KERAGAAN BEBERAPA GENOTIPE
CABAI (Capsicum annuum L.) DAN KETAHANANNYA
TERHADAP ANTRAKNOSA, HAWAR PHYTOPHTHORA,
DAN LAYU BAKTERI SERTA PARAMETER GENETIKNYA
NURWANITA EKASARI PUTRI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Keragaan Beberapa Genotipe Cabai (Capsicum annuum L.) dan Ketahanannya terhadap Antraknosa, Hawar
Phytophthora, dan Layu Bakteri serta Parameter Genetiknya adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2010
Nurwanita Ekasari Putri
ABSTRACT
NURWANITA EKASARI PUTRI. Performance of Several Chilli (Capsicum annuum L.) Genotypes and Its Resistance to Anthracnose, Phytophthora Blight, Bacterial Wilt, and Their Genetic Parameters. Under direction of SRIANI SUJIPRIHATI, MUHAMAD SYUKUR, and WIDODO.
The aims of this research are to estimate genetic parameters to yield character and their resistance to anthracnose, phytophthora blight, and bacterial wilt. This research was conducted from June 2008 until November 2009 and consisted of three stages i.e. yield evaluation, resistance evaluation to anthracnose, phytophthora blight, and bacterial wilt, and genetic parameter estimation using diallel analysis. The characters being observed for yield evaluation were fruit weight per plant, weight per fruit, fruit length and fruit diameter. Genotype IPBC7 has the highest fruit weight per plant. Genotype IPBC28 has the highest weight per fruit. Genotypes with longest fruit were IPBC2, IPBC28, IPBC50, IPBC51, and IPBC64. Genotype IPBC5 has the biggest fruit diameter. The character being observed for resistance evaluation was disease incidence (DI). Genotypes IPBC15 and IPBC131 were resistant and moderately resistant to C. acutatum PYK04 isolate, respectively. Genotypes IPBC12, IPBC13, IPBC15, IPBC128 and IPBC130 were resistant while IPBC9, IPBC14, and IPBC19 were susceptible to
P. capsici TG01 isolate. IPBC19 and IPBC127 genotypes were slightly susceptible; IPBC12, IPBC15, IPBC125, and IPBC126 were slightly resistant genotypes; and other genotypes were resistant to R. solanacearum CHG7 isolate. Six genotypes were selected based on yield, disease resistance, and easy to cross, i.e. IPBC2, IPBC9, IPBC10, IPBC14, IPBC15, and IPBC20. These genotypes were used for diallel analysis in order to obtain genetic parameters estimation. Fruit length and phytophthora blight resistance were affected by inter-gene interaction (non allelic) except for fruit weight per plant, weight per fruit, and resistance to anthracnose and bacterial wilt. All observed-characters are controlled by partially-dominant gene action except for phytophthora blight resistance which is controlled by over-dominant gene action. Both broad and narrow-sense heritabilities are high for all characters, except for narrow-sense heritability of phytophthora resistance. Narrow heritability of phytophthora resistance is very low because there is no additive variance to control it. General combining ability and specific combining ability play important role to all observed-characters except to phytophthora blight resistance. There were no maternal effects to fruit weight and resistance to anthracnose. Additive variance is bigger than dominance variance for fruit weight per plant, weight per fruit, fruit length, and resistance to anthracnose and bacterial wilt. Thus, chilli breeding program is more directed to develop open pollinated variety. On the other hand, phytophthora blight resistance is more directed to develop hybrid variety. Based on both broad and narrow-sense heritabilities, selection can be done in early generation.
RINGKASAN
NURWANITA EKASARI PUTRI. Keragaan Beberapa Genotipe Cabai (Capsicum annuum L.) dan Ketahanannya terhadap Antraknosa, Hawar
Phytophthora, dan Layu Bakteri serta Parameter Genetiknya. Dibimbing oleh SRIANI SUJIPRIHATI, MUHAMAD SYUKUR, dan WIDODO.
Informasi genetik sangat diperlukan dalam merakit varietas cabai yang berdaya hasil tinggi dan tahan terhadap penyakit (antraknosa, hawar phytophthora,
dan layu bakteri). Untuk mendapatkan informasi genetik ini maka dilakukan penelitian dengan tiga kegiatan, yaitu 1) evaluasi daya hasil; 2) evaluasi ketahanan terhadap antraknosa, hawar phytophthora, dan layu bakteri; 3) pendugaan parameter genetik menggunakan analisis dialel. Penelitian dilakukan Juni 2008 – November 2009 di lapangan dan laboratorium.
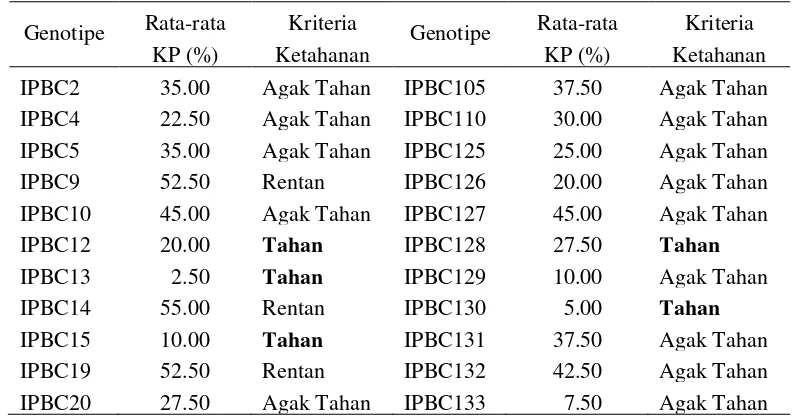
Kegiatan evaluasi daya hasil menggunakan 28 genotipe cabai koleksi Bagian Genetika dan Pemuliaan Tanaman, IPB. Kegiatan ini bertujuan untuk mendapatkan informasi mengenai daya hasil dari karakter yang diamati, yaitu bobot buah per tanaman, bobot per buah, panjang buah, dan diameter buah. IPBC7 memiliki bobot buah per tanaman paling besar. IPBC28 merupakan genotipe yang memiliki bobot per buah yang paling besar. Buah terpanjang dimiliki oleh IPBC2, IPBC28, IPBC50, IPBC51, dan IPBC64. Diameter buah terbesar adalah IPBC5. Keragaman genetiknya luas pada semua karakter yang diamati kecuali diameter buah sehingga karakter diameter buah tidak diikutsertakan dalam pendugaan parameter genetik.
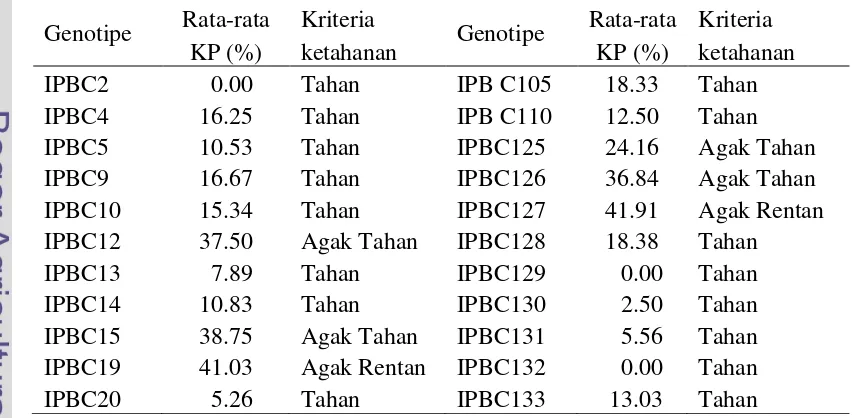
Evaluasi ketahanan pada 25 genotipe cabai terhadap antraknosa menggunakan C. acutatum isolat PYK04. IPBC15 dan IPBC131 berturut-turut merupakan genotipe tahan dan moderat sedangkan genotipe lainnya adalah rentan. Ketahanan 22 genotipe cabai terhadap hawar phytophthora menggunakan
P. capsici isolat TG01. IPBC12, IPBC13, IPBC15, IPBC128, IPBC130 merupakan genotipe tahan sedangkan IPBC9, IPB14, dan IPBC19 merupakan genotipe rentan dan sisanya adalah genotipe agak tahan. Pengujian ketahanan 22 genotipe cabai terhadap layu bakteri menggunakan R. solanacearum isolat CHG7. IPBC19 dan IPBC127 adalah genotipe agak rentan. IPBC12, IPBC15, IPB125, dan IPBC126 adalah genotipe agak tahan dan genotipe lainnya tergolong tahan.
Enam genotipe dipilih berdasarkan daya hasilnya, ketahanan terhadap penyakit (antraknosa, hawar phytophthora, layu bakteri) serta kemudahan dilakukan persilangan, yaitu IPBC2, IPBC9, IPBC10, IPBC14, IPBC15, dan IPBC20. Genotipe ini digunakan sebagai tetua dalam persilangan dialel untuk menduga parameter genetiknya. Keragaman genetik pada keenam genotipe yang dipilih adalah luas pada karakter bobot buah per tanaman, bobot per buah, dan panjang buah, namun ketahanan terhadap penyakit yang diamati keragamannya sempit. Karakter panjang buah dan ketahanan terhadap hawar phytophthora
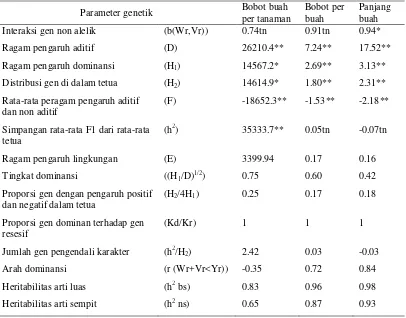
buah, dan panjang buah masing-masing adalah tiga, satu, dan satu kelompok gen. Ketahanan terhadap antraknosa dan hawar phytophthora dikendalikan masing-masing oleh satu dan empat gen.
Ragam DGU dan DGK berperan pada semua karakter yang diamati kecuali ketahanan terhadap hawar phytophthora hanya dipengaruhi oleh ragam DGK. Ragam DGU merupakan indikasi adanya ragam aditif dan ragam DGK merupakan manifestasi ragam dominan. Pengaruh maternal tidak ada pada bobot buah per tanaman dan ketahanan terhadap antraknosa.
Ragam aditif yang lebih besar dibandingkan ragam dominan pada karakter bobot buah per tanaman, bobot per buah, panjang buah, ketahanan terhadap penyakit (antraknosa dan layu bakteri) maka pemuliaan cabai diarahkan membentuk varietas open pollinated sedangkan ketahanan terhadap P. capsici
lebih diarahkan membentuk hibrida. Heritabilitas arti luas dan sempit semua karakter tinggi kecuali ketahanan terhadap hawar phytophthora yang memiliki heritabilitas arti sempit yang sangat kecil akibat tidak adanya ragam aditif yang berperan. Berdasarkan nilai heritabilitas arti luas dan sempit, karakter-karakter yang diamati dapat diseleksi pada generasi awal.
© Hak Cipta milik IPB, tahun 2010
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
KERAGAAN BEBERAPA GENOTIPE
CABAI (Capsicum annuum L.) DAN KETAHANANNYA
TERHADAP ANTRAKNOSA, HAWAR PHYTOPHTHORA,
DAN LAYU BAKTERI SERTA PARAMETER GENETIKNYA
NURWANITA EKASARI PUTRI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Keragaan Beberapa Genotipe Cabai (Capsicum annuum L.) dan Ketahanannya terhadap Antraknosa, Hawar Phytophthora, dan Layu Bakteri serta Parameter Genetiknya
Nama : Nurwanita Ekasari Putri
NIM : A253070061
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Sriani Sujiprihati, M.S. Ketua
Dr. Muhamad Syukur, SP.M.Si. Dr. Ir. Widodo, M.S. Anggota Anggota
Diketahui
Ketua Mayor Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
PRAKATA
Alhamdulillahi Rabbil ‘alamin atas rahmat dan hidayah Allah SWT yang memberikan kemudahan dan kelapangan sehingga penulis dapat menyelesaikan
tesis yang berjudul Keragaan Beberapa Genotipe Cabai (Capsicum annuum L.)
dan Ketahanannya terhadap Antraknosa, Hawar Phytophthora, dan Layu Bakteri
serta Parameter Genetiknya. Tesis ini merupakan tugas akhir dalam menyelesaikan
Program Magister pada Sekolah Pascasarjana Institut Pertanian Bogor.
Ucapan terima kasih dan penghargaan yang setinggi-tingginya penulis
sampaikan kepada:
1. Prof. Dr. Ir. Sriani Sujiprihati, MS., Dr. Muhamad Syukur, SP.MSi., dan
Dr. Ir. Widodo, MS. selaku komisi pembimbing yang telah memberikan
arahan dan bimbingan sejak perencanaan dan penyusunan penelitian hingga
penyelesaian tulisan.
2. Dr. Ir. Yudiwanti Wahyu Endro Kusumo, MS. sebagai penguji luar komisi
pembimbing atas`masukan dan saran untuk penyempurnaan tulisan ini.
3. Dr. Ir. Trikoesoemaningtyas, MSc. selaku Ketua Mayor Pemuliaan dan
Bioteknologi Tanaman SPs IPB yang selalu memberi arahan dalam
pelaksanaan studi selama perkuliahan hingga pelaksanaan tugas akhir.
4. Dr. Rahmi Yunianti, SP.MSi yang selalu meluangkan waktu untuk berdiskusi
dan memberikan masukan serta motivasi selama perkuliahan hingga
penyelesaian tugas akhir.
5. Rektor Universitas Andalas, Dekan Fakultas Pertanian, dan Ketua Jurusan
Budidaya Pertanian yang telah memberikan izin belajar.
6. Dirjen DIKTI yang telah memberikan beasiswa BPPS.
7. Prof. Dr. Ir. Sriani Sujiprihati, MS. sebagai Ketua Tim Hibah Pascasarjana
Tahun 2009 dan Dr. Muhamad Syukur, SP.MSi. sebagai Ketua Tim Hibah
KKP3T Deptan Tahun 2009 yang telah membiayai penelitian ini.
8. Ketua Tim Pemuliaan Cabai Bagian Genetika dan Pemuliaan Tanaman
Departemen Agronomi dan Hortikultura IPB atas bantuan bahan genetik dan
9. Dr. Ir. Widodo, MS atas sumbangan isolat Colletotrichum acutatum dan
Phytophthora capsici.
10. Kepala Laboratorium Bakteriologi Tumbuhan Departemen Proteksi Tanaman
yang memberikan ijin untuk menggunakan isolat Ralstonia solanacearum.
11. Ady Daryanto SP, Tiara Yudilastari SP, Abdul Hakim, Endah SP, Purwati SP,
Swisci Margareth SP, Ir. Dwi Wahyuni Ganefianti MS, Undang SP, Siti
Marwiyah SP, Abdulllah bin Arif SP, Avicenna, Rohim Firdaus SP, Esa Hari
Wibowo SP, Andra, Warno, dan Pak Darwa atas kebersamaan bantuan dan
kerjasama serta kecerian selama penelitian di lapang dan laboratorium.
12. Ir. Gusti Adhi, MS yang telah membuat dan memberi ijin penggunaan listing
SAS dalam analisis dialel.
13. Teman-teman PBT 2007: Amin Nur, Hairinsyah, Rokhana Faizah, Yusie
Arisanti, Alfin Widiastuti, Heni Safitri, Erni Suminar, Fifin N. Khairunisa,
Isnaini dan Siti Nurrohmah atas kebersamaan dan saling memotivasi sejak
awal kuliah hingga penyelesaian tugas akhir.
14. Adik-adik di Blobo atas keceriaan dan kebersamaan terutama Nurul Fuadi,
Risa, dan Indah yang juga turut membantu panen di lapang.
15. Ayahanda Ir. Bustimar dan Ibunda Andi Rasnah serta adik-adikku Puput dan
Yuyuk yang tiada hentinya selalu mendoakan dan memberikan kasih sayang,
pengorbanan, dan dukungan serta motivasi.
16. Suami tercinta Ardian Amien atas doa, kasih sayang, pengorbanan dan
pengertiannya.
17. Keluarga besar Ardan Amien (alm.) atas doa, dukungan dan kasih sayang.
18. Para sahabat dan semua pihak yang telah banyak membantu penulis.
Penulis berharap semoga tulisan ini dapat dimanfaatkan guna memperkaya
khazanah pengembangan ilmu pengetahuan.
Bogor, Februari 2010
RIWAYAT HIDUP
Penulis dilahirkan dilahirkan di Padang pada tanggal 1 Agustus 1978
sebagai putri pertama dari ayah Ir. Bustimar dan ibu Andi Rasnah M. Penulis
menikah dengan Ardian Amien SH pada tanggal 2 Agustus 2008.
Penulis menyelesaikan pendidikan sarjana di Program Studi Pemuliaan
Tanaman Jurusan Budidaya Pertanian Fakultas Pertanian IPB pada tahun 2001.
Pada tahun 2005, penulis diterima sebagai staf pengajar di Program Studi
Pemuliaan Tanaman Jurusan Budidaya Pertanian Faperta Universitas Andalas.
Penulis berkesempatan melanjutkan pendidikan Program Magister di Sekolah
Pascasarjana IPB pada Mayor Pemuliaan dan Bioteknologi Tanaman dengan
beasiswa BPPS pada tahun 2007.
Artikel yang berjudul Analisis Dialel pada Komponen Daya Hasil Cabai
telah disajikan pada Seminar Nasional Perhimpinan Ilmu Pemuliaan Indonesia
pada Tanggal 18-19 November 2009 di Cimanggu, Bogor. Artikel ini merupakan
DAFTAR ISI
Halaman
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN ... xv
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 4
Kerangka Pemikiran dan Pengajuan Hipotesis ... 4
Ruang lingkup penelitian ... 5
TINJAUAN PUSTAKA ... 8
Klasifikasi Cabai ... 8
Botani Cabai ... 8
Penyakit Antraknosa ... 9
Penyakit Hawar Phytophthora ... 11
Penyakit Layu Bakteri ... 13
Perakitan Varietas Tahan ... 16
Metode Silang Dialel ... 17
Heterosis ... 20
Heritabilitas ... 21
BAHAN METODE ... 23
Percobaan 1: Karakterisasi Daya Hasil ... 23
Percobaan 2: Pengujian Ketahanan Terhadap Antraknosa, Hawar Phytophthora dan Layu Bakteri ... 24
Percobaan 3: Pendugaan Parameter Genetik Menggunakan Analisis Dialel ... 29
HASIL DAN PEMBAHASAN ... 34
Karakterisasi Daya Hasil ... 34
Evaluasi Ketahanan Terhadap Penyakit Antraknosa, Hawar Phytophthora, dan Layu Bakteri ... 38
Analisis Silang Dialel ... 44
PEMBAHASAN UMUM ... 72
SIMPULAN DAN SARAN ... 81
Simpulan ... 81
Saran ... 82
DAFTAR PUSTAKA ... 83
LAMPIRAN ... 89
DAFTAR TABEL
Halaman 1 Pengelompokan ras penyakit Ralstonia solanacearum... 142 Analisis ragam rancangan acak kelompok ... 23
4 Kriteria ketahanan cabai terhadap hawar Phytophthora ... 27
5 Kriteria ketahanan cabai terhadap layu bakteri ... 29
6 Populasi full diallel dalam menduga parameter genetik untuk daya hasil 30 7 Populasi full diallel dalam menduga parameter genetik untuk ketahanan terhadap antraknosa ... 32
8 Populasi half diallel dalam menduga parameter genetik untuk ketahanan terhadap penyakit hawar phytophthora dan layu bakteri ... 32
9 Nilai kuadrat tengah beberapa sifat kuantitatif pada genotipe cabai yang diamati ... 34
10 Nilai rata-rata beberapa sifat kuantitatif pada genotipe yang diamati... 35
11 Parameter genetik pada bobot buah per tanaman, bobot per buah, panjang buah, dan diameter buah ... 36
12 Korelasi pada beberapa karakter cabai yang diamati ... 37
13 Ketahanan beberapa genotipe cabai terhadap penyakit antraknosa yang disebabkan oleh C. acutatum isolat PYK04 ... 40
14 Ketahanan beberapa genotipe cabai terhadap penyakit hawar Phytophthora yang disebabkan oleh Phytophthora capsici LEONIAN isolat TG01 ... 42
15 Ketahanan beberapa genotipe cabai terhadap penyakit layu bakteri yang disebabkan oleh Ralstonia solanacearum ... 43
16 Parameter genetik ketahanan terhadap antraknosa, hawar phytophthora, dan layu bakteri ... 44
17 Genotipe-genotipe yang dijadikan tetua dalam persilangan dialel ... 45
18 Parameter genetik dari enam genotipe yang dipilih ... 45
19 Populasi full diallel dengan menggunakan enam genotipe terpilih ... 46
20 Nilai kuadrat tengah beberapa sifat kuantitatif cabai ... 47
21 Pendugaan parameter genetik pada beberapa sifat kuantittaif cabai ... 48
22 Rata-rata tetua dan F1 pada karakter yang diamati ... 49
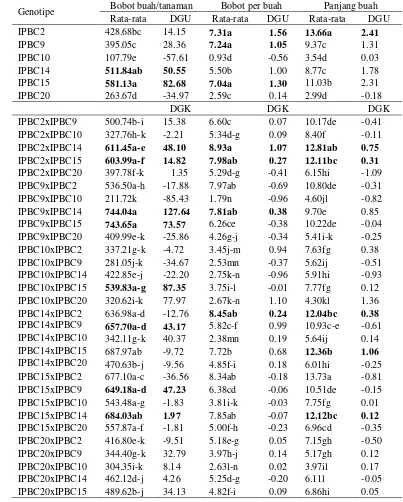
23 Analisis ragam DGU, DGK dan resiprokal pada karakter bobot buah per tanaman, bobot per buah, dan panjang buah ... 54
24 Nilai rata-rata, DGU, dan DGK pada bobot buah per tanaman, bobot per buah, dan panjang buah ... 55
25 Nilai heterosis dan heterobeltiosis pada karakter cabai yang diamati... 58
26 Kuadrat tengah kejadian penyakit antraknosa... 59
27 Populasi half diallel dalam menduga parameter genetik untuk ketahanan terhadaphawar phytophthora ... 60
28 Kuadrat tengah ketahanan terhadap Phytophthora capsici ... 60
29 Populasi half diallel dalam menduga parameter genetik untuk ketahanan terhadap layu bakteri ... 61
30 Nilai kuadrat tengah ketahanan terhadap layu bakteri ... 61
31 Pendugaan parameter genetik ketahanan cabai terhadap antraknosa (C. acutatum) isolat PYK04 dan hawar phytophthora ... 62
antraknosa ... 66
33 Nilai rata-rata, DGU, dan DGK serta kriteria ketahanan cabai terhadap antraknosa ... 67
34 Analisis DGU dan DGK pada ketahanan terhadap P. capsici ... 68
35 Nilai rata-rata, DGU, dan DGK serta kriteria ketahanan terhadap Hawar phytophthora... 68
36 Kriteria ketahanan populasi half diallel terhadap layu bakteri ... 70
37 Komponen analisis ragam analisis silang dialel... 94
38 Pendugaan ragam dan peragam ... 94
39 Komponen ragam untuk daya gabung pada Metode I Griffing ... 98
40 Komponen ragam untuk daya gabung pada Metode II Griffing ... 99
DAFTAR GAMBAR
Halaman 1 Bagan alir penelitian ... 72 Inokulum C. acutatum isolat PYK04 ... 25
3 Tahapan pelaksanaan inokulasi C. acutatum isolat PYK04 ... 25
4 Inokulum P. capsici isolat TG01 ... 26
5 Inokulasi P. capsici pada tanaman berumur 28 hari ... 27
6 Persiapan dan inokulasi R. solanacearum penyebab layu bakteri ... 29
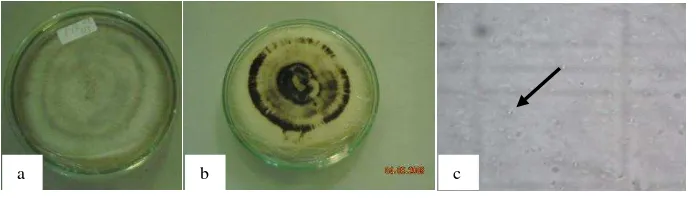
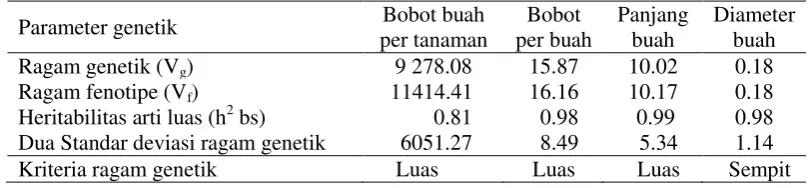
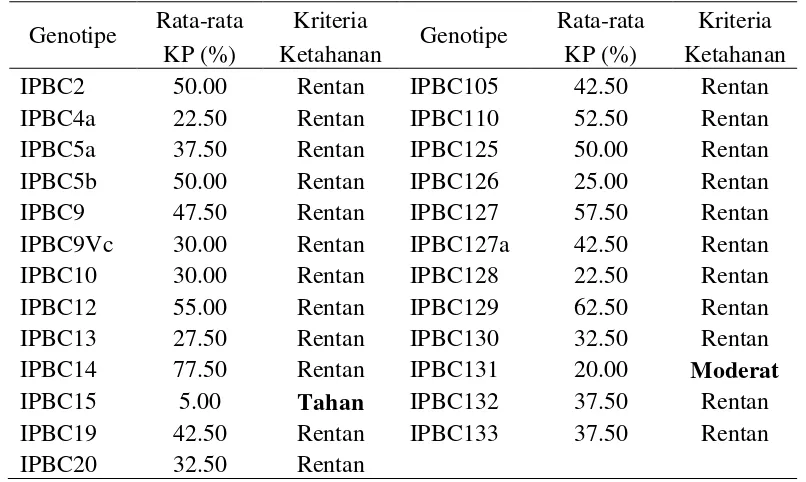
7 Serangan antraknosa yang disebabkan C. acutatum pada buah ... 39
8 Serangan hawar phytophthora yang disebabkan P. capsici pada fase bibit ... 40
9 Serangan layu bakteri yang disebabkan oleh R. solanacearum ... 42
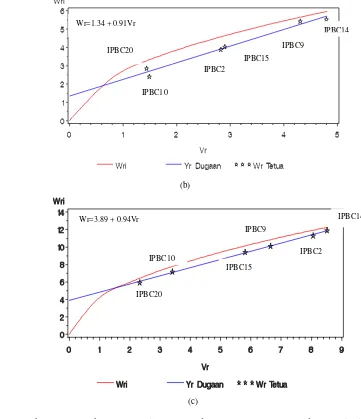
10 Hubungan varians (Vr) dan peragam (Wr) pada populasi F1 silang dialel.. ... 52
11 Hubungan varians (Vr) dan peragam (Wr) pada populasi F1 silang dialel pada ketahanannya terhadap antraknosa ... 64
12 Hubungan varians (Vr) dan peragam (Wr) pada populasi F1 silang dialel pada ketahanannya terhadap hawar phytophthora ... 65
13 Bulu batang pada cabai ... 91
15 Bentuk buah ... 92
16 Bentuk pangkal buah ... 93
17 Bentuk ujung buah ... 93
DAFTAR LAMPIRAN
... Halaman 1 Genotipe yang digunakan dalam karakterisasi daya hasil ... 892 Genotipe yang digunakan dalam pengujian ketahanan terhadap antraknosa, hawar phytophthora, dan layu bakteri ... 90
3 ... Descriptor for Capsicum ... 91
4 Analisis menggunakan Metode Hayman ... 94
5 Analisis menggunakan Metode Griffing ... 98
6 Karakerisasi beberapa genotipe cabai berdasarkan karakter kualitatif ... 101
7 Hasil sidik ragam ketahanan terhadap penyakit antraknosa, hawar ... Phytophthora, dan layu bakteri ... 103
PENDAHULUAN
Latar Belakang
Tanaman cabai (Capsicum sp.) merupakan salah satu komoditas hortikultura
yang banyak digemari masyarakat. Salah satu spesies cabai yang banyak
dibudidayakan adalah cabai merah (Capsicum annuum L.). Selain dapat
dikonsumsi segar, cabai dapat dikonsumsi kering sebagai bumbu masakan dan
juga sebagai bahan baku industri. Rubatzky dan Yamaguchi (1997) menyatakan
bahwa cabai juga digunakan dalam industri pangan, pakan unggas, dan farmasi.
Bosland dan Votava (2000) melaporkan bahwa cabai mengandung zat-zat gizi
antara lain protein, lemak, karbohidrat, kalsium, fosfor, besi, vitamin (A, C, dan
B1), dan senyawa alkaloid seperti capsaicin, plavonoid, dan minyak esensial.
Areal pertanaman cabai pada tahun 2007 adalah 187 236 ha dan mengalami
penurunan menjadi 107 326 ha pada tahun 2008, namun produktivitas cabai naik
dari 5.7 ton/ha menjadi 6.3 ton/ha (BPS 2009). Kenaikan produktivitas tersebut
masih jauh dari potensi produksinya. Menurut Duriat (1996) cabai memiliki
potensi produksi 12-20 ton/ha.
Rendahnya produktivitas per tanaman cabai dipengaruhi oleh faktor biotik
dan abiotik. Salah satu faktor biotik yang sangat mempengaruhi hasil cabai adalah
hama dan penyakit. Bosland dan Votava (2000) menegaskan bahwa salah satu
penyebab tidak tercapainya potensi produksi cabai adalah gangguan hama dan
penyakit. Suryaningsih et al. (1996) menyatakan bahwa penyakit yang paling
dominan menyerang pertanaman cabai adalah antraknosa (Colletotrichum sp),
hawar (Phytophthora capsici), layu bakteri (Ralstonia solanacearum), dan virus.
Penyakit antraknosa di Indonesia dapat menurunkan hasil sampai 75%.
Penyakit yang disebabkan oleh cendawan Colletotrichum sp menyerang buah baik
yang baru terbentuk maupun yang sudah matang sehingga menimbulkan kerugian
yang cukup besar. Patogen ini menyerang tanaman cabai di dataran tinggi dan
dataran rendah (Kusandriani & Permadi 1996). Cendawan C. acutatum
merupakan salah satu cendawan penyebab penyakit antraknosa di Indonesia
Penyakit hawar yang disebabkan oleh patogen P. capsici menyerang semua
fase dan bagian tanaman. Serangan pada pangkal batang ditandai dengan busuk
batang berwarna coklat kehitaman, tanaman layu tanpa terjadi penguningan daun
terlebih dahulu. Patogen ini menyebabkan kerugian yang sangat besar dan
bergantung terutama pada suhu dan kelembapan, intensif tidaknya tindakan
penyemprotan dengan fungisida, umur tanaman saat diserang, serta tingkat
ketahanan yang dimiliki oleh varietas cabai yang ditanam (Yunianti 2007).
Penyakit layu bakteri disebabkan oleh bakteri R. solanacearum yang dahulu
bernama Pseudomonas solanacearum (Machmud 1993). Patogen ini memiliki
kisaran inang dan daerah sebaran yang luas dan kemampuan bertahan hidup dalam
tanaman yang luar biasa di dalam tanah dan rizosfer tanaman yang bukan
inangnya, dan variabilitasnya sangat tinggi serta daya rusak yang besar (Machmud
et al. 1999). Penyakit layu bakteri dapat menyebabkan kematian tanaman dan
kegagalan panen sehingga menimbulkan penurunan hasil yang cukup besar
(Semangun 2000). Pada tanaman kentang dan tomat, penyakit layu bakteri dapat
menurunkan produksi berturut-turut sebesar 75% dan tomat 100% (Persley et al.
1985).
Pengendalian patogen penyebab antraknosa, hawar phytophthora, dan layu
bakteri tidaklah mudah karena patogen ini sering ditemukan bersama di lahan
petani. Varietas yang tahan terhadap suatu penyakit kadangkala ketahanannya
dapat dipatahkan oleh munculnya ras baru dari suatu patogen. Bahkan tanaman
tidak dapat bertahan dan berproduksi baik akibat banyaknya serangan penyakit
yang datang sekaligus. Pengendalian secara kimia dapat membahayakan petani,
masyarakat sebagai konsumen dan lingkungan serta biaya yang mahal. Alternatif
yang dapat dilakukan adalah penggunaan varietas yang tahan.
Perakitan varietas cabai dapat ditempuh dengan menggabungkan beberapa
galur yang memiliki ketahanan terhadap penyakit antara lain tahan terhadap
antraknosa, hawar, dan layu bakteri. Bagian Genetika dan Pemuliaan Departemen
AGH IPB telah mempunyai bahan tanaman cabai yang memiliki karakter
ketahanan terhadap beberapa penyakit yang telah diuji oleh peneliti sebelumnya.
Syukur (2007) melaporkan bahwa diantara genotipe yang diuji hanya genotipe
sedangkan lainnya tergolong rentan. Hasil penelitian Yunianti (2007) menyatakan
bahwa genotipe IPBC2 merupakan genotipe yang tergolong agak tahan terhadap
Phytophthora ras 1. Yulianah (2007) melaporkan bahwa IPBC9 dan IPBC10
merupakan genotipe yang tahan terhadap layu bakteri.
Perbaikan produksi tanaman dapat ditempuh melalui kegiatan pemuliaan
tanaman untuk menghasilkan genotipe-genotipe cabai berproduksi tinggi. Namun,
usaha tersebut dihadapkan kepada masalah dalam memilih tetua-tetua yang
mempunyai daya gabung yang tinggi. Genotipe yang akan digunakan sebagai tetua
dalam program pemuliaan didasarkan atas penampilan keturunan yang dihasilkan
dari persilangan tertentu. Untuk itu perlu diperoleh informasi kemampuan tetua
untuk bergabung dengan tetua lainnya dalam membentuk turunannya dan metode
yang umum digunakan adalah metode silang dialel.
Metode silang dialel merupakan suatu metode yang menggunakan semua
kombinasi persilangan yang mungkin dari beberapa tetua yang digunakan.
Menurut Singh dan Chaudhary (1979), metode ini memberikan informasi daya
gabung umum (general combining ability) dan daya gabung khusus (spesific
combining ability) serta dapat menduga parameter genetik. Poehlman dan Sleeper
(1990) menyatakan bahwa daya gabung umum adalah kemampuan individu tetua
untuk menghasilkan keturunan yang unggul untuk suatu karakter tertentu yang
disilangkan dengan sejumlah tetua lainnya. Daya gabung khusus adalah
kemampuan individu tetua untuk menghasilkan turunan yang unggul jika
disilangkan dalam kombinasi yang spesifik dengan tetua lainnya. Menurut Darlina
et al. (1992) daya gabung sangat diperlukan untuk mengindentifikasi kombinasi
tetua yang akan menghasilkan keturunan yang berpotensi hasil tinggi. Baihaki
(2000) menyatakan pengujian daya gabung dapat dilakukan dengan analisis silang
dialel. Analisis ini juga berguna dalam menduga ragam genetik dan heritabilitas
serta memberikan informasi mengenai aksi gen yang terdapat dalam suatu
populasi.
Keuntungan menggunakan analisis dialel adalah dapat menduga parameter
genetik pada populasi F1 tanpa harus membentuk populasi F2, BCP1 dan BCP2
sangat sistematik dalam mengevaluasi genetik pada generasi awal sehingga
diketahui persilangan yang berpotensi untuk diseleksi (Johnson 1963).
Dalam penelitian ini dilakukan evaluasi terhadap populasi cabai yang
merupakan koleksi dari Bagian Genetika dan Pemuliaan Departemen AGH IPB
guna memperoleh informasi daya hasil dari berbagai genotipe. Genotipe-genotipe
yang terpilih diuji ulang ketahanannya terhadap penyakit antraknosa, hawar
phytophthora dan layu bakteri. Selanjutnya, tetua terpilih dibuat persilangan
dialelnya. Dengan analisis dialel dapat diperoleh informasi yang dapat digunakan
dalam menentukan metode seleksi yang akurat dalam merakit varietas tahan dan
berproduksi cukup tinggi.
Tujuan Penelitian Penelitian ini bertujuan untuk :
1. Memperoleh informasi tentang karakter daya hasil beberapa genotipe cabai
koleksi Bagian Genetika dan Pemuliaan Departemen AGH IPB.
2. Menguji ulang ketahanan terhadap penyakit antraknosa, hawar phytophthora
dan layu bakteri pada beberapa genotipe cabai.
3. Mendapatkan informasi parameter genetik beberapa genotipe cabai untuk
karakter produksi dan ketahanannya terhadap penyakit antraknosa, hawar
phytophthora, atau layu bakteri.
4. Mendapatkan informasi daya gabung beberapa genotipe cabai untuk karakter
produksi dan ketahanannya terhadap penyakit antraknosa, hawar
phytophthora, atau layu bakteri.
Kerangka Pemikiran dan Pengajuan Hipotesis
Perbaikan produksi cabai baik secara kualitas maupun kuantitas dapat
dilakukan dengan kegiatan pemuliaan tanaman. Kegiatan ini memerlukan
pengetahuan mengenai informasi genetik yang bekerja mengendalikan suatu
karakter. Informasi genetik yang diperlukan dapat dilakukan salah satunya dengan
analisis dialel. Analisis ini dapat menduga parameter genetik pada populasi F1
tanpa harus membentuk F2 dan backcrossnya. Menurut Johnson (1963), analisis
merupakan pendekatan sistematik dan secara analitik merupakan evaluasi genetik
menyeluruh yang berguna dalam mengidentifikasi persilangan pada awal generasi.
Penelitian mengenai ketahanan terhadap penyakit antraknosa, hawar
phytophthora, dan layu bakteri telah banyak dilaporkan oleh peneliti sebelumnya.
Namun, informasi mengenai kendali genetik pada masing-masing penyakit
berbeda-beda. Hal ini mungkin terjadi karena populasi dan metode yang digunakan
pun berbeda. Oleh karena itu, untuk mengintegrasikan gen-gen ketahanan terhadap
ketiga penyakit kepada genotipe yang berdaya hasil tinggi membutuhkan
informasi genetik dari populasi yang kita gunakan sehingga dapat diketahui
metode yang efektif dan efisien dalam merakit varietas berdaya hasil tinggi dan
tahan terhadap beberapa penyakit.
Berdasarkan serangkaian kegiatan penelitian maka hipotesis yang diajukan
adalah sebagai berikut:
1. Terdapat beberapa genotipe yang memiliki daya hasil yang tinggi.
2. Terdapat genotipe yang tahan, moderat dan rentan terhadap penyakit
antraknosa, hawar phytophthora, atau layu bakteri.
3. Terdapat satu atau beberapa genotipe yang mempunyai daya gabung yang
tinggi untuk produksi dan ketahanannya terhadap penyakit antraknosa, hawar
phytophthora, atau layu bakteri.
Ruang Lingkup Penelitian
Penelitian terdiri atas tiga percobaan, yaitu 1) karakterisasi daya hasil,
2) evaluasi ketahanan terhadap penyakit antraknosa, hawar phytophthora, dan layu
bakteri, dan 3) pendugaan parameter genetik menggunakan analisis dialel.
Pada percobaan pertama dilakukan karakterisasi daya hasil pada koleksi
plasma nutfah cabai milik Bagian Genetika dan Pemuliaan Tanaman Departemen
Agronomi dan Hortikultura IPB. Koleksi ini terdiri dari varietas lokal, galur-galur
hasil kegiatan pemuliaan, dan juga introduksi. Dari percobaan pertama akan
diperoleh informasi genotipe yang berdaya hasil tinggi, sedang, dan rendah.
Beberapa genotipe koleksi Bagian Genetika dan Pemuliaan Tanaman
Departemen Agronomi dan Hortikultura IPB telah diidentifikasi tahan terhadap
yang dilakukan oleh peneliti-peneliti sebelumnya. Pada percobaan kedua
dilakukan uji ulang ketahanan terhadap penyakit antraknosa, hawar phytophthora,
dan layu bakteri pada beberapa genotipe dari plasma nutfah cabai termasuk yang
belum diidentifikasi ketahanannya terhadap ketiga penyakit tersebut. Hasil
percobaan kedua akan diperoleh informasi genotipe-genotipe yang tahan, moderat,
dan rentan pada ketiga penyakit yang diamati.
Pada percobaan ketiga akan dilakukan persilangan dialel dengan
menggunakan enam genotipe sebagai tetua. Pemilihan enam genotipe ini
berdasarkan informasi daya hasil yang diperoleh dari percobaan pertama,
informasi ketahanannya terhadap antraknosa, hawar phytophthora, dan layu
bakteri yang diperoleh dari percobaan kedua dan kemudahannya dalam bersilang (’easy to cross’) dengan genotipe lain. Hasil dari percobaan ketiga ini akan memberikan informasi mengenai paramater genetik dan kemampuan daya gabung
dari genotipe yang digunakan. Keseluruhan kegiatan penelitian dirangkum pada
Gambar 1 Bagan alir penelitian. Plasma nutfah cabai
(koleksi dan introduksi)
Percobaan 1:
Karakterisasi daya hasil
Percobaan 2:
Uji ulang ketahanan terhadap penyakit:
Antraknosa
Hawar phytophthora
Layu Bakteri
Percobaan 3:
Pendugaan parameter genetik menggunakan analisis dialel
Informasi parameter genetik
Metode seleksi yang efektif dan efisien
Genotipe tahan terhadap antraknosa, hawar
phytophthora, dan layu bakteri
Genotipe terpilih Genotipe dengan
TINJUAUAN PUSTAKA
Klasifikasi Cabai
Cabai (Capsicum sp) merupakan tanaman herba tropika yang biasanya
ditanam sebagai tanaman tahunan (Bosland dan Votava 2000). Genus Capsicum
termasuk ke dalam famili Solanaceae. Capsicum dikelompokan oleh ahli
taksonomi menjadi lima spesies yaitu C. annuum L., C. frutescens L., C.
chinense, C. pubescens, dan C. baccatum yang berasal dari pusat penyebaran yang
berbeda. C. annuum L. berasal dari Meksiko, dua spesies berikutnya berasal dari
Guatemala dan dua spesies terakhir berturut-turut dari Bolivia dan Peru (Greenleaf
1986).
Spesies cabai liar yang pernah diidentifikasi adalah sebagai berikut:
(1) C. praetermisum Heiser & Smith yang menyebar di Brasil Selatan; (2)
C. chacoense A.T. Hunz yang menyebar di Argentina Utara, Bolivia dan
Paraguay; (3) C. galapagoense A.T. Hunz menyebar di Pulau Galapagos; (4)
C. cardenasii Heiser & Smith menyebar di Bolivia; (5) C. eximiun A.T Hunz
menyebar di Argentina Utara dan Bolovia; (6) C. tovarii Eshbaugh, Smith,
Nickrent menyebar di Andes, Peru (Greenleaf 1986).
Tanaman cabai merupakan tanaman menyerbuk sendiri namun dapat
terjadi persilangan secara alami sekitar 7.6-36.8% dengan bantuan lebah.
Persilangan antar spesies mungkin terjadi. Namun Greenleaf (1986) menerangkan
bahwa terdapat beberapa spesies yang sulit disilangkan misalnya C. annuum L. x
C. frutescens, dan C. annuum x C. pubescens.
Botani Cabai
Seperti famili Solanaceae lainnya, tanaman cabai memiliki bunga
berbentuk terompet. Umumnya, bunga tunggal dan tumbuh pada ujung ruas dan
merupakan bunga sempurna (hermaprodit). Mahkota bunga berwarna putih atau
ungu tergantung varietasnya, helaian mahkota bunga lima atau enam helai. Setiap
bunga memiliki satu putik (stigma) dengan kepala berbentuk bulat dan berwarna
kuning kehijauan. Benang sari cabai berkisar 5-8 helai dengan kepala sari (anther)
Bunga pertama terbentuk pada umur 23-32 hari setelah tanam (HST) dan buah
pertama mulai terbentuk pada umur 29-40 HST. Sekitar 45 hari setelah pembuahan
buah yang dihasilkan sudah bisa dipanen. Struktur buah terdiri atas kulit, daging
buah, dan sebuah plasenta tempat melekatnya biji. Daging buah umumnya renyah
atau kadang-kadang lunak pada beberapa kultivar. Biji cabai berwarna kuning
jerami (Greenleaf 1986).
Daun tumbuh secara spiral pada batang utama (Kusandriani 1996b). Batang
yang berupa kayu memiliki percabangan yang banyak. Pembentukan kayu pada
batang utama mulai terbentuk pada umur 30 HST. Pada setiap ketiak daun tumbuh
tunas baru mulai 10 HST. Tipe percabangan tegak atau menyebar (Rubatzky dan
Yamaguchi 1997).
Akar tanaman cabai diawali dengan akar primer yang panjangnya 35-50
cm. Kemudian muncul akar lateral yang tumbuh ke samping yang panjangnya
mencapai 34-45 cm. Sistem perakaran cabai termasuk tipe dangkal (Rubatzky dan
Yamaguchi 1997).
Penyakit Antraknosa
Serangan penyakit antraknosa dapat mengurangi produksi cabai karena
patogennya menggugurkan buah baik yang masih hijau ataupun yang sudah
matang. Selain itu, benih yang dihasilkan dari tanaman yang terinfeksi antraknosa
dapat menularkannya melalui benih (seed borne). Patogen penyebab tular benih
biasanya berupa miselium atau spora yang dorman pada permukaan benih, atau
berupa miselium yang dorman pada embrio. Colletotrichum pada buah masuk ke
dalam ruang biji dan menginfeksi biji (Semangun 2000). Agrios (1997)
menerangkan bahwa cendawan ini dapat menyebabkan infeksi ringan pada daun
dan batang muda yang mungkin tidak terdeteksi namun cendawan ini mampu
bertahan dan berkembang biak pada tahapan tertentu hingga buah mulai masak
menjadi rentan terhadap infeksi.
Antraknosa disebabkan oleh cendawan dari genus Colletotrichum.
Kim et al. (1999) menggolongkan Colletotrichum menjadi 6 spesies utama, yaitu
Colletotrichum gloeosporioides, C. acutatum, C. dematium, C. capsici, dan C.
coccodes. Yoon (2003) melaporkan bahwa C. gloesporioides dan C. acutatum
Suryaningsih et al. (1996) menyatakan bahwa C. capsici dan C. gloeosporioides
adalah spesies yang paling banyak dijumpai di Indonesia. C. capsici banyak
menyerang pertanaman cabai di dataran tinggi dan C. gloeosporioides di dataran
rendah.
C. acutatum mempunyai miselium berwarna putih hingga abu-abu. Jika
dibalik, koloni berwarna orange hingga merah muda. Konidia berbentuk silindris
dengan ujung runcing, berukuran 15.1 (12.8-16.9) x 4.8 (4.0-5.7) μm. Suhu
optimal adalah 280C dengan rata-rata pertumbuhan 10.3 mm/hari (AVRDC 2003).
Gejala yang timbul akibat serangan antraknosa pada buah adalah
terdapatnya bercak coklat kehitaman pada permukaan buah, yang selanjutnya
menjadi busuk lunak. Pada bagian tengah bercak terdapat kumpulan titik-titik
hitam yang terdiri dari sekelompok seta dan konidium jamur. Serangan yang berat
dapat menyebabkan buah mengering dan keriput sehingga buah yang seharusnya
berwarna merah menjadi seperti jerami (Semangun 2000).
Suryaningsih et al. (1996) menyatakan infeksi cendawan Colletotrichum
bersifat laten dan bertahan dalam jaringan tanaman dalam bentuk aservulus.
Aservulus dapat bertahan dan tumbuh lama dalam biji kemudian miselium tumbuh
di luar kulit biji. Miselium dan aservulus dapat tumbuh ± 9 bulan dalam biji.
Varietas tahan masih dapat terserang penyakit antraknosa jika ditanam pada tanah
yang sudah terinfeksi cendawan ini.
Cendawan Colletotrichum dapat menyerang buah yang masih hijau
maupun yang sudah masak. Gejala yang ditimbulkan adalah bintik-bintik kecil
berwarna kehitaman dengan tepi berwarna kuning, membesar dan memanjang,
bagian tengah menjadi semakin gelap. Dalam keadaan lembab, cendawan
membentuk badan buah (aservulus) dalam lingkaran-lingkaran sepusat yang
membentuk massa spora (konidium) berwarna merah jambu. Penyakit masih
berkembang terus pada waktu buah cabai disimpan atau diangkut (Semangun
2000).
Colletotrichum merupakan penyakit tular biji dan dapat bertahan di dalam
tanah. Cendawan ini memiliki banyak inang (Suryaningsih et al. 1996). Serangan
penyakit antraknosa sangat bergantung pada kondisi lingkungan. Serangan paling
dan suhu di atas 300C (AVRDC 1998). Penanaman cabai yang terus menerus
dalam areal yang sama merupakan faktor yang memudahkan tersebarnya
cendawan penyebab antraknosa. Penyakit antraknosa akan berkembang sangat
cepat jika tidak dilakukan pengendalian yang efektif.
Penyakit antraknosa sulit dikendalikan karena patogennya bersifat laten
dan sistemik serta menyerang semua fase pertumbuhan tanaman (Sinaga et al.
1992). Pengendalian penyakit ini dengan fungisida kurang efisien dan bersifat
sementara (Wijaya 1991). Selain itu dapat menyebabkan pencemaran lingkungan,
berbahaya baik bagi petani maupun konsumen, biaya relatif mahal dan tekanan
seleksi dapat menimbulkan ras-ras patogen baru yang resisten (Suryaningsih dan
Suhardi 1993).
Penyakit Hawar Phytophthora
Penyakit hawar phytophthora pada pertanaman cabai disebabkan oleh
Phytophthora capsici Leonian (Kurt dan Emir 2004). P. capsici termasuk famili
Pythiaceae ordo Peronosporales, kelas Oomycetes (Alexopoulus dan Mims 1979).
Menurut Sinaga (2003) Phytophthora merupakan cendawan penghuni tanah (
soil-inhabitant fungi) yang menggunakan sebagian hidupnya dalam tanah dan bertahan
sebagai saprofit.
Siklus hidup P. capsici terdiri dari fase pertumbuhan aktif dalam jaringan
dan fase dormansi (Lamour dan Hausbeck 2001a). Fase dormansi berupa oospora
berdinding tebal yang diproduksi selama reproduksi seksual (Lamour dan
Hausbeck 2001b). Oospora membutuhkan periode dormansi sebelum berkecambah
yang akan menghasilkan Coenocytic mycelium, yang secara langsung menginfeksi
atau berdiferensiasi menjadi cadicous sporangia dalam kondisi sesuai. Sporangia
dapat menyebabkan infeksi secara langsung atau dimediasi air bebas dengan
melepaskan 20-40 zoospora (Lamour dan Hausbeck 2001a).
Zoospora sebagai inokulum bila sampai ke permukaan jaringan tanaman
maka inokulum tersebut akan menempel dengan bantuan protein atau glikoprotein
yang ada pada permukaan jaringan (Coffey dan Gees 1991). P. capsici yang telah
masuk ke dalam jaringan akan mengganggu aktivitas fotosintesis dengan
terjadinya disfungsi biosintesis C18:3 yang merupakan komponen utama asam
lemak pada membran tilakoid tanaman (Souli et al. 1989). Gangguan pada
komponen kloroplas tersebut akan menyebabkan penurunan kapasitas fotosintesis
inang (Yunianti 2007).
Cendawan P. capsici akan membentuk struktur hifa primer dalam sel inang
dan menyebar melalui ruang interseluler. Dari hifa primer akan dibentuk hifa
sekunder (haustorium) yang menembus ke dalam sel. Hifa selanjutnya akan
berkembang membentuk sporangiofor. Struktur tersebut muncul dari stomata
inang dan selanjutnya membentuk cabang-cabang pendek. Pada cabang-cabang
tersebut dibentuk sporangium. Cendawan mampu membentuk ribuan sporangium
pada tempat penetrasi hanya dalam waktu beberapa hari. Di dalam sporangium
dibentuk zoospora yang akan dilepas melalui papila setelah dinding sporangia
pecah (Agrios 1997).
Perbedaan interaksi isolat dengan genotipe cabai mengindikasikan bahwa
ada ras-ras patogenik pada isolat P. capsici (Hwang et al. 1996). AVRDC telah
mengidentifikasi tiga ras yaitu ras 1, 2, dan 3. Ras 1 adalah yang paling virulen
diikuti oleh ras 2 dan ras 3. Galur-galur diferensial cabai yang digunakan untuk
identifikasi ras adalah PI 188478, PBC 602, PBC 137, Early Calwonder (AVRDC
2000).
Cendawan P. capsici menyerang semua fase dan bagian tanaman. Serangan
pada pangkal batang ditandai dengan busuk batang berwarna coklat kehitaman,
tanaman layu tanpa terjadi penguningan daun terlebih dahulu (Yunianti 2007).
Pada tanaman dewasa, infeksi ditandai dengan munculnya lesio berwarna ungu
gelap dekat dengan permukaan tanah. Pada buah akan muncul bercak kecil, berarti
atau basah dan berwarna pudar. Perkembangan lesio lebih cepat biasanya
sepanjang ujung buah dibandingkan bagian tengah buah. Bagian jaringan yang
terinfeksi akan menjadi kering, cekung dan terdapat garis-garis (Erwin dan
Ribeiro 1996). Pada daun akan memperlihatkan gejala berupa bercak gelap dengan
bentuk dan ukuran yang tidak beraturan, jaringan mengering dan tampak seperti
terbakar matahari (Doolittle 1953).
Yunianti (2007) melaporkan bahwa gejala nekrosis pada bagian pangkal
dengan cendawan P. capsici. Marwiyah et al. (2006) melaporkan bahwa kematian
cabai dapat mencapai 65.3% setelah diinokulasi dengan isolat P. capsici ras 1.
Harahap (1991) menyatakan bahwa masa inkubasi penyakit berlangsung 2-3 hari
setelah inokulasi dan mengakibatkan buah gugur pada hari kelima serta kematian
tanaman pada hari ketujuh. Agrios (1997) menegaskan bahwa masa inkubasi
bervariasi waktunya dipengaruhi oleh interaksi antara inang dan patogennya,
stadium perkembangan inang, dan keadaan lingkungan.
Penyakit ini menyebabkan kerugian yang sangat besar. Perkembangannya
bergantung terutama pada suhu dan kelembapan, intensif tidaknya tindakan
penyemprotan dengan fungisida, umur tanaman saat diserang, serta tingkat
ketahanan yang dimiliki oleh varietas cabai yang ditanam (Yunianti 2007).
Penyakit Layu Bakteri
Penyakit layu bakteri disebabkan oleh Rasltonia solanacearum yang
semula bernama Pseudomonas solanacearum. Penyakit ini menyerang tanaman
tembakau pada tahun 1890-an dan menyerang kacang tanah pada tahun 1905.
Penyebaran patogen ini meluas pada tanaman kentang, pisang, jambu mete, jahe,
dan cengkeh (Machmud 1993). Penyakit layu bakteri dapat menurunkan produksi
pada tanaman kentang sebesar 75% dan tomat 100% (Persley et al. 1985). Patogen
ini memiliki kisaran inang yang luas dan bersifat soil borne. Kemampuannya yang
mudah berubah strain menyebabkan penyakit ini sulit untuk dikendalikan
(Hayward 1991).
Bakteri R. solanacearum memiliki inang sebagian besar dari famili
Solanaceae, Musaceae, Asteraceae, dan Fabaceae (Hayward 1991). Pada tahun
1994, Hayward melaporkan bahwa jumlah inang patogen ini mencapai 50 famili.
Menurut Machmud et al. (1999) patogen penyakit ini memiliki kisaran inang dan
daerah sebaran penyakit yang luas dan kemampuan bertahan hidup dalam tanaman
yang luar biasa di dalam tanah dan rizosfer tanaman yang bukan inangnya, dan
variabilitasnya sangat tinggi serta daya rusak yang besar.
Bakteri R. solanacearum dikelompokkan menjadi lima ras berdasarkan
kisaran inangnya dan lima biovar berdasarkan kemampuannya mengkonsumsi
sulit dilakukan terutama pada tingkat sub spesies karena sifat patogen ini yang
kompleks dan sangat heterogen (Machmud et al. 1999).
Tabel 1 Pengelompokan ras penyakit R. solanacearum (Persley et al. 1985)
Ras Inang Daerah Sebaran
1 Kacang tanah, famili solanaceae dan beberapa spesies gulma
Dataran rendah tropik dan sub tropik
2 Pisang, Heliconia, dan tanaman famili Musaceae lainnya
Asia, daerah tropik Amerika
3 Khusus menyerang kentang dan beberapa inang alternatif
Daerah tropik dan subtropik
4 Jahe 5 Mulberry
Yulianah (2007) melaporkan bahwa pada penelitiannya, gejala layu terlihat
pada daun, pangkal batang serta akar menghitam dan tampak basah. Pada genotipe
yang rentan terhadap layu bakterti akan terlihat gejala layu mulai 4 hari setelah
inokulasi. Martin dan French (1985) menyatakan gejala khas penyakit layu adalah
terjadinya kelayuan, daun menguning dan tanaman menjadi kerdil. Pada tanaman
tahan, kelayuan berkurang namun terjadi pengerdilan pada tanaman muda.
Hayward (1983) menerangkan bahwa gejala layu pertama kali terlihat biasanya
pada tanaman yang berumur kurang lebih 6 minggu dan dimulai pada daun-daun
muda. Pada tanaman muda dan sukulen dari varietas rentan, kelayuan yang hebat
disertai dengan robohnya batang tanaman. Batang tanaman yang sakit jika
dipotong akan terlihat berkas pembuluh yang berwarna coklat dan jika ditekan
akan mengeluarkan eksudat berupa lendir yang berwarna putih susu. Semangun
(2000) menyatakan bahwa jika potongan batang yang sakit dimasukkan ke dalam
air steril maka berkas pembuluh akan mengeluarkan aliran massa bakteri yang
berwarna putih. Ini merupakan ciri penting penyakit layu bakteri yang
membedakannya dengan penyakit layu yang disebabkan oleh cendawan Fusarium
sp.
R. solanacearum cenderung membentuk koloni tidak virulen atau tingkat
virulensinya lemah pada media biakan. Koloni virulen dan tidak virulen dapat
dideteksi dengan menumbuhkannya pada media yang mengandung 2,3,5
muda dengan bentuk tidak beraturan. Koloni yang tidak virulen berbentuk bulat
kecil dengan pusat merah tua. Pada media cair, bakteri virulen biasanya non motil
dan yang aktif bergerak merupakan bakteri yang tidak virulen (Hooker 1981).
Kelembaban dan suhu tanah mempengaruhi bertahannya bakteri penyebab
penyakit layu. Akiew (1985) melaporkan bahwa populasi R. solanacearum
menurun tajam pada peningkatan suhu tanah serta menurunnya kelembaban tanah.
Sedangkan pada kelembapan tanah tinggi dan suhu rendah, bakteri mampu
bertahan hidup lama dalam tanah. Kelman (1953) menerangkan bahwa suhu
optimum untuk perkembangan penyakit adalah 27-37 0C.
Kondisi pertumbuhan tanaman inang mempengaruhi ketahanannya
terhadap penyakit. Agrios (1997) menyatakan bahwa berkurangnya ketersediaan
nitrogen dapat meningkatkan kerentanan tanaman tomat terhadap layu Fusarium
dan layu R. solanacearum, tanaman tumbuh lebih lama, lebih lambat dan lebih
cepat tua. Kelebihan nitrogen menyebabkan pertumbuhan tanaman menjadi
sukulen dan dapat menyebabkan tumbuhan menjadi lebih rentan terhadap patogen
yang menyerang jaringan tumbuhan tersebut.
Pengujian ketahanan cabai terhadap layu bakteri dapat dilakukan dengan
cara menanam genotipe cabai pada lahan yang sudah terinfeksi secara alami dan
dapat juga dilakukan dengan inokulasi buatan. Khairul (2005) melaporkan bahwa
metode inokulasi dengan perendaman akar bibit yang telah dilukai lebih efesien
dibandingkan inokulasi dengan metode penyiraman suspensi ke pangkal batang
dan pengguntingan daun. Metode perendaman akar bibit ini dapat mempercepat
munculnya gejala layu (9.3 HSI) dengan kejadian penyakit yang lebih tinggi yaitu
sebesar 65%.
Perakitan Varietas Tahan
Pengendalian penyakit dengan menggunakan bahan kimia seperti yang
banyak dilakukan petani sangat tidak menguntungkan. Selain berbahaya bagi
petani dan konsumen, bahan kimia tersebut mahal dan tidak ramah lingkungan.
mempunyai modal terbatas sehingga dianjurkan pengendalian penyakit dengan
menanam varietas toleran, bibit yang sehat dan rotasi tanaman. Alternatif yang
baik dalam pengendalian penyakit adalah penggunaan varietas tahan.
Tahapan yang paling penting dalam program pemuliaan tanaman untuk
menghasilkan varietas yang tahan terhadap penyakit adalah mendapatkan sumber
ketahanan dan menentukan pola pewarisan sifat ketahanan tanaman serta sifat
genetik dari interaksi antar tanaman dengan patogen (Allard 1960). Varietas
tanaman yang tahan penyakit akan dihasilkan melalui seleksi plasma nutfah dan
persilangan dengan tetua terpilih. Sifat tahan ini dapat berasal dari varietas yang
berbeda, varietas komersil, atau spesies liar (Kallo 1988).
Tanaman tahan dan rentan dapat dibedakan dengan mudah jika ketahanan
dikendalikan oleh satu atau dua gen mayor, pada keadaan tersebut varians
ketahanan akan menunjukkan kurva sebagian terputus atau diskontinyu. Seringkali
pada ketahanan yang dikendalikan oleh banyak gen dan tidak ada perbedaan yang
jelas antara individu tanaman tahan dan tanaman rentan dalam populasi yang
bersegregasi (Allard 1960). Penentuan ketahanan terhadap sutau penyakit dapat
dilihat dari kejadian penyakitnya dan intensitas serangan. Russel (1981)
menyatakan bahwa penting untuk melakukan pengukuran atau estimasi terhadap
besarnya intensitas serangan dengan sistem pemberian nilai skor atas gejala yang
muncul.
Akhir-akhir ini varietas tahan yang banyak beredar di petani mulai dapat
dipatahkan ketahanannya akibat munculnya ras-ras baru dari suatu patogen
penyakit. Varietas tahan yang ada umumnya memiliki ketahanan vertikal yang
hanya tahan pada satu ras penyakit dan tidak bertahan lama. Kondisi ini semakin
diperburuk oleh kehadiran beberapa penyakit sekaligus di areal pertanaman petani.
Penyakit yang dominan menyerang pertanaman cabai adalah antraknosa, hawar
phytophthora, layu bakteri, dan virus.
Untuk mendapatkan varietas yang tahan pada beberapa penyakit maka
perlu dirakit suatu varietas multiline. Agrios (1997) menyatakan bahwa varietas
multiline adalah campuran individu-individu yang memiliki sifat agronomis yang
sama namun memiliki gen ketahanan yang berbeda. Perakitan varietas multiline
gen-gen ketahanan yang berbeda dengan genotipe yang memiliki sifat agronomi
yang baik. Gen-gen tersebut dipindahkan melalui hibridisasi.
Metode Silang Dialel
Pongoh (1987) menyatakan bahwa persilangan yang melibatkan dua tetua
sebagai induk persilangan akan membentuk rekombinasi gen yang sangat terbatas
dan tidak semua gen yang dikehendaki terkumpul pada galur yang diuji.
Disamping itu, keadaan demikian dapat mempersempit dasar genetik yang ada dan
mempercepat terjadinya pemantapan blok linkage serta kerapuhan genetik. Oleh
karena itu, sebaiknya setelah persilangan perlu diikuti dengan suatu metode seleksi
yang terarah atau memperbesar jumlah tetua persilangan yang akan digunakan.
Berkaitan dengan jumlah tetua yang digunakan maka dikenal istilah metode silang
dialel.
Analisis dialel membantu para pemulia menentukan pola heterosis antar
populasinya serta memilih bahan dan metode yang akan digunakan dalam program
pemuliaan. Persilangan dialel adalah persilangan yang melibatkan sejumlah
genotipe (varietas, galur, klon) dalam semua kombinasi sehingga masing-masing
genotipe mempunyai kesempatan untuk disilangkan dengan genotipe lain, dan
dapat juga dilakukan persilangan sendiri pada genotipe itu. Dengan analisis dialel
dapat diketahui kombinasi mana yang baik untuk menghasilkan suatu varietas
yang lebih baik. Penggunaan metode dialel harus memenuhi asumsi yaitu (1)
segregasi diploid, (2) tidak ada perbedaan antara persilangan resiprokal, (3) tidak
ada interaksi antara gen-gen yang tidak satu alel, (4) tidak ada multiallelisme, (5)
tetua homozigot, dan (6) gen-gen menyebar secara bebas diantara tetua (Singh &
Chaudhary 1979; Roy 2000).
Greenleaf (1986) menyatakan bahwa cabai merupakan tanaman diploid.
Hal ini menunjukan bahwa cabai memiliki gen-gen yang bersegregasi diploid
sehingga memenuhi asumsi untuk dilaksanakannya analisis dialel.
Singh dan Chaudary (1979) menyatakan bahwa pengujian nilai koefisien
regresi b pada garis Wr (peragam antara tetua dan keturunann dari array ke-r)
terhadap garis Vr (ragam dalam array ke-r) dapat menunjukkan ada atau tidaknya
dijelaskan bahwa tidak terdapat interaksi non allelik dan sebaliknya, jika diperoleh b≠1 maka mengindikasikan adanya peran interasi non allelik dalam mengendalikan suatu sifat.
Analisis silang dialel dapat dilakukan dengan dua pendekatan yaitu Hayman
dan Griffing. Pendekatan yang dikembangkan oleh Hayman dapat memberikan
informasi mengenai parameter genetik dari tetua-tetua yang digunakan dalam
persilangan sedangkan pendekatan Griffing memberikan informasi kemampuan
daya gabung dari tetua-tetua yang digunakan dalam persilangan (Singh dan
Chaudhary 1979).
Informasi yang dapat diperoleh dari pendekatan Hayman adalah (1) nilai D
yaitu keragaman akibat pengaruh aditif; (2) F yaitu nilai tengah Fr genotipe
(rata-rata Fr untuk semua array), F peragam pengaruh aditif dan non aditif pada array
ke-r; (3) H1 yaitu keragaman karena pengaruh dominansi; (4) H2 yaitu menduga
sebaran gen dalam tetua dan digunkan untuk menduga proporsi gen negatif dan
positif pada tetua; (5) h2 merupakan pengaruh dominan (sebagai jumlah aljabar
dari semua persilangan saat heterozigous); (6) E yaitu keragaman karena pengaruh
lingkungan; (7) rata-rata tingkat dominansi; (8) proporsi gen-gen dengan pengaruh
positif dan negatif dalam tetua; (9) proporsi gen-gen dengan pengaruh dominan
dan resesif dalam tetua; (10) jumlah kelompok gen yang mengendalikan sifat dan
menimbulkan dominansi; (11) heritabilitas arti luas dan sempit (Singh dan
Chaudary 1979).
Menurut Griffing (1956) dalam melakukan analisis daya gabung
diperlukan tiga materi genetik, yaitu tetua, F1, dan resiproknya. Dengan
menggunakan ketiga set materi genetik tersebut, Griffing memberikan empat
metode dalam analisa daya gabung, yaitu Metode I yang disebut juga full diallel
yang melibatkan n tetua dan F1 hasil silangan serta resiprokalnya masing-masing
sejumlah [n(n-1)/2 persilangan; Metode II disebut juga metode half diallel yang
melibatkan n tetua dan F1 saja dengan jumlah [n(n-1)/2] persilangan; Metode III
melibatkan F1 hasil silangan serta resiprokalnya tanpa tetua dengan jumlah
silangan [n(n-1)] dan Metode IV hanya melibatkan F1 dengan jumlah silangan
Daya gabung adalah ukuran kemampuan suatu tetua bila disilangkan dengan
galur lain yang akan menghasilkan hibrida dengan penampilan superior (Allard
1960). Singh dan Chaudary (1979) menjelaskan bahwa daya gabung dibedakan
menjadi dua, yaitu gabung umum (general combining ability) dan daya gabung
khusus (spesific combining ability). Daya gabung umum adalah kemampuan suatu
genotipe yang menunjukkan kemampuan rata-rata keturunan bila disilangkan
dengan sejumlah genotipe lain. Daya gabung khusus adalah kemampuan individu
tetua untuk menghasilkan turunan yang unggul jika disilangkan secara spesifik
dengan tetua lainnya.
Daya gabung umum (DGU) yang besar dan positif menunjukkan bahwa
tetua tersebut mempunyai daya gabung yang baik. Nilai DGU yang negatif berarti
tetua yang bersangkutan mempunyai daya gabung (rata-rata) yang lebih rendah
dibandingkan dengan tetua-tetua lainnya. Daya gabung khusus (DGK) yang positif
menunjukkan bahwa tetua tersebut mempunyai kombinasi hibrida yang tinggi
dengan salah satu tetua yang digunakan. Sebaliknya, apabila DGK negatif berarti
tetua tersebut tidak mempunyai kombinasi hibrida yang tinggi dengan salah satu
dari tetua-tetua yang digunakan (Sujiprihati 1996).
Informasi genetik yang diperoleh dari pengujian DGU, DGK, dan
resiprokalnya akan berguna untuk menentukan tetua dan metode pemuliaan
tanaman yang sesuai dalam rangka perbaikan sifat-sifat tanaman (Sujiprihati
1996).DGU adalah nilai rata-rata dari galur-galur dalam seluruh kombinasi
persilangan bila disilangkan dengan dengan galur-galur lain. DGK adalah
penampilan kombinasi pasangan persilangan tertentu. Bila nilai pasangan tertentu
lebih baik dari pada nilai rata-rata keseluruhan persilangan yang terlibat, dikatakan
DGKnya baik (Poehlman dan Sleeper 1990). DGU yang besar menunjukkan tetua
yang bersangkutan mempunyai kemampuan bergabung dengan semua tetua
sedangkan nilai DGU yang rendah menunjukkan bahwa tetua tersebut mempunyai
kemampuan bergabung yang kurang baik terhadap semua tetua yang lain.
Menurut Falconer (1981) efek DGU dan DGK adalah indikator penting
dari nilai potensial suatu galur murni dalam kombinasi hibrida. Welsh (1981)
menambahkan bahwa DGU terutama merupakan hasil dari aksi gen aditif,
Heterosis
Allard (1960) menggambarkan heterosis sebagai vigor hibrida dari F1 yang
melebihi rata-rata kedua tetuanya. Sebenarnya vigor hibrida menerangkan
pertambahan dalam ukuran dan vigor suatu tanaman sedangkan heterosis
digunakan untuk pertambahan maupun pengurangan ukuran dan vigor suatu
tanaman. Istilah heterobeltiosis lebih menggambarkan vigor hibrida F1 yang
melebihi dari tetua terbaiknya.
Shull (1908) pertama kali mendeskripsikan fenomena ini, dimana dikatakan
bahwa individu yang heterozigot mengalami pembagian sel, pertumbuhan dan
mempunyai proses fisiologis yang lebih baik dari pada individu yang homozigot.
Kusandriani (1996a) melaporkan bahwa pada tanaman cabai juga ditemukan
fenomena heterosis sehingga dimungkinkan dibentuknya hibrida cabai. Greenleaf
(1986) menerangkan bahwa hibrida akan memperlihatkan penampilan yang lebih
baik dari rata-rata kedua tetuanya. Untuk itu perlu dibentuk tetua dari populasi
galur murni yang berbeda secara genetik. Fenomena inilah yang menjadi dasar
dibentuknya varietas hibrida.
Poespodarsono (1998) menerangkan bahwa terdapat teori dasar yang
menjadi landasan terjadinya heterosis, yaitu adanya interaksi antar alel dalam satu
lokus dimana fungsi masing-masing gen berbeda sehingga menambah nilai
interaksi dan ini dikenal dengan efek heterozigositas dalam arti dominan lebih
(overdominan). Selain itu, heterosis juga merupakan akumulasi alel dominan,
sehingga persilangan antara kedua tetua akan menambahkan alel dominan pada
F1nya. Teori terakhir yang menjelaskan tentang heterosis adalah adanya interaksi
antar alel dari lokus yang berbeda (interaksi nonalelik) dan ini berkaitan juga
dengan terjadinya epistasi.
Dikenal dua istilah ynag menjelaskan fenomena heterosis yaitu heterosis itu
sendiri dan heterobeltiosis. Heterosis merupakan peningkatan atau penurunan
penampilan hibrida dibandingkan nilai rata-rata kedua tetuanya. Heterobeltiosis
adalah peningkatan penampilan hibrida dibandingkan tetua terbaik yang digunakan
Heritabilitas
Heritabilitas adalah proporsi besaran ragam genetik terhadap besaran ragam
fenotipe suatu karakter tertentu (Makmur 1992). Roy (2000) menyatakan bahwa
heritabilitas arti sempit (narrow sense) menggambarkan pengaruh aditif dari alel
yang diwariskan dari tetua kepada turunannya. Seleksi sifat yang dikendalikan
oleh gen aditif diharapkan mendapatkan kemajuan seleksi yang besar dan cepat.
Crowder (1993) menegaskan bahwa heritabilitas merupakan parameter genetik
yang efektif dalam melakukan seleksi yang efektif.
Heritabilitas dapat digunakan dalam penentuan metode seleksi. Basuki
(1995) menjelaskan bahwa jika nilai heritabilitas arti sempit tinggi maka lebih
tepat menggunakan seleksi massa. Sebaliknya jika nilai heritabilitasnya rendah
maka dapat digunakan seleksi silsilah, uji kekerabatan (sib-test), dan uji keturunan
(progeny test). Selanjutnya bila ragam epistasis tinggi maka lebih tepat
menggunakan metode seleksi diantara famili dan pemuliaan galur (line breeding).
Bila gen dominan berperan maka program pemuliaan diarahkan untuk pembuatan
galur silang-dalam untuk membentuk hibrida. Bila ragam interaksi lingkungan
(GE) besar maka program pemuliaan diarahkan untuk mendapatkan varietas yang
sesuai dengan wilayah ekologis tertentu. Heritabilitas arti sempit dapat digunakan
untuk kemajuan genetik harapan akibat seleksi. Semakin besar nilai heritabilitas
suatu sifat maka semakin besar kemungkinan sifat tersebut diturunkan ke generasi
selanjutnya.
Roy (2000) menyatakan bahwa heritabilitas dapat diduga langsung dari
pendugaan komponen ragam dan secara tidak langsung melalui pendugaan nilai
regresi (b) serta korelasi antar kelas (t). Pendugaan komponen ragam dapat
diperoleh dari studi populasi dasar (P1, P2, F1, F2, dan backcross) dan berbagai
rancangan persilangan. Pada tanaman yang memperbanyak diri secara aseksual
seperti klon, seluruh ragam genetik diturunkan pada turunannya sehingga hanya
dapat diduga heritabilitas arti luas (h bs), Jika heritabilitas arti sempit mempunyai
nilai yang kecil melalui pendugaan komponen ragam maka diduga adanya
pengaruh dominansi atau pengaruh lingkungan yang besar. Namun kelemahan
menduga heritabilitas dari komponen ragam adalah standar error tidak dapat
BAHAN DAN METODE
Penelitian ini terdiri dari tiga percobaan, yaitu 1) karakterisasi daya hasil;
2) pengujian ketahanan terhadap penyakit (antraknosa, hawar phytophthora, dan
layu bakteri); dan 3) pendugaan parameter genetiknya menggunakan analisis
dialel. Penelitian ini dilakukan mulai Juni 2008 sampai November 2009 yang
bertempat di KP Leuwikopo, KP Alam Sinar Sari-Cibereum, dan Lab. Dik
Pemuliaan Tanaman Departemen AGH Faperta IPB.
Percobaan 1: Karakterisasi Daya Hasil
Karakterisasi daya hasil dilakukan terhadap 28 genotipe cabai yang berasal
dari koleksi Bagian Genetika dan Pemuliaan Tanaman Departemen AGH IPB,
galur-galur introduksi dari AVRDC dan cabai lokal yang sudah digalurkan.
Genotipe-genotipe yang digunakan dapat dilihat pada Lampiran 1. Percobaan
dilakukan dengan menggunakan Rancangan Kelompok Lengkap Teracak (RKLT)
faktor tunggal dengan 3 ulangan, masing-masing satuan percobaan terdiri dari 20
tanaman. Peubah yang diamati terdiri sifat kualitatif dan kuantitatif berdasarkan
Descriptors for Capsicum (IPGRI 1995) disajikan pada Lampiran 3.
Analisis data yang dilakukan adalah sebagai berikut:
1. Analisis ragam dengan menggunakan software SAS versi 9.0 dan bila
hasilnya menunjukkan hasil yang berbeda nyata maka dilakukan dengan uji
lanjut dengan DMRT taraf α =0.05. Sumber keragaman dalam RKLT
disajikan pada Tabel 2.
Tabel 2 Analisis ragam rancangan acak kelompok
Sumber Keragaman
Derajat Bebas (db)
Kuadrat Tengah (KT)
Nilai Harapan KT
F hitung
Ulangan r-1
Genotipe t-1 KTg σ2+ rσg2 KTg/KTe
Galat (r-1)(t-1) KTe σ2
2. Ragam genetik mengikuti formula Singh dan Chaudhary (1979) σ2
Keterangan: σ2
g = ragam genetik
KTg = kuadrat tengah genotipe KTe = kuadrat tengah galat r = ulangan
3. Standar deviasi ragam genetik
σ
σ2g = √[2/n2][(KTg2/(dbg +2)) + (KTe2/(dbe+2))]Keterangan:
σ
σ2g = standar deviasi ragam genetikKTg = kuadrat tengah genotipe
KTe = kuadrat tengah galat
n = ulangan
dbg = derajat bebas genotipe
dbe = derajat bebas galat
4. Heritabilitas arti luas
h2bs= (σ2g / σ2f ) x 100%
5. Analisis korelasi (r) menggunakan software Minitab versi 14. dengan
koefisien korela