PERKEMBANGAN OOSIT KARANG LUNAK
Sarcophyton crassocaule
HASIL FRAGMENTASI DI GOSONG PRAMUKA,
KEPULAUAN SERIBU, JAKARTA
VIDIA CHAIRUN NISA
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa Skripsi yang berjudul:
PERKEMBANGAN OOSIT KARANG LUNAK
Sarcophyton
crassocaule
HASIL FRAGMENTASI DI GOSONG PRAMUKA,
KEPULAUAN SERIBU, JAKARTA
adalah benar merupakan hasil karya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir Skripsi ini.
Bogor, Februari 2011
VIDIA CHAIRUN NISA
RINGKASAN
VIDIA CHAIRUN NISA. C54051988. Perkembangan Oosit Karang Lunak
Sarcophyton Crassocaule Hasil Fragmentasi Di Pulau Pramuka, Kepulauan Seribu, Jakarta. Dibawah bimbingan MUJIZAT KAWAROE dan ADI WINARTO
Sarcophyton sp. merupakan salah satu dari beberapa jenis karang lunak yang saat ini menjadi pusat perhatian di bidang famasi . Sarcophyton
crassocaule mengandung bioaktif yang mengandung senyawa terpen yang memi-liki banyak fungsi diantaranya sebagai antimikroba, antikanker, antitumor, dan anti inflammantori. Potensi yang dimiliki oleh Sarcophytoncrassocaule memi-cu adanya kekhawatiran terganggunya keseimbangan alam karena pengambilan untuk skala produksi masal dari Sarcophyton sp. yang dapat menyebabkan keru-sakan ekosistem bahkan kepunahan dari spesies ini, maka dilakukanlah kegiatan fragmentasi karang lunak tersebut untuk memperbanyak jumlah koloni dari ka-rang lunak.
Tujuan dari penelitian ini adalah untuk mengamati perkembangan Oosit
Sarcophytoncrassocaule hasil fragmentasi dan alam, termasuk di dalamnya mengkaji jumlah dan ukuran gamet karang lunak Sarcophyton sp. hasil fragmen-tasi dan non fragmenfragmen-tasi (alam), serta mengkaji pengaruh perbedaan kedalaman lokasi penanaman hasil fragmentasi Sarcophytoncrassocaule terhadap repro-duksi karang lunak Sarcophyton sp.
Pengambilan sampel dilakukan secara acak dengan memotong bagian tubuh pada tiap koloni karang lunak. Total populasi koloni yang diambil nyak 72 koloni, terdiri dari cabang koloni karang lunak hasil fragmentasi seba-nyak 24 koloni untuk kedalaman 3 m, 24 koloni untuk kedalaman 12 m, dan 24 koloni karang lunak non fragmentasi. Koloni karang lunak tersebut digunakan untuk sampel guna mengkaji gamet pada karang lunak non fragmentasi dan hasil fragmentasi kedalaman 3 dan 12 m.
Pengambilan sampel dilakukan saat umur fragmentasi mencapai 8 dan 10 bulan berdasarkan fase bulan Qomariah. Pembuatan preparat histologis dilaku-kan dengan metode parafin dengan cara memotong jaringan karang lunak setebal ±5 µm dan mewarnainya dengan pewarna ganda Hematoksilin Eosin (Kiernan, 1990 dan Gunarso, 1989).
© Hak cipta milik Vidia Chairun Nisa, tahun 2011
Hak cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah dengan tidak merugikan kepentingan yang wajar dari Pencipta
PERKEMBANGAN OOSIT KARANG LUNAK
Sarcophyton crassocaule
HASIL FRAGMENTASI DI GOSONG PRAMUKA,
KEPULAUAN SERIBU, JAKARTA
VIDIA CHAIRUN NISA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Ilmu Kelautan
Departemen Ilmu dan Teknologi Kelautan Pada Fakultas Perikanan dan Ilmu Kelautan
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Judul : PERKEMBANGAN OOSIT KARANG LUNAK
Sarcophyton crassocaule HASIL FRAGMENTASI DI
GOSONG PRAMUKA, KEPULAUAN SERIBU JAKARTA
Nama : VIDIA CHAIRUN NISA
NRP : C54051988
Disetujui,
Pembimbing Utama Pembimbing Anggota
Dr. Ir. Mujizat Kawaroe, M.Si NIP. 19651213 199403 2 002
drh. Adi Winarto, Ph.D NIP. 19580516 198601 1 001
Mengetahui,
Ketua Departemen Ilmu dan Teknologi Kelautan
Prof.Dr. Ir. Setyo Budi Susilo, M. Sc. NIP. 19580909 1983031 003
KATA PENGANTAR
Puji dan syukur penulis panjatkan kepada Allah SWT yang telah melimpahkan rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan dengan baik penelitian dan penulisan skripsi yang berjudul ”Perkembangan
Oosit Karang Lunak Sarcophyton crassocaule Hasil Fragmentasi Di Gosong
Pramuka, Kepulauan Seribu, Jakarta” . Pada kesempatan ini penulis ingin
menyampaikan rasa terima kasih kepada:
1. Kedua orang tua, suami, serta adik atas segala doa dan dukungannya
2. Dr. Ir. Mujizat Kawaroe, M.Si selaku dan drh. Adi Winarto, Ph.D sebagai pembimbing skripsi atas bimbingan, masukan, kritik yang sangat berharga dalam penyusunan skripsi.
3. Prof. Dr. Ir. Dedi Soedharma, DEA selaku dosen penguji yang telah memberikan koreksi dan masukan dalam menyelesaikan skripsi ini.
4. Marsheilla Tjahjadi , sebagai rekan dalam kegiatan transplantasi dan monitoring karang lunak.
Penulis menyadari bahwa penulisan skripsi ini masih jauh dari kata sempurna, untuk itu saran dan kritik tetap penulis harapkan untuk menjadikan tulisan ini lebih baik. Penulis berharap semoga skripsi ini dapat memberikan manfaat bagi pengembangan ilmu pengetahuan maupun sebagai tambahan informasi untuk memperkaya ilmu di kemudian hari.
Bogor, Mei 2011
DAFTAR ISI
2.4 Faktor yang Mempengaruhi Reproduksi Karang Lunak (Octocorallia)... 15
2.4.1 Faktor Internal ... 15
2.4.2 Faktor Eksternal ... 16
2.5 Faktor – faktor Pembatas Pertumbuhan Karang Lunak (Octocorallia) .... 18
III. BAHAN DAN METODE ... 20
3.1 Waktu dan Lokasi Penelitian ... 20
3.3 Metode Kerja ... 21
3.3.1 Pembuatan Preparat Histologis ... 21
3.3.2 Pengamatan Mikroskopis ... 22
3.3.3 Pengukuran Parameter Lingkungan ... 23
3.4 Analisis Data ... 24
3.4.1 Analisis Deskriptif ... 24
3.4.2 Analisis Ragam ... 24
IV. HASIL DAN PEMBAHASAN ... 25
4.1 Kondisi Lingkungan Stasiun Penelitian ... 25
4.2 Pengamatan Mikroskopis terhadap Sarcophyton crassaule ... 28
4.3 Oogenesis dan Ukuran Gamet Betina Karang Lunak Sarcophyton crassocaule ... 29
4.5 Pengaruh Kedalaman Lokasi Fragmentasi pada Reproduksi Seksual
Sarcophyton crassocaule . ... 39
V. KESIMPULAN DAN SARAN ... 45
5.1 Kesimpulan ... 45
5.2 Saran ... 45
DAFTAR PUSTAKA ... 46
DAFTAR TABEL
DAFTAR GAMBAR
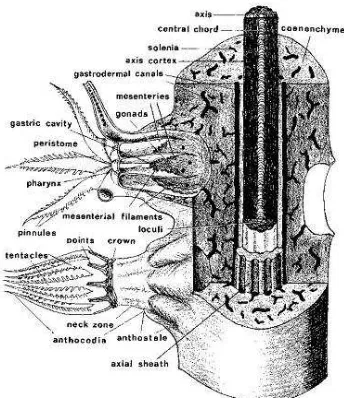
Gambar 1. Penampang vertikal polip karang lunak (Bayer, 1956) ... 5
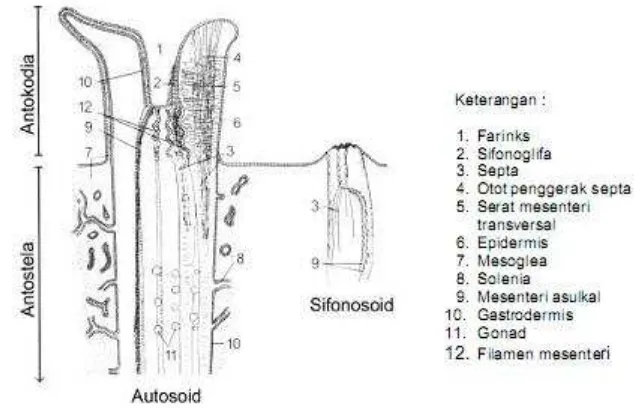
Gambar 2. Penampang vertikal autozooid (Fabricius dan Alderslade, 2001)... 6
Gambar 3. Sarcophyton crassocaule a. keadaan saat polip keluar ... 8
Gambar 4. Gambaran umum dari polip betina ... 10
Gambar 5. Gambaran umum dari polip jantan ... 13
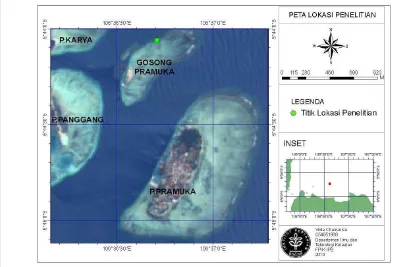
Gambar 6. Peta perairan Pulau Pramuka sebagai lokasi penelitian ... 20
Gambar 7. Oogenesis tahap I karang lunak Sarcophyton crassocaule yang menempel pada saluran mesenteri... 30
Gambar 8. Oogenesis tahap II karang lunak Sarcophyton crassocaule yang menempel pada saluran mesenteri ... 31
Gambar 9. Rata-rata jumlah oosit pada karang lunak non fragmentasi dan fragmentasi umur 8 bulan setelah fragmentasi ... 35
Gambar 10. Komposisi rata-rata jumlah oosit tahap1 dan tahap 2 pada karang lunak non fragmentasi (alam) dan hasil fragmentasi umur 8 bulan setelah fragmentasi ... 36
Gambar 11.Rata-rata jumlah oosit pada karang lunak non fragmentasi & fragmentasi umur 10 bulan setelah fragmentasi ... 37
Gambar 12.Komposisi Rata-rata jumlah O1 dan O 2 pada karang lunak non fragmentasi dan fragmentasi umur 10 bulan setelah fragmentasi .. 38
Gambar 13.Rata-rata jumlah oosit pada karang lunak fragmentasi pada kedalaman 3 meter dan 12 meter pada umur 8 bulan setelah fragmentasi ... 40
Gambar 14.Komposisi Rata-rata jumlah O1 dan O2 pada karang lunak fragmentasi pada kedalaman 3 dan 12 meter 8 bulan setelah fragmentasi ... 41
Gambar 15.Rata-rata jumlah oosit pada karang lunak fragmentasi pada kedalaman 3 meter dan 12 meter pada umur 10 bulan setelah fragmentasi ... 42
DAFTAR LAMPIRAN
Lampiran 1. Data rata-rata jumlah telur pada bulan ke – 8 ... 51 Lampiran 2. Jumlah oosit pada setiap stage dan jumlah total telur pada 4 fase
bulan yang berbeda pada umur 8 dan 10 bulan . ... 59 Lampiran 3. Analisis ragam antara perlakuan non fragmentasi dan fragmentasi
jenis Sarcophyton crassocaule pada umur 8 bulan. ... 60 Lampiran 4. Analisis ragam antara perlakuan non fragmentasi dan fragmentasi
jenis Sarcophyton crassocaule pada umur 10 bulan. ... 61 Lampiran 5. Analisis ragam antara perlakuan produk fragmentasi dengan
kedalaman berbeda jenis Sarcophyton crassocaule pada umur 8 bulan. ... 62 Lampiran 6. Analisis ragam antara perlakuan produk fragmentasi dengan
kedalaman berbeda jenis Sarcophyton crassocaule pada umur 10 bulan. ... 63 Lampiran 7. Uji t-stat perlakuan non fragmentasi dan fragmentasi jenis
Sarcophyton crassocaule pada umur 8 bulan. ... 64 Lampiran 8. Uji t-stat perlakuan non fragmentasi dan fragmentasi jenis
Sarcophyton crassocaule pada umur 10 bulan. ... 66 Lampiran 9. Uji t-stat antara perlakuan produk fragmentasi dengan kedalaman
berbeda jenis Sarcophyton crassocaule pada umur 8 bulan. ... 68 Lampiran 10. Uji t-stat antara perlakuan produk fragmentasi dengan kedalaman
I. PENDAHULUAN
1.1 Latar Belakang
Ekosistem terumbu karang yang terdapat pada perairan tropis dan subtropis
pada umumnya didominasi oleh biota karang batu. Namun selain itu terdapat
pula biota berupa karang lunak atau yang lebih dikenal sebagai Alcyonaria
(Alcyionarian corals), merupakan salah satu jenis coelentrata dan memiliki pera-nan yang tidak kalah penting perapera-nannya dalam pembentukan fisik ekosistem
terumbu karang. Seperti halnya dengan karang batu, karang lunak merupakan
Coelenterata yang berbentuk polip yaitu bentuk seperti bunga yang kecil.
Namun tidak seperti karang batu, tubuh Alcyonaria lunak tetapi disokong oleh
sejumlah besar duri-duri yang kokoh, berukuran kecil dan tersusun sedemikian
rupa sehingga tubuh Alcyonaria lentur dan tidak mudah putus. Duri-duri ini
mengandung karbonat kalsium dan disebut spikula. Secara sepintas lalu
Alcyonaria nampak seperti tumbuhan, karena bentuk koloninya yang
bercabang-cabang seperti pohon dan melekat pada substrat yang keras.
Karang lunak juga memiliki potensi yang sangat besar dalam bidang farmasi
(obat-obatan). Potensial dikarenakan karang lunak (Alcyonaria) mangandung
se-nyawa terpen yang dapat dimanfaatkan sebagai obat-obatan dalam bidang farmasi
sebagai antibiotika , anti fungi hinga senyawa anti tumor.
Kajian tentang senyawa karang lunak yang telah banyak diteliti adalah
kan-dungan kimianya. Senyawa terpen dari beberapa jenis karang lunak. Senyawa
terpen merupakan senyawa kimia yang dihasilkan secara alamiah oleh tumbuh
tumbuhan dan mengandung aroma atau bau yang harum. Senyawa terpen ini
alamiah karena dapat digunakan dalam bidang farmasi sebagai antibiotika, anti
jamur dan senyawa anti tumor. Sedangkan kegunaannya bagi karang lunak itu
sendiri ialah sebagai penangkal terhadap serangan predator, dalam hal
mempere-butkan ruang lingkup, dan dalam proses reproduksi. Senyawa terpen ini pada
karang lunak dihasilkan oleh zooxanthella yaitu alga uniseluler yang bersimbiosis
dengan karang lunak.
Besarnya potensi yang dimiliki oleh karang lunak inilah dikhawatirkan
ter-jadi eksploitasi besar-besaran pada karang lunak langung dari alam tanpa
mem-perhatikan penurunan populasi yang ditimbukan serta kerusakan ekosistem akibat
penurunan popolasi karang lunak tersebut. Karena sampai saat ini belum banyak
usaha pembudidayaan karang lunak untuk produksi masal.
Menjawab tantangan di atas maka pembudidayaan adalah solusi untuk
meng-hindari adanya kerusakan ekosistem. Salah satu pendekatanyang
dilakukan untuk membudidayakan karang lunak adalah dengan metode
fragmentasi dan fragmentasi buatan. Namun pengaruh fragmentasi terhadap
perkembangan reproduksi seksual karang lunak belum diketahui sehingga
dibutuhkan adanya penelitian lebih lanjut mengenai pengaruh fragmentasi tersebut
terhadap reproduksi seksual karang lunak.
1.2 Tujuan
Tujuan dari penelitian ini adalah :
a. Mengamati perkembangan Oosit karang lunak (Sarcophyton crassocaule)
hasil fragmentasi dan non fragmentasi (alam).
c. Mengkaji pengaruh perbedaan kedalaman lokasi penanaman hasil
II. TINJAUAN PUSTAKA
2.1 Sistematika Karang Lunak
Seperti halnya karang batu, karang lunak termasuk filum Coelenterata,
kelas Anthozoa yaitu hewan yang bentuknya seperti bunga, dan disebut polip.
Kelas Anthozoa dibagi dalam dua sub-kelas yaitu subkelas Zoantharia atau
Hexacorallia atau Scleractinia dan sub-kelas Octocorallia atau lebih populer
dengan Alcyonaria. Karang lunak termasuk dalam sub-kelas Alcyonaria.
Sub-kelas Alcyonaria dibagi dalam enam bangsa (ordo) dan salah satu diantaranya
ordo Alcyonacea yang merupakan karang lunak yang sebenarnya. Urut-urutan
klasifikasi karang lunak adalah sebagai berikut :
Filum : Coelenterata
Kelas : Anthozoa
Sub-Kelas : Octocorallia (Alcyonaria)
Ordo : Alcyonacea
Sub ordo : Alcyoniina
Famili ; Alcyoniidae
Genus : Sarcophyton
Species : crassocaule
(Prat, 1903 in Manupputy, 2002)
2.2 Morfologi Karang Lunak
Seperti namanya karang lunak memiliki tubuh dengan struktur yang lunak
terutama karang mati. Walaupun zat penyusun karang lunak dan karang lunak
dan karang keras sama yaitu zat kapur , tubuh karang lunak ini lebih lunak dan
kenyal. Hal ini dikarenakan karang lunak tidak memiliki kerangka kapur yang
keras seperti halnya karang batu.
Gambar 1. Penampang vertikal polip karang lunak (Bayer, 1956)
Bagian atas tangkai disebut kapitulum, bentuknya bervariasi antara lain
seperti jamur, bentuk lobus atau bercabang-cabang. Variasi bentuk inilah yang
menentukan bentuk koloni secara keseluruhan, hal mana sangat membantu dalam
pengenalan jenis di lapangan. Kapitulum mengandung polip sehingga disebut
bagian fertil, sedangkan tangkainya lebih banyak mengandung spikula yaitu
duri-duri kecil dari karbonat kalsium yang berfungsi sebagai penyokong jaringan
tubuh, sehingga disebut bagian steril. Polip dapat dibagi menjadi tiga bagian
Antokodia merupakan bagian yang terdapat di permukaan koloni dan
bersifat retraktil, yaitu dapat ditarik masuk ke dalam jaringan tubuh. Apabila
antokodia ditarik ke dalam, maka yang nampak dari atas adalah pori-pori kecil
seperti bintang. Bangunan luar dari pori-pori inilah yang disebut kaliks.
Gambar 2. Penampang vertikal autozooid (Fabricius dan Alderslade, 2001)
Daerah pada antokodia ditemukan tentakel yang berjumlah delapan dengan
deretan duri-duri di sepanjang sisinya. Duri-duri ini disebut pinnula, fungsinya
untuk membantu mengalirkan air dan zat-zat makanan ke dalam mulut. Selain
tentakel, ditemukan mulut sifonoglifa) yang melanjutkan diri membentuk septa.
Antokodia juga mengandung spikula yang letaknya berderet sampai ke ujung
masing-masing tentakel. Pada pangkal tentakel terdapat mulut yang berbentuk
kepingan yang disebut stomodeum. Lanjutan mulut berupa saluran pendek
disebut farinks atau esofagus. Bagian dalam farinks disusun oleh sel-sel epitel
membantu mengalirkan air ke dalam rongga perut pada proses respirasi. Sel-sel
epitel tadi tersusun sedemikian rupa sehingga bagian dalam farinks berbentuk
alur-alur yang disebut sifonoglifa. Bagian polip dimana sifinoglifa terletak
disebut bagian ventral, sebaliknya yang berseberangan dengannya disebut bagian
dorsal.
Rongga gastrovaskuler atau rongga perut ditemukan pada daerah kaliks,
terusan dari farinks (yang terbagi menjadi delapan dan disebut septa),
benang-benang septa dan organ reproduksi atau gonad. Septa membagi rongga perut
menjadi delapan ruangan. Ujung akhir septa menebal membentuk benang septa
dan menggantung bebas di dalam rongga perut. Dua di antara delapan septa tadi
lebih panjang dan melebar ke bagian basal polip, mengandung banyak flagela dan
fungsinya untuk membantu menyalurkan air dan sisa-sisa makanan ke atas untuk
dibuang ke luar. Sedangkan enam septa lainnya pendek-pendek, mengandung
selsel kelenjar yang fungsinya membantu proses pencernaan makanan.
Masing-masing septa mempunyai otot retraktor yang fungsinya membantu kontraksi
antokodia. Beberapa jenis karang lunak dari marga Sarcophyton dapat menjadi indikator arus di perairan sekelilingnya. Bila arus cukup deras antokodia akan
mencuat keluar untuk mengambil air dan zat-zat makanan ke dalam.
Jenis karang lunak yang akan kami bahas dalam penelitian ini adalah
A
B
Gambar 3. Sarcophyton crassocaule a. keadaan saat polip keluar b. bentuk koloni
2.3 Reproduksi Karang Lunak
2.3.1 Reproduksi Aseksual
Reproduksi aseksual pada habitat alami merupakan mekanisme penting
dalam meningkatkan jumlah individu dalam suatu koloni. Reproduksi ini
dilakukan dengan cara pertumbuhan koloni, fragmentasi, tunas, pembelahan
melintang, dan pencabikan pedal (Sprung dan Delbeek, 1997dalam Setyawan,
2008).
a. Fragmentasi, penempelan fragmen buatan akan berhasil dengan baik bila
kondisi lingkungan pun optimal dan substrat dasarnya pun baik. Karang lunak
yang paling mudah diperbanyak adalah genus dari Sarcophyton, Sinularia, Xenia, dan Anthelia. Fragmentasi dapat juga terjadi karena adanya predator dan
Sarcophyton dapat merusak koloni. Namun, penggunaan fragmentasi mampu menghasilkan sejumlah keturunan dari sisa jaringan.
b. Pembentukan tunas, biasa terjadi pada karang lunak masif seperti
Sarcophyton di bagian dekat dasar tangkai atau pada bagian pinggir kapitulum. Jika pertunasan terjadi pada koloni yang masih kecil, maka anak dan induk akan
tumbuh bersama-sama untuk membentuk koloni bertangkai banyak. Bila koloni
induk yang bertunas sudah berukuran besar maka tunas yang tumbuh akan tetap
kerdil karena terhalang oleh koloni induk.
c. Pembelahan melintang, terjadi pada Xenia spp, dimana pembelahan diawali dengan terpisahnya tangkai mulai dari dasar terus memanjang ke arah vertikal
diantara dua cabang terbesar, hingga akhirnya dapat menghasilkan dua koloni
berukuran sama. Proses ini memakan waktu beberapa bulan untuk sampai
benar-benar terpisah. Namun untuk Xenia spp hanya membutuhkan waktu satu minggu saja.
d. Pencabikan pedal (pedal laceration), koloni benar-benar bergerak melintasi substrat mengikuti jaringan bagian basalnya. Selanjutnya, jaringan ini dapat
terus menempel atau menjadi terlepas dan menjadi individu baru.
2.3.2 Reproduksi Seksual
Seksualitas karang lunak (Alcyonacea) dapat dikelompokkan menjadi dua
yaitu hermaprodit dan gonokhorik (Hwang dan Song, 2007).
a. Hermaprodit, yaitu koloni atau polip karang lunak yang mampu
meng-hasilkan gamet jantan dan betina selama hidupnya. Tipe hermaprodit ditemukan
b. Gonokhorik, merupakan tipe paling umum pada karang lunak. Polip atau
koloni karang lunak gonokhorik hanya menghasilkan gamet jantan atau betina
saja selama hidupnya. Tipe gonokhorik dapat ditemukan pada Anthelia,
Sinularia, Sarcophyton, Lobophytum, Cladiella, Dendronephthya, dan sebagainya.
2.3.3 Gametogenesis
Gametogenesis pada umumnya terjadi pada polip autozooid yang memiliki
alat kelamin atau gonad.
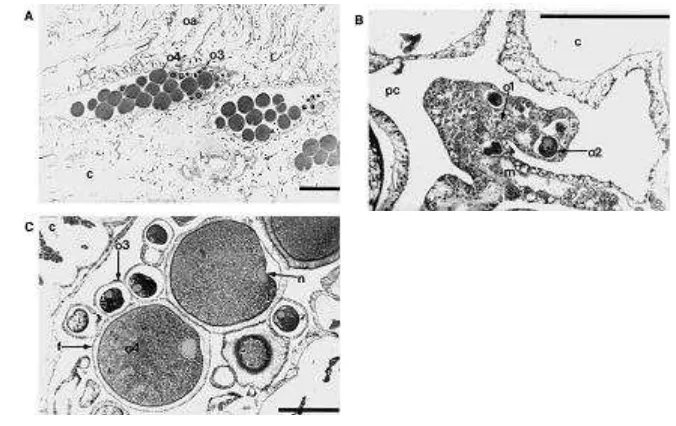
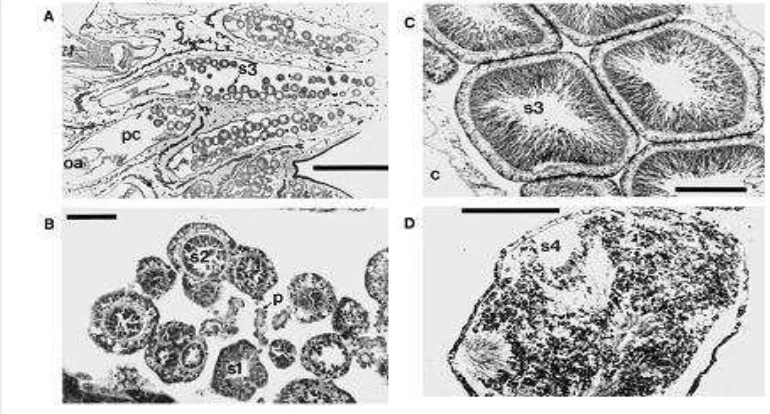
Gambar 4. Gambaran umum dari polip betina (Scale bars ¼ 1000 lm (A) and 250 lm(B and C). (D.G. Fautin et all , 2004 )
Simpson (2008) menjelaskan bahwa secara umum, baik pada polip betina
atau jantan, gamet berkembang di sepanjang non asulkal mesenteri dan seringkali
ditemukan pada bagian dasar polip karang lunak. Dan Gambar 4 memperlihatkan
dari polip betina yang menunjukkan tatanan dan perbedaan ukuran oosit pada
stage III dan IV . Sedangkan pada Gambar B Stage I dan II oosit yang tertanam
di mesenteri . Gambar C , stage III dan IV oosit dalam rongga polip. Huruf c
menunjukkan Coenenchyme, Sedangkan huruf m adalah mesentery filamen; huruf n menunjukkan nucleus ; o1 , adalah stage awal dari dari oosit; o2 , stage II oosit ; o3 , stage III oosit, o4 fase matang oosit IV; oa bagian oral dari polip ; pc rongga polip ; f lapisan folikel sel.
Gamet berasal dari gastrodermis dan akan melekat pada mesenteri dengan
bantuan tangkai pedikel pada awal masa perkembangannya. Selama proses
perkembangan, gamet seringkali dibungkus oleh lapisan folikel yang berasal dari
sel-sel yang terspesialisasi pada gastrodermis. Dengan ukuran yang semakin
meningkat, gamet akan terlepas menuju rongga gastrovaskular atau tetap bertahan
pada mesenteri hingga proses pematangan gamet selesai.
Studi 2-tahun di reproduksi dalam spesies yang paling umum, Sarchophyton glaucum., yang dilakukan di KwaZulu-Natal mengungkapkan gametogenesis dalam koloni jantan membutuhkan waktu 9-10 bulan sedangkan untuk koloni
betina membutuhkan waktu 16-18 bulan (D.G. Fautin et all , 2004) (1) Oogenesis
Proses oogenesis ditandai dengan adanya proses vitellogenik (Simpson,
2008) yang melibatkan pembentukan lemak heterosintetik. Gelembung
bermembran pada ooplasma bersatu membentuk butiran lemak yang akan mengisi
ooplasma. Menurut Hwang dan Song (2007), perkembangan oosit (oogenesis)
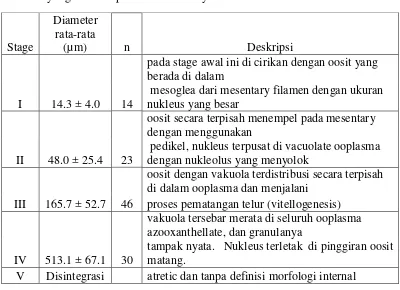
dapat dibedakan menjadi 5 tahap. Tabel 1 adalah pembagian stage oosit telur
penelitian mereka di Kwazulu Natal dengan spesies Sarchophyton glaucum
dengan perbedaan diameter dan ciri-ciri seperti yag di tampilkan pada Tabel 1.
Tabel 1. Ukuran diameter oosit pada setiap tahap perkembangan S. glaucum yang di ambil pada Sodwana Bay
Stage
Diameter rata-rata
(µm) n Deskripsi
I 14.3 ± 4.0 14
pada stage awal ini di cirikan dengan oosit yang berada di dalam
mesoglea dari mesentary filamen dengan ukuran nukleus yang besar
II 48.0 ± 25.4 23
oosit secara terpisah menempel pada mesentary dengan menggunakan
pedikel, nukleus terpusat di vacuolate ooplasma dengan nukleolus yang menyolok
III 165.7 ± 52.7 46
oosit dengan vakuola terdistribusi secara terpisah di dalam ooplasma dan menjalani
proses pematangan telur (vitellogenesis)
IV 513.1 ± 67.1 30
vakuola tersebar merata di seluruh ooplasma azooxanthellate, dan granulanya
tampak nyata. Nukleus terletak di pinggiran oosit matang.
V Disintegrasi atretic dan tanpa definisi morfologi internal
Oosit tahap I ditandai dengan nukleus yang berukuran besar. Oosit
primordial ini melekat pada mesoglea pada mesenteri. Oosit tahap II terlihat
pada rongga gastrovaskular dengan posisi masih melekat pada mesenteri dengan
bantuan pedikel. Proses vitellogenik dimulai pada oosit tahap III dengan ukuran
oosit yang makin membesar. Pembesaran ini disebabkan oleh pembentukan
butiran lemak di dalam oosit. Pada tahap ini, oosit biasanya sudah mulai terlepas
dari mesenteri menuju rongga gastrovaskular.
Oosit tahap IV ditandai dengan semakin membesarnya ukuran oosit karena
menjadi terang. Pada oosit tahap V, oosit atau telur telah matang dan mencapai
ukuran maksimum dengan warna yang telah menjadi terang. Telur yang matang
ditandai dengan banyak butiran-butiran lemak di dalamnya.
(2) Spermatogenesis
Hwang dan Song (2007) membedakan perkembangan spermatogenesis
menjadi 4 tahap. Tahap I biasanya ditandai dengan berkumpulnya
spermatogonia di mesoglea pada mesenteri. Pada tahap II (spermatosit) sudah
memiliki batas dan bentuk yang jelas dan melekat pada mesenteri dengan bantuan
pedikel. Tahap III, ukuran kista sperma menjadi semakin besar. Spermatosit
berkembang menjadi spermatid yang jumlahnya sangat banyak dan tersusun di
bagian tepi dari kista. Pada tahap IV, spermatosit telah matang dengan
berkembang menjadi spermatozoa yang telah memiliki ekor.
Gambar 5 menunjukkan perkembangan perkembangan spermatit pada
Sarcophyton glaucum. (A) Gambaran umum dari polyp jantan menunjukkan perkembangan sperma, (B) Sperma dengan spermatogonia dan spermatosit, (C)
Spermatosit dengan spermatid , (D) Sperma matang dengan ekor . Huruf c
menunjukkan Coenenchyme; oa bagian oral dari polip ; pc rongga polip ; s1, Stage I Spermatositdengan spermatogonia; s2 , Stage II .spermatosit yag
menempel pada pedikel ; s3 , Stage III; s4 , Stage IV sperma yang telah matang.
2.3.4 Pemijahan dan Fertilisasi
Karang lunak alcyonacea memiliki tiga cara reproduksi untuk menjamin
kesuksesan reproduksinya yaitu pemijahan gamet ke kolom perairan (broadcast spawning), internal brooding, dan external brooding (Hwang dan Song, 2008).
a. Pemijahan gamet ke kolom perairan, merupakan cara reproduksi yang
paling umum terjadi pada karang lunak alcyonacea. Cara ini akan disertai
dengan proses fertilisasi dan perkembangan embryo di kolom perairan. Proses
pemijahan pada karang lunak biasanya mengikuti pemijahan massal secara
serempak dengan organisme lain di ekosistem terumbu karang sebagai suatu
bentuk strategi untuk mengurangi tekanan predasi pada gamet yang baru saja
dikeluarkan (Simpson, 2008).
b. Internal brooding biasa terjadi pada genus Xenia, Heteroxenia, dan
c. External brooding, terjadi pada genus Alcyonium dan Capnella. Telur akan dikeluarkan di permukaan koloni karang lunak dan menunggu hingga
terjadi proses fertilisasi. Cara ini merupakan strategi terhadap rendahnya
kesuburan gamet sebagai upaya untuk meningkatkan kelangsungan hidup larva
dari bahaya predasi.
2.4 Faktor yang Mempengaruhi Reproduksi Karang Lunak (Octocorallia)
Pola reproduksi karang lunak yang di dalamnya termasuk gametogenesis
dan strategi atau cara dalam melakukan reproduksi dipengaruhi oleh beberapa
faktor. Faktor-faktor yang berpengaruh bisa berupa faktor eksternal dan faktor
internal.
2.4.1 Faktor Internal
Ukuran dan Umur Koloni
Karang lunak memiliki ukuran dan umur yang bervariasi dalam proses
kematangannya untuk melakukan reproduksi. Di dalam satu spesies, ukuran
koloni merupakan salah satu indikator penting dalam menentukan kematangan
reproduksi. Lobophytum crassum dengan diameter kurang dari 18 cm belum bisa menghasilkan gamet untuk bereproduksi sementara Heteroxenia fuscescens
tidak akan mencapai kematangan reproduksi hingga ukuran koloninya mencapai
volu-me 10 cm3 . Hal ini merupakan strategi dalam melakukan reproduksi dimana energi/sumber daya dalam koloni dialokasikan terlebih dahulu untuk
pertumbuhan koloni hingga mencapai ukuran minimum untuk melakukan
Penelitian yang dilakukan di KwaZulu-Natal yang menggunakan
Sarchophyton glaucum, mengungkapkan bahwa gametogenesis dalam koloni laki-laki membutuhkan waktu 9-10 bulan sedangkan untuk koloni betina
membu-tuhkan waktu 16-18 bulan (D.G. Fautin et all , 2004 ).
2.4.2 Faktor Eksternal
(1) Ketersediaan Makanan
Ketersediaan sumber makanan yang terdapat di perairan mempengaruhi
waktu terjadinya gametogenesis dan proses spawning pada beberapa karang
lunak. Hartnoll (1975) menjelaskan bahwa perkembangan gamet pada
Alcyonium digitatum terjadi pada awal musim semi dan panas dimana sumber makanan planktonik sangat melimpah di perairan. Proses spawning pada spesies
ini terjadi pada pertengahan musim dingin dimana pada saat awal musim semi,
larva dari hasil proses pembuahan di kolom perairan akan memiliki banyak
persediaan makanan untuk berkembang. Benayahu (1991) menjelaskan bahwa
proses pembentukan larva planula pada Acabaria biserialis di Laut Merah terjadi setelah terjadinya blooming musiman pada fitoplankton.
(2) Fase Bulan
Karang lunak yang tumbuh di perairan dangkal, tingkah laku pengeluaran
telur ke perairan (spawning) memiliki hubungan dengan fase bulan. Gamet dilepas ke perairan pada saat atau menjelang bulan purnama dan pada sekitar atau
menjelang fase bulan baru/mati. Bahkan proses spawning juga terjadi pada fase bulan tiga perempat (Simpson, 2008). Namun, hasil yang berkebalikan
fase bulan dengan pelepasan larva planula pada karang lunak Heteroxenia fuscescens.
(3) Suhu Perairan
Suhu merupakan salah satu faktor yang berpengaruh dalam reproduksi
gamet pada karang lunak. Choi dan Song (2007) menjelaskan bahwa jumlah
oosit pada setiap polip meningkat setiap bulan hingga suhu perairan mencapai
nilai tertinggi dan menurun seiring dengan turunnya suhu perairan. Begitu juga
dengan jumlah gamet jantan yang meningkat pada bulan Agustus dan Desember
dimana suhu perairan meningkat. Pada bulan oktober, jumlah gamet jantan mulai
menurun karena suhu perairan menurun secara cepat.
(4) Intensitas Cahaya
Intensitas cahaya berpengaruh terhadap reproduksi karang lunak
dihubungkan dengan tingkat kejernihan perairan dan sedimentasi. Kojis dan
Quinn (1984) menjelaskan bahwa kedua faktor tersebut bisa menurunkan tingkat
kesuburan polip. Hal ini disebabkan energi yang digunakan untuk melakukan
reproduksi menjadi berkurang karena menurunnya intensitas cahaya untuk proses
fotosintesis oleh alga zooxanthellae di dalamnya dan pengalokasian energi /
sumber daya untuk membersihkan diri dari sedimen.
(5) Habitat
Jenis-jenis karang lunak hidup di daerah pasang surut sampai kedalaman
200 m. Umumnya syarat-syarat hidupnya sama dengan karang batu. Hewan ini
menyukai perairan yang hangat atau sedang terutama di Indo-Pasifik. Ada
Dan secara umum, karang lunak di daerah tropis melakukan reproduksi
dengan cara melepaskan gametnya ke kolom perairan secara serempak dengan
organisme lain di ekosistem terumbu karang sedangkan pada karang lunak di
daerah subtropis menunjukkan proses gametogenesis yang lama dan melakukan
reproduksi secara brooding (Cordes et al., 2001).
Pengaruh habitat pada reproduksi karang lunak juga terlihat pada tipe
seksualitas yang terjadi pada spesies Heteroxenia elizabethae. Di Great Barrier Reef, spesies ini menunjukkan tipe seksualitas gonokhorik sedangkan di Laut
Merah, spesies ini menujukkan tipe seksualitas hermaprodit.
2.5 Faktor – faktor Pembatas Pertumbuhan Karang Lunak (Octocorallia)
Faktor pembatas adalah faktor- faktor lingkungan yang dapat
mempenga-ruhi laju pertumbuhan suatu individu di dalam habitatnya. Pertumbuhan dan
per-kembangan karang lunak dipengaruhi oleh :
(1) Suhu
Menurut Nybakken (1992), pertumbuhan karang mencapai maksimum
pa-da suhu optimum 25-29 °C pa-dan bertahan hidup sampai suhu minimum 15°C pa-dan
maksimum 36°C. Pertumbuhan optimal terjadi di perairan yang memiliki
rata-rata suhu tahunan 23-25°C . Suhu ekstrim yang masih dapat ditoleransi adalah
36-40°C.
(2) Kecerahan dan Kedalaman
Hewan karang pembentuk terumbu membutuhkan sinar matahari bagi
zooxanthellae untuk berfotosintesis. Cahaya adalah suatu faktor yang paling
zooxanthellaes simbiotik dalam jaringan karang (Nybakken, 1992). Menurut
Nybakken (1992), terumbu karang tidak dapat berkembang di perairan yang lebih
dalam dari 50-70 meter. Zooxanthellae sebagai alga simbiotik yang memerlukan
cahaya matahari sehingga terjadi sedikit pertumbuhan di bawah kedalaman 46
meter dan di bawah kedalaman 90 meter terumbu karang sudah sangat jarang.
Faktor kecerahan dan kedalaman pada karang lunak berperan untuk melakukan
proses fotosintesis, hal ini dikarenakan karang lunak membutuhkan cahaya yang
cukup.
(3) Salinitas
Salinitas rata-rata di daerah tropis adalah 35 ‰ dimana masih berada pada
kisaran optimum untuk pertumbuhan karang yaitu 34-36‰ (Supriharyono, 2000).
Nybakken (1992) menyatakan bahwa toleransi organisme karang terhadap
salinitas berkisar antara 32-35‰.
(4) Derajat Keasaman (pH)
Derajat keasaman menunjukkan aktivitas ion H+ dalam air. Menurut
Tomascik (1997), habitat yang cocok bagi pertumbuhan karang memiliki kisaran
pH 8, 2-8, 5.
(5) Pergerakan Arus
Pergerakan arus sangat diperlukan untuk tersedianya aliran suplai makanan
(dalam bentuk jasad renik) dan suplai oksigen yang segar, serta menjaga agar
III. BAHAN DAN METODE
3.1 Waktu dan Lokasi Penelitian
Sampel karang lunak yang digunakan untuk penelitian di laboratorium
diperoleh dari stasiun pengamatan yang berada di Area Perlindungan Laut, Pulau
Pramuka, Kepulauan Seribu. Pengambilan dilakukan dalam dua tahap, yaitu
tahap I (umur 8 bulan setelah fragmentasi) dilakukan pada tanggal 8, 15, 22, dan
29 Mei 2009 serta tahap II (umur 10 bulan setelah transplantasi) dilakukan pada
tanggal 7, 14, 21, dan 28 Juli 2009. Pengolahan sampel, pembuatan, dan
pengamatan preparat histologis dilakukan di Laboratorium Histologi, Fakultas
Kedokteran Hewan, Institut Pertanian Bogor dari bulan Maret hingga Oktober
2009.
3.2 Alat dan Bahan
Alat dan bahan yang digunakan dalam penelitian ini adalah alat yang di
gunakan untuk pemotongan sampel , pewarnaan dan penghitungan telur.
Adapun alat dan bahan tersebut akan disajikan dalam Tabel 2.
Tabel 2. Alat dan bahan yang digunakan pada saat penelitian
Pengolahan Sampel
Alat Bahan
Alat pemanas air Eyela Water Bath
SB-650 Formalin 10%
Basket jaringan
Karang lunak Sarcophyton crassocaule
Blok kayu Air destilasi dan air kran
Botol film dan plastik sampel Enthellan
Pisau cutter
Etanol (70%, 80%, 90%, dan 100%)
Embedding Consule Tissue-Tek TEC Asam Formik Gelas objek dan cover glass Asam Klorida Inkubator Eyela Soft Incubator SL-450N Parafin
Inkubator Memmert Pewarna Hematoxylin dan Eosin
Jarum Xylol
Mikrometer objectif dan okuler Mikroskop cahaya dan stereo Olympus
CH 20
Mikrotom Spencer 820
Tutup Pagoda
3.3 Metode Kerja
3.3.1 Pembuatan Preparat Histologis
Pembuatan preparat histologis dilakukan dengan metode parafin (Gunarso,
1989 dan Kiernan, 1990) dengan tahapan mencakup: (1) Pengambilan jaringan
(70 – 100% ); (5) Penjernihan (clearing) menggunakan xylol; (6) Infiltrasi parafin (infiltration) menggunakan parafin cair dalam inkubator bersuhu 65 Penanaman (embedding) menggunakan parafin; (8) Penyayatan (sectioning) menggunakan mikrotom (±5µm); (9) Afiksing (afixing); (10) Deparafinasi (deparaffination) menggunakan xylol; (11) Rehidrasi (rehydration)
menggunakan alkohol bertingkat (100-70%) dan air (12) Pewarnaan (staining) menggunakan pewarnaan Hematoksilin-Eosin (HE), tahap akhir adalah mounting
dengan menggunakan entelan. Pembuatan preparat histologi dilakukan sebanyak
6 sayatan untuk setiap cabang koloni karang lunak yang diolah. Selanjutnya,
pengamatan struktur histologis dilakukan terhadap kehadiran dan perkembangan
gonad (telur dan sperma). Tahap terakhir dilakukan pemotretan atau
mikrofotografi menggunakan mikroskop yang dilengkapi kamera dengan
perbesaran 40x , 100x , dan 200x
3.3.2 Pengamatan Mikroskopis
Pengamatan preparat histologis dilakukan di Laboratorium Histologi,
Fakultas Kedokteran Hewan, Institut Pertanian Bogor. Pengamatan pada
mikro-skop dilakukan dengan perbesaran lensa sebesar 40x, 100x, 200x, dan 400x.
Objek yang diamati pada pengamatan ini, antara lain:
a. Tahap perkembangan oosit
Perkembangan oosit diamati berdasarkan ciri morfologi dan ukuran dalam
b. Jumlah oosit
Jumlah oosit ditentukan berdasarkan jumlah gamet yang ditemukan dalam
setiap cabang koloni (jumlah oosit per cabang koloni)
c. Diameter oosit
Oosit yang diukur adalah oosit yang tampak nukleus atau nukleolusnya.
Panjang oosit diukur pada bagian oosit yang paling panjang sedangkan
lebarnya diukur secara tegak lurus terhadap garis panjangnya.
3.3.3 Pengukuran Parameter Lingkungan
Parameter lingkungan yang diukur adalah parameter fisika dan kimia yang
dilakukan secara in situ dan pengamatan melalui analisis laboratorium. Parameter yang akan diukur, satuan, dan alat/metode yang digunakan dalam
pengukuran pa-rameter lingkungan dapat dilihat pada Tabel 3.
Tabel 3. Parameter fisika-kimia perairan yang diukur
Parameter Satuan Alat/ metode
Suhu ◦C Termometer Hg
Kecerahan meter (m) Secchi disk
Kecepatan arus cm/s Floating Droadge
Kedalaman meter (m) Depth gauge
Derajat keasaman Ph Kertas Ph
3.4 Analisis Data
3.4.1 Analisis Deskriptif
Pengamatan yang dilakukan dengan mikroskop untuk menganalisa
kembangan gamet dilakukan dengan perbesaran lensa sebesar 400x. Tahap
per-kembangan gamet dan pengaruh fase bulan Qomariyah dianalisis secara deskriptif
berdasarkan keberadaan gamet pada setiap fase bulan Qomariah dengan
meng-amati karakter gamet pada preparat histologis dan gambar hasil mikrofotografi
kemudian membandingkannya dengan pustaka terbaru dan jurnal terkait.
3.4.2 Analisis Ragam
Jumlah gamet dinyatakan dengan jumlah gamet yang ditemukan per cabang
koloni. Data yang diperoleh kemudian dianalisis menggunakan uji t ,
menggunakan selang kepercayaan 95% yang dioperasikan dengan bantuan
software Microsoft Excel 2007. Analisis ini dilakukan untuk melihat pengaruh penggunaan metode fragmentasi dan kedalaman lokasi fragmentasi terhadap
perkembangan reproduksi karang lunak Sarcophyton crassaule dalam setiap fase bulan Qomariyah.
Analisis ragam akan menunjukkan beda nyata atau tidak pada
perkembangan reproduksi karang lunak Sarcophyton crassaule berdasarkan kedalaman dan penggunaan metode fragmentasi. Jika nilai thit < t one tail maka terima Ho berarti tidak ada perbedaan yang nyata pada perkembangan reproduksi
karang lunak terhadap kedalaman ataupun penggunaan metode fragmentasi. Jika
nilai thit > t onetail maka tolak Ho yang berarti ada perbedaan yang nyata pada perkembangan reproduksi karang lunak terhadap kedalaman ataupun penggunaan
IV. HASIL DAN PEMBAHASAN
4.1 Kondisi Lingkungan Stasiun Penelitian
Parameter fisika dan kimia yang diukur untuk mengetahui kondisi stasiun
penelitian meliputi; suhu, kecerahan, kecepatan arus, pH, oksigen terlarut, nitrat,
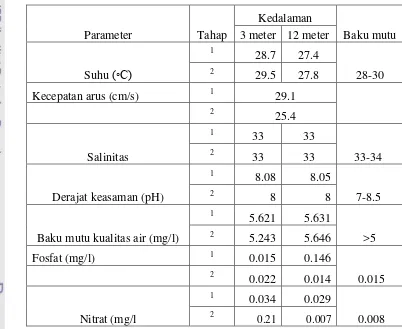
dan fosfat. Hasil pengukuran dapat dilihat pada Tabel 4.
Tabel 4. Hasil pengukuran parameter fisika dan kimia di stasiun penelitian
Parameter Tahap
Baku mutu kualitas air (mg/l) 1
Berdasarkan data yang di dapatkan dari lingkungan stasiun penelitian di
dapatkan beberapa informasi penting, diantaranya adalah suhu perairan pada
adalah sebesar 27.4 oC dan 27.8 oC pada tahap 1 dan 2 pengukuran, perbedaan suhu tersebut dapat disebabkan karena perbedaan kedalaman sehingga terjadi
layetring suhu di perairan , untuk kedalaman 3 meter besar suhunya masih dalam
kisaran baku mutu , sedangkan untuk kedalaman 12 meter kisaran suhunya berada
di bawah standar baku mutu namun masih dalam pertumbuhan maksimum
karena menurut pertumbuhan karang mencapai maksimum pada suhu optimum
25-29 °C ( Nybakken , 1992).
Tabel di atas menunjukkan nilai kecepatan arus pada kedalaman 3 meter
sebesar 29.1 cm/s sedangkan pada kedalaman 12 meter kecepatannya sebesar 25.4
cm/s. Penyebab arus permukaan lebih cepat di bandingkan kecepatan arus pada
perairan dalam, hal ini di sebabkan karena arus pada daerah dangkal di pengaruhi
oleh angin. Bagi karang arus berperan untuk membawa masuk makanan ke
habitat koral dan mengangkut limbah dari koloni karang dan merangsang
terjadinya fotosintesis (Fabricius, K dan P.Alderslade ) . Namun jika dilihat dari
kondisi rak-rak fragmentasi dapat dilihat bahwa terjadi penumpukan pertumbuhan
alga di sekitar rak-rak transplant yang menjadi indikator bahwa bahan organik
pada daerah tersebut berada dalam jumlah yang besar. Banyaknya bahan
organik pada suatu perairan dapat dikarena dekatnya lokasi perairan dengan darat
dan kurang berperannya kecepatan arus pada lokasi tersebut untuk membawa
limbah atau bahan organik keluar dari habitat karang itu sendiri.
Kondisi di atas diperkuat dengan data dari kandungan senyawa fosfat dan
nitrat yang ada pada lokasi penelitian yang di tampilkan pada table di atas yang
hasilnya adalah sebagai berikut nilai kandungan fosfat pada kedua kedalaman
baku mutu kualitas air. Nilai kandungan nitrat pada kedalaman 3 meter
menunjukkan selang nilai 0.034 - 0.21 mg/ l nilai ini jauh melampaui nilai baku
mutu yang hanya sebesar 0.008 mg/l. Hal ini juga terjadi pada kedalaman 12
meter dimana nilai dari kandungan nitratnya antara 0.07- 021mg/l .
Fosfat adalah indikator jumlah nutrient yang berada pada kolom perairan
yang dibutuhan oleh fitoplankton termasuk alga untuk meakukan fotosintesis ,
Selain nilai kandungan Fosfat nilai kandungan nitrat juga menjadi indikator
apakah perairan tersebut subur atau kaya nutrient. Tingginya kandungan nitrat
dan fosfat pada kolom perairandapat memacu pertumbuhan alga yang akhirnya
akan memacu kompetisi cahaya dan ruang bagi pertumbuhan koral itu sendiri.
Tingginya nilai kedua nutrient seperti yang telah di jelaskan pada paragraph
sebelumnya bahwa hal ini dibuktikan dengan keruhnya perairan di sekitar lokasi
penanaman karang lunak hasil transplantasi yang menandakan tingginya bahan
organik yang berada di kolom perairan dan tumbuhnya banyak alga di rak
fragmentasi.
Data salinitas yang didapatkan dari hasil pengukuran pada kedua tahap
pengambilan pada dua kedalaman air adalah sama yaitu 33 o/oo. Dan nilai ini masih berada dalam kisaran baku mutu salinitas untuk karang yaitu 33-34 o/oo . Maka perairan tersebut dapat dikatakan baik karena masih dalam kisaran salinitas
yang di maksud.
Informasi untuk nilai derajat keasaman didapatkan kisaran ph yaitu antara 8
– 8.08 yang dan kisaran tersebut masih berada dalam kisaran baku mutu kualitas
air masih berada pada yaitu antara 7 – 8.5. Begitupun dengan nilai pengukuran
atas 5 mg/l dan hal ini sesuai dengan baku mutu kualitas air untuk karang.
Nutrien seperti nitrat dan fosfat mempengaruhi pertumbuhan alga pada karang.
Alga cenderung tumbuh lebih baik pada perairan yang kaya akan nutrien.
Gambar 7 mrnunjukkan kondisi alga yang tumbuh di sekitar rak penanaman
fragmentasi karena pengaruh kandungan bahan organik perairan.
4.2 Pengamatan Mikroskopis terhadap Karang Lunak Sarcophyton
crassaule
Menentukan seksualitas dari karang lunak maka dibutuhkan pendekatan
yang dilakukan dengan melakukan pengamatan histologis tentang keberadaan
gamet jantan (spermatosit) dan gamet betina (oosit) pada setiap bagian tubuh
koloninya. Mengingat pengambilan dilakukan tanpa memperhatikan jenis
kelamin di koloni pada area pertumbuhan di alam upaya identifikasi dalam koloni
dan system reproduksi Sarcophyton crassocaule hingga kini belum banyak di laporkan
Pengamatan yang dilakukan terhadap sayatan menegak bagian tubuh koloni
karang lunak menunjukkan gamet di temukan tidak merata pada seluruh bagian
sayatan yang di amati. Gamet dalam hal ini Oosit ditemukan dalam posisi soliter
(terpisah) dan berkelompok dengan gamet lain pada rongga gastrovaskuler yang
tersebar pada bagian tubuh koloni. Hasil yang di dapatkan adalah posisi gamet
yang berada di rongga gastrovaskuler cenderung menempel pada mesenteri , dan
untuk gamet yang berada pada tahap perkembangan awal cenderung
adanya stage lanjutan sehingga posisi untuk stage akhir tidak dapat di gambarkan
pada penelitian kali ini.
Hasil lain yang didapatkan dari pengamatan selain melihat posisi dan pola
penyebaran dari Oosit adalah semua koloni (72 koloni sampel) yang diamati
secara mikroskopis semuanya adalah koloni betina. Hal ini dibuktikan dengan
tidak di temukan spermatosit pada saat pengamatan dilakukan. Keseragaman dari
jenis kelamin karang dikarenakan karang yang digunakan untuk melakukan
fragmentasi diambil pada lokasi yang sama baik untuk penanaman 3 m, 12 m
maupun karang lunak control (non transplant) . Hanya ditemukannya Oosit pada
semua koloni yang diteliti memberikan informasi bahwa pada lokasi tersebut
Sarcophyton crassocaule bersifat gonokhorik (Hwang dan Song, 2007; Simpson, 2008).
4.3 Oogenesis dan Ukuran Gamet Betina Karang Lunak Sarcophyton
crassocaule
Penelitian yang dilakukan di KwaZulu-Natal mengungkapkan
gametogenesis Sarchophyton glaucum dalam koloni laki-laki membutuhkan waktu 9-10 bulan sedangkan untuk koloni betina membutuhkan waktu 16-18
bulan (D.G. Fautin et all , 2004 ). Dan perkembangan dari Oosit dapat diamati dengan melihat perbedaan diameter gamet dan ciri morfologi yang kita amati
dalam preparat histologis karang lunak yang telah di buat.
Tahap oogenesis dapatditentukan dengan mengidentifikasi berdasarkan
ciri dan morfologi dari telur. Oogenesis yang ditemukan pada penelitian kali
hanya di temukan dalam dua tahap pertumbuhan yaitu oosit tahap I dan oosit
tahap II . Tabel berikut akan menunjukkan tahapan oosit dan diameter dari oosit
yang ditemukan pada penelitian kali ini.
Tabel 5. Ukuran diameter oosit pada setiap tahap perkembangan
Tahap Diameter (µm)
Rata±SD n
I 6.36±1.26 50
II 9.2 ±1.02 50
(Sumber : Diolah dari lampiran 11)
Gambar 7. Oogenesis tahap I karang lunak Sarcophyton crassocaule yang menempel pada saluran mesenteri (Tanda panah menunjukkan O1 dengan skala garis 20 µm)
Gambar 8. Oogenesis tahap II karang lunak Sarcophyton crassocaule yang menempel pada saluran mesenteri (Tanda panah menunjukkan O2 dengan skala garis 20 µm)
Dua gambar di atas menunjukkan kecenderungan pada oosit tahap I
dite-mukan berkelompok dengan oosit tahap I lainnya dan menempel pada mesoglea
di mesenteri filament. Kita dapat melihat pada Gambar 7 bahwa Oosit tahap I
memiliki ciri-ciri yaitu batasan antara nucleus, nucleolus dan sitoplasmanya
belum jelas. Dan dari Tabel 5 didapatkan bahwa pada oosit tahap I memiliki
diameter rata-rata 6.36±1.26 µm (n=50).
Oosit tahap dua digambarkan dengan cukup jelas pada Gambar 8, jika
dibandingan dengan Oosit tahap I maka pada Oosit tahap II batasan antara
nukleus, nukleoulus dan sitoplasma sudah jelas terlihat . Pada Oosit tahap II telur
di temukan di ronga gastrovaskuler atau masih melekat pada mesenteri yang
terhubung melalui pedikel (Gambar 8) . Pada oosit tahap II memiliki ukuran
diameter 9.2 ±1.02 µm (n=50).
Hasil yang didapatkan penelitian kali ini adalah tidak di temukan
perkembangan gametogenesis yang lengkap (Stage I hingga Stage V) Hal ini
dapat dikaitkan dengan beberapa faktor diantaranya adalah faktor internal yang
memepengaruhi perkembangan gamet karang yang menyebutkan bahwa karang
lunak memiliki ukuran dan umur yang bervariasi dalam proses kematangannya
untuk melakukan reproduksi. Di dalam satu spesies, ukuran koloni merupakan
salah satu indikator penting dalam menentukan kematangan reproduksi.
Lobophytum crassum dengan diameter kurang dari 18 cm belum bisa
menghasilkan gamet untuk bereproduksi sementara Heteroxenia fuscescens tidak akan mencapai kematangan reproduksi hingga ukuran koloninya mencapai
volume 10 cm3 . Hal ini merupakan strategi dalam melakukan reproduksi dimana energi/sumber daya dalam koloni dialokasikan terlebih dahulu untuk
pertumbuhan koloni hingga mencapai ukuran minimum untuk melakukan
reproduksi (Gutiérrez dan Lasker, 2004).
Fragmentasi yang menggunakan jenis Sarcophyton crassocaule ini meng-ambil sampel dengan diameter kapitulum antara 5-6 cm . Dari ukuran diameter
tersebut maka dapat dilihat bahwa ukuran dan umur dari Sarcophyton crassocaule
yang di ambil untuk fragmentasi masih jauh dari ukuran untuk bereproduksi hal
inilah yang menyebabkan tidak ditemukannya gametogenesis secara lengkap
da-lam tubuh Sarcophyton crassocaule. Sedangkan jika di kaji dari segi umur menu-rut penelitian yang dilakukan di KwaZulu-Natal mengungkapkan
waktu 9-10 bulan sedangkan untuk koloni betina membutuhkan waktu 16-18
bulan (D.G. Fautin et all , 2004 ). Sedangkan umur karang lunak Sarcophyton crassocaule yang di-gunakan untuk fragmentasi kali ini adalah 8 dan 10 bulan. Maka jika dikaji dari segi umur maka dapat dikatakan bahwa faktor umur
menjadikan Sarcophyton crassocaule ini belum layak untuk dikaji dari proses gametogenesis yang berlangsung pada koloni yang di fragmentasi.
Tidak hanya faktor internal yang mempengaruhi kematangan gonad
ter-nyata faktor eksternal berupa intensitas cahaya berpengaruh terhadap reproduksi
karang lunak dihubungkan dengan tingkat kejernihan perairan dan sedimentasi.
Kojis dan Quinn (1984) menjelaskan bahwa kedua faktor tersebut bisa
menurun-kan tingkat kesuburan polip. Hal ini disebabmenurun-kan energi yang digunamenurun-kan untuk
melakukan reproduksi menjadi berkurang karena menurunnya intensitas cahaya
untuk proses fotosintesis oleh alga zooxanthellae di dalamnya dan pengalokasian
energi atau sumber daya untuk membersihkan diri dari sedimen. Hal ini
mempe-ngaruhi kematangan gonad Sarcophyton crassocaule pada proses fragmentasi, terbukti dari tingginya nilai pospat dan nitrat yang di dapatkan pada pembahasan
sebelumnya mengindikasikan bahwa pada stasiun pengamatan tersebut kandungan
bahan organiknya tinggi sehingga mempengaruhi pertumbuhan alga pada rak
frag-mentasi sehingga kematangan gonad dari karang lunak sulit dicapai karena
ener-ginya digunakan untuk berkompetisi dengan cahaya dengan alga karena sulitnya
4.4 Pengaruh Fragmentasi pada Reproduksi Seksual Sarcophyton
crassocaule
Menurut Sprung dan Delbeek (1997) dalam Setyawan ( 2008) Fragmentasi
adalah penempelan fragmen buatan akan berhasil dengan baik bila kondisi
lingkungan pun optimal dan substrat dasarnya pun baik. Tujuan utama
fragmentasi karang adalah untuk memperbaiki kualitas terumbu karang seperti
meningkatnya tutupan karang hidup, keanekaragaman hayati dan keunikan
topografi karang (Clark dan Edwards, 1998).
Penelitian ini ingin meliihat pengaruh dari jumlah oosit antara produk hasil
fragmentasi dengan alam. Nilai yang kami bandingkan adalah nilai jumlah
rata-rata oosit yang pada karang non fragmentasi yang berada pada kedalaman yang
sama (12 m) dengan karang fragmentasi.
Gambar 9 menunjukkan bahwa rata-rata jumlah oosit pada karang lunak
fragmentasi memiliki nilai yang sedikit lebih tinggi dibandingkan dengan nilai
karang lunak non transplantasi (pada 3 fase bulan yaitu purnama, tiga perempat
dan mati baru) , namun jika dihitung secara statistik nilai keduanya tidak berbeda
nyata. Hal ini di perkuat lagi dengan dilakukan analisis ragam yang
menggunakan uji t. Hasil yang di dapatkan dari analisis ragam terhadap
penggunaan metode fragmentasi pada rata-rata jumlah oosit menunjukkan bahwa
Gambar 9. Rata-rata jumlah oosit pada karang lunak non fragmentasi (alam) dan hasil fragmentasi umur 8 bulan setelah fragmentasi
Diterimanya Ho menjelaskan bahwa antara rata-rata jumlah oosit karang
lunak pada hasil fragmentasi dengan non fragmentasi tidak berbeda nyata pada
selang kepercayaan 95%. Namun adanya nilai yang lebih tinggi pada karang
lunak hasil fragmentasi dapat disebabkan sebagai proses homeostasis yang terjadi
pada organisme yang mengalami keadaaan stress akan berusaha memproduksi
lebih banyak telur , namun hanya pada tingkat awal perkembangan karena untuk
mematangkannya dibutuhkan banyak energi dan energi tersebut diprioritaskan
oleh karang lunak untuk menyembuhkan dirinya dan bertahan dari lingkungan
Gambar 10. Komposisi rata-rata jumlah oosit tahap1 dan tahap 2 pada karang lunak non fragmentasi (alam) dan hasil fragmentasi umur 8 bulan setelah fragmentasi
Keterangan : (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat
Gambar 10 menunjukkan bahwa walaupun secara statistik rata-rata jumlah
oosit pada karang lunak hasil fragmentasi dengan karang lunak non fragmentasi
(alam) tidak berbeda nyata terlihat bahwa komposisi dari keseluruhan jumlah
oosit (O1 dan O2 ) antara karang lunak non fragmentasi dengan hasil fragmentasi
setelah umur 8 bulan didominasi dengan oosit tahap satu (O1) pada setiap fase
bulan pada kedua jenis karang lunak tersebut.
Gambar 11. Rata-rata jumlah oosit pada karang lunak non fragmentasi (alam) dan hasil fragmentasi umur 10 bulan setelah fragmentasi
Gambar 11 menunjukkan nilai rata-rata jumlah oosit pada karang lunak non
fragmentasi (alam) dan hasil fragmentasi pada kedalaman yang sama (12 m)
namun pada umur yang berbeda (10 bulan). Seperti halnya yang terjadi pada
Gambar 9 yaitu pada umur 8 bulan dari tabel dapat dilihat terdapat perbedaan nilai
antara karang lunak hasil fragmentasi dengan karang lunak non trasplantasi pada
fase bulan tiga perempat dan seperempat . Dimana nilai pada fase bulan tersebut
karang lunak hasil fragmentasi memiliki nilai jumlah yang lebih besar di
bandingkan dengan karang non fragmentasi.
Hasil di atas apabila diuji secara statistik maka hasil yang didapatkan adalah
terima Ho atau tidak ada perbedaan nyata antara nilai jumlah karang lunak hasil
fragmentasi dengan non fragmentasi karena hasil yang di dapatkan dari analisis
ragam terhadap penggunaan metode fragmentasi pada rata-rata jumlah oosit
memiliki ritme yang sama dengan karang lunak non transplant yaitu mengalami
kenaikan pada fase bulan tiga perempat, penurunan pada fase bulan mati baru dan
terus turun hingga fase bulan seperempat. Hal ini dapat diasumsikan karena
pada umur 10 bulan karang lunak hasil fragmentasi berada pada kondisi
pemulihan yang jauh lebih baik dibandingkan dengan dengan karang lunak pada
umur 8 bulan. Proses penyembuhan dari luka yang terjadi pada saat proses
fragmentasi telah berjalan baik sehingga ritme tubuh karang lunak hasil
fragmentasi menjadi lebih normal.
Gambar 12. Komposisi Rata-rata jumlah oosit tahap1 dan tahap 2 pada karang lunak non fragmentasi (alam) dan hasil fragmentasi umur 10 bulan setelah fragmentasi
Keterangan : (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat
Gambar 12 menunjukkan komposisi O1 dan O2 pada karang lunak
fragmentasi (12 meter) dan non fragmentasi pada umur 10 bulan. Komposisi
rata-rata jumlah pada kedua tahap tersebut didominasi oleh oosit tahap pertama (O1)
0
10 Bulan Setelah Transplantasi
pada setiap fase bulan baik pada karang lunak non fragemntasi maupun karang
lunak hasil fragmentasi. Hal ini menunjukkan baik karang lunak non fragmentasi
maupun fragmentasi belum mencapai kemaanganseksual.
Fragmentasi di gunakan sebagai salah satu solusi untuk memperbanyak
koloni karang lunak tanpa menunggu proses alami yang cenderung lama dan
terpengaruh banyak faktor baik internal maupun eksternal . Pada hasil yang di
dapatkan jumlah gamet pada masa awal fragmentasi cenderung bersifat labil atau
tidak sama dengan jumlah oosit yang ada di alam , hal ini di buktikan pada
Gambar 10 dimana jumlahnya cukup berbeda dan tidak sesuai dengan keadaan
pada alam (terjadi kenaikan jumlah pada fase bulan seperempat sementara pada
karang lunak non fragmentasi mengalami penurunan). Hal ini dapat disebabkan
karena karang lunak fragmentasi masih melakukan proses penyesuaian terhadap
lingkungan dan proses penyembuhan luka yang terjadi pada tubuhnya, sehinga
reproduksinya menjadi tidak stabil. Ketika kondisi karang telah mulai pulih
maka yang terjadi adalah ritme positif dari jumlah telur perfase bulan pada karang
lunak hasil fragmentasi menjadi sama dengan karang hasil non fragmentasi .
4.5 Pengaruh Kedalaman Lokasi Fragmentasi pada Reproduksi Seksual
Sarcophyton crassocaule .
Karang lunak hasil fragmentasi diberikan perlakuan yang berbeda yaitu rak
ditanam pada kedalaman 3m dan 12 meter. Hal ini dilakukan untuk melihat
apakah ada pengaruh kedalaman terhadap reproduksi karang lunak yang dapat
dilihat dari kepadatan oosit pada masing-masing koloni fragmentasi pada
0
purnama tigaperempat mati baru seperempat
R kedalaman mana yang paling baik untuk melakukan fragmentasi tanpa
menggangu proses reproduksi seksualnya.
Gambar 13 memperlihatkan bahwa terjadi ritme yang sama antara karang
lunak pada kedua kedalaman fragmentasi dimana nilai dari jumlah oositnya
cenderung naik dari fase bulan purnama ke fase bulan tigaperempat pada umur 8
bulan namun pada fase bulan mati baru menuju seperempat untuk karang lunak
yang di fragmentasi pada kedalaman 12 meter mengalami penurunan rata-rata
jumlah oosit.
Gambar 13. Rata-rata jumlah oosit pada karang lunak fragmentasi pada kedalaman 3 meter dan 12 meter pada umur 8 bulan setelah fragmentasi
Ketidakteraturan ritme jumlah oosit pada setiap fase bulannya dapat
dikarenakan kondisi karang yang belum stabil dalam mengobati luka dan berjuang
dengan lingkungan barunya. Hal ini diperkuat dengan dilakukannya analisis
hasil yang di dapatkan adalah pada selang kepercaaan 95% memperlihat-kan nilai
P value > 0.05 (Lampiran 9 dan 10) . Hal ini berarti Terima Ho atau bisa
diinterpretasikan bahwa tidak ada perbedaan yang nyata antara jumlah oosit pada
kedalaman karang lunak hasil fragmentasi pada kedalaman 3 meter dengan 12
meter pada umur 8 bulan.
Gambar 14. Komposisi Rata-rata jumlah oosit tahap1 dan tahap 2 pada karang lunak fragmentasi pada kedalaman 3 dan 12 meter , 8 bulan setelah fragmentasi
Keterangan: (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat.
Gambar 14 menunjukkan komposisi antara oosit tahap pertama dan oosit
tahap kedua pada karang lunak hasil fragmentasi yang ditanam pada kedalaman
yang berbeda yaitu 3 dan 12 meter. Komposisi rata-ratajumlah oosit
0
purnama tigaperempat mati baru seperempat
R
menunjukkan bahwa komposisi oosit tahap pertama mendominasi oosit tahap
kedua pada setiap fase bulan dikedua kedalaman lokasi penanaman tersebut.
Gambar 14 menunjukkan pengaruh kedalaman lokasi penanaman fragmen
pada umur 10 bulan. Jika dilihat pada gambar tersebut maka kita masih melihat
adanya perbedaan jumlah oosit antara kedalaman tiga dan 12 meter . Pada
kedalaman 3 meter rata-rata jumlah oosit jauh terlihat lebih besar jika
dibandingkan dengan rata-rata jumlah oosit pada kedalaman 12 meter. Hal ini
dapat disebabkan karena posisi kedalaman 3 meter memiliki karakteristik habitat
yang lebih menantang bagi karang lunak itu sendiri , dari segi arus perairan dan
pertumbuhan alga akibat banyaknya bahan organik yang terdapat pada kedalaman
tersebut sehingga keadaanya jauh lebih stress dibandingkan dengan karang lunak
hasil fragmentasi yang di tanam pada kedalaman 12 meter.
Gambar 15. Rata-rata jumlah oosit pada karang lunak fragmentasi pada kedalaman 3 meter dan 12 meter pada umur 10 bulan setelah fragmentasi
Kita melakukan analisis ragam untuk mendukung hasil pada Gambar 14 ,
hasil yang di dapatkan adalah pada selang kepercaaan 95% memperlihatkan nilai
diinterpretasikan bahwa tidak ada perbedaan yang nyata antara jumlah oosit pada
kedalaman karang lunak hasil fragmentasi pada kedalaman 3 meter dengan 12
meter pada masa fragmentasi 10 bulan. Dari hasil ini maka dapat diambil
informasi bahwa tidak ada perbedaan yang nyata antara lokasi penanaman karang
pada kedalaman 3 meter dengan 12 meter sehingga kedalaman lokasi penanaman
frgamen karang lunak tidak berpengaruh pada perkembangan reproduksi karang
lunak Sarcophyton crassocaule.
Gambar 16. Komposisi Rata-rata jumlah oosit tahap1 dan tahap 2 pada karang lunak fragmentasi pada kedalaman 3 dan 12 meter , 10 bulan setelah fragmentasi
Keterangan : (O1) Oosit tahap I, (O2) Oosit tahap II, (P) Fase bulan Purnama, (T) Fase bulan Tiga per empat, (M) Fase bulan Mati, (S) Fase bulan Seperempat
Gambar 16 menunjukkan adanya peningkatan komposisi jumlah oosit
tahap kedua jika dibandingkan dengan 8 bulan setelah fragmentasi. Untuk karang
lunak yang difragmentasi dan ditanam pada kedalaman 3 meter komposisi O2 dan
O1 cukup seimbang sedangkan pada karang lunak yang ditanam pada kedalaman 0
10 Bulan Setelah Transplantasi
12 meter komposisi O1 masih mendominasi O2 dari rata-rata jumlah oosit yang
V. KESIMPULAN DAN SARAN
5.1 Kesimpulan
Perkembangan Oosit karang lunak Sarcophyton crassocaule antara hasil fragmentasi dan non fragmentasi (alam) tidak berbeda nyata pada umur 8 bulan maupun 10 bulan setelah fragmentasi.
Fragmentasi tidak mempengaruhi perkembangan Oosit pada karang lunak baik pada bulan ke -8 maupun ke-10 setelah fragmentasi.
Kedalaman penanaman hasil fragmentasi tidak menunjukkan perbedaan yang nyata antara penanaman di kedalaman 3 ataupun 12 meter.
5.2 Saran
Perlu diadakan penelitian minimal 16-18 bulan agar didapatkan rangkaian
proses gametogenesis yang lengkap dari karang lunak.
Ukuran karang yang dijadikan untuk fragmentasi harus sesuai dengan
DAFTAR PUSTAKA
Bayer, F.M., 1956. Octocorallia. in: Treatise on Invertebrata Paleontology, Part F Coelenterata. University of Kansas Press, Lawrence.
Benayahu, Y. 1991. Reproduction and developmental pathways of Red Sea Xeniidae (Octocorallia, Alcyonacea). Hydrobiologia 216/217: 125-130. (R.C. Moore ed.). Geological Society of America and Univ. Kansas Press, Lawrence.
Birkeland, C. 1997. LifeandDeath of Coral Reefs. Chapman and Hall. International Thamson publishing. New York.
Choi, E.J. dan J.I Song, . 2007. Reproductive Biology of the Temperate Soft Coral Dendronephthya suensoni (Alcyonacea: Nephtheidae). Integrative Biosciences 11: 215-225.
Cordes, E.E., Nybakken, dan D. G Van. 2001. Reproduction and growth of
Anthomastus ritteri (Octocorallia: Alcyonacea) from Monterey Bay, California, USA. MarBiol 138: 491-501.
D.G. Fautin, J.A. Westfall, P. Cartwright, M. Daly & C.R. Wyttenbach (eds). 2004. Coelenterate Biology 2003: Trends in Research on Cnidaria and Ctenophora. Hydrobiologia 530/531: 399–409 . Kluwer Academic Publishers.. Amsterdam.
Fabricius, K. dan P, Alderslade. 2001. Soft Coral and Sea Fans : A Comprehensive guide to the tropical shallow-water genera of the CentralWest Pasific, the Ocean and the Red Sea. Australian Institute of Marine Science. Townsville.
Gunarso, W. 1989. Bahan Pengajaran Mikroteknik. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Gutiérrez, C. dan H.R Lasker, . 2004. Reproductive biology, development, and planula behavior in the Caribbean gorgonian Pseudopterogorgia
elisabethae. Invertebr Biol 123(1): 54–67
Haris, A. 2001. Laju pertumbuhan dan tingkat kelangsungan hidup fragmentasi buatan karang lunak (Octocorallia: Alcyonacea) Sarcophyton
Harrison, P. L. dan C. C. Wallace. 1990. Reproduction, dispersal and
recruitment of scleractinian corals in Z. Dubinsky (ed.), Coral Reefs. Ecosystems of the world. Volume 25. Elsevier. Amsterdam.
Hartnoll, R.G. 1975. The annual cycle of Alcyonium digitatum. Estuarine and Coastal Marine Science 3: 71–78 .
Hwang, S.J. dan J.I Song, . 2007. Reproductive biology and larval development of the temperate soft coral Dendronephthya gigantea
(Alcyonacea: Nephtheidae). Mar Biol 152:273–284
Kiernan, J.A. 1990. Histological and Histochemical Methods: Theory and Practice. 2nd Edition. Pergamon Press. Oxford.
Kojis, B.L. dan Quinn, N.J. 1984. Seasonal and Depth Variation in Fecundity of Acropora palifera at Two Reefs in Papua New Guinea. Coral reef
3:165172
Manuputty, A.E.W. 2002. Karang Lunak (Soft Coral) Perairan Indonesia. Lembaga Ilmu Pengetahuan Indonesia. Pusat Penelitian Oseanografi. Jakarta.
Margono, W. 2009. Perkembangan Dan Pertumbuhan Karang Jenis
Lobophyllia hemprichii Yang Difragmentasikan di Pulau Pramuka, Kepulauan Seribu, Jakarta. Skripsi Fakultas Perikanan dan Ilmu Kelautan.
Michalek-Wagner, K. dan B. L. Willis. 2001. Impacts of bleaching on the soft coral Lobophytum compactum. I. Fecundity, fertilization and off spring viability. Coral Reefs. 19: 231–239.
Nybakken, J.W., 1992. Biologi Laut : Suatu Pendekatan Ekologi. Terjemahan oleh Eidman, M., D. G. Bengen, Koesoebiono, M. Hutomo dan Sukristijono. PT. Gramedia , Jakarta.
Setyawan , E, 2008 . Perkembangan Gamet Karang Lunak Sinularia dura Hasil Fragmentasi Di Pulau Pramuka, Kepulauan Seribu, DKI Jakarta. Skripsi Fakultas Perikanan dan Ilmu Kelautan IPB.
Sukarno, M., M. Hutomo, K. Moosa dan P. Darsono.1981. Terumbu Karang di Indonesia: Sumberdaya, Permasalahan dan Pengelolaannya. Proyek Penelitian Potensi Sumberdaya Alam Indonesia. LON – LIPI. Jakarta. Supriharyono. 2000. Pengelolaan Ekosistem Terumbu Karang. Penerbit