l Na Na K K m i C v v g v v h m g v v n g w C a dt
dw
(24)
n v n v dt dn n n( )(1 ) ( )
(9a) m v m v dt dm m m( )(1 ) ( )
(9b) h v h v dt dh h h( )(1 ) ( )
Sekarang persamaan (21) telah diubah menjadi persamaan diferensial biasa berode satu yang terkopel sehingga terdapat lima variabel (v, w, n, m, h) yang merupakan fungsi waktu. Dengan teknik trial and error akan dihitung besarnya kecepatan propagasi potensial aksi pada axon gurita, sebagai berikut :
Gambar 7. Propagasi Potensial aksi pada saat K = 10470
Dengan memasukan data tentang akson tak bermyelin dengan a = 0.0238
cm, 1
) . ( 4 . 35 /
1
cm
, 2
/ 1 F cm
MODEL DINAMIKA NEURON FITZHUGH-NAGUMO
4.1 Model Dinamika Neuron Fitzhugh-Nagumo
Dalam papernya pada tahun 1961, Fitzhugh mengusulkan untuk menerangkan model Hodgkin-Huxley menjadi lebih umum, yang di dalamnya terdapat eksitasi dan osilasi. Pendekatan serupapun telah dilakukan secara terpisah oleh Nagumo (1962), sehingga model itu disebut persamaan Fitzhugh- Nagumo.
Untuk menghindari kesalahan persepsi, maka harus ditegaskan bahwa tujuan utama model ini tidak untuk menggambarkan kandungan kuantitatif secara akurat dari impuls axon, variabel dari persamaan tersebut memiliki apa yang disebut ketidaktelitian dan hubungan relasi tidak berhubungan secara eksak dari fakta fisiologi atau hanya perkiraan saja. Juga, sistem ini menjadi lebih sederhana. Dimana, kita bisa menunjukkan interaksi singkat antara variabel yang menunjukkan sebuah eksitasi dan osilasi (impuls berulang). Persamaan Fitzhugh (persamaan tak berdimensi),
) (
3 1 3
bw a v dt dw
I w v v dt dv
(25)
4.2 Teori Dasar Sistem Dinamika
4.2.1 Sistem Dinamika dan Deterministik
Dinamika berhubungan dengan perubahan perilaku sistem terhadap waktu. Sistem dinamika dapat bersifat konservatif atau disipatif. Sistem yang konservatif memiliki energi yang konstan terhadap waktu, sedangkan sistem yang disipatif kehilangan energi terhadap waktu. Salah satu sistem yang konservatif adalah bandul sederhana. Pada bandul sederhana gesekan udara diabaikan sehingga energi potensial dan kinetik sistem konstan untuk setiap waktu. Sebaliknya jika gesekan udara diperhitungkan, ada energi dalam sistem yang terus menerus berkurang terhadap waktu dalam bentuk energi panas atau gesekan maka sistem ini bersifat disipatif (Guckenheimer J& Holmes P 1983).
Sebuah sistem yang perilakunya dimasa depan ( atau dimasa lalu ) dapat diperkirakan bila kondisi awalnya diketahui adalah sistem yang deterministik. Setiap sistem mekanik klasik adalah deterministik. Contohnya pada hukum gerak Newton, jika posisi dan momentum pada suatu waktu dapat ditentukan maka perilaku sistem dapat ditentukan untuk waktu-waktu lainnya. Sedangkan sistem non-deterministik menggunakan konsep probabilitas untuk menggambarkan perilakunya terhadap waktu. Molekul gas dalam termoDinamika, teori kinetik gas, gerak brown, dan kuantum merupakan contoh sistem probabilistik (Guckenheimer J & Holmes P 1983).
4.2.2 Persamaan Differensial Orde Pertama
Sistem persamaan differensial orde pertama interaksi dua persamaan differensial terkopel (Hirsch MW et al 2004) dapat dinyatakan sebagai:
1
2 ( , )
( , ) dx
f x y dt
dy
f x y dt
(26)
f1 dan ,f2 adalah fungsi kontinu bernilai real dari x dan y, dengan laju perubahan x dan y sendiri dan tidak mengandung t di dalamnya. Sistem persamaan differensial disebut sebagai sistem persamaan differensial mandiri (Autonomous).
4.2.3 Titik Kritis (critical point)
Analisis sistem persamaan differensial sistem dua persmaan terkopel sering digunakan untuk menentukan solusi yang tidak berubah terhadap waktu (Hirsch MW et al 2004), yaitu untuk tiap dx/dt 0,dy/dt 0. Titik kritis ( * *
,y
x )
dari sistem dapat diperoleh dengan menentukan
0 / , 0
/dt dy dt
dx (27)
4.2.4 Konstruksi Matrik Jacobi
Dengan melakukan pelinieran pada persamaan interaksi dua persamaan terkopel maka diperoleh matriks Jacobi (Hirsch MW et al 2004) berikut :
2 2 1 2 2 1 1 1 x f x f x f x f
Ji (28)
4.2.5 Vektor Eigen dan Nilai Eigen
Diberikan matrik dengan koefisien konstan J berukuran nn dan SPD homogen berikut:
Jx
x
, x(0)x0 (29)Suatu vektor tak nol x dalam ruang n disebut vektor eigen dari J jika untuk suatu skalar berlaku:
x
Jx
(30)Untuk mencari nilai eigen dari matrik J maka persamaan (30) dapat ditulis kembali sebagai:
0
)
(
J
I
x
(31)Dengan I matrik diagonal satuan. Persamaan (31) mempunyai solusi tak nol jika dan hanya jika
0 )
det( )
( J I J I
p (32)
Persamaan (32) disebut persamaan karakteristik dari matrik Jacobi (Hirsch MW et al 2004).
4.2.6 Orbit Kestabilan
Berdasarkan uraian di atas maka kestabilan titik kritis memiliki tiga kondisi (Hirsch MW et al 2004), yaitu
Stabil, jika :
a. tiap nilai eigen real adalah negatif (i 0 untuk semua i )
b. tiap komponen nilai eigen kompleks adalah lebih kecil atau sama dengan nol, Re(i)0 untuk setiap i.
Tak Stabil, jika :
a. tiap nilai eigen real adalah positif (i 0untuk semua i )
b. tiap komponen nilai eigen kompleks adalah lebih besar dari nol, Re(i)0 untuk semua i.
Saddle, jika :
(a) (b) (c)
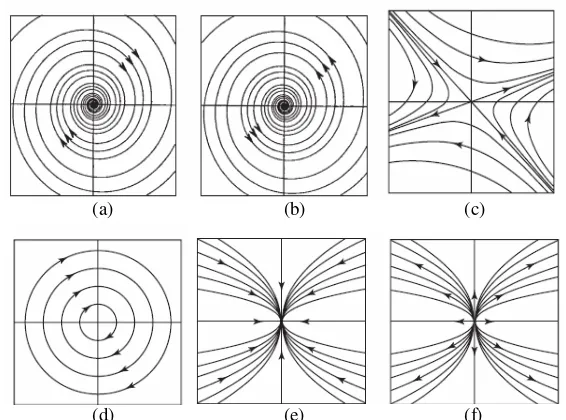
[image:41.612.177.460.96.306.2](d) (e) (f)
Gambar 8. Orbit kestabilan disekitar titik kritis; (a) spiral stabil, (b) spiral tak stabil, (c) titik
saddle, (d) center, (e) titik stabil dan (f) titik tak stabil (Hirsch MW et al 2004)
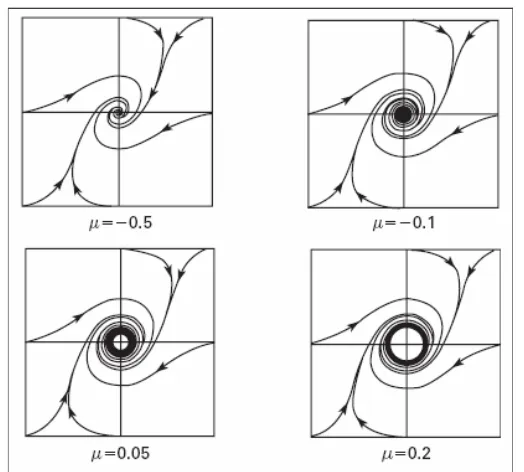
4.2.7 Bifurkasi Hopf
Bifurkasi secara sederhana dapat diartikan sebagai suatu perubahan karakteristik orbit kestabilan disuatu titik kritis yang biasanya ditandai dengan kehadiran suatu limit cycle. Sebagai contoh sederhana terjadinya bifurkasi pada persamaan van der Pol berupa persamaan diferensial pada R2 (Hirsch MW et al 2004).
x dt dx
x x y dt dx
3
(33)
Dengan parameter berada pada interval [-1, 1]. Dengan menggunakan Linierisasi diperoleh nilai eigen berikut :
4
2
1 2
(34)
Gambar 9. Bifurkasi pada persamaan van der Pol ketika parameter divariasikan (Hirsch MW et al 2004)
4.3 Penentuan Titik Kritis Model Fitzhugh-Nagumo
Untuk memperoleh letak titik kritis dapat ditentukan melalui analisis nullcline dari tiap persamaan Fitzhugh-Nagumo ,yaitu sebagai berikut,
v nullcline terjadi pada saat w 0, sehingga diperoleh;
I v v
w 3
3
1 (35)
w nullclline terjadi pada saat v0, sehingga diperoleh;
b a v
w (36)
Dari persamaan (35) dan (36) di peroleh persamaan kubik sebagai berikut,
0 1
3
1 3
I
b a v
v (37)
4.4 Matrik Jacobian Model Fitzhugh-Nagumo
Dengan mensubstitusikan persamaan (25) kedalam persamaan Jacobi (28) diperoleh matriks Jacobi untuk model Fitzhugh-Nagumo sebagai,
b v
J 1 1
2
(38)
4.5 Nilai Eigen dan Syarat Kestabilan Model Fitzhugh-Nagumo
Dari persamaan (32) diperoleh persamaan karakteristik untuk persamaan Fitzhugh-Nagumo : 0 ) ˆ ( ) ˆ 1
( 2 2
2
v b b b v (39)
Sehingga nilai eigen dari persamaan karakteristik tersebut dapat ditulis sebagai 2 ) ˆ ( 4 ) ˆ 1 ( ) ˆ 1
( 2 2 2 2
2 , 1
v b v b b b v (40)
Maka kondisi stabil dari ruang fase akan diperoleh jika memenuhi ketentuan, 0 ˆ 0 ˆ 1 2 2 v b b b v (41)
4.6 Analisis Kestabilan Titik Kritis Model Fitzhugh-Nagumo
besarnya arus eksternal yang diberikan I dan parameter tetap yaitu a = 0.7, b = 0.8,= 0.08. Dengan menggunakan software maple 11 diperoleh hasil
numerik sebagai berikut :
Tabel 1. Analisis numerik kestabilan titik kritis model Fitzhugh-Nagumo
No Variasi Ieks Titik kritis Nilai eigen kestabilan
1 0.00 -1.1994,-0.6243 -0.25130.211900 i Spiral stabil
2 0.32 -0.9769,-0.3461 -0.0091760.2774 i Spiral stabil
3 0.33 -0.9685,-0.3357 -0.0010450.2757 i Limit cycle
4 0.50 -0.1311,-0.8048 +0.14410.191500 i Spiral tak stabil
5 1.25 1.8810, 0.8048 +0.14410.191500 i Spiral tak stabil
6 1.42 2.0857, 0.9685 -0.0010450.2757 i Spiral stabil
7 1.43 0.9769, 2.0961 -0.0091760.2774 i Spiral stabil
8 1.45 0.9933, 2.1166 -0.025320.28020 i Spiral stabil
9 1.50 2.1656, 1.0325 -0.065010.282800i Spiral stabil
10 2.00 1.3341, 2.5426 -0.6412, -0.2026 Stabil node
4.6.1 Kasus Arus Stimulus I = 0
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
(a) (b)
Gambar 10. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 0 ; (a) bidang fase antara v dan w
Dari perhitungan numerik pada tabel 1 diketahui bahwa ketika arus yang eksternal yang diberikan I = 0 maka menghasilkan nilai eigen berupa nilai kompleks dengan bagian real bernilai negatif menunjukkan bahwa titik kritis tersebut bersifat spiral stabil artinya berapapun kondisi awal yang diberikan maka trayektorinya akan menuju titik kritis tersebut membentuk spiral. Namun, jika dilihat pada grafik Dinamikanya terhadap waktu maka pada saat I = 0 tidak tejadi osilasi karena potensial aksi dan potensial recovery langsung menuju kestabilan yaitu pada saat neuron berada pada fase istirahat. Gambar 10 model Fitzhugh-Nagumo menunjukkan suatu kemiripan secara kualitatif dengan gambar 3 pada model Hodgkin-Huxley.
4.6.2 Kasus Arus Stimulus I = 0.33
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
(a) (b)
[image:46.612.126.500.78.347.2]
(c) (d)
Gambar 11. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 0.33 ; (a) bidang fase antara v dan
w bersifat stabil limit cycle , (b) Dinamika v, w terhadap waktu t = 100(c) Dinamika v, w terhadap waktu
t = 100 dan (d) grafik 3D v,w terhadap t
Melalui analisis bifurkasi Hopf (Hirsch MW et al 2004; Medvedev GS, Yoo Y 2007), pada parameter I = 0.33 merupakan parameter kritis terjadinya transisi dari orbit spiral stabil menjadi orbit trayektori“limit cycle“ dan ketika parameter I dinaikan menjadi I = 0.5 mulai terjadi transisi dari stabil“limit cycle“ menjadi spiral tak stabil sebagaimana terlihat dalam tabel 1.
4.6.3 Kasus Arus Stimulus I = 1.25
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
terbentuk trayektori yang bergerak menjauhi titik kritisnya dengan lintasan tertutup sehingga sifat titik kritisnya merupakan spiral tidak stabil.
(a) (b)
Gambar 12. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 1.25 ; (a) bidang fase antara v dan
w bersifat spiral tak stabil (b) Dinamika v, w terhadap waktu t.
Dari gambar 11 (c) dan 12 (b) memperlihatkan fenomena osilasi pada potensial aksi yang frekuensinya bertambah besar seiring dengan bertambah besarnya arus eksternal yang melewati membran, hal ini sejalan dengan hasil yang didapat pada model Hodgkin-Huxley. Melalui analisis bifurkasi Hopf (Hirsch MW et al 2004; Medvedev GS, Yoo Y 2007), pada parameter I = 0.25 merupakan parameter kritis terjadinya transisi dari orbit spiral tak stabil menjadi orbit trayektori spiral stabil menuju keadaan istirahat.
4.6.4 Kasus Arus Stimulus I = 1.43
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
dimana berapapun arus diperbesar tidak akan mempengaruhi potensial aksi yang bergerak menuju kestabilan pada keadaan istirahat.
(a) (b)
Gambar 13. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 1.43 ; (a) bidang fase antara v dan
w bersifat spiral stabil (b) Dinamika v, w terhadap waktu t.
4.6.5 Kasus Arus Stimulus I = 1.45
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
(a) (b)
Gambar 14. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 1.45 (a) bidang fase antara v dan
w bersifat spiral stabil (b) Dinamika v, w terhadap waktu t.
yang bergerak mendekati titik kritisnya dengan lintasan tertutup sehingga sifat titik kritisnya merupakan spiral stabil. Pada gambar 14 (b) mulai memperlihatkan fenomena berkurangnya osilasi akibat adanya ateunasi dimana berapapun arus diperbesar tidak akan mempengaruhi potensial aksi yang bergerak menuju kestabilan pada keadaan istirahat. Pada konsisi ini penambahan arus eksternal hanya akan menambah kecepatan potensial membran menuju stabil pada keadaan istirahat.
4.6.6 Kasus Arus Stimulus I = 2
Melalui simulasi numerik menggunakan software Matlab 7.01, dengan mensubstitusikan nilai parameter ke persamaan (25) dapat diperoleh grafik hubungan antara v dan w serta Dinamika v, w terhadap waktu t.
(a) (b)
Gambar 15. Sistem Dinamika membran model Fitzhugh-Nagumo saat I = 2 ; (a) bidang fase antara v dan w
bersifat stabil node (b) Dinamika v, w terhadap waktu t.
Gambar 15 memperlihatkan suatu kodisi bidang fase bersifat stabil asimtotik. Jika dilihat dari nilai eigennya dari tabel 1. menunjukkan ketika arus eksternal yang diberikan I = 2 maka akan menghasilkan titik kritis yang memiliki nilai eigen real bernilai negatif sehingga terbentuk trayektori yang bergerak mendekati titik kritisnya tanpa osilasi. Keadaan ini memperlihatkan bahwa potensial membran sudah stabil sehingga arus yang diperbesar tidak lagi berpengaruh.
SINKRONISASI CHAOTIK NEURONAL NETWORK
5.1 Gejala Chaos Model Neuron Fitzhugh Nagumo
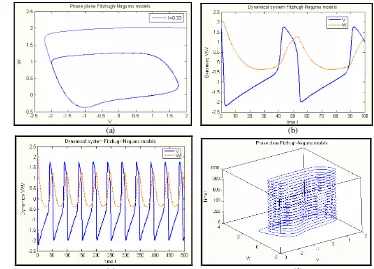
Jika pada pembahasan sebelumnya, yang menjadi parameter adalah arus stimulasi eksternal kontinu dapat menghasilkan bidang fase yang bersifat periodik, maka jika arus stimulasi eksternal berupa arus periodik sinusoidal yang merupakan fungsi waktu akan diperoleh grafik bidang fase yang merupakan gejala chaos, dengan ciri utama memiliki sensitifitas yang yang tinggi terhadap perubahan kondisi awal meski tetap bersifat deterministik, dan munculnya attractor asing (Guckenheimer J & Holmes P 1983).
Persamaan Dinamika model Fitzhugh-Nagumo untuk neuron tunggal tak terkopel disajikan dalam persamaan (25). Parameter yang digunakan dalam simulasi model tersebut antara lain a = 0.7, b = 0.8 dan = 0.08. serta dengan memvariasikan arus stimulan yang diberikan. Telah diperoleh dari tabel 1 titik equilibrium pada saat I = 0 berada pada (v*, w*) = ( -1.1994, -0.6243 ) dengan nilai eigen pada titik ini sebesar (-0.25130.211900 i ). Jadi dengan memvariasikan arus stimulan akan diperoleh kestabilan asimtotik di sekitar titik equilibrium pada I = 0, dan dengan memvariasikan arus stimulan yang diberikan maka akan menghasilkan beberapa sifat bifurkasi seperti pada tabel 1.
Dalam pembahasan ini, arus stimulan eksternal yang diberikan berupa arus periodik sinusoidal dengan fungsi berikut (Mishra D et al 2006),
) cos( ) / ( )
(t A t
I (42)
(a) (b)
(c) (d)
[image:52.612.112.517.77.598.2]
(e) (f)
Gambar 16. Model Fitzhugh-Nagumo saat A = 0.7 ; (a) respon waktu saat = 0.4 (b) bidang fase saat
= 0.4 , (c) respon waktu saat = 0.8145 (d) bidang fase saat = 0.8145 , (e) respon waktu saat
= 0.93, (f) bidang fase saat = 0.93
perilaku dinamikanya dari sifat periodik pada gambar 16.a dan 16.b sampai pada perilaku dinamika yang bersifat chaotik seperti terlihat pada gambar 16 e dan 16 f.
5.2 Sensitivitas Terhadap Kondisi Awal
Telah terlihat pada gambar 16 terjadinya bifurkasi pada bidang fase dari kondisi periodik seperti gambar 16.a menjadi kondisi chaotik seperti terlihat pada gambar 16.e. Pada kondisi chaotik ini memiliki sifat yang khas yaitu amat sensitif terhadap perubahan kondisi awal. Pada gambar berikut diperlihatkan adanya gejala sensitivitas terhadap perubahan kondisi awal dari potensial aksi yang merupakan ciri gejala chaotik (Guckenheimer J & Holmes P 1983).
Gambar 17. Model Fitzhugh-Nagumo saat A = 0.7 dan = 0.945 serta memvariasikan kondisi awal potensial aksi V0
Dalam gambar 17. memperlihatkan adanya perubahan Dinamika potensial aksi ketika kondisi awal potensial aksinya sedikit diubah yaitu V0 = 2.0000,
V0 = 2.0001 dan V0 = 2.0002.
jaringan syaraf berdasarkan teori sinkronisasi yang terjadi antar neuron (Aziz-Alaoui MA 2006).
Sinkronisasi secara umum merupakan karakteristik banyak proses dalam sistem alam dan sains nonlinier serta meruapakan salah satu dasar fenomena nonlinier yang dipelajari dalam matematika, fisika, teknik dan ilmu biologi.
Sinkronisasi berasal dari bahasa Yunani dari kata syn = umum dan chronos = waktu, sehingga dapat diartikan berbagi waktu secara umum atau
terjadi pada waktu yang sama, yang merupakan korelasi atau kesesuaian waktu dari proses yang berbeda (Aziz-Alaoui MA 2006).
Kemudian sinkronisasi dua sistem Dinamika secara umum diartikan suatu sistem dengan cara tertentu terkait dengan gerakan (Dinamika) sistem yang lain.
Osilator Chaotik ditemukan dalam banyak sistem Dinamika. Perilaku sistem dikarakterisasi dengan ketidakstabilan, dan sebagai hasilnya tidak dapat diprediksi (terbatas pada rentang waktu kecil). Secara kasar sistem dikatakan chaotik jika tetap deterministik (tidak acak), jika memiliki karakter takperiodik dalam rentang waktu yang besar, dan jika memiliki sensitifitas yang tinggi terhadap perubahan kondisi awal yang diberikan.
Konsekuensinya, untuk sebuah sistem chaotik, lintasan awal berubah-ubah mendekati satu sama lain berbeda eksponensial terhadap waktu, dan dengan cepat menjadi tak terkait. Hal ini menyebabkan dua sistem chaotik yang identik tak dapat disinkronisasi. Hal ini berarti bahwa mereka tak dapat memproduksi sinyal chaotik identik, terkecuali jika kondisi awal secara eksak dibuat sama, yang secara umum menurut fisika tidaklah mungkin. Sehingga keadaan chaos dipercaya takbisa terkontrol dan kemudian takbisa digunakan (Aziz-Alaoui MA 2006).
kimia, atau untuk mengkode pesan elektronik untuk keamanan komunikasi (Aziz-Alaoui MA 2006).
5.3 Kopling Neuron Model Fitzhugh-Nagumo
Mari kita perhatikan sebuah jaringan dari n buah neuron Fitzhugh-Nagumo. Neuron ini terkopel oleh variabel v. Jaringan ini dapat dimodelkan sebagai sistem :
) ( ) , ( 3 1 3 i i i j i i i i i bw a v dt dw v v h I w v v dt dv (43)
Dimana , i j, i1,...,n, j1,...,n dan arus stimulus ( I ) yang diberikan berupa fungsi sinusoidal bergantung waktu.
Fungsi kopling h diberikan sebagai :
n j j ij s s i ji v v V g c v
v h 1 ) ( ) ( ) ,
( (44)
Dimana kopling sinaptik dimodelkan sebagai fungsi sigmoid (Lange E et al 2005; Belykh I & Shilnikov A 2008) dengan batas :
)) ( exp( 1 1 ) ( s j j v v (45) s
adalah ambang yang dicapai oleh setiap potensial aksi dari sebuah neuron. Neuron-neuron tersebut dibuat identik dan sinapsis terjadi sangat cepat dan saat itu juga. Parameter gs berkaitan dengan kekuatan kopling sinaptik.
Sinapsis tereksitasi, karena itu kenapa potensial pembalik Vs harus lebih besar dari
) ( ij
n c
C adalah matrik koneksi nn: 1
ij
c jika i dan j terkoneksi, i = 1,...,n, j = 1,...,n
0
ij
c jika i dan j tidak terkoneksi
Matrik C ini dapat simetri ataupun tidak, sehingga kopling satu arah atau dua arah dimungkinkan.
Solusi sinkronisasi ada jika dan hanya jika setiap neuron memiliki input yang sama dari neuron yang lain (Lange E et al 2005; Belykh I & Shilnikov A 2008).
5.3.1 Model Dua Neuron Terkopel Fitzhugh-Nagumo
Langkah awal dalam hal ini adalah mengadaptasi metode sebelumnya untuk dua neuron Fitzhugh-Nagumo. Dalam kasus ini, kita gunakan dua arah koneksi karena jumlah input harus sama