PENAPlSAN
B M E R I PROBXOTIK UNTUK
BIQKONTROL,
VtBRlQSXS PADA
U R V A
UDANG
WINDU:
KQNSTRUKSX PENANDA MOLEKULER
DAN ESEI PELEKATAN
SE
KOWH PASCASARJANA
WDANARNI,
Penapisan BaMeri
Probatik untuk Biokontrol Vibriosis pada Larva Udang Windu: Konstruksi PenandaMalekuler
dan Esei Pelekatan. Dibimbing aleh AMTONIUS SUWAMTO, SUKENDA, dan BLSlANA WiDLYATIM Y .
BaMeri prubiatik untuk biokantrol vibriasis pada
larva
udang windu telah diisatasi darilarva
udang windu dan lingkungan perneliharaannya. fjampir sernua isalat yang diperaleh dapat rnengharnbat pertumbuhan V.harveyi
ABSTRACT
WIDANARNI,
Screening
of Probiotic Bacteria for Biocuntrol of Vibriosis in Tiger Shrimp Larvae: Construction of Molecular Marker and Adherence Assay. Under the direction of ANTONlUS SUWANTO, SUKENDA, and BIBIANA WIDIYATI M Y .Probiotic ba&eria far biocuntrol
of
vibriosis were isolated from tiger shrimplarvae
and hatchery environment. Almost all probiotic isolates wuld inhibit the growth ofpathogenic
Vibn'o haweyi. SST-b isolate fromSkeIetonema
was very effective in inhibiting V , harveyi and significantly reduced larvalmortaltty
in pathogen challenge assays. These prospective probiotic bacteria, at concentration 10'-10~ CFUfml, did not show pathogenicity toshrimp larvae.
SKT-b was Gram negative, short rodshape,
exhibited yellow colonies an TCBS and swarming an SWC-agar media, motile, utilizedglucose and
sucrose but not lactose; produced extmcellular proteaseancl
amylase, but did notproduce
&itinasen
Partial sequencing of 16s-rRMA gene showed that SKT-b was similar to Vib* algindyticus.To
analyse mechanism of tiisease suppression by SKT-b,especially
on
their competition foradhesion
sites, we constructed recombinantSURAT
PERNYATAAN
Dengan
ini saya rnenyatakan bahwa disertasi yang berjudui:PENAPISAN BAKTERI PROBlOTlK UNTUK 810KQWTR01, VIBRIOSIS
PADA
LARVA UDANG WIHDU: KONSTRUKSIPENANDA
MOLEKULERDAN
ESEl PEEKATANAdalah benar merupakan hasil karya sendiri
dan
belum
p m a h dipublikasikan oranglain.
Semua surnber
data dan infwmasi yang digunakanielah dinyatakan
secara jalas datr ciapat diperiksa kebnarannya.
PENAPlSAN W
R
I
PROBIOTIK UNTUK
BIOKOMTRUL VlBRlQSIS PADA
U R V A
UDANG WXIYDU:
KUNSTRUKSI PEWNDA MOLEKULER
DAM ESEl PELEKATAN
OLEEI:
WIDANARNI
Disertasi
sebagai salah satu syarat untuk memptofeh gelar
Ooktar pada
Program Studi Biobgi
SEKOUH
PASCASAWANA
INSTITUT PERTANIAN BOGOR
Judul Uisertasi
:
Penapisarr
bakteri probiotik untuk biakuntraivibriosis
pada
larva udang windu: konstruksipenanda rnolekuler dan esei pelelcatan
Narna
:
WidanarniNRP
:P17600007
Program Studi : Biologi
Prof.
Dr.
ir. Antonius Suwanto, MSc
Ketua
_C_C.''
Prof. Dr. Dh. Bibiana
W. Lay. MSc
Anggota
2.
Ketua Program Studi BiaiagiA D e k a n
!
Sekolah
Pascasarjana
Dr.
Ir.
Dedy Duwadi S.,DEA
7
MSc
Penulis dilahirkan di Blitar pada
tanggal
27 September 1967 dari ayah Goded Nistamardan ibu 5ri
Muharti. Pendidikan sarjana di tampuh di Program Studi Budidaya Perairan, Fakuttas Perikanan dan llmu Kelautan (PB, lutus pads tahun 1991. Pada tahun 1992-1993 penulis mengikuti research student diLaboratory of Fish Phydogy,
Depattment
of Fisheries,Facufiy
of Agricuftum/, University of Tokyo, Jepang. Pada tahun 1996, penutisditerima
di Program Studi Bialagipada Program
Pascasarjana 1P8 dan rnenamatkannya pada tahun 1999.Kewmpatan untuk
rnelanjutkan
ke program doktar pada program studi dan pada perguruan tinggi yang sama diperoleh pada tahun ZQOO. Beasiswa pendidikanpascasarjana diperoleh dad
Departemen Pendidikan Nasional.Penulis k k e j a sebagai stat
pengajar
di Departemen Budiiaya Perairan, Fakultas Perikanan dan llmu Kelautan, IPBsejak
tahun 1 994. Pada tahun 1996- 1998 penulis rnenjadi peneliti utama pada Riset Unggulan Terpadu (RUT IV) bidang Teknolugi Hasii Pertanian dan Rise!Dasar pada tahun
7999-2000.P&cy of Vibrio isdates for biocontrol of vibn'osis in tiger shdmp
(Penaeus
monodon)
lawae,
telah diterbitkanpada
jurnal Biotropia (20:11-23) pada tahun 2003 dan (5)Mo/ecu/ar
analysis of pmbiatic bacteria fur bimnfrof of vibn'asis in shrimp larvae, telah disajikan padaMarine
Biotechrtology Conference di Chiba,Puji syukur penulis panjatkan kehadirat Allah SWT, atas rahrnat dan hidayah-Nya penulis dapat menyelesaikan peneliian dan penulisan
disertasi
dengan judul: Psnapisan balrteit probiotik untuk biokontral vibriosispada
larvaurfang
wtndu: konstruksi penanda malekuferdan
emi pelekatan,Ucapan
terima kasih
dan penghargaan yang setinggi-tingginya penulis sarnpaikan kepada Prof.Dr.
Ir. Antonius Swanto, M Scsebagai
gum dan pernbirnbing, atas bimbingan clanarahan
yang diberikan sejak kuliah, penyusunanproposal,
pelaksanaan pmalitian, dan penulisan d i r t a s i ini. Beliau jugs telah memberi kesemgatan cian kepercayaan kepada penufis untuk mernbantu dalarn proyek penetitian rnengenai "frubiotik untuk Udang" yang didanaiofeh
Dl P SEAMEO-BIOTRQP dan Intemasionaf Foundation for Science(IFS),
Sweden,
sejak tahun 2001 hingga 2003. Atas birnbingan yang sangatintensif
dari bliau pulasehingga
bebrapa bagian dari dimrtasi ini dapatdisajikan pada seminar internasbnal
serta
dipublikasikan pada jurnaiinternasional.
Penuiis juga rnenyarnpaikan
t&ma
kasihdan penghargaan
yang setinggi- tingginya kepada: Dr. lr. Sukenda, MSc dan Prof. Dr. Dh. BibianaW.
Lay,
MSc sebagai guru dan pernbimbing,am
birnbingan dan saran yang dibtiiran sejak kuliah, penyusunan proposal, pelaksanaan penelitian, dan penulsan disehsi ini. Kepada Dr.Ir.
b d y Duryadi S.,0€A
sebagai W u a Program Studi Wiologi penulis juga menyampaikantetirna
kasih atas birnbingan dan nasihat yang dikrikan. Ucapan terirna kasih juga penuiis sampaikan kepada Prof, Dr, Takashi Aoki,Or.
lkuo Hirona, Dr. Tsuyushi Ohira, dan Ryasuke Yasawa, Laboratmyof
Ucapan terima kasih yang sebsar-besamya juga penulis sampaikan kepada: Rektar lnstitut Pertanian Bogor dan
Dekan
Sekolah Pascasarjana lP8 atas kesernpatanyang
dibrikanuntuk
rnengikuti pendidikan di Sekolah Pascasarjana IPB;Direktorat
Jenderai Pendidikan Tinggi rnelaiui BPPS yang telah rnernbrikan biaya pndidikan; DIP SEAMEO-BfOTROP dan IFSyang
telahmenanggung
sernua
biaya penelitian; dan Direktur SEAMEO-BIOTROP yang blahrnenyediakan
fasifitas pranelitian pada Labaratorlurn Biologi Malekuler.Kepada
ternan-teman mahasiswa: BuNyurnan,
Bu
Selly, Mbak Ella, Mbak Etty, lrawan, Yogi, Pak Hem, Pak Mardi, Yusuf, Peter, Nana, Cecil, Bu hi, Esti, Wiwit, dan adik-adik mahasiswa S-1 serta teknisi Pak Huwndan
Pak Ran&, penulis rnengucapkan terirna kasih yangsebesar-besamya
atas banbanclan kerjasarnzt
yang baikselarna
pelaksanaan penelitian di Laboratorium BiologiMolekuler,
SHMEG1310"FROP danLaboratoriurn
Kesehatan Ikan, Fakultas Perikanan dan ltmu Kelautan,1PB.
Ucapan terima kasih penutis sampaikan pula kepada Dr.
Ir.
KetutSugama,
MSc dan Dr. !r. I Wayan Teguh Wibawan, MSi sabagai penguji tuar kornisi yang telah rnemkrikan krifik dan saran &mi perbaikan disertasi ini.Terima kasih yang tulus pnulis sampaikan kepada Ayah, Ibu, Nenek,
dan
adik-adik atas daa dan kasih sayang yang dihrikan demi kesuksesan studi penulk. Ucapan terima kasih dan penghargaan yang setinggi-tingginya penulis sarnpaikankepada
suami tercinta Prof, Dr. Ir. MuhammadZairin
Jr, MSc yang dengan setiadan
sangat sabar rnendarnpingi penulisdan
rnernbimbing anak anak kami tercinta Gentiga Muhammad Zairin dan Dwiputra MuhammadZairin.
Kepada
sernua
pihak yang tefah rnembantu dan tidak dapat disebutkan satu prsatu penulis msngucapkan terima kasih atas segata bantuannya, sernoga Affah SWT akan rnernberikan balasan yang setimpal, Amin.Patogenisitas Vibn'u Kandidat Probiutik
...
37Uji fn V h V i r i o Kandidat Prabiutik
...
39...
.
Konstruksi danEsei
Penggunaan
Penanda gfp pada V harvtayi 43...
Konstfuksi Plasmid Pernbawa DsRed untuk Vibrio SKT-b 53 ... Esei Pelekatan V .harveyj
dart Vibrio SKT-b pada Larva Udang 54 Aplikasi Vibrio SKT-b pad# Lawa Udang Melatui Pengkayaan Artemia...
60a
.
Pengkayaan
Artemia dengan Vjbria SKT-b...
60
b
.
Patogenisitas Vibrio S K T b pada Artemia...
61c
.
Pertumbuhan dan Kelangsungan Hidup Larva Udang Windu dengan Pengkayaan Artemia...
62...
...
...
KaraMerisasi dan Identifikasi Vibrio SKT-b.
.
.
.
64KESIMPUIAN DAN SARAN
...
66Kesirnpulan ...
66
Saran
...
67No. Teks
Halaman
.1. Galur baMeri dan plasmidyang
digunakan pad#...
penelitian
ini 172. Sandi dan
asal
isoiat VibriQ kandidat probiotik yangdigunakan dalam penelitian
ini
...
.
.
...
31 3, Patogenisitas V. harveyiR ~ R
dart tipe liarnya...
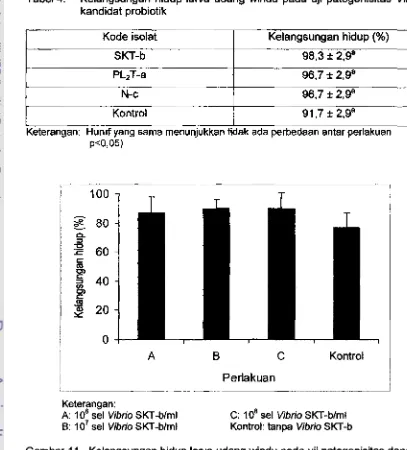
35 4. Uelangsungan hiduplarva
udang windupada
ujipatoganisitas Vibrio kandidat probiotik
...
.. .. . .
38...
5. Patoganisitas V. hatveyi (pWGOl) dan tipe liarnya 47 6. Transposisi pWGU3 dad E. d i S 1 7-3 Apir
ke
V.haw@
dan R. sphaemides2.4.1
...
50'7. Transposisi pLOFKmgfp
dari
E.
cofi ShRIQ,k pirke
V. harveyidan SKT-b
...
51 8. Perturnbuhan bumdan prnnjartglarva udang
windupada
pedakuan kontral dan Vibrio SKT-bselarna
dust...
minggu pemeliharaan, 62
9.
Jurnlah
Vibrio SKT-b di air mediapemeliharaan
clan
No.
TeksHalaman
1
.
Diagram seleksi bakteri grabiotik untuk perneliharaanlarva
hewanakuatik (Garnez-Gil
dan Raque, 1998)...
142. Konstntksi
plasmid
parnbawa gfp (pWGO1) untuk ekspresi ekstrakromosorn pada V. h a a ~ y i...
.
.
...
213. Konstruksi plasmid perantara
pembawer
gfp (pWG02)...
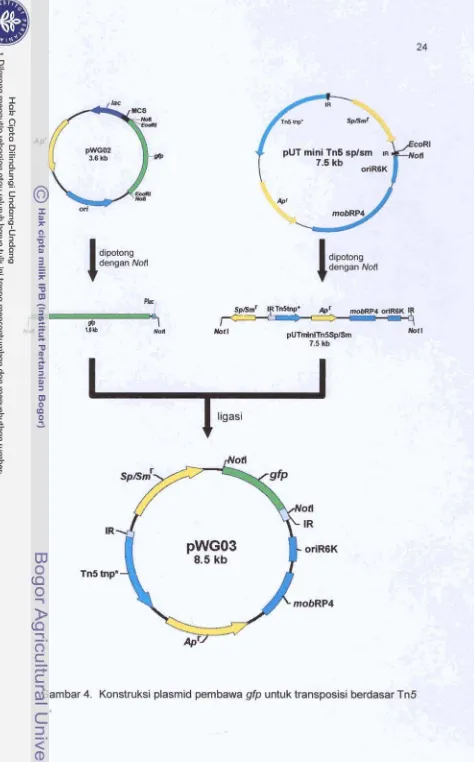
234. Konstruksi ptasrnid pernbawa gfp untuk transposisi berdasar Tn5
...
245. Konstruksi plasmid pembawa &Red (pM0-3)
...
266. PenampiIan Vibna kandidat probiotik pads media TCBS-agar (A) dan SWC-agar
(8)
...
..,..,.
...
327. Profil DNA
genorn
V. hanreyi G3,G?,
dan MR5339
berdasarkan hasil pmotangan dengan endm restriksi Notf. M adalah DNA genom Rhodobacter sphaeroides 2.4.1 yang dipotong dengan enzim restriksi Aset sebagai marker...
32
8.
PenampiIan V.
harveyi
pada media TCBS-agaryang
diamati pada kandisitwang
(A) dan gefap (B)...
339. Perbandingan perturnbuhan V.
fiarveyi
@
dan t i p liarnya (wt)...
3410. Pewhambatan
. .
V. haarvyyi MR5339RP
oleh Vfbriakandidat
probrotrk pada uji in vitro...
36l I. Ketangsungan hidup
larva
udang windu pada uji patogenisitas...
dengan
berbgai tingkat konsentrad Vibrio SKT-b 38 12. Ketangsungan hidup larva udang windu pada uji h vivo Vibrio...
...
kandidat probiotik , 39 13. Jurnlahsel
Vib* sp. dan V. harveyi MR5339RP
pada
air mediapernefiharaan
larvaudang
...
,....
4114,
Jurntah
sel Vi6rio sp. dan V. harveyiM R
5339 W s e ~ t aakurnutasi
larvamati
pada uji in vivo... ...
...
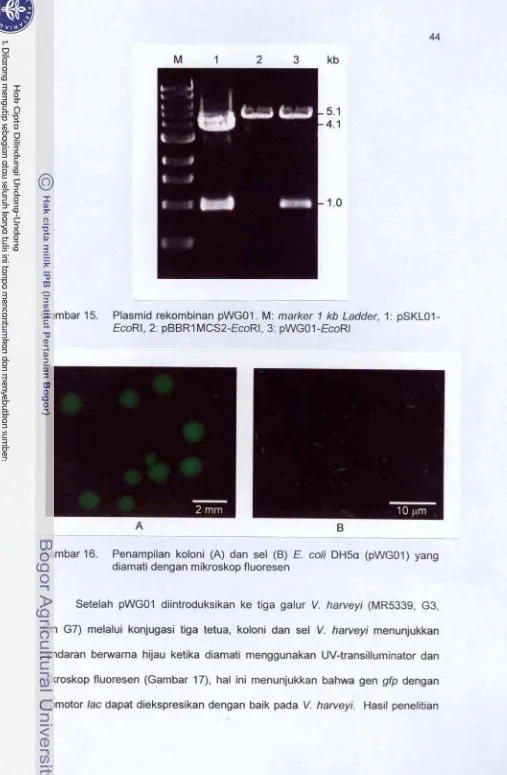
42l 5 . Plasmid rekornbinan pWGO'I. M: marker l kb Ladder, 1 : pSKLU1 -EmRf
.
2: pBBR1 MCS2-EcoR t.
3: pWGO t -EmRI...
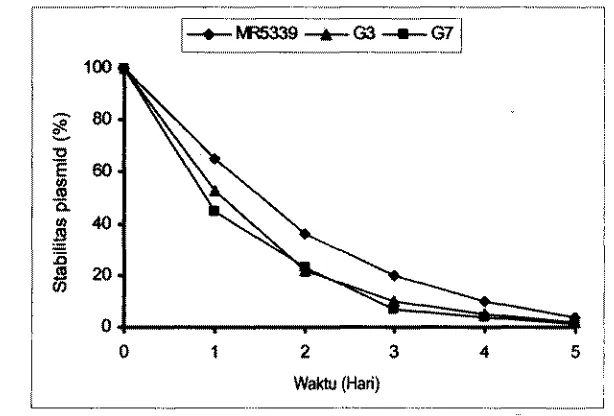
4417. PenampiIan set V. haweyi (pWGO1)
yang
diamati dengan mikroskapfluwesen
...
45 18. Perbandingan perturnbuhan V. harveyi (pWGO?)dan
tipe liarnya...
4619. Stabititas plasmid pWGO4 pada V. harveyi MR5339, G3,
clan
G7
...
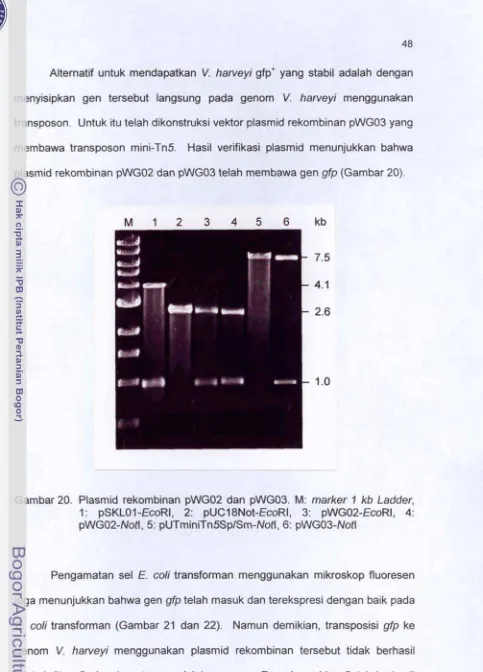
4720. Plasmid rekarnbinan pWG02 dan pWG03. An:
marker
1 kb Ladder,I: gSKL0.t -EcoRI, 2: pUC'l8Rl~i-€coRl, 3: ~ W G U ~ - E C O R ~ ,
4: pWGO2-Notl, 5: pUTminiTnSSplSm-NO& 6: pWE03-#dl
...
48 21. Penampilankulani (A)
dansel(8)
E coli DHSa (pWGO2)yang
...
diarnati dengan rnikroskop
ftuaresen
4922. Penampitan koloni (A) dan el (8) E.
cdi
S17--11 pir (pWG03)yang diamati dengan rnikraskop fluaresen
...~...
49 23.Penarnpilan
kolafii (A) dansel(0)
V. harveyi G3-Tn 1C)gfp yang...
diamati
dengan
rnikroskup fluoresen 51...
24.
Konstruksi
gfp
pada pLOFUmgfp (Stretton et a/.., 1998) 53 25. Prafil DNA genorn bebrapa mutan V.harveyf
G3
(M: markerR. sphaemides 2.4.1 yang dipatong
den
W
an
endm restti
ksi Asef ,1 dan 8: G3 tipe liar, 2: G%Tnl@fp (Km dan mengekspresikan
...-,... ...
...
gfp), 37: mutan
G3
(KrnR)
,.
.
5226.
Kelangsungan hidup larva udang windupada
uji patogenisitas...
V. harveyiG3 tipe
liar dan G3-TnfOgfp
52...
27. Stabifitas gfp pada V. harveyi G3-Tn f Ugf pdan
G3 f pWGO1)53
28, Penarnpilan sei E, &/ DHficx (pWRO1) yang diamati dengan...
rnikroskop
fluoresen 5429.
Usus l a mudang
wincju pada bebrapaperlakuan-
inokutasiV. harveyiG3-TnfqXfp
...
5530.
Usus larvaudang
w i d u yang dikolonisasi dengan V. harveyi...
G3-Tn f Qgfp 56
31. Poputasi V. harveyi G3
RP
di air mediaperneliharaan,
larva hidup,dan
larva mati pada perlakuan denganpenambahan
SKT-b
~ f f f
dantanpa
SKT-bR ~ R
...
57
32. Poputasi V. Aarvayj G3
RP
di air media pernelihara-dan,larva
hidup, dan larva mati pada prlakuan dengan penarnbahan33.
Jumfah
Vibrio SKT-b pada Artemia selarna 6 jammasa
pengkayaan ... 60
34. Kelangsungan
hidup
Arfemia pada uji patogenisitasVibrio SKT-b
...
6135. Kalangsungan hidup fawa udang windu pada perlakuan
bntrol
PENDAHULUAN
Udang windu (Penaeus monadon),
rnerupakan
salah satu kornoditas ekspor unggulan Rasil perikanan. Pemerintah, rnelaluiProgram
Peningkatan10.19
milyar dari
perikanan, Darijurntah
tersebut sebagian besar digantungkan kepada budidaya udang yang diharapkan rnarnpu menyumbangkan devisa sebanyak US $6.79 rnifyar (Basoeki2000).
penunman
k u a l h lingkungan budidaya masih rnerupakan icendala utama untuk rnencapai tujuan tersebut. %ranganpenyakit
bairterial pada tingkatpemknihan
yang
paling&us
dan sering rnenyebabiran tetjadinya kematian massal pada larvaltdang windu adalah serangan b a k t d bwpmiar yang diideiWikasi -ai V i b h harveyi (Lavilla-Pitogo et a/. 1 1990; Karunasagar ef a/. I 994; Ruang pan et a!. 1 998). Baktwi ini p d a umurnnya
menyerang larva
udang pada sMia zma,m p i s
danawal pascalanra (Lavilla-Pitogo
ei
a!. 1990) sehingga rnerupakankendala
dalarn pnyediaan bmih udangyew
sehatdahm
jurnlahb a r
yang diperlukan untuittenrs-menerus dengan dosis suboptimat tefah naengakibatican harveyi menjadi resisten (Karunasagar
et
a/. 1994; Tjahjadi et a/. 1994; Teoet
a!,2000).
Penggunaan
vaksin
jugasuli
diterapkan karma galur V. hwveyj yangmenyerang
larva uclang sangat kmriasi (Suwanto
ef
at/. 41998). *lain iki, degradasi vaksinDengan adanya kehmahan-itelmahan dad brbagai upaya
yang telah
dilakukan, pmggunaan bkteri prubiik sebagai agen biontrctl pada pembenihn udang menawatltan atternatifp r n m h a n
untukmenanggulangi
pernasalahan
tersebut.Dasar
penclekatan ini adalah dengan rnenggunakan aldivitas rnikroorganisrneyang
dapat rnenekan ataurnengtrarnbat
prturnbuhan V , haw8yi tanpamenirnbulkan
darnpak bumk terhdap sistem keseimbangttn ekolqis rnikrob. Cam initefah
terbuMi brhasil danbanyak
cjigunzrlran pada usahahewan
tsrnak (Fuller 1992; Ohhim et al. 11996), narnun k r u akhir-akhir ini ditelitiuntuk
ciiapliicasikan pada sistern hdidayia perairan, rnisalnya gada bucfiaya ikan (Skjemo dan Vacistein 3999; Gram ef a!. 1999), ktsrang-kerangan (Dwtlbt dan
Langdon ?
9W;
Riuelme et BI. 1997)
dan udang Fjahyadiet
a!. 1 994; Rengpipatef
a/. I. 9gSa, f W b , 2000; GOMBZ-GiIet
a[. ZQOQ),M u r u t
Blmrnberg et a/. (19931, tahappelakatan
bairteri patogen pada tubuh inang mtupahn prasyarsltyang
akan rnenerttukan k-asilan bakteri tersebut dalarn mlakukan kolonisasi dan rnensek~sikanfaktor-faktor virulensi.
Virulensi V. harveyi m i t a ndengan
protein
ekstraseluk dan turninesensiyang
diatur
secara krsarna
metalui mkanisrne quwum sensinginterselubr
(Manefield%t
a/. 2800). Adanya kmpetisi tempat plekafanantara
V. haw8yi &wan baM&problotiic pads tubuh udang
diharapkan
dapat m e w a h tercapainya quonrm sen-sehlgus
s e b Faldar-faker virubnsi.Dewan
dmikian, bawd yangrnarnpu
rnenghambatpelekatan,
kolanisasi clan
prkurnbuhan
V. harvsyi swta tidakbemifat patqen terhadap larva udang,
patensial
digunakan set3agai biokontrol untuk melawan Vibrio patogen tersebut.Karena tam laclang bbas V i m tidak &media (Warneed t
993;
Widanarni dan Suwanto20001,
makauntuk
mehkukan esei pelekatanV.
harveyi
p d alawa
3
adalah
gen gfp (green ffmsc3ent protein) (Manning 1997; Janssan 2003). Gen tersebut jika terekspresi akan menghasilkan protein GFP yang h w n d a r hijau jika diarnati dengan cahaya W (Tsien 1998), sehinggapengarmatan ekspresinya
relatif mudah. Selain itu, -ai pnandarnoMukr
geng@
rnerniliki bebrapa keuntungan, antara lain tidak rnernbutuhkan kofaktar atau penamkhan substrat untuk visualisasinya,sensitif,
stabil, firfak toksik, setttn tidak mengganggu fungsi dan peftumtwhansel
(Jmnhans eta!.
1998; Ling et a/. 2000). Penanda resisten terhadap antibiotikrifampisin juga
dapat digunakan untuk V. harveyikafena
padaurnurnnya Vibnio t m & &
sensitif
terhaclap antibiotik rCfarnpisin frjahyadi etal.
1994). manmama mabkufertersebut,
V B h uji dapatdibedakan dad
V i mlain
yaw
sebeiurnnyatelah
terclapat pada larva udang atau air media pemeliharaannya mhingga eseipelekatan
d a m dilakukan.Tufuan
Pemlitian ini bertujuan untuk;:
1.
Mernpfajari
kemampuangenghambatan
Vibri0 kandidat probiotik tertradap pertumbuhan V. harveyipatogen
pada larva udang.2.
Memplajari pelekaian
dan patogenisitas V. harveyipada
larva
udang menggunakan gensnda maleiruler sebagaigen
plapur.3. Mempelajari
penggunaan
pnanda molekuler dalam essipefekatan
unkrk penapisan baMeri prabioti k.Kegunaan Penelaan
Pemlitian ini diharapkan dapat digunakan sebagai suatu metode unkrk menguji patogenisitas V. harveyi pada larva udang windu
dan
juga sebagai suatu metodeuntuk menapis
bakteri probiotik yang patensial rnarnpu rnenghambat Vibriopatogen.
Secar-d
keseluruhan, hasii pnetitian ini diharapkan dapat rnernhrikan infomasi penting daiamusaha
pnanggulangan penyakit vibriosisWbrfo hanreyi dan Penyakit Udang Berpandar
V.
harveyt
adalah
bakteri yang hidup di lingkungan perairan laut,dan
termasuk bakteri yang dapal berpendadar (Baurnann et al. 1994). Beberapa nama lain dari bakteri ini adalah Beneckea haweyidan
Lrrcibactefium
haweyi, dan kernudian diwbut sebagai Vn hhffeyi atau V. carcireriae(Pedersen
et al, 1 998).Ciri-ciri
morfologl
dan ffsiologi V. harveyjpada
medium NrPtrien Agar dengan NaCl 1,596dan
seawater cempIet8-agar fSWC-agar) antara lain: benkrk kolonibulat
dengan etevasi cembung, krwarna krem, clandiarnsternya
2-3 mm setelah inkubsi 24jam
pada suhu 28°C. Jika diamati di ruang gelap, V. hhweyitarnpak
berpendar dan pendarannya ,dapat bedahan hingga2-3
hari. Bakteritersebut
termasuk Gram negati,sd
tunggal b h n t u k M a n g pendek, bemifatmat il,
aksidase
pasitif,sensitif
tehaclap uji vi briostatik 011 29 (2,4-diamina-67, diisopmpylpfen'dinephosphate), tidak r n m b n t u k gas dari fermentasi
terhadap
0-glukasa,tidak
marnpraduksi H2S, tumbuh pada media dfangan penamkhan 1 hingga6% NaCCf,
dan mernpunyai flageta pada salah satu kutub selnya (Baumann et a!. Ig94; Lavilla-Pitoga ef a/, 1.990; Suwarrto et a!. 1998).Pada medium sefektiif untuk
genus
Vibrio, y&u TCBS-agar {ThiosuIfafe Carate Bile-Safi Sucrose), kolani V.harveyi
be~warna hijaudan
hrpendar jika diamati pada ruang getap (Lavilla-Pitogoef
a!. 1990), Kemarnpuan berpendar merupakan hasif aktivitasenzim
luciferase
yang dapat brfungsi sebagai katalisator dalam proses oksidasi reduksi. Proses oksidasi rnefibatkan flavinrnononukhticla
dan a k h i d alifatik rantai panjangsebagai
sutsstratnya.perpendaran ini disandikan datam suatu aperun yang disebut dengan
owran
lux (Meighen 1991; Ruby 1996).V. haweyi pada urnumnya bersifat patagen apartunistik, yaitu organisme yang
dafam
keadaannormal
adadafam
lingkungan
pemeliharaandan
bekernbang dari sifatsaprofitik
menjadi patogenik apabila kondisi lingkungan dan inang rnemburuk. Pada saat tejadinya wabah, populasi bakteri ini dapat rneningkat menjadiribuan
kali dalarn wadahlbak pemefiharaan larva,dan ha1
ini tejadi setelah usaha budidaya udang windu Writembangsemra
intensif (Lavilla- Pitoga ef a/. 1990).Menurut Saulnkr
et
a!. (2000) beberapa gatur V. harveyi rnenrpakan patogen wbnamya dan penyebabutama
penyakit vibrbsis pada udang windu. Kesimpulan tersebut diambil tserdasarican studi virulensi dari beberapagalur
V. harveyi yang diisalasidari
larva sekarat (man'bund)dan
diinfeksikan kmbafi pada udangsahat.
Beberapa dari gafur tersebut dapatmanyebabkan kematian total larva udang dengan dosis yang sangat rendah (102
CFU/ml).
Di antara spies-spsies bakteri k p n d a r yang diisolasi dari areal
pertarnbakan
udang, panti-panti prnbenihan, dan pada tubuhudang yang
sekarat, V, harvayi adalah spesies yang paling
=ring
dapat dlsolasi daripadaspesbs Vr'bn'o fainnya (Ruangpan 1998; Tendencia dan da la Pena 2001).
Bakteri ini juga dapat diigolasi dari air, kotoran dan sitsoskeleton induk udang, air penetasan
gaitan
alarni, Arfemia,serta
dariusus
udangsehat
(Lavilla-Pitogo et a/. 1982). Bebrapa pnelitirnenyatakan
batrwa penyakit vibriosis terjadi karena adanya infeksi sekuder olehpatogen
oportunistik. Sedangkan penyebatr utamanya adalah infeksi oieh patogenlain,
defisiensinutrisi,
kondisi lingitungan, dan stress (Lavilla-Piagoet
a!. 1992,Lihtner eta/+
1992).6
Tengah, Jawa
Barat,
Sumatera Utara, dan Sulawesi Selatan (Tjahjadiet
a!. 1994; Prayitno dan Latchford 1995; Suwanto et a!. 1998). Dinegara
lainwabah
penyakifin!
juga terjadi di Thailand (Pasharawipaset
a!.
1998), di Philipina (Lavilla-Pifogo at a/. 1990), dan di India (Karunasagar et a!. 1994). W a h h penyakit ini padaumumnya rnernuncak
pada rnusirn penghujan(Lavilla-Pitago
et
at. t990).Sejumlah pnelitian rnenunjukkan bahwa pnggunaan anti biotik untuk pernberantasan penyakit vibriasis
telah
menysbabkan resistensi bsakteri Vibriaterhadap
bebrapajsnis
antibiotik (Tjahjadief
al1994;
Kanrnasagar et a!. 1994; Teo et a!. 2000; Tendsnda dan de fa Pena 2UU1).Penggunaan
anfibitik juga iturang efektif itarena V.harveyi
dapat
rnernbentuk biafrfm yang dapat rnelindungi sel-sel bakteri ifadbahan
aktifantibiotik
dan
desinfektan sehingga tetap hidup{Karunasagar
et a!. 1996).Penggunaan
vaksin dan irnunostimulanuntuk
rnengelrclalikan penyakit vibriosis
ternyata
dapat mengurangi tingkat kmatian ikan dan udang (Vadstein 1997; Devaraja ef al. 1998; Alabief
a/. 1999). Akan tetapi tidak sernua s p i e s Vibrio penyekbpenyakit
vibriasis dapat dikermdalikan dengan rnenggunakan vaksin, oleh karma sering kali galur-galur Vibrio dalam satu spesies tidak hornogsn. Hasil penelitianSwanto
eta!.
(.1998)
rnenunjukkan k h w a galur V. harvayi berpendaryang
wnyerang farvaudang
sangat bewafiasi pada musirn dan takasiyang
k W a , sehinggasuli
memtukan galuryang
tepat sebagaibahan
vaksin.Petekatan clan
Patownisitas V,frameyi
Menurut Bloembrg
tat
d. ((1993), tahap pekkatan bakteri patogen pada inang merupakan pcasprat yangakan
menentuhn
kekrbsilan baM& tersebut dalam mebkukan $colonis& dan rnmsekresikanfairtor-faMw
virubnsi. Virulensi1999), sehingga penggunaan senyawa yang brsifat
antaganis
terhadapsignal
i n t d u l e r tersebut potwsial untuk mqontrol %Pangan V. harveyi pada larvaudang
(Manefield et a/. 2000).Quorum
sensing pada V.hafveyi
terjadi dengan rnenggunakan senyawacyf Iromosffnne
lacton
(AHL)tertentu,
ya kni N-(3-fiydoxybut~noif~L-homose##e factone (HBHL) sehgai sinyalnya (Whitebadet
a/. 2001). Bila jumlahsd
kkteri telah mencapai kepadatantertentu
maka HBHL itu akan rnernbentuk kornwks dewan protein pengaktr khusus yang akhirnya krfungsiuntuk
mengaktiin skspmsisejurnlah
gen-gwtwnyancli
emimn;rim untuk luminesensi, s d m kiinase, dan protease ektrasetuler, serta MQT-faktw patagenisitas lainnya f Manefrefdet
a!-2000:
Whitehead et a!. 2001 ). Oengan dernikian,adanya
karnpetisitempat pekkatan
antam
V.harveyi
clan hkteri probiotik padalarva
udang diharapkan dapatrnencegah
tercapainyaq u m
sensing sekaligus sekresi faktur- faktor vintlensi.Menun&
Whitehead et al. (2001), V. haw~yi rnerniliki kbih dati satu quorum sensingdengan
duarnokkul
sinyaf, yakni rnolekuf sinyal Ai-l dan Ab2.Molekut
sinyal
Al-ldigunakan
untuk
komunikasiintm-specks
(dalam satu s p s k s )sedangkan Al-2 untuk komunikasi inter-specks (antar s p s k ) . Lebh lanjut dijelaskan bahwa banyak s p i e s lain seperti V. & d m , V. p a r a h ~ ~ k u s ,
V.
an@i/~~tlm,
V. a l g i ~ i a r s , V. nahgens, d m Phdobacferrum pbsphmumjuga rnsnghasilkan molekul sinyaf dengan aktivitas seperti A!-2. Bahkan hasil analisis lebih lanjut menunjukkan bahwa N-2
yaw
dihasilkan oleh E. cdcofi, SalmmIfa typhimurium, dan V. chdera merniliki homologi yang sangat tinggidengan
Al-2 V. bweyi. ihngan demikian, anfara V.harveyi
dan bkWi kandiia£DNA dengan
kanjugasi,
simbiosis, dan produksi antibiotik dikwrtrol oleh rnekanisme quorum sensing (Bassbr 2003).Lavilla-Pitogo
et
al. (4992) melaporkan bahwa ko#onisasi bakteri tarjadi khusus padaorgan-organ
saluranpencemaan dan jarang
diternukan pada aksaskeletun. Penggunaanteknik
antibodi flouresen menunjukkan bahwa V. vulnificus berada pada hernotimfa dan saluran pencemaanudang
windu (Sungdan
Sang 1996). Hasil penelitian Soto-Rodriguez et a!, (2003) rnenggunakanpnanda 5-(4,6-dicAIomfn'azi17-2-y t)
aminoflu~~esceh
(5-WAF,D-16),
rnenunjukkan bhwa V.harveyi
meqkolonisasi saluran pencemaan larva (zoea dan mysis) udang vaname(Litopenaeus
vannelmefl. V,/,harveyi jugs diketahui memilih aaittivitasenzim
kitinase,
protease, dan lipase (Baurnanrt et a/. 19%). Endm kitinase memungkinkan bagi harveyi untuk menguraikan Kitin dan krkolonisasi di dalarn tubuh larva udang windu.Salah
satu mekanisms yang rnungkin terjadl untukmen-ah
blonisasibaMeri
patogen padainang adalah
dengan rnenarnbahkan bakterilain
yangrnarnpu
krkompetisi dengan bakteri patagen dalam rnendominasi ternpatplekatan
(Verschure faf sf. 2000), teeet
al.
(2000) manambahkan bahwa keWhasiIan baktwi probiotik berkomptisi dengan bakteri patogen ditentukan hrdasakan kernampuannya mengkolonisasiinang
dsngan melakat pada mukosa usus.Dengan
demikhn, VibrEo sp. yangrnampu
mengtrambat pelekatan, kohisasi, dan pefturnbuhan V. harvsyi serta tidak bersifat patogen terhaclap larvaudang,
potensial
cfigunakan sebagai Wukontroluntuk
melawan V i b h patogen tersebut.itu, karaktwhsi
dan
identifiicasi baketi probiotik pmting dilaitukan untuk memkdakan dengan galur lain yangpatensial
bemifat patqsn. Karakterisasi dan identifikasijuga
pnting dilakukan unkrk kontrol kualitasdan
kebutuhan paten(Garnez-Eill
et
al. 2UU0).B a k d Prubiotik wbagaf Agen Biokontrol pada
Larva
UdangMenurut fuller (1 9921, probiotik adalah rnikrab hidup
yang
ditambsahkan kedalam pakan yang dapat rnernberikan pengaruh rnenguntungkan
bagi
hewan hang dengan rnernperbaiki iceseimbangan rnikrob ususnya. Pada h e w n akuatik, =lainsaluran
pencemaan,
air di sekeliling organisme tersebut jugamemegang
peranan penting. Sehingga probiotik untuk hewan akuatikadalatr
age# rnikrob hidupyang
mernhrikan penganth menguntungkan pada inang dengan rnernocfifikasikomunitas
mikrob atau berasosiasidengan
inang, menjamin perkikan dalam penggunaan pakan ataumemperbaiki
nilai nutrisinya, rnernperbaiki respan inang terhadap penyakit, atau memperbaiki kualitaslingkungan arnbangnya
(Verschuere et a!. 2000).Biakantrol menunrt
@mz-Gil
ef
a/. (2000)adabfr penggunaan
rnusuhalarniah untuk rnengumngi kerusakan yang disebabkan
oteh
organismekrbahaya
sampai tingkat y a q &pat di&&rir, atau lebih tepat lagi pengaturn papulasi
penyakit ofeh musuh alamiahnya. L&ih lanjut ditamkhkan bahwa
komunitas
mi&&
di dalarn saluranp m a a n
hewan sampai batas tertentu membri ketahanan atau perlinduwan terhadap penya)rit. Demikian pula pacla populasi alamiah hewan akuatik, rnikrobiota di dalam saluran pencemandapat
menmrminkan fingkungan akuatikternbut.
Narnun demikian, pada perneli haraansistem
saluran p e m a a n
larva. Pascafarva yang dipetihara didalarn
lingkungan yang refati steril di suatu pembenihn tidak dapat tumbuh dengan baik dan rnernperfihatkan daya hidup yang rendah bila terpaparpada
karnpleks populasi rnikrob di petak pndederan atau pembesaran. Pascalarva tersebut akan mudah terserang penyakit M aterkena
stress lingkungan atau terinfeksi oleh bakteri yangpatensial
bersifat patagen.Banyak spsies bakteri tetah digunakan sebagai pfabiatik
rnaupun
biakontraf pada larvaudang,
wlaupun
hasilnyaM u m semuanya
memuaskan.
Tjahjacliet
ab (1994) menunjukkan bahwa papulasi bkteri V. harveyi dilingkungan
pemeliharaan udang dapat &&&an dengan caw mengintroduksikan bakteri tertenkr yang diisolasi dari perairanlaut
disekitar
tambak atau pembenihan udang.Muliani
et #I.(2003)
jugamelaporkan
bahwa isolat B L S 2 yang kemudian diidantifikasi sebagai Pseudo~ftemmonas sp. Edssp-l efe)ctif rnenghambat pertumbuhan V, harveyjMR5339
secarain
witm rnaupun in vivo pada larva udang windukarma
senyawa antirnikrob yang dihasilkannya. Widiyanta etd
(1998) melaporitan bahwa kitteri fotosintetik afioksigenik MW4 dan MW5 hrpotensi untuk dikembangkan wbagai biokontrolpada
tarnbak udang karma mampu rnenekan perturnbuhan V. harveeyi masing-masingsebesar
90
dan 70%.Metranisme
Kerja
dan %leks$ Bakterf PrubiotilrMenurut Vefsctrwre
ef
a/.
(2000),
mekanismekerja
bakteri probiatik dapat dibagi menjadi bebrapa cara, yaitu: (1) produksi senyawa inhibitor; (2) kornpetisi terhadapsenyawa
kirnia atau sumbr energi (nutrisi); (3) kornpetisi terhadap tempat ppelekatan; (4) peningkatan respon imun (kekebalan); ( 5 )perbaikan kualitas air; dan (6) interaksi dengan fitaptankton.
Suatu kelompok mikrob dapat manghasilkan
senyawa
antirnikrob yangrnenghambat
perlumbuhan miitrob lain dan dapat diigunaitanuntuk
pengabataninfeksi rnikrob pada manusia dan
h e w n .
Riquslms etal.
f 1997) melaparkan bahwa Vibrio sp. yang cfiemukan brasosiasi dewan larva kerarg-kerangan menjadiprobatik yang
potensial pada budidaya icerang-kerangankarma rnarnpu
menghambat pertumbuhan V. anguifIamrn yang tefah diketahui bersifat patogen pada larva tersebut. Selanjutnya, pertumbuhan V. anguillamm dapat diharnbatoleh
Pseudomonas fluumscens AH2yang
diturnbuhkan pada media dengankonsenfrasi
besiterbatas.
Pada konsentrasi b s i terbatas, pruduksi stderofordan aCrtwitas antikirteri Pseudomonas fluomsmns AH2 rneningkat, sehingga dapat rnenghambaf prtumbuhan V. sngguifIarum (Gram
ei
at/. 1999).permbenitran udang windu whiqga dapat diperoleh probiotik yang potensiat seperti yang diperobh Riquelme et a]- (1997)
pada bubudiya
kerang-kerangan.Irnunostirnulan adalah suatu senyawa kimia yang dapat rnengaktiian sistern imun dan rneningkatkan resistensi inang terhadap serangan virus, bakteri, fungi, dan parasit. Larva ikan, udang, dan invertebrata lainnya pada umurnnya rnerniliki sistern imun
yang
kurang krkembang dibrandingkan ikan ckwasa. Bebrapa penetitian blah dilakukan dalam ugaya meningkatkan respon imunhewan
akuatik, baik rnenggunakan rnikrob hidup maupun ekstrak selwadstein,
1997; Devaraja et a]., 1998; Afabi et a!., 1999; Skjermodan
Vadstein, 19991,Hasilnya
befum wmuanyamemuaskan,
walaupun bebrapa dari inang resistensinya rneningkat terhadag serangan patogen.Peranan
rnikrobdararn m e r n p h i i t i kualitas
air media budidayatelah
banya k dipetajari. Bakteri niirifikasi (Nitrosomonas
clan
Nitmbacfer), tela h diketahui b e ~ r s l n dalam aksidasiammonia menjadi nitrat, dernikian juga
baMeri fotasinte4ik anoksigenik (Rhadopseud~manasdan
Rhodobactee yang brperan dalam mendegradasi H2S (Madigan cafa!.,
1997). Ammonia dan H2S adalah duasenyawa
yangsangaf
toksik bagi hewan akuatik, termasuk ikan dan udang.Hasil penelitian Widiyanto et BI. (1998) menunjukkan b a h bakeri btosintetik anoksigenik MW4 dan
M W 5 ,
selain rnampu msnekan petturnbuhan V. haweyi, juga mampu menurunkan konmntrasiti&
masing-masingsebesar
60 dan 30°h, sehingga potensial digunakan sshgai biokondisioner di tambakudang.
l ntera ksi bakteri
probiofik-fit~planicton
w r a
tida k langsungdapat
meningkatkan prturnbuhandan
kelangsungan hidup organisme budidaya. Hasil pemlitian Dwillet dan Langdwl(t
994) rnenunjukkan bahwa penambahan bakteri galur CA2 ke dafam kutkrralga dapat
meningkatkan nilai nutrisi dari alga tersebutsenyawa antirnikrab yang diekstrak dari diatom, Skeiefonema wstatum, terbukti dapat mewhambat V. anguiIIarum dan spesies-spesies Vibrio lain yang bemifat
patogen tarhadap
ikan, udang, dan kerang.Menurut
Gamez-Eil (ZUOU), seleksi bakteripfabiotik
biasanya rnentpakan suatu proses empiris yang didasarkan pada sedikit buktiilrniah. Banyak
kegagalan penelltianhkteri
prabiotik yang tejadikarma
ketiru rnemilih rnikraarganisrne. Tahapan seleksi memang sudah ditentukan, tefapi tahapan ini masih p r l u disesuaikan menuntt spesies inangdan
lingkungan, sehingga p r l u untu k memahami rnekanismeaksi
probioti k dan msnsntukan kdteria seleksi kitteri probiotik potensiai. Kriteria umum sefeitsi tenrtarna ditentukanberdasarkan
pertirnkngan keamanan batogis (biosafety), meto& pruduksidan
pengalahan,metode
pemktian
probiotik, s&a lokasi di dalarn tubuh dirnana rnikroorganisrnetersebut diharapkan
a@#.Metade
seleksi bakteri probiotiiruntuk kegiafan prnetiharaan
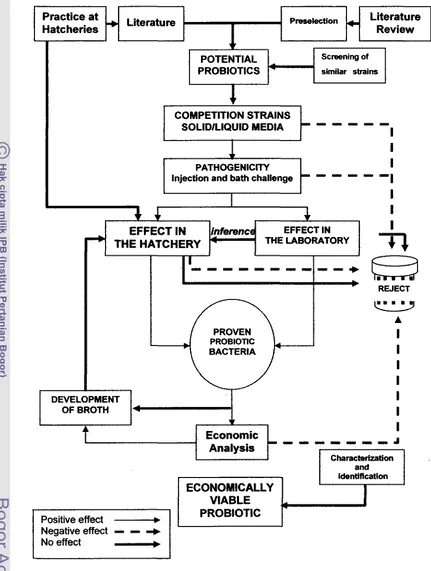
larva hewanPmsekblon Literature Review
screening of
PROBIOTlCS swlar strains
COMPETITIQM STRAINS
SULlDKWUiD MEDlA " "" " " " "
1
1
1
PATHOGENICITY
1
InjeeWn and bath chalienge
- -
lC- -
I
I
I
r
J
1i
EFFECT IN EFFECT IN
M E HATCHERY THE LABQC~ATORY
DEVELOPMENT
/
OFBROTHEconomic
3 1 1 1 1 1 - 1 C JX
VIABLE PRUBiOTIC
Negativeeffect R R
+
[image:30.612.86.517.50.621.2]No effect
Penggunaan Penanda
Mofekukrsebagai
Gen PelaporOalam bialogi rnalekuler dikenat beberapa gen
yang dapat
digunakan sebagai penandamoiekuler
(Manning 1997;Jansson
2003). Gen yang akhir- akhir ini banyak digunakan untuk rnenandai bakteri unfuk rnernpelajari patogenisitas bakteri pada inang adalah gen g f p (Ling et a/. 2000; Sukenda dan Wabbayashi 2001). Gen yang diisolasi dari ubur-ubur (Aequwea victoris) iniapabila
tefekspfesiakan
rnenghasiliran protein yang krpendarhijau
(Tsien, 7998).Pratein
ini menyerap cahaya biwdengan
absorbansi rnaksirnum 395 nm dan memancadcan catraya hijau dengan abeiorbansi maksimurn5U9
nrn
(Manning 1997; Tsien 1998).Protein
GFP
diasumsikan berbentuk silinder yang pada bagian tengahnyaterdapat 15 asam amino,
memknkrk
kol oc-heliks. Tiga dari asam amino iniyakni nomar 65
(serin),
66 (tyrosin) dan 67 (gtysin) mernhntuk kromofor yaitu bagian molekul yang mernancarkancahaya.
Kromofor yangrnengikat
sisi-sisi silinder ini akan mefindungiprotein
tersebut dad enzim-enximsel
inangdan
radikal-radikal bebas (Manning 199'7; Tsien 1998).Sekgai
penanda
mole kufsrgen
gfp rnerniliki bebra pa keuntungan.Keberadaan gen ini di dalam sel tidak kfbahaya karena tidak mengganggu fungsi dan pertumbuhan
wl
wrta proteinnya tidak bemifat tuksiic. Ueuntungan fain dari gen ini adiala htidak
membutuhkan kofaMor atau pena'm bahansubsirat
untuk
visuafisasin ya, sensitif,stabit,
seFta
ekspresin ya dapat dideteksi sarnpai tingkat seltunggal
(Jassnhans et a/. 7998; Ling et a!.ZQQQ).
g%p juga telah berhasil digunakan sebagai
penanda
bakteri asamtalctat
(Lacfohaciffus planfarumdan
L. factis) untuk
mem pelajarikemung
kinan penggunaan baktari tersebut sebagai pernbawa vaksin hidup (GeofFmyet
a/.2000).
Pada penelitian ini,
gen
gfp yang digunakan adalah gfpuv yang dikembangkanoteh
Crarneri et a/. (3996) dan dioptimasi untuk fluotesansirnaksimal
jika disksitasi dengan sinar UV(360400
nm). Fluoresensi E. coii yangmengeicspresikan gfpuv sekiiar 18 kali lebih cerah dibandingkan
dengan
E. ccdiyang
mngekspresikan gfp tipe fiar. Fluaresensi gfpuv inipada
kotuni baker! dapat diarnati dengan rnenggunakan sinar UV,Untuk
memonitor ke-aan bbih dari satu bakbri uji dalarn satu tubuh inangdapat
digunakan
beberapa penanda yang rnemiliki pendamn wama yang brbeda (Soto-Rdriguez et 81.2UU3;
Janssan2003).
Suatttu penanclarnolekuler
yang banr-baru ini juga banyak diplajari adalah gen DsRed.
Gen
tersebutdiisalasi dari kefampok anernan laut
(Discusuma
sp.)dan
jib terrakspresi akanmemanearkan pendaran krwarna mernh (Janswn 2003). Namun pmggunaan DsRed sabagai penanda rnolekufer rnasih rnernertukan optimasi labih
lanjut
karenawaMu
prnatangan
yang diperlukan sangat lama, yakni antam W r a pjam hingga beberapa had (Janssan 2003).
Penanda resisten tefhadap antibiotik rifarnpism juga dapat d'igunakan
untuk
V.hatveyi
karena pa& umumnya Vibnia tersebut sensitif terhadap antibikBAI-IAN DAN
METODE
Bakteri dsn Plasmid
[image:33.611.93.534.189.700.2]BaMeri dan plasmid yang digunakan pada penelitian ini disajikan pada Tabei 1 .
Tabel 1.
Galur
baktcteridan
plasmidyang
digunakan pada penelitian ini Galur bakteri dan Karakteristik yang relevan Sumber/ReferensiTipe liar Tipe liar
UP
RF
g w , ~ m "
gfp',
urn;
gfp*, Krn
G3:: mini-Tn JOKrngfp, ~ m " , gfp'
E* coll
DHSa F', lacZAMl5, hsdR17, m A l , gyrA,
thi-
HB101 Res-, Mod-, recA13, ~ m "
S17-Ihpir Pro", Red,
ad'.
r e d , diIisogenidengan ba k t h f a g e hpir
~ b ' , rep*, MCS p0luescript I I , h"
Cot E l replimn, tm*, ~ r n ~
Ptac dan gfp d i s i s p n pada MCS
pBBRfMCS2, Urn , gfp+
Identik dengan pUC18 dengan dua situs unik Nan mengapit MCS
Vekfor integrasi kromosom hrnrnan
Tn5
pLUfKrn dengan
gfp tanpa promotok diklan di bagian
upstream kan, Km
PIac dan @p ffisisipkan pada MCS
pUClbNot, ApR, gfp'
Plac dan gfp disisipkan p d a MCS pUTmini-Tn5 SplSm, ApR, gfp",
swm"
P I ~ C , A$, DSR&+
Plac dan DsRed disisipkan pads
MCS aB8R-l MCSZ. ~ m " . DsRed*
Koleksi lab (Maros-Sulsel) UuIelcsi lab (EondoCBati) Kubksi b b fGandol-Bali) Penelitian ini
Pmalitian ini Penelitian ini PenaBan ini Penelitian ini Penelitian ini Penelitian ini
Sambrook ef al. (I 989)
Sambrook
ef
al.
(1 989)l-lerrsro et a/. (1990)
Sukenda & Wakabayashi
tzml)
Kovac et a). (1 9%)
D i et Eal. (1 980) Panelitian ini
Peneiitian ini
Panelitian ini
Isolasi M M o Kandidat Prubiotik
V l b h kanrficlat prokotik diisolasi dari berbagai stadia larva udang windu dan media perneliharaannya, termasuk dari
ku&ur
pakan alaminya, di ternpatpernbenihan
udang, Labuhan, Pangandaran, dan Lamgung. Masing-masing sarnpl dimbarpada
mediaseleMi
Thiosulphafe CitmteBi/&a#
Sucrose (TCBS- agar, Qxoid) dan selanjutnya diinkubasi padasuhu
ruang (28-31)OC selama 24 jam. Kalani yangterpisah
kernudian dipilih secara acak untukrnendapatkan
isolat mumi untuk dipehjarl lebih lanjut.Sensitivitas W h o twrhadap AnebWk Rifampisin
Isoht mumi
VibM icandidat probiotik dan V, haweyi patogan diuji sif& sensWitasnya terfradap antibiotik rifampisin dengancara menurnbuhkan
isolat- isolat tersebut m a media seawater mprete (SWC-agar) (5 g bacSapeptone, 1 g yeastextract, 3
ml glycerol, 15 gagar,
750 mt air hut, dan 250 mlaquades)
dengan suplementasi rifarnpisin50
@mi.
Seteiatr diinkuksi pada suhu ruang -lama 24jam, rwpon rwbtensi dikehhui dengan rnengarnati
turnbuhnya
koloni pada media temM.Konstruirsi dan Esei
Penggunaan
Penancla pada V.haweyi
rnengandung rif;arnpisin 50 d r n l (SWC-1-Rfl. Mutan yarag diperoleh selanjutnya diuji patcgenisitasnya pada larva udang. Mutan yang membearihn patogenisitas tertinggi dan sama seprti t i p liarnya kernudian digunakan pada uji in vitro dan
in v h .
Pengujian In Wtro Wbn'o Kandidat Problotik
EEek
pngharnbatan
masing-masing isalat Vibfio kndidat probiotik diujitantang dengan
V. haarvyyi MR5339w.
Setiap isolatVibrio
kandidat probiatik ditumbuhhn pada media SWC-broth dengan kepaclatan 10f5 CFUiml. Pada tabung yang sama dirnbahkanI@
CFU/ml
V. harveyi MR5339RfCZ.
Setelah
diinkubasim a l a m
@aShaker
bergayang dan suhu 2% kufturt
d
disebar padamedia
SWC*Rf sehingga hanya V.harveyj
MR5339 yang turnbuh. ApabilaV. harveyj MR5339
pada tabung
kofitrul
(tanpa isalat probiotik) tumbuh jauh leWh banyak dibanditq pada kuttur camputan (V. hameyi MR5339fiP
dicarnpur dngan M a t prabtik), brarti isolat protriotiktersebut
mampu rnenghambat perturnbutran V. harveyi MR5339fifR.
Patogenisitas
Wbn'o
Kandidat ProbiutikSebetum dilakukan uji tantang dengan V. harveyi pada
larva
udang, hberapa isuiatyang
palingpotensial
berdasarkan uji in vitn, diuji patogenisitasnyapada
lawa wfang. Fengujian dilakukan denganmenamkhkan
Pengujhn In Vwa Wbria Kandkiat Prabiotik
Tiga isalat Vj6tio kandidat probiotik yang paling patensial berdasarkan uji
in
vifro dan tidak bersifat patugen diuji efektivitasnya dalarn menghambatserangan
V. harveyi pada larva udang (Widanam! et a!. ZQQ3a). lsalat Vibn'o kandidat prabiotik dengan kansentrasi 1oe
CFU/rnl dirnasukkan ke dalamwadah
pernsliharaan udang seharisetelah
larva udang dimasukkan. Setelah kokultivasi dengan larvaudang
salarna 6 jam, V. haweyi MR5339R ~ R
dimasukkan dsngan kansentrasiw3
CFUlml. Percobaan dilakukan dengan tiga ulangantemasuk
kantrol (tanpa penamkhanV.
herveyj
maupun Vib#u prabotik). Pengainatan ditakukan selama 5 had, dan pada akhir per&# dihitung kelangsungan Kidup larva udang. Setamapercobaan
larva udang dibri pakan Artemia sebanyak 2-3 individulml dengan kekwensi 4 keli sehari.Konstruksi dan Esei Penggunaan Pananda gfp pada V ,
haweyi
a. K o ~ i r s i gfp untuk E b p m i Elrstmkmmsmt
VeMw dasar dari plasmid yaw dibangun ufrtuk rnembawa gen gfp
adalah
pBBRlMCS2, yaifu pksrnid bspk.b-um inang basdengan
pnancta resistenantihtik kanarnisin, serta memilib
g w
mob agar dapat ditransferke
V. harveyi rneialui konjugasi. Sedangkangm
gfp
yaw
d i m diisolasi dari pSWl dengan pananda antibiotik gentamisin dan prarnatw 1%.Gen
gfp clan promatar lacyang
metode shndar
(Sarnkmk et a!. 1989) dan instruksi sebagairnana disamnkan olah prdusennya. V. harveyirekombinan
(gfp*)
yang dipraleh kernudian diuji perturnbuhan, stabilitas plasmid, dan patagenisitasnya pada larva udang serta dibandingkandengan
t i pliarnya (Wldanarni
et a!. 22003b). Rekombimn yang memiliki stabilitas piasmiddan
patogenisitas tertinggi dan sama sewrti tipe liarnya kernudian digunakan pala esei petekatan.b. Konstruksi gfp untuk Txrtnsposisi
pada
a n o m V.harveyi
Gen gfp clan promatar lac yang diisalasi dari pSKLO? diligasikan dengan pUC18Not yang tdah dilinieritan
dengan
d m EcoRI (Gambar 3). Ptasmicl reicarnbnan (pWGM) yang dipmleh kmudian dipotmgdengan
d m N d sehingga dipmkh fragmen gfp danpromatar
fac yang diapitaM situs
Natl. Fragmsn tersebut kemudian diligasikan dengan pUTminbTn5 SplSm yang telahdilinierkan m a n errzjm Noif (Gambar 4). Uasil ligasi selanjutnya cfitransfomasiiran
ke
E coli Sly-lhpir. Tmnsforman E d i Sf?-4Apir yang rnernbawa plasmid rekombnan fpWG03) wlanjutnya dikanjugasikan dengan V. haweyi (MR5339,G3,
danE7)
dengankkonjugasi
dua tetua(Swanto
1993).Transfer
g@
ke
pnom V. harveyijuga
dilakukan rnenggunakan mini TnlO (pLUFKmgfp)yang
dikmtruksi olsh W o n eta/,
1, (1 998).Konatruksi Piasmid
PembwaD s R d
untuir Wbdo Prubioffk
Untuk merrgamati adanya kornptisi tempat pekkatan antara V. harveyi dan Vibrio prabiotik
secara
langsung pada larvaudang,
dipertukan
penandarnalakuter
yang kfbecfa untuk V.harveyj
dan Vibrio prubiatik. Pada penelitian ini, V.harveyi
dibeti penanda gfp yangbrpendar
hijau, sedangkan untuk Vibrio probatik diberi pnanda DsRscf yang berpendar rnerah.promator lac,
G m
DsR& dan promotor lac diisolasi dafi pOsFied menggunakan enzirn restriksi Pvull d m S p l . Fragmen tersebut selanjutnya diligasikandangan
p88RlMCS2 yang tefah diliniekan dengan enzim restriksi Spl dan EcuRV {Garnbar 5). Plasmid rekombinan hasil konstmksi (pWRU1) selanjutnya diintmduksikan ke ViMo probiotik yang paling potensial rnelalui kanjugasit i a
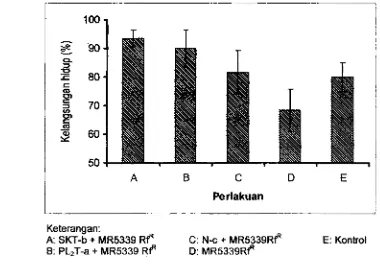
tetua dewan pRK2013 sebagai plasmid penalong (Swanto 1993).Esei Pabkatan V. haweyi
dan
W M o ProbJotik padaLarva
UrtangEsei ini dilakukan untuk mengetahui efeMivitas pengharnbatan ViMo kandidat prabiatik terhadap pslekatan dan kolonisasi V. harveyi pada larva udang. Isofat V i m kandidat probiotik
yang
paling patensiat berdasarkan uji invifro
dan in vivowrta
V.harveyi
~ f f f
dan V. harveyi gfp"yang
memili ki stabilitasdan
patogenisitas
tettitqgi dipifih pada pngujian ini. Konsentrasi Vibrio kandidat probiotik dan V.haweyi
yang digunakan pada prcobaan inimasing-masing
10' CFUlrnl. Pengamatan dilakukan terhadap populasi Vibrio prohotik dan V.harveyi,
baik pada airprnetiharaan rnaupun
pada larva udang. Papulasi Vjhrio pfobiotik dihitung dsngan rnemkrikan penandafiP
seperti pada V. harveyi, Keberadaan V. harveyi gfp4 juga diarnatisecara
langsung padalarva
udang menggunakan rnikroskap fluaresen.Aplibsi Wbrh Probioffk pada Larva Udang Yelaiui Pengkayadln Artemk a. Pengkayaan
Memk
dengan Wbrr'o Probio##kI
diptong denganMI dan !$el
waktu pengksyaan yang
menghasilkan
akumulasi Vibn'o probiotik tertinggiselanjufnya digunakan
@a penelitian ini.b. PatogenbRas WbrSa Probiatik pada Artemk
Uji patogenisitas Vjbrio prabiatik SKT-b pada Artemia dilakukan
dengan
tiga ulangan dan dua perlakuan, yaitu Artemia yang diberi Vibria SKT-bdengan
dosis 10' CfUlrnl dan tanpa SKT-b(kantral).
Ademia dipelihara&lam
botal air mineralyang
diisi dengan 100 rnf air lautsteril dengan
kepadatan Arfemia 100 individulml.Kefangsungan
hidup pada kedua perfakuan dihitungsetiap jam,
sampai jam ke-6 clan setiap 6 jam,rnulai
jam ke12 sampai 24 jam berikutnya.Jumlah Artemia
yang hidup
dihitung dengan menggunakanmetude
sampling vofurnetrik. Airpernetiharaan
Artemia diarnbilsebanyak
100 )11 dan dihitung jurnlah Arfemia yang terctapat didalamnya.
NDai kstangsungan hidup dari kedua pedakuankemudian
dibandingkan untuk rnengetahuipatogenisitas
Vibrio prohotik tersebut.c.
Pertumbuhan
dan Kalangsungan
Hidup h w a Udang Windu denganPen9
lxayaan AHerniaPercabaan
dilakukan dengan dua perlakuan, yaitu
prlakuanprtarna
larva
udang diberi pakan Ademia yang diperkaya dengan Vibrio probiotik SKT-bKeierang
an
:a : Laju
perturnbuhan harian
udang
(%) t :Lama
waHu pemeliharaan udang(hari)
Wt : 8 o M rata-rata akhir udang(mg)
Wo : Bobot rata-rat8 awal udang (mg) Lt :
Panjang
rats-rats akhirudang
(mm)
Lo : Panjang rats-rata
awal
udang(mm)
Sedangkan kelangsungan hidup larva udang
dihitung
dengan rnenggunakan Furnus sebagaiberikut:
SR
: Tingkat kelangsungan hiidup (%)Nt : Jumlah udang yang hidup pada akhir perlakuan (elror) Mo : Jumtah udang yang
hdup
padaawal
pertakuan (&or)
Krrrakbrisad dan Idenmkasi
Wbdo
Prubiotik TerpilihKaraMetisasi fisiolagi
dan
bidimia terfiadap isolat probiotik terpilih ditakukan rnenggunakan analisis fisiabgi MICRUBAC~ ( W e d Science, Australia)dan
idwtifikasinya bedasarkan hasil sekuensing
gen
16s rRNA (WlaMesi et rtl,Wairtu &n Tempat PenelDtian
lsolasi V i b h Kandidat Problo#jk
VibHo kandidat probiotik yang krhasil diisolasi dari larva udang windu dan lingkungan pemetiharaannya di Unit Pembenihan Udang, Proyek Udang Nasianal,
Labuan; Hatchery
Berkah, Pangandaran; dan PT Central Perkiwi Bahari, Larnpung ada 31 isafat (Tabel 2). Dari jumlah tersebut, 26 isolat ktasaf dari telur, larva, pascalarva, dan media pemeliharaan masing-masing stadia udang; 1 isotat brasaI dariair
la& yang dipersiapkanuntuk
kegiatan pembenihan;dan
4 isotat kberasal dari pakan alarni (2 isalat dari Skeletmema dan 2isalat
dari Memia).Semua
kalani yang diperaleh menampilkan wamakuning
pada media TCBS-agar, tidzrk berpendar, dan bersifatmenyebar
pada media SWC-agar (Gambar 6 ) .V. harveyi
yang
digunakan
dalarn genelZtian iniaclalah
V. hharvyi MR5339, G3, danG7
yang secara genetik M aberdasarkan schizotyping Notl (Earnbar7).
V.
harveyj
MR5339 diisolasi dariudang
sakit,rnerupakan
kolsksi dari Batai Penelitian PerikananPantai,
Maros, Sulawesi Selatan dan teiah diuji bersifatpafogen
pada larva udang windu. Sedangkan V. harveyiG3
danG7
masing- masing diisaiasi dari nauplius dan pasca