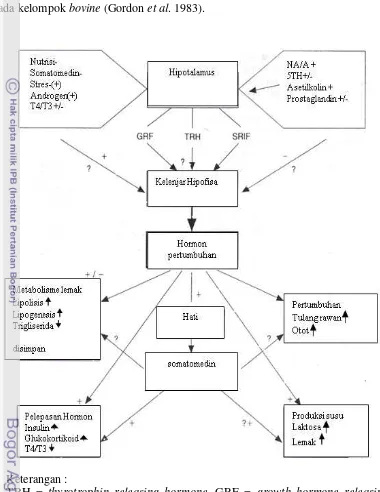
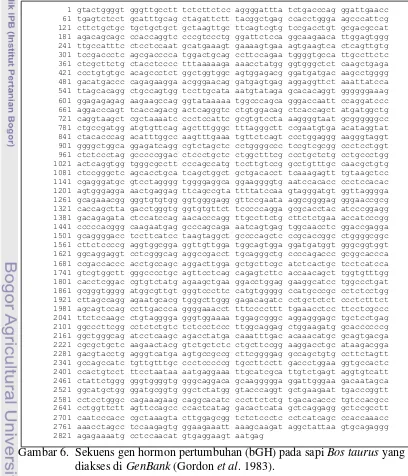
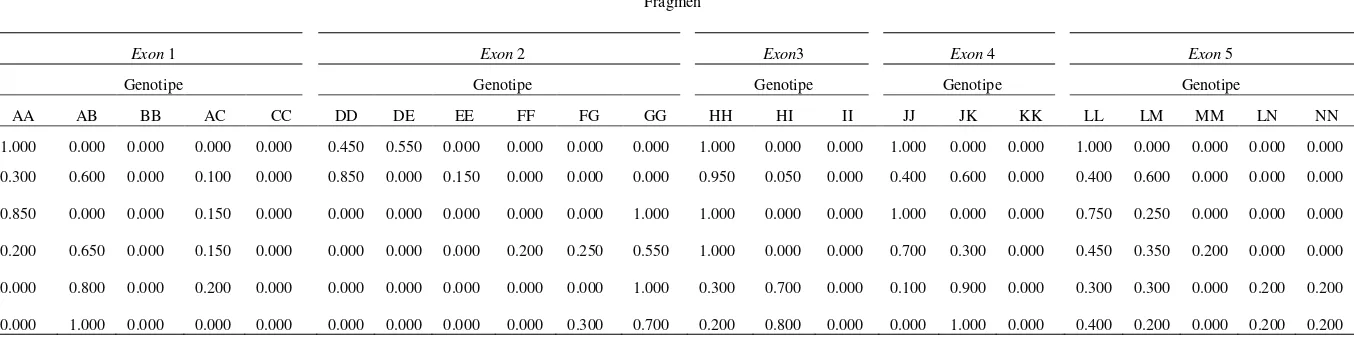
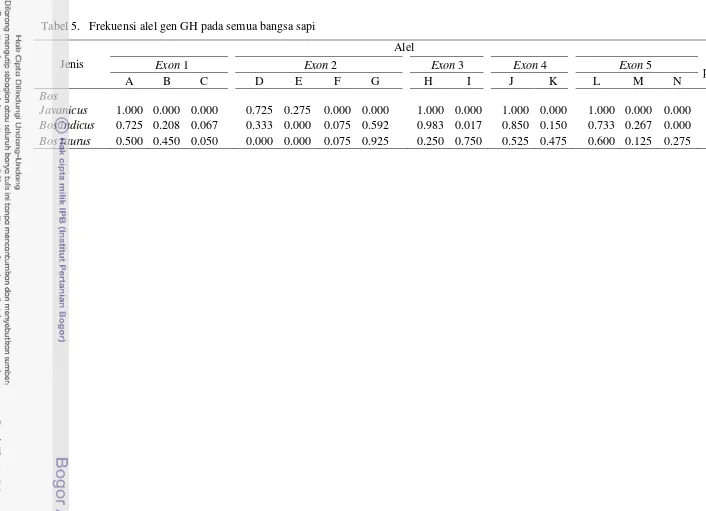
Identification of Growth Hormone Gene Variation in the Exon Region of Indonesian Local Cattle Based on PCR SSCP Method
Bebas
98
0
0
Teks penuh
Gambar


+7

Dokumen terkait
