KAJIAN GENETIKA KETAHANAN TANAMAN KAKAO
(Theobroma cacao L.) TERHADAP PENYAKIT BUSUK BUAH
(Phytophthora palmivora Butl) DI INDONESIA
R U B I Y O
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ii Dengan ini saya menyatakan dengan sebenarnya bahwa semua pernyataan dalam disertasi saya yang berjudul:
” Kajian Genetika Ketahanan Tanaman Kakao (Theobroma cacao L)
terhadap Penyakit Busuk Buah (Phytophthora palmivora Butler) di
Indonesia”
Merupakan gagasan atau hasil penelitian disertasi saya sendiri, dengan pembimbingan oleh para komisi pembimbing, terkecuali yang dengan jelas ditunjukkan rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain.
Semua data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, Oktober 2009
Rubiyo
iii ABSTRACT
RUBIYO, Genetic Study of Cacao (Theobroma cacao L.) Resistance against
Black Pod Disease (Phytophthora palmivora Butl) in Indonesia. Supervised by
SUDARSONO, AGUS PURWANTARA, TRIKOESOEMANINGTYAS, and SATRIYAS ILYAS.
Cacao (Theobroma cacao L.) is one of the estate crops having important
role in economy Indonesia. Cacao cultivation faces a lot of constraints, such as crop pests and diseases which can reduce the quality and production of cacao. One of the main diseases which attack cacao in Indonesia is black pod disease caused by Phytophthora palmivora (Butl.) Butl. The disease caused yield losses ranging from 40 to 50% in Indonesia and worldwide.
The research on genetic resistance of cacao against the disease caused by
P. palmivora in Indonesia is still very limited. This research can assist the effort of cacao breeding to produce cacao clones or hybrids which are resistant against P. palmivora and to establish heritability model of F1 progeny to produce crop materials having some high qualities, so that the production of cacao can be
improved. The objectives of this research: (1) To identify P. palmivora species
and to study its genetic variation in cacao production centre of some provinces in Indonesia (2) To establish a standard inoculation method for screening in cacao
resistance (3) To measure pathogenicity of P. palmivora upon cacao (4) To test
cacao germplasm collection against P. palmivora and use them as parental clones
to construct F1 hybrids (5) To study the correlation of resistance levels and observed quantitative characters of several cacao clones (6) To study the potency of high general and specific combining abilities and its heterosis effect on diallel
crossing of cacao clones so that F1 hybrid with high yield and resistance to P.
palmivora and its heritability can be obtained.
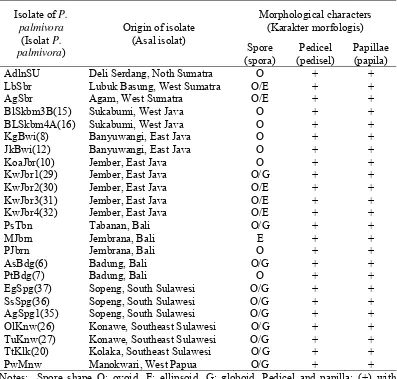
In the first part of this research, 24 indigenous isolates of P. palmivora had been isolated from 13 districts and eight provinces in Indonesia. The indigenous isolates produce ellipsoid, globoid, or ovoid sporangia with distinct papillae and pedicel, typical of P. palmivora. Among indigenous isolates, there was no distinct difference in the papilla and their pedicel. Even though the isolates showed similar in morphology, they showed variation in pathogenicity on cacao clones GC 7, ICS
60 and TSH 858. Phytophthora palmivora isolate from Lubuk Basung, West
Sumatra was very pathogenic to pods of the three cacao clones. While the isolates JkBwi (12) and KgBwi (8) from Banyuwangi, East Java; PtBdg (7) from Badung, Bali; SsSpg (36) and AgSpg1 (35) from Sopeng, South Sulawesi, and also Pwmnw from Manokwari, West Papua were pathogenic or very pathogenic.
The second part showed that inoculation using mycelia inoculum was
more efficient than using zoospore, and wounding treatment could assist in accurately detecting cacao seedling resistance against the P. palmivora infection. The estimation of resistance using detached pod was in line with the result of evaluation using cacao seedlings so that the seedlings can be used for an alternative for resistance evaluation against P. palmivora. TSH 858 clone is better to be used as female parent and crossed with Sca 12 as male parent to establish the
population of F1 hybrids which are resistant to the P. palmivora infection and
iv laboratory for resistance screening gave the similar degree of resistance. Cacao clones showing susceptibility in laboratory have the same susceptibility in the field.
The fourth part indicated that among 35 cacao clones, there were 10 clones which were resistant against P. palmivora infection based on pod inoculation test in laboratory. The clones are: ICCRI 1, ICCRI 3 PA 300, UIT 1, NIC 4, DR 38, ICS 13, TSH 858, Sca 6 and ICS 60. Whereas, the cacao clones showing susceptibility based on inoculation were: RCC 72, KKM 22, NIC 7, DRC 15, DRC 16, RCC71,BL 300, BL 301, KEE2, TSH 908. Genotypes used for the parental clones for future selection process were eight clones: ICCRI 1, ICCRI3, ICS 13, TSH 858, UIT 1, PA 300, NIC 4 and DR38.
The fifth part, based on stomata observation of 10 clones, stomata density in pod and leaf did not show high correlation to the resistance. The number of stomata on resistant and susceptible clones is not significantly different, indicating that cacao clones which are resistant do not always have low stomata density compared to that of susceptible, and vice versa cacao clones which are susceptible do not always have high stomata density in pod and leaf. Study on the activities of chitinase and peroxidase enzymes upon tested clones indicated that there was chitinase role to the resistance of cacao against the infection by P. palmivora. The increase of chitinase activity in resistant clones generally intensified consistently, and so did peroxidase enzyme. Susceptible cacao clones whose peroxidase enzyme activity did not increase were DRC 15 and DRC 16 and they belong to very susceptible clones.
The sixth part of the research showed that there was no gene interaction
v RINGKASAN
RUBIYO, Kajian Genetika Ketahanan Tanaman Kakao (Theobroma cacao L.)
terhadap Penyakit Busuk Buah (Phytophthora palmivora Butl) di Indonesia.
Dibimbing oleh SUDARSONO, AGUS PURWANTARA,
TRIKOESOEMANINGTYAS, dan SATRIYAS ILYAS.
Kakao (Theobroma cacao L.) merupakan salah satu komoditas perkebunan
yang mempunyai peran penting dalam perekonomian Indonesia. Budidaya kakao menghadapi banyak kendala di lapangan, antara lain penyakit dan hama tanaman yang dapat menurunkan kuantitas dan kualitas produksi kakao. Salah satu penyakit utama yang menyerang tanaman kakao di Indonesia adalah penyakit
busuk buah (black pod) yang disebabkan oleh Phytophthora palmivora (Butl).
Butl. Penyakit busuk buah kakao mengakibatkan kerugian antara 40 sampai 50% di Indonesia, dan di seluruh dunia.
Penelitian genetika ketahanan kakao terhadap penyakit P. palmivora di
Indonesia belum banyak dilakukan. Penelitian ini dapat membantu usaha pemuliaan untuk memperoleh bahan tanam yang tahan terhadap penyakit busuk buah dan model pewarisan terhadap F1 nya untuk menghasilkan bahan tanam yang mempunyai beberapa sifat unggul, sehingga produksi kakao dapat ditingkatkan. Penelitian ini bertujuan untuk: (1) Melakukan identifikasi spesies P. palmivora dan mengetahui keragaman patogenisitasnya pada lokasi sentra kakao di Indonesia (2) Mengetahui metode inokulasi untuk penapisan ketahanan kakao
(3) Mengetahui patogenitas P. palmivora terhadap tanaman kakao (4) Mengetahui
dan mendapatkan tanaman kakao yang tahan dan rentan terhadap penyakit P.
palmivora di koleksi plasma nutfah kakao untuk digunakan sebagai tetua untuk perakitan hibrida F1 (5) Mengetahui korelasi tingkat ketahanan beberapa klon kakao untuk karakter kuantitatif yang diamati (6) Mengetahui potensi daya gabung umum dan khusus yang tinggi serta efek heterosisnya pada persilangan dialel klon kakao sehingga diperoleh potensi pada hibrida F1 serta heritabilitasnya.
Pada bagian pertama dari penelitian ini telah diperoleh 24 isolat indigenus P. palmivora yang diisolasi dari 13 kabupaten dan delapan provinsi di Indonesia. Isolat indigenus yang didapat mempunyai bentuk sprora ellipsoid, globoid, atau ovoid, tipikal P. palmivora. Di antara isolat tidak terdapat perbedaan yang jelas
pada papila dan pediselnya. Meskipun isolat P. palmivora yang didapat secara
morfologis hampir sama, terdapat perbedaan yang besar dalam tingkat
patogenisitasnya terhadap kakao klon GC 7, ICS 60 atau TSH 858. Isolat P.
palmivora LbSbr dari Lubuk Basung, Sumatra Barat diketahui sangat patogenik terhadap ketiga kultivar kakao yang diuji. Isolat JkBwi (12) dan KgBwi (8) dari
Banyuwangi, Jawa Timur; PtBdg (7) dari Badung, Bali; SsSpg (36) dan AgSpg1
(35) dari Sopeng, Sulawesi Selatan, serta PwMnw dari Manokwari, Papua Barat bersifat patogenik atau sangat patogenik.
Bagian kedua dari penelitian ini menunjukkan bahwa inokulasi dengan
menggunakan inokulum miselia lebih efisien dibandingkan dengan zoospora P.
Palmivora, dan perlakuan pelukaan lebih mampu secara akurat menduga respon
ketahanan bibit kakao terhadap infeksi P. palmivora. Hasil pendugaan ketahanan
vi
terhadap P. palmivora. Klon TSH 858 lebih baik untuk digunakan sebagai induk
betina dan disilangkan dengan Sca 12 sebagai induk jantan untuk menghasilkan
populasi hibrida F1 yang resisten terhadap infeksi P. palmivora dan berpotensi
berdaya hasil tinggi.
Bagian ketiga menunjukkan bahwa inokulasi untuk mengetahui ketahanan klon kakao di laboratorium maupun di lapangan menghasilkan ketahanan yang sama. Terdapat perbedaan dalam perkembangan luas bercak dan masa inkubasinya. Umumnya rata-rata luas bercak dan perkembangan yang dihasilkan pada inokulasi di laboratorium lebih besar dibandingkan dengan luas bercak yang di hasilkan pada uji inokulasi di lapangan. Masa inkubasi umumnya di lapangan lebih lamban rata-rata 2 hari dibandingkan dengan inokulasi di laboratorium.
Bagian keempat hasil penelitian menunjukkan bahwa berdasarkan hasil uji ketahanan terhadap 35 klon kakao, terdapat 10 klon kakao yang resisten terhadap
infeksi P. palmivora berdasarkan uji inokulasi buah di laboratorium. Klon kakao
tersebut adalah: ICCRI 1, ICCRI 3, PA 300, UIT 1, NIC 4, DR 38, ICS 13, TSH 858, Sca 6 dan ICS 60. sedangkan klon kakao yang menunjukkan hasil rentan berdasarkan hasil inokulasi adalah: RCC 72, KKM 22, NIC 7, DRC 15, DRC 16, RCC71, BL 300, BL 301, KEE2, TSH 908. Genotipe yang digunakan untuk tetua dalam rangka proses seleksi lebih lanjut atau untuk bahan tanam klonal ada delapan klon yaitu: ICCRI 1, ICCRI 3, ICS 13, TSH 858, UIT 1, PA 300, NIC 4 dan DR38.
Bagian kelima menunjukkan bahwa berdasarkan hasil pengamatan stomata pada 10 klon, kerapatan stomata pada daun maupun buah tidak memberikan korelasi yang tinggi terhadap ketahanan. Jumlah stomata tidak berbeda nyata antara kelompok klon yang tahan maupun rentan. Klon kakao yang tahan tidak selalu menghasilkan jumlah kerapatan stomata yang lebih sedikit dibandingkan dengan yang rentan. Klon kakao yang rentan tidak selalu memiliki jumlah stomata yang banyak di daun maupun pada buah. Aktivitas kitinase dan peroksidase terhadap klon kakao yang diuji mengindikasikan ada peran kitinase terhadap
ketahanan kakao dari infeksi P. palmivora. Peningkatan aktivitas kitinase klon
yang tahan umumnya lebih meningkat, begitu juga pada enzim peroksidase. Klon kakao yang rentan, dan tidak memiliki peningkatan aktivitas enzim peroksidase adalah klon DRC 15 dan DRC 16, sehingga klon tersebut masuk dalam kelompok sangat rentan.
Bagian ke enam hasil penelitian menunjukkan bahwa tidak ada interaksi
gen yang terjadi dalam menentukan ketahanan terhadap penyakit P. palmivora.
vii ©Hak cipta milik Institut Pertanian Bogor, tahun 2009
Hak cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
viii
(Theobroma cacao L.) TERHADAP PENYAKIT BUSUK BUAH
( Phytophthora palmivora Butl.) DI INDONESIA
R U B I Y O
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ix Penguji pada Ujian Tertutup : Dr.Ir.Yudiwanti Wahyu E.K, MS.
Dr.Ir.Muchdar Sudarjo, M.Sc.
Penguji pada Ujian Terbuka: Prof.Dr.Ir. Sudirman Yahya
x (Theobroma cacao L.) terhadap Penyakit Busuk Buah (Phytophthora palmivora Butl) di Indonesia
Nama : Rubiyo
Nomor Pokok : A 161060011
Program Studi : Agronomi
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Sudarsono, M.Sc. Dr. Ir. Agus Purwantara, APU.
Ketua Anggota
Dr. Ir. Trikoesoemaningtyas, M.Sc. Anggota
Prof. Dr.Ir. Satriyas Ilyas, M.S. Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, M.S. Prof. Dr. Ir. Khairil A.Notodiputro, M.S.
xi PRAKATA
Bismillahirrahmanirrahiim. Alhamdulillah, puji dan syukur penulis sampaikan kehadirat Alloh SWT atas segala karunia dan petunjuk-NYA, sehingga penelitian dan penulisan disertasi ini dapat diselesaikan. Solawat dan salam semoga senantiasa tercurah kepada Nabi Muhammad SAW, yang telah membawa cahaya dan petunjuk bagi kehidupan umat manusia hingga akhir zaman.
Disertasi dengan judul “Kajian Genetika Ketahanan Tanaman Kakao
(Theobroma cacao L.) terhadap Penyakit Busuk Buah (Phytophthora
palmivora Butl) di Indonesia “ disusun berdasarkan penelitian–penelitian yang dilakukan di lapangan yang meliputi sembilan propinsi (Sumatra Utara, Sumatra Barat, Jawa Barat, Jawa timur, Bali, Sulawesi Selatan, Sulawesi Barat, Sulawesi Tenggara dan Papua Barat) saat pengambilan isolat di Indonesia. Penelitian di
laboratorium Balai Penelitian Bioteknologi Perkebunan Bogor, laboratorium fitopatologi dan kebun percobaan Kaliwining Pusat Penelitian Kopi dan Kakao Indonesia Jember Jawa Timur Lembaga Riset Perkebunan Indonesia.
xii penulisan disertasi dapat diselesaikan. Karena itu ungkapan dan penghargaan ucapan terimakasih penulis sampaikan kepada:
1. Ketua komisi pembinaan tenaga Badan Penelitian dan Pengembangan Pertanian Departemen Pertanian atas kepercayaan, biaya dan fasilitas yang diberikan selama tugas belajar sehingga penulis dapat menyelesaikan pendidikan Program S3 di Institut Pertanian Bogor.
2. Kepala Badan Penelitian dan Pengembangan Pertanian yang telah memberikan beasiswa dan biaya penelitian, ijin dan kesempatan tugas belajar S3 kepada penulis di Institut Pertanian Bogor
3. Rektor Institut Pertanian Bogor, Dekan Sekolah Pascasarjana, Ketua PS Agronomi Sekolah Pascasarjana Institut Pertanian Bogor atas kesempatan dan dukungan yang diberikan mulai dari perkuliahan sehingga penulis dapat menjalankan penelitian dan menyelesaikan penulisan disertasi ini dengan baik.
4. Dr. H. Darmono Taniwiryono dan Dr.Hj. Endang Nurhayati yang telah menguji penulis pada ujian lisan Prakualifikasi Program Doktor.
5. Kepala BPTP Bali dan staf atas dukungannya selama penulis mengikuti tugas belajar program Doktor di Institut Pertanian Bogor.
6. Direktur Balai Penelitian Bioteknologi Perkebunan Bogor-Lembaga Riset Perkebunan Indonesia atas ijin dan fasilitas yang diberikan selama penelitian berlangsung di Laboratorium maupun rumah kaca.
8. Direktur Pusat Penelitian Kopi dan Kakao Indonesia atas ijin dan fasilitas yang diberikan selama penelitian berlangsung baik di kebun percobaan Kaliwining maupun Laboratorium dan perpustakaan dalam studi literatur.
xiii 10. Staf Pengajar Program Studi Agronomi Sekolah Pascasarjana Institut
Pertanian Bogor yang telah memberikan ilmu selama penulis mengambil kuliah untuk program S3 di Institut Pertanian Bogor.
11. Dr.Ir. H. Sutanto Abdulah, SU., Ir.Sri-Sukamto,MP., Ir.Dedy Suhendi, MS., Ir. Sudarsianto, Ir. Nurkolis, atas bantuannya selama penelitian berlangsung di laboratorium dan lapangan yang diberikan, juga diskusi-diskusi yang sangat membantu sehingga sangat dimungkinkan penelitian berlangsung dengan lancar.
12. Ucapan yang sama juga disampaikan kepada Dr.M.Syukur, Atekan, SP, M.Si; Alfian Futuhulhadi, S.Si, M.Si atas bantuan dalam analisa data penelitian ini.
13. Ir.Endang Mufrihati yang telah banyak membantu dalam pengambilan isolat di Provinsi Sumatera Utara, Sumatra Barat dan Papua Barat, Ir.Suparti Disbun Sopeng yang telah membantu dalam pengambilan isolat di daerah Sulawesi Selatan, Ucapan yang sama juga disampaikan kepada Ir. Ramlan, MP Peneliti di BPTP Sulbar yang membantu dalam pengambilan isolat di Sulawesi Barat, Imran, SP peneliti BPTP Sulawesi Tenggara juga membantu pengambilan isolat di daerah Sultra dan Mas Rahmat (yang dengan tekun membantu dalam isolasi isolat dan
inokulasi di Laboratorium dan Green house Balai Penelitian Bioteknologi Perkebunan Bogor).
14. Rekan–rekan di laboratorium dan kebun percobaan Kaliwining: Supandi, SP., Ir.Dody Sulistyo, Suliono, Sukarmin, Sarkawi, Adi Hario, mbak Khotijah, Mbak Imsiah, bu Aan, mas Sumarto dan pak Karmidin, dan berbagai pihak yang telah banyak membantu di kebun maupun di laboratorium, mulai dari persiapan persilangan hingga inokulasi dan tabulasi data.
15. Rekan di Lab.Biomol IPB (mbak Minarti, Susiani dan Mas Agus serta Mas Joko) yang telah banyak membantu dalam penelitian ini.
xiv kedua orang tua penulis: Bapak Yasareja [Alm] dan Ibu Mukinah atas perjuangan, pengorbanan dan doa yang tidak pernah terhenti dalam membesarkan dan mendidik penulis.
18.Terimakasih kepada Bapak dan Ibu mertua (Bapak Ismail Mahardi [alm] dan Ibu Siti Juariah) yang telah mendidik dan memberikan istri yang sangat baik bagi penulis.
19.Rasa terimakasih yang sebesar-besarnya juga penulis sampaikan kepada istri tercinta Ir. Endang Mufrihati serta ananda Nicho Nurdebyandaru, S.Si, Briliandaru Mahardhiyasa Pribadi, Dhimas Upadyandaru SB, Prabandaru Nuriza Daksa B dan F.Adellia Virgiandaru Huzna yang telah melengkapi dan memberikan kebahagiaan bagi kehidupan penulis serta dengan segala pengertian, pengorbanan, kesabarannya dan doa yang tidak pernah terlupakan selama penulis menyelesaikan studi S3 ini. 20.Terimakasih juga penulis sampaikan kepada kakak dan adik (Mas Ir.
Zaenal Arifin, mbak Ir.Susilowati, Mas Samsul Hadi, mbak Rohma, dik Min, dik Mini dik Tugiya, SE, dik Mery beserta keluarga) serta sanak famili yang tidak dapat penulis sebutkan satu persatu.
21.Terimakasih kepada berbagai pihak yang telah membantu pelaksanaan
dan penyelesaian disertasi penulis yang tidak dapat disebutkan satu persatu dalam tulisan ini.
Semoga bimbingan, bantuan, dukungan dan doa dari berbagai pihak akan menjadi amal baik dan mendapatkan balasan yang berlipat ganda dari Alloh SWT. Akhir kata, semoga tulisan ini bermanfaat bagi pengembangan dan kemajuan ilmu pengetahuan khususnya perkakaoan di Indonesia untuk kehidupan dan kemakmuran kita bersama. Amin.
Bogor, Oktober 2009
xv RIWAYAT HIDUP
Penulis dilahirkan di Gunung Kidul Yogyakarta tanggal, 11 November 1963 sebagai anak sulung pasangan Yasareja dan Mukinah. Pendidikan sarjana ditempuh di Program Studi Budidaya Pertanian, Fakultas Pertanian Universitas Moch. Sroedji Jember, lulus pada tahun 1989. Pada tahun 2004 menyelesaikan pendidikan di Sekolah Pascasarjana Universitas Udayana program studi pertanian lahan kering. Kesempatan untuk melanjutkan ke program Doktor pada Program Studi Agronomi Sekolah Pascasarjana Institut Pertanian Bogor diperoleh pada tahun 2006. Beasiswa pendidikan Pascasarjana S3 diperoleh dari Badan Penelitian dan Pengembangan Pertanian – Departemen Pertanian.
Penulis mulai bekerja di Balai Penelitian Perkebunan Jember sekarang Pusat Penelitian Kopi dan Kakao Indonesia tahun 1984 - 1997 sebagai staf pada bidang pemuliaan tanaman kakao. Mulai tahun 1998 penulis memilih jalur PNS dan ditempatkan di Balai Pengkajian Teknologi Pertanian (BPTP) Kendari lingkup Badan Penelitian dan Pengembangan Pertanian sebagai Staf Peneliti. Tahun 2001 sampai sekarang sebagai peneliti di BPTP Bali. Pada tahun
2002-2004 sebagai Pemimpin Proyek PAATP Bali. Jabatan fungsional Peneliti pertama diperoleh pada tahun 2000 sebagai Asisten Peneliti Madya bidang budidaya pertanian dan tahun 2003 sebagai Ajun Peneliti Madya, kemudian jabatan Peneliti Madya diperoleh pada tahun 2005. Selama menjadi peneliti karya tulis yang dipublikasikan di Jurnal Ilmiah Nasional sebagai penulis utama 12 buah dan sebagai penulis kedua 15 buah.
Dua buah karya ilmiah berjudul (1) “Response of 35 Cacao Collections of
Indonesian Coffee and Cacao Research Institute against Phytophthora palmivora
Butl. Infection Based on Detached Pod Assays “ dan (2) Judul” Heritability and
Genetic control of black pod disease caused by P. palmivora infection in cacao”
diterima sebagai makalah dan akan dipresentasikan pada International Cacao Research Conference (ICRC) di Bali November 2009. Dua artikel telah
xvi
kedua (2) dengan judul: “Uji ketahanan kakao (Theobroma cacao L.) terhadap
penyakit busuk buah dan efektivitas metode inokulasi” pada Jurnal Pelita
Perkebunan. Karya-karya ilmiah tersebut merupakan bagian dari program S3 penulis. Sebagai seorang peneliti pemuliaan kakao bersama dengan peneliti yang lain pada tahun 2004 dan 2005 telah melepas 4 klon kakao (ICCRI 1, ICCRI 2, ICCRI 3 dan ICCRI 4) sebagai klon unggul nasional.
xvii DAFTAR ISI
Halaman HALAMAN PENGESAHAN ... I KATA PENGANTAR ... Ii DAFTAR ISI ... Iii
RINGKASAN ... 1
PENDAHULUAN... 1
Latar Belakang ... 1
Perumusan masalah ... 5
TUJUAN PENELITIAN ... 7
MANFAAT PENELITIAN... 7
HIPOTESIS ... 8
TINJAUAN PUSTAKA... 9
A.MorfologiTanaman kakao ... 9
B. Keragaman GenetikTanaman kakao... 11
C. Penyakit Busuk Buah Kakao ... 12
D. Pengendalian Penyakit Busuk Buah... 16
E. Mekanisme Ketahanan... 17
F Mekanisme Ketahanan Struktural ... Mekanisme Ketahanan Biokimia ... 18 20 G. Genetika Ketahan Kakao terhadap Penyakit P.palmivora... 22
H. Analisis Daya Gabung... 24
I. Heterosis ... 25
J.Heritabilitas... 26
JUDUL 1. ISOLATION OF INDIGENOUS Phytophthora palmivora FROM INDONESIA, THEIR MORPHOLOGICAL AND PATHOGENICITY CHARACTERIZATIONS ... 28
Abstrak ... 28
Introduction ... 31
Material and Methods... 33
Results and Discussion... 36
Conclusions ... 44
xviii TERHADAP PENYAKIT BUSUK BUAH DAN EFEKTIVITAS
METODE INOKULASI ... 47
Abstrak ... 48
Pendahuluan ... 49
Bahan dan Metode... 50
Inokulasi pada Buah Kakao... 51
Inokulasi pada Bibit ... 52
. Pengaruh Genotipe Kakao... 52
Hasil dan Pembahasan... 54
Simpulan... 65
Daftar Pustaka ... 66
JUDUL 3. UJI KETAHANAN KAKAO DI LAPANGAN DAN LABORATORIUM TERHADAP PENYAKIT BUSUK BUAH (Phytophthora palmivora Butler)... 68
Abstract ... 69
Pendahuluan ... 70
Bahan dan Metode... 72
Penelitian uji ketahanan tanaman kakao di laboratorium dan lapangan .. 72
Uji Detached Pod di laboratorium... 73
Uji ketahanan di lapangan ... 73
Hasil dan Pembahasan... 74
Uji ketahanan kakao di laboratorium ... 74
Uji ketahanan di lapangan ... 76
Simpulan... 78
Daftar Pustaka ... 79
JUDUL 4. PENELITIAN RESISTENSI KLON KAKAO TERHADAP INFEKSI Phytophthora palmivora Butl RESPON 35 KLON KAKAO BERDASARKAN UJI DETACHED POD ... 82
Abstract ... 82
Pendahuluan ... 84
Bahan dan Metode... 86
Hasil dan Pembahasan... 89
Uji Detached Pod di Laboratorium ... 89
xix
Pendugaan nilai duga ragam genetik... 97
Simpulan... 98
Daftar Pustaka ... 98
JUDUL 5. AKTIVITAS ENZIM KITINASE , PEROKSIDASE SERTA KERAPATAN STOMATA PADA KETAHANAN KAKAO (Theobroma cacao L) TERHADAP PENYAKIT BUSUK BUAH ( P. Palmivora) ... 100
Apstrak ... 101
Pendahuluan ... 102
Bahan dan Metode... 104
A. Penelitian Kerapatan stomata terhadap buah dan daun akao ... 104
Pengamatan stomata buah kakao... 104
B. Penelitian Aktivitas Kitinase dan Peroksidase Daun pada Beberapa Klon kakao ... 105
Ekstraksi Protein ... 106
Analisis Total Protein Terlarut (TPT) ... 106
Analisis Aktivitas kitinase... 106
Analisis Aktivitas Peroksidase ... 107
Hasil dan Pembahasan... 107
Simpulan... 111
Daftar Pustaka ... 112
JUDUL 6. PENDUGAAN PARAMETER GENETIK UNTUK KARAKTER KETAHANAN TANAMAN KAKAO (Theobroma cacao L) TERHADAP PENYAKIT BUSUK BUAH (Phytophthora palmivora.)... 115
Abstrak ... 115
Pendahuluan ... 117
Bahan dan Metode... 119
Lokasi dan Waktu Penelitian... 119
Bahan Tanaman yang Digunakan ... 120
Uji Ketahanan Populasi Hibrida F1 Hasil Persilangan Dialel... 122
Analisis Data ... 124
Hasil dan Pembahasan... 130
Pendugaan parameter genetik... 132
Interaksi Gen ... 132
xx
Tingkat Dominansi. ... 134
Proporsi gen dominan terhadap gen Resesif ... ... 134
Arah dan urutan dominan ... ... 135
Jumlah Gen Pengendali karakter... 136
Heritabilitas ... 136
Daya Gabung Umum (DGU) ... 137
Daya Gabung Khusus (DGK) ... 137
Heterosis... 139
Simpulan... 140
Daftar Pustaka. ... 141
PEMBAHASAN UMUM ... 144
SIMPULAN DAN SARAN ... 157
SARAN ... 158
DAFTAR TABEL
Halaman
1 Daftar lokasi pengambilan contoh buah kakao terinfeksi penyakit
busuk buah kakao, jumlah isolat cendawan dan jumlah isolat P.
palmivora indigenus yang teridentifikasi dari masing-masing lokasi.... 37
2 Karakteristik morfologis isolat P. palmivora indigenus berdasarkan
bentuk spora dan keberadaan pedisel serta papila... 39
3 Pengelompokkan patogenisitas isolat P. palmivora indigenus yang
diisolasi dari berbagai pusat produksi kakao di Indonesia berdasarkan respons buah kakao klon GC7 (rentan), ICS60 (agak resisten), dan TSH858 (resisten – terhadap infeksi P. palmivora)... 41
4 Pathogenicity grouping of indigenous isolates of Phytophthora
palmivora isolated from various cacao production centers in Indonesia based on the response of pods of cacao clones GC7 (susceptible), ICS60 (moderately resistance), and TSH858 (resistance - against P. palmivora infection)... 42
5 Pengaruh jenis inokulan terhadap persentase bibit kakao klon GC 7
dan Sca 12 yang terinfeksi P. palmivora dan kisaran panjang bercak
pada batang yang dihasilkan. Pengamatan panjang bercak dilakukan 28 hari sesudah inokulasi batang dari bibit kakao yang diuji. ... 60
6 Pengaruh pelukaan terhadap persentase bibit kakao klon GC 7 dan
Sca 12 yang terinfeksi P. palmivora dan kisaran panjang bercak
pada batang yang dihasilkan. Pengamatan dilakukan 28 hari sesudah inokulasi bibit kakao yang diuji. ... 61
7 Jumlah bercak bibit kakao hibrida F1 (ICS 60 x Sca 12) dan hibrida
F1 (TSH 858 x Sca 12) serta zuriat kakao klon Sca 12 yang
terinfeksi P. palmivora dan rataan panjang bercak pada daun yang
dihasilkan. Pengamatan panjang bercak dilakukan 7 (I), 14 (II), and 21 (III) hari sesudah inokulasi daun bibit kakao yang diuji ... 64
8 Persentase bibit kakao hibrida F1 (ICS 60 x Sca 12) dan hibrida F1
(TSH 858 x Sca 12) serta zuriat kakao klon Sca 12 yang terinfeksi P. palmivora dan kisaran panjang bercak pada daun yang dihasilkan. Pengamatan lebar bercak dilakukan 14 (I), 21 (II), and 28 (III) hari sesudah inokulasi daun bibit kakao yang diuji... 64
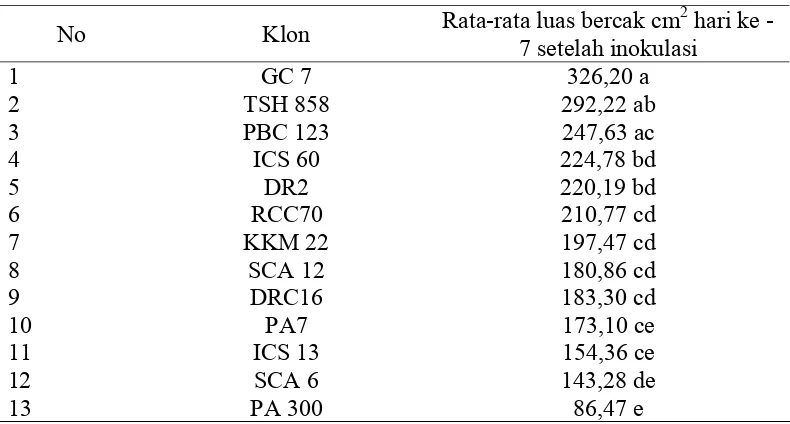
9 Rata-rata luas bercak (cm2) hasil inokulasi beberapa klon kakao di
Laboratorium 7 hari setelah inokulasi P. palmivora... 74
10 Rata-rata luas bercak (cm2) hasil inokulasi beberapa klon kakao di
Lapangan 9 hari setelah inokulasi P. palmivora... 76
11 Material Genotipe Kakao yang digunakan sebagai uji Evaluasi
vi
diinokulasi dengan miselia P. palmivora menggunakan uji detached
pod di laboratorium. Pengamatan dilakukan pada tiga sampai
dengan tujuh hari sesudah inokulasi (HSI)………... .. 91
13 Rataan lebar bercak pada permukaan buah kakao yang diinokulasi
dengan miselia P. palmivora menggunakan uji detached pod di
laboratorium. Pengamatan dilakukan pada tiga sampai dengan tujuh hari sesudah inokulasi (HSI) ... 93
14 Rataan luas bercak pada buah yang diuji dan pengelompokan
respons terhadap infeksi P. palmivora berdasarkan uji detached pod di laboratorium. Pengamatan dilakukan pada 3 - 7 hari sesudah inokulasi (HSI) ... 94
15 Persentase buah tanpa gejala, rataan luas bercak dan pengelompokan
respons terhadap infeksi P. palmivora berdasarkan uji detached pod di laboratorium. Pengamatan dilakukan pada 7 hari sesudah inokulasi (HSI) ... 95
16 Tipe kakao, bentuk buah dan pengelompokan respons klon kakao
koleksi Puslit Kopi dan Kakao Indonesia terhadap infeksi P.
palmivora berdasarkan uji detached pod di laboratorium. Penentuan respons didasarkan pada luas bercak yang diamati pada 7 hari sesudah inokulasi (HIS) ... 95
17 Nilai duga ragam genetik luas bercak setelah inokulasi hari ke 6 dan
ke 7 ... 97
18 Rata-rata Kerapatan Stomata Daun dan buah (cm2) Pada Beberapa
Klon Kakao ... 108
19 Kandungan dan aktivitas kitinase ( µM pNP/mg protein/jam) pada
daun kakao sehat dan terinfeksi penyakit busuk buah P. Palmivora... 110
20 Kandungan dan Aktivitas Peroksidase ( µM pNP/mg protein/jam)
pada daun kakao sehat dan terinfeksi penyakit busuk buah P.palmivora... 110
21 Karakteristik klon sebagai tetua untuk pembentukan populasi hibrida
F1... 121 22 Persilangan setengah dialel menggunakan lima tetua ... 122 23 Komponen Analisis ragam analisis silang dialel... 123 24 Pengelompokan ketahanan ... 124 25 Komponen Analisis Ragam untuk populasi dialel ... 125 26 Persilangan dialel ketahanan kakao terhadap P. palmivora... 125
27 Komponen analisis ragam untuk daya gabung menggunakan
vii 29
Pendugaan parameter genetik ketahanan genotip kakao terhadap P.
palmivora ……… 133
30 Heritabilitas dalam arti luas (h2bs) dan heritabilitas dalam arti sempit
(h2ns ) komponen ketahanan berdasarkan luas bercak dan Intensitas
Penyakit terhadap P. palmivora. ... 136
31 Anova daya gabung karakter luas bercak terhadap penyakit
Phytopthora palmivora pada tanaman kakao... 137
32 Nilai efek daya gabung umum (DGU) dan Daya Gabung Khusus
(DGK) genotipe tanaman kakao berdasarkan luas bercak dan intensitas penyakit hasil inokulasi ketahanan terhadap penyakit Phytopthora palmivora... 138
33 Penampilan tetua, F1, nilai heterosis rata-rata tetua, dan heterosis
rata-rata tetua tertinggi persilangan genotipe tanaman kakao berdasarkan luas bercak... 139
34 Penampilan tetua, F1, nilai heterosis rata-rata tetua, dan heterosis
Halaman
1 Beberapa Type Kakao trinitario yang berkembang di Indonesia ... 12
2 A dan B Bentuk Sporangium P.palmivora dan pedise... 14
3 (A). Serangan P.palmivora pada buah kakao dan (B).Serangan
P.palmivora pada bibit ... 15
4 (A) Kerapatan stomata pada epidermis buah kakao (B) Tabung
kecambah saat penetrasi pada stomata ... 19
5 (A) buah kakao klon GC7 (rentan), (B). Buah kakao klon DRC 16
(resisten) ... 23
6 Tahapan isolasi P. palmivora indigenus dari contoh buah kakao
terinfeksi busuk buah dari lapangan dan karakter morfologis isolat.
(a) Contoh buah kakao terinfeksi; (b) Baiting step – inokulasi buah kakao sehat dengan miselia potongan contoh buah sakit dari lapangan, (c) Kemunculan bercak gejala pada buah sehat yang diinokulasi dengan potongan buah bergejala dari lapangan dan potongan buah pada perbatasan jaringan yang bergejala dan tidak bergejala hasil baiting yang digunakan sebagai inokulum pada tahapan isolasi; (d) Koloni cendawan yang tumbuh dari inokulum pada medium PDA; (e) Contoh pembentukan spora pada isolat
cendawan yang diduga P. palmivora. Pengamatan mikroskopik
untuk kemampuan membentuk spora spesifik tersebut digunakan untuk mengidentifikasi isolat cendawan yang dievaluasi sebagai isolat P. palmivora; dan (f) Contoh morfologi spora - ovoid (O) dan ellipsoid (E). ... 34 7 Variasi patogenisitas isolat P. Palmivora indigenus yang berasal dari
sentra produksi kakao di Indonesia berdasarkan luas bercak pada buah kakao klon GC7 (a,b,c), ICS60 (d,e,f), dan TSH858 (g,h,i).
Setiap buah kakao diinokulasi dengan satu isolat P. Palmivora
indigenus. Gejala dicatat pada hari ke 3 (a,d,g), 5 (b,e,h), atau 7 (c,f,i) sesudah inokulasi dengan masing-masing isolat ... 35
8 Inokulasi P. palmivora pada buah dan daun kakao. (a) Buah kakao
terinfeksi P. palmivora dari lapangan yang digunakan sebagai
sumber isolat; (b) Kultur P. palmivora dengan miselia yang aktif
tumbuh; (c) Sporangia P. palmivora; (d) Gejala infeksi P. palmivora pada buah dan (e) pada daun kakao hasil inokulasi buatan... 52
9 Persentase buah kakao klon GC 7 dan Sca 12 total yang terinfeksi P.
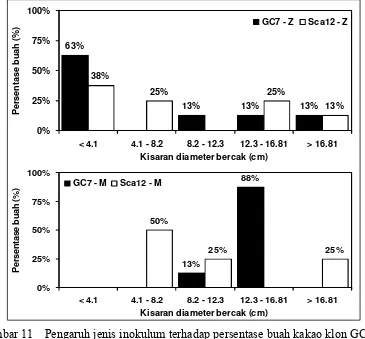
palmivora dengan kisaran diameter bercak yang ditimbulkan. Pengamatan dilakukan 7 hari sesudah inokulasi buah. ... 54 10 Pengaruh pelukaan buah terhadap persentase buah kakao klon GC 7
dan Sca 12 yang terinfeksi P. palmivora pada berbagai kisaran
2
dengan pelukaan buah sebelum diinokulasi. GC 7 – TPl dan Sca 12 – TPl: buah kakao klon GC 7 dan Sca 12 tanpa pelukaan buah sebelum diinokulasi... 55 11 Pengaruh jenis inokulum terhadap persentase buah kakao klon GC 7
dan Sca 12 yang terinfeksi P. palmivora pada berbagai kisaran
diameter bercak yang ditimbulkan ( dengan pelukaan, 7 hari sesudah inokulasi buah). GC 7 – M dan Sca 12– M: buah kakao klon GC 7 dan Sca 12 yang diinokulasi dengan miselia. GC 7 – Z dan Sca 12 – Z: buah kakao klon GC 7 dan Sca 12 yang diinokulasi dengan zoospora. ... 56 12 Representasi perkembangan luas bercak 5 klon kakao inokulasi di
laboratorium ... 75 13 Representasi perkembangan luas bercak 5 klon kakao
inokulasi di Laboratorium ... 76 14 Representasi perkembangan luas bercak tiga klon kakao PA 300,
DR 2 dan GC7 inokulasi di lapangan dan laboratorium beberapa
klon kakao terhadap penyakit busuk buah P.palmivora... 76
15 Luas bercak (cm2) hasil inokulasi di Lapangan dan Laboratorium
beberapa klon kakao terhadap penyakit busuk buah P. palmivora
Stomata daun kakao klon TSH 858, ICCRI , GC 7 dan ICS 13... 77
16 Stomata daun kakao klon TSH 858, ICCRI , GC 7 dan ICS 13... 108 17 Hubungan kerapatan stomata daun dan buah dengan luas bercak
yang disebabkan oleh infeksi P. palmivora... 109
18 Hubungan peragam (Wr) dan Ragam (Vr) 5 klon kakao sifat ketahanannya
PENDAHULUAN
Latar Belakang
Kakao (Theobromacacao L.) merupakan salah satu komoditas perkebunan
yang mempunyai peran penting dalam perekonomian Indonesia. Pada tahun 2002, dari 776 ribu ha areal kakao Indonesia, sekitar 668 ribu ha atau 86 % adalah kakao rakyat (Anonim, 2004). Hal ini mengindikasikan peran penting kakao baik sebagai sumber lapangan kerja maupun pendapatan bagi petani. Disamping itu, areal dan produksi kakao Indonesia meningkat pesat pada dekade terakhir, dengan laju 7,99% per tahun (Ditjen Perkebunan, 2008).
Volume dan nilai ekspor kakao Indonesia pada periode 1997-2002 meningkat masing-masing dengan laju 12 % dan 10,84 %/tahun, suatu pertumbuhan yang sangat pesat. Hasil penelitian juga mendukung bahwa industri kakao patut dikembangkan sebagai salah satu andalan karena mempunyai koefisien keterkaitan ke depan dan ke belakang yang lebih besar dari satu, efek
penggandaan dan lapangan kerja yang relatif besar, serta efek distribusionalnya
yang cukup baik (tersebar) (Zainudin et al., 2005). Sejalan dengan peran penting
tersebut, peluang pasar kakao Indonesia masih cukup terbuka. Potensi untuk menggunakan industri kakao sebagai salah satu pendorong pertumbuhan dan distribusi pendapatan cukup terbuka dan sangat menjanjikan.
Permintaan biji kakao terus meningkat, terutama dari Amerika Serikat dan negara-negara Eropa Barat. Berbagai Negara tersebut dikenal sebagai produsen makanan yang menggunakan kakao sebagai komponen utamanya. Indonesia sebagai salah satu produsen perlu memanfaatkan peluang tersebut untuk meningkatkan devisa negara dengan meningkatkan ekspor biji kakao. Berorientasi pada pasar ekspor, peluang besar kakao Indonesia relatif masih terbuka. Beberapa hasil studi menunjukkan bahwa daya saing produk kakao Indonesia, khususnya biji kakao masih baik sehingga Indonesia masih mempunyai peluang untuk meningkatkan ekspor dan mengembangkan pasar domestik.
4
di atas laju perdagangan kakao dunia. Pada periode 1997-2002, laju ekspor kakao (volume) Indonesia adalah sekitar 12,0% per tahun, sedangkan laju pertumbuhan dunia hanya 3,51% per tahun (Zainudin & Baon, 2004). Walaupun mempunyai kelemahan dan komposisi komoditas dan distribusi pasar, daya saing biji kakao Indonesia cukup baik yang dicerminkan dengan koefisien daya saing lebih besar dari satu (1,62). (b) Memiliki daya saing yang cukup baik, Indonesia diperkirakan akan mampu memanfaatkan peluang pasar yang masih cukup terbuka pada masa mendatang. Beberapa studi menunjukkan bahwa peluang ekspor kakao Indonesia pada periode 2000-2008 masih tumbuh dengan laju sekitar 3,3% per tahun sampai dengan tahun 2008. Laju tersebut tertinggi di antara negara eksportir dan jauh di atas rata-rata laju ekspor dunia yang hanya 1,7%. (c) Liberalisasi perdagangan juga diperkirakan akan memperkuat posisi kakao Indonesia di pasar Internasional. Beberapa negara produsen utama kakao seperti Pantai Gading dan Ghana harus mengurangi berbagai bentuk dukungan dan subsidi pada agribisnis kakaonya. Di sisi lain, agribisnis kakao di Indonesia hampir tidak diproteksi atau mendapat subsidi. Indonesia diperkirakan merupakan salah satu yang akan memperoleh manfaat liberalisasi perdagangan tersebut ( Zainudin & Baon, 2004)
Peningkatan produksi dan perbaikan mutu kakao Indonesia dapat dilakukan melalui intensifikasi dan ekstensifikasi. Penerapan kedua program tersebut di
Indonesia memerlukan tersedianya bibit dan benih kakao unggul, sehingga pengembangan kultivar atau klon kakao unggul secara terprogram perlu segera dilakukan. Umumnya bahan tanam kakao yang digunakan untuk pengembangan di Indonesia menggunakan benih hibrida F1, yang diperoleh dari kebun benih. Kebun benih dirancang khusus untuk menghasilkan benih hibrida F1, dengan menggunakan tetua (sebagai induk betina dan jantan) yang telah diketahui daya dan mutu hasilnya serta sifat-sifat penting seperti ketahanan terhadap penyakit
utama (Phytophthora palmivora dan Vascular-Streak Dieback/VSD).
Budidaya kakao menghadapi banyak kendala di lapangan, antara lain penyakit dan hama tanaman yang dapat menurunkan kuantitas dan kualitas produksi kakao. Salah satu penyakit utama pada tanaman kakao di Indonesia
adalah penyakit busuk buah (black pod) yang disebabkan oleh Phytopthora
kakao di berbagai negara penghasil kakao. Penyakit busuk buah di lapangan menyebabkan kerugian yang bervariasi besarnya antara satu daerah dengan daerah lainnya di Indonesia bahkan di antar negara. Secara umum, besarnya kerugian antara 20-30% per tahun dapat terjadi akibat infeksi penyakit busuk buah pada pertanaman kakao di lapangan (Wood & Lass, 1985). Berdasarkan data tahun 1997 dilaporkan infeksi penyakit busuk buah menyebabkan menurunnya total kakao dunia hingga sebesar 44 %/tahun (Van der Vossen, 1997).
Pengendalian penyakit busuk buah yang telah dipraktekkan di lapangan seringkali memberikan hasil yang tidak konsisten karena perkembangan penyakit di lapangan dipengaruhi oleh banyak faktor, antara lain: (i) Pertanaman kakao dibudidayakan di daerah yang mempunyai kondisi iklim cocok untuk perkembangan penyakit busuk buah, (ii) Tanaman kakao yang diusahakan pada umumnya mempunyai ketahanan sedang sampai rendah, (iii) Perkembangan sejak
penyerbukan hingga panen kakao memerlukan waktu antara 5,0-5,5 bulan, (iv) P.
palmivora dapat menyerang semua organ kakao dan serangan pada buah terjadi
pada semua tahap pertumbuhannya, (v) Inokulum P. palmivora banyak ditemukan
di lapangan sehingga pada kondisi lingkungan yang optimum untuk perkembangannya, serangan patogen busuk buah dapat terjadi sepanjang tahun,
(vi) P. palmivora diketahui mempunyai banyak tanaman inang. Dengan kondisi
agroekosistem yang sangat sesuai tersebut sangat dimungkinkan patogen P.
palmivora di daerah sentra produksi kakao di Indonesia akan menghasilkan tingkat patogenisitas yang berbeda. Hal ini semakin menambah sulitnya pengendalian secara umum terhadap patogen tersebut. Oleh karena itu diperlukan informasi isolat-isolat dari daerah yang berbeda pada sentra kakao di Indonesia untuk dapat membantu menyelesaikan masalah tersebut. Berbagai hal tersebut menjadi penguat perlunya pengembangan metode pengendalian penyakit busuk buah kakao yang efektif di lapangan.
6
ekonomis, sehingga perlu dicarikan alternatif pengendalian lain yang secara bertahap dapat mengurangi ketergantungan pada fungisida.
Pemuliaan untuk mengembangkan varietas kakao unggul yang resisten
terhadap P. palmivora sangat penting untuk dilakukan dan perlu mendapatkan
perhatian khusus jika Indonesia tetap ingin menjadi produsen terbesar komoditas ekspor ini. Tersedianya varietas tahan membantu meringankan ongkos produksi, sehingga berpotensi meningkatkan pendapatan. Jika telah dikembangkan,
penanaman kultivar unggul kakao yang resisten terhadap P. palmivora dapat
menjadi solusi terbaik yang tersedia bagi petani dan produsen kakao di Indonesia untuk mengatasi masalah penyakit busuk buah di lapangan.
Di Indonesia, arah pemuliaan tanaman kakao ditujukan untuk mengembangkan kultivar unggul dengan sifat-sifat sebagai berikut: (i) mempunyai daya hasil yang tinggi, (ii) kualitas biji bermutu tinggi, dan (iii) resisten terhadap hama seperti: penggerek buah kakao (PBK) dan penyakit busuk
buah yang disebabkan oleh P. palmivora dan VSD (Iswanto & Winarno, 1992).
Program pemuliaan untuk memperoleh kultivar resisten terhadap P. palmivora
merupakan tujuan kegiatan yang dilakukan di berbagai negara produsen kakao (van der Vossen, 1997). Namun demikian, kemajuan yang didapat untuk mencapai tujuan tersebut masih terbatas sebagai akibat (i) belum tersedianya
informasi tentang keragaman genetik/tingkat keragaman plasma nutfah kakao, (ii) belum dilakukannya strategi pemuliaan yang efektif, (iii) belum tersedia
informasi dasar tentang genetika dan mekanisme dari sifat resisten terhadap P.
palmivora yang ada pada plasma nutfah kakao. Untuk itu perlu tersedia data tentang ketahanan dari inang, dengan demikian pemahaman mengenai genetika ketahanan tanaman sangat membantu usaha pemuliaan ketahanan.
hibrida dengan sifat-sifat kualitatif dan kuantitatif yang diinginkan perlu dilakukan studi pendugaan daya waris terhadap klon-klon yang ada dengan melakukan persilangan dialel. Dengan demikian informasi tentang daya gabung umum (DGU), daya gabung khusus (DGK), heterosis serta dayawaris menjadi sangat penting untuk tanaman kakao yang heterosigot.
Perumusan Masalah
Penyakit busuk buah kakao yang disebabkan oleh P. palmivora merupakan
penyakit penting di Indonesia. Pengembangan kakao di daerah – daerah sentra kakao sangat riskan sebagai tempat endemik penyakit ini. Mengingat banyak tanaman inang dari patogen tersebut, sangat dimungkinkan isolat dari daerah akan memberikan tingkat patogenisitas yang berbeda. Isolat yang diambil dari pertanaman kakao di Indonesia akan menjadi informasi yang penting untuk membantu merakit kultivar yang resisten terhadap patogen ini.
Jenis kakao yang ditanam menunjukkan tingkat ketahanan yang berbeda
terhadap P. palmivora. Umumnya seleksi ketahanan kakao terhadap penyakit
busuk buah dilakukan dengan inokulasi alami ataupun buatan, yang didasarkan pada jumlah organ sakit dan keparahan penyakit (Rocha, 1974). Indikator ini menunjukkan reaksi jaringan terhadap serangan patogen, tetapi tidak mengungkapkan secara tepat mekanisme ketahanan yang bekerja pada satu atau
beberapa tahap dari daur penyakit busuk buah. Pengujian ketahanan dilakukan
pada buah yang dipetik (detachedpod) maupun buah di pohon (attachedpod). Uji
pertama banyak diminati, namun hasilnya kurang sesuai dengan kondisi lapangan karena uji ini mengabaikan pengaruh lingkungan. Dengan demikian metode inokulasi yang baku sangat penting dilakukan untuk mendapatkan metode uji ketahanan yang cepat dan akurat. Hal ini sangat membantu dalam proses seleksi tanaman kakao yang berumur panjang. Disamping itu, hal lain yang penting adalah tersedianya plasma nutfah yang cukup.
Mekanisme ketahanan tanaman terhadap penyakit dibedakan atas mekanisme struktural dan biokimia (Agrios, 1998). Kedua mekanisme tersebut
dapat berperan dalam ketahanan sebelum penetrasi (preexisting defense) dan
8
kakao melalui mulut kulit (Tarjot, 1974), namun terdapat laporan kontradiktif mengenai peran mulut kulit sebagai mekanisme struktural ketahanan kakao
terhadap patogen ini (Tarjot, 1972; Iwaro et al., 1997; Iwaro et al., 1999;
Phillips-Mora, 1999). Permukaan buah kakao mempunyai alur primer yang diperkirakan dapat mempengaruhi penyebaran, deposisi, dan pertumbuhan pra-penetrasi inokulum. Mekanisme biokimia tergantung pada reaksi biokimia yang terjadi dalam sel tanaman. Seperti protein yang berhubungan dengan respon ketahanan tanaman terhadap patogen adalah kitinase dan peroksidase. Kitinase dapat mendegradasi senyawa kitin yang merupakan komponen utama penyusun dinding sel cendawan. Sebagian besar cendawan filamentus mengandung senyawa kitin pada dinding sel hifanya (Kasprzewska, 2003). Peroksidase merupakan enzim yang terlibat dalam respon tanaman terhadap patogen dan termasuk ke
dalam PR-9 (pathogenesis related protein) (Lagrimini et al., 1997). Aktivitas
peroksidase yang tinggi pada tanaman terkait dengan ketahanan tanaman yang lebih tinggi terhadap patogen seperti yang pernah dilaporkan pada kacang tanah (Pujihartati et al., 2006).
Berdasarkan uraian di atas dirumuskan bahwa mekanisme ketahanan tanaman kakao terhadap penyakit busuk buah dibedakan atas mekanisme ketahanan struktural dan mekanisme biokimia. Mekanisme struktural diarahkan
pada kerapatan mulut kulit buah dan mulut daun kakao. Mekanisme ketahanan biokimia diarahkan pada aktivitas enzim kitinase dan peroksidase.
melakukan uji ketahanan terhadap beberapa plasma nutfah kakao yang ada, bertujuan untuk mendapatkan genotipe yang tahan dan rentan, juga dapat digunakan sebagai tetua sebagai langkah awal untuk merakit bahan tanam kakao yang baru.
Terkait untuk mengidentifikasi pasangan tetua yang dapat menghasilkan hibrida dengan sifat-sifat kualitatif dan kuantitatif yang diinginkan, perlu dilakukan studi pendugaan daya waris terhadap klon-klon yang ada dengan melakukan persilangan dialel. Untuk merakit hibrida unggul informasi tentang daya gabung umum (DGU) dan daya gabung khusus (DGK), heterosis, heritabilitas atau daya waris menjadi sangat penting untuk tanaman kakao yang heterosigot.
Tujuan Penelitian
1. Memperoleh identitas spesies P. palmivora dan informasi keragaman
patogenisitasnya pada lokasi sentra kakao beberapa propinsi di Indonesia.
2. Memperoleh metode inokulasi untuk penapisan ketahanan kakao
3. Memperoleh informasi patogenisitas P. palmivora terhadap tanaman kakao.
4. Mendapatkan tanaman kakao yang tahan dan rentan terhadap penyakit P.
palmivora di koleksi plasma nutfah kakao untuk digunakan sebagai tetua untuk perakitan hibrida.
5. Memperoleh informasi korelasi tingkat ketahanan beberapa klon kakao untuk
karakter kwantitatif yang diamati.
6. Memperoleh informasi tentang potensi daya gabung umum dan khusus yang
tinggi serta efek heterosisnya pada persilangan dialel klon kakao sehingga diperoleh potensi pada hibrida.
Manfaat Penelitian
Penelitian genetika ketahanan kakao terhadap penyakit P. palmivora ini
dapat membantu usaha pemuliaan untuk memperoleh bahan tanam kakao yang
tahan terhadap penyakit P. palmivora. Dengan mendapatkan klon unggul yang
10
diharapkan akan dapat memperbaiki mutu hasil dan peningkatan produktivitas kakao nasional.
Penelitian mengenai kajian genetika ketahanan tanaman kakao terhadap penyakit busuk buah dapat membantu usaha pemuliaan untuk memperoleh bahan tanam yang tahan terhadap penyakit busuk buah dan model pewarisan terhadap F1 nya serta menjadi informasi yang penting untuk menghasilkan bahan tanam yang mempunyai beberapa sifat unggul, sehingga produksi kakao dapat ditingkatkan.
Manfaat lain adalah memberikan alternatif pengendalian yang diharapkan saling melengkapi dengan pengendalian kimiawi sehingga secara bertahap penggunaan fungisida dapat dikurangi dan biaya pemeliharaan menjadi lebih murah.
Hipotesis
Berdasarkan telaah beberapa pustaka tersebut di atas penelitian ini disusun dengan beberapa hipotesis sebagai berikut:
1. Isolat P. palmivora yang menyerang kakao di Indonesia adalah salah satu
spesies P. palmivora.
2. Terdapat satu metode penapisan yang efisien dalam menentukan derajat
ketahanan kakao.
3. Terdapat sedikitnya satu isolat P. palmivora yang mempunyai tingkat
patogenitas tinggi, dan dapat digunakan sebagai sumber inokulum untuk uji ketahanan.
4. Tetua yang membawa gen sumber ketahanan dapat dicari dengan cara
identifikasi tingkat ketahanan beberapa klon kakao pada koleksi plasma nutfah kakao.
5. Terdapat beberapa karakter kuantitatif struktur tanaman kakao yang
memcerminkan ketahanan terhadap P. palmivora.
6. Terdapat tetua yang mempunyai daya gabung umum dan daya gabung khusus
A. Morfologi Tanaman Kakao
Tanaman kakao yang mempunyai nama ilmiah Theobroma cocoa L.
merupakan anggota dari familia Sterculiaceae (Wood, 1975; Tjitrosoepomo,
1988). Kakao merupakan jenis tanaman asli hutan hujan tropis Amerika Selatan (Wood, 1975) dan telah lama dibudidayakan di Indonesia yaitu sejak jaman “culturstelsel” tahun 1826 (Sunaryo & Situmorang, 1978). Diperkirakan kakao
berasal dari hulu sungai Amazon, tempat Theobroma dan jenis sekerabatnya
terdapat dalam populasi yang paling besar. Tanaman kakao tersebut merupakan
satu-satunya species diantara 22 jenis dalam genus Theobrama yang diusahakan
secara komersial. Sistematika tanaman kakao secara lengkap dapat
diklasifikasikan dalam taksa-taksa sebagai berikut: Divisio: Spermatophyta, Sub
divisio: Angiospermae, Klassis: Dicotyledoneae, Ordo: Malvales, Familia:
Sterculiaceae, Genus: Theobroma, Spesies: Theobroma cocoa, L. (Cheesman,
1944).
Sebagai tanaman yang masuk dalam anggota dari klas Dicotyledonae, benih tanaman kakao mempunyai tipe perkecambahan yang epigeus yang pada waktu kecambah daun kotilnya terangkat ke atas serta membentuk akar tunggang yang tumbuh lurus ke bawah masuk ke dalam tanah (Prawoto, 1991). Sedangkan akar lateralnya banyak tumbuh dan berkembang di dekat permukaan tanah pada kedalaman sekitar 0-30 cm.
Pertumbuhan batang kakao bersifat dimorfisme yang berarti mempunyai dua macam bentuk pertumbuhan batang, yaitu pertumbuhan batang utama yang bersifat ortotrop yang tumbuh tegak dengan rumus daun 3/8, dan pertumbuhan ke samping seperti cabang primer disebut plagiotrop, mempunyai rumus daun ½ (Prawoto, 1991).
10
pangkal daun dan ujung tangkai daun. Adanya persendian ini memungkinkan daun membuat gerakan untuk menyesuaikan dengan arah datangnya sinar matahari (Prawoto, 1991). Pertumbuhan daun pada cabang plagiotrop berlangsung serempak dan berkala. Tunas baru disebut dengan flush, dan pada saat flush setiap tunas dapat membentuk 4-6 lembar daun baru sekaligus.
Tanaman kakao bersifat kaolifloris yang berarti bunga dan buahnya tumbuh dan berkembang pada batang atau cabang. Sifat penyerbukan kakao adalah
menyerbuk silang. Bekas ketiak daun, tempat tumbuhnya bunga atau buah
tersebut lama kelamaan menebal dan membesar disebut dengan bantalan bunga atau bantalan buah. Bunga kakao mempunyai rumus K5C5A5+5G(5). K5 berarti
bunga tersusun atas 5 daun kelopak yang bebas satu sama lainnya. C5 bunga
kakao memiliki daun mahkota yang lepas atau tidak berlekatan satu sama lain.
A5+5 berarti bunga kakao memiliki 10 tangkai sari yang tersusun atas dua
lingkaran masing-masing lingkaran tersusun atas 5 tangkai sari steril yang disebut
dengan staminodia dan 5 tangkai sari yang fertil. G5 bunga kakao mempunyai 5
daun buah yang bersatu (Lass & Wood, 1985).
Bentuk buah kakao bervariasi, dari bulat ke lonjong dan meruncing dengan permukaan yang halus sampai kasar. Permukaan buah kakao mempunyai alur primer dan alur sekunder. Sering kali alur sekunder tidak tampak. Permukaan
buah kakao berlilin, kaku (rigid), mempunyai rambut-rambut tegak dan mulut
kulit yang agak terangkat (Cuatrecasas, 1964). Pengelompokan kakao dapat didasarkan pada bentuk buah (Pound, 1932), gabungan karakteristik buah dan sebaran geografi (Cheesman, 1944), bentuk buah dan struktur permukaan buah (Ostendorf, 1956; Engels, 1986). Bentuk buah kakao tersebut antara lain: amilonado, cundeamor, angoleta, calabasilo, criolo dan pentagona.
Buah kakao mempunyai karakteristik termodinamika yang menarik. Waktu siang hari buah menjadi panas dan dingin waktu malam hari, menjelang dini hari suhu buah sama dengan suhu lingkungan. Peningkatan suhu udara pada dini hari memacu kondensasi uap air pada seluruh permukaan buah yang dapat menjadi
suatu inkubator mikro yang baik bagi perkecambahan spora patogen termasuk P.
palmivora (Fulton, 1989).
(i) Epikarp, terdiri atas: lapisan epidermis dengan mulut kulit dan trikoma. Parenkim dengan sel yang relatif kecil (diameter 10-20 µm), terbagi dalam 2 zona, yaitu: lapisan luar yang tidak mengandung klorofil, terdiri atas 2-4 lapis sel, lapisan dalam yang mengandung klorofil, terdiri atas 6-12 lapis sel. Parenkim dengan sel yang relatif besar. Diameter sel paling luar 30-40 µm dan bertambah kearah dalam. (ii) Mesokarp, terdiri atas sel yang agak berserat dan (iii) Endokarp, terdiri atas sel parenkim yang besar dengan berkas pengangkutan (Tarjot, 1974).
B. Keragaman Genetik Tanaman Kakao
Menurut Las & Wood (1985) berdasarkan type populasinya, tanaman kakao dapat dibagi menjadi tiga kelompok besar yaitu tipe Criolo, Forastero dan Trinitario. Criollo berasal dari penyebaran melintasi pegunungan Andes ke arah dataran rendah Venezuela, Kolumbia, dan Ekuador, dan ke arah utara ke Amerika Tengah dan Meksiko. Sifat-sifat tipe Criolo antara lain pertumbuhan tanaman kurang kuat, daya hasilnya lebih rendah dibanding Forastero, dan relatif lebih rentan terhadap gangguan hama dan penyakit. Kulit buahnya tebal tetapi lunak
sehingga mudah dibelah. Criollo menghasilkan kakao mulia (fineflavourcocoa).
Warna buah hijau atau agak merah karena adanya pigmen antosianin; perikarp agak kasar, tipis dan lunak, mesokarp mengandung lignin, biji bulat dan kotiledon putih. Kelompok ini cenderung rentan terhadap penyakit (Soria, 1974; Opeke,
1982). Kadar lemak di dalam biji lebih rendah dibandingkan dengan Forastero tetapi ukuran bijinya lebih besar, bulat, memberikan citarasa khas yang unggul. Dalam tataniaga kakao Criolo termasuk dalam jenis kakao mulia, sedangkan tipe Forastero termasuk dalam jenis kakao lindak.
Forastero dihasilkan oleh penyebaran ke lembah Amazon, ke arah Brazil bagian barat dan Guyana (Alvim, 1997). Forastero menghasilkan kakao bermutu
sedang, dikenal dengan kakao lindak (bulk cocoa). Warna buah hijau, tidak ada
12
[image:37.595.121.474.327.540.2]Tipe Trinitario merupakan hibrida antara Criolo dan Forastero. Sifat morfologi dan fisiologinya sangat beragam, demikian pula sifat daya hasil dan mutu hasilnya. Dalam tataniaga kakao kelompok Trinitario termasuk dalam kakao mulia atau kakao lindak tergantung dari mutu biji yang dihasilkannya. Seperti klon DR menghasilkan kakao mulia, sedangkan klon ICS banyak menghasilkan kakao lindak (Mawardi, 1982; Opeke, 1982). Trinitario mempunyai buah berwarna merah atau hijau dan bervariasi, tekstur keras; warna biji bervariasi dari ungu muda sampai ungu tua (Wood & Lass, 1985). Pertumbuhan pohon gigas. Contoh kelompok ini adalah klon-klon ICS 60, ICS 84, ICS 95, DR 1, DR 2, DR 38, dan DRC 16.
Gambar 1. Beberapa tipe kakao Trinitario yang berkembang di Indonesia.
Selanjutnya Lanaud (1987; Laurent, 1993; N’Goran, 1994 cit, Sounigo et
al., 2000) memisahkan kelompok Forastero, antara genotip yang berasal dari
lembah hulu sungai Amazon dan lembah hilir sungai Amazon. Trinitario lebih dekat ke genotip Amazone hilir daripada Amazone hulu.
C. Penyakit Busuk Buah Kakao
Busuk buah (black pod atau pod rot) merupakan penyakit yang paling
merugikan di banyak negara produsen kakao. Masalah penyakit ini bisa bersifat ICS 95
ICS 13
lokal, regional atau bahkan global. Busuk buah pada kakao terutama disebabkan Phytophthora palmivora (Butl.) Butl. Sejak tahun 1979, setelah Brasier dan
Griffin mempublikasikan kajian taksonomi Phytophthora, diketahui ada spesies
lain yang patogenik terhadap kakao, tetapi hanya menimbulkan masalah lokal ataupun regional, misal P. arecae di Vanuata, P. capcisi di Kamerun dan Brazil, P. citrophthora di Brazil, P. faberi dan P. megakarya di Afrika Barat (Zadock, 1997). Busuk buah kakao di Indonesia, Malaysia, dan Papua New Guinea
disebabkan oleh P. palmivora (Waterhouse, 1974; Prior, 1992; van der Vossen,
1997). Sebagai patogen tropika, berdasarkan penyebaran inang aslinya,
diperkirakan P. palmivora berasal dari Amerika Tengah/Selatan atau Indo-Pasifik
(Zentmyer, 1988).
Terdapat tiga bentuk morfologi P. palmivora yang dapat menyebabkan
penyakit pada kakao, namun sekarang bentuk morfologi tersebut mewakili tiga jenis yang berbeda, yaitu P. capsici, P. megakarya, dan P. palmivora, dan ada
pula tiga jenis tambahan yang dapat menyebabkan penyakit ini, yaitu P. heveae,
P. megasperma, dan P. citrophthora (Thurston, 1998).
Menurut Chee (1974), P. palmivora mempunyai 138 jenis tumbuhan inang,
antara lain karet, lada, kelapa, sukun, pala, jeruk, kapas, pepaya, anggrek, mangga, alpokat, dan durian. Patogen ini dapat menyerang semua organ atau bagian
tanaman kakao, seperti akar, daun, batang, ranting, bantalan bunga, dan buah pada semua tingkatan umur. Serangan pada buah paling merugikan dan di Indonesia penyakit ini perlu mendapat perhatian (Opeke & Gorenz, 1974; Pawirosoemardjo & Purwantara, 1992).
Selama daur hidupnya, P. palmivora menghasilkan beberapa inokulum yang
14
Gambar 2. (A) Sporangium P. palmivora berbentuk ovoid dengan pedisel
yang jelas. (B) Zoospora
P. palmivora bertahan sebagai klamidospora dalam tanah dan miselium
pada bantalan bunga, buah muda (cherelle), batang pohon kakao, dan sisa-sisa
tanaman yang tersebar di tanah. Busuk buah dapat berasal dari inokulum yang bertahan di tanah, sisa-sisa tanaman, bantalan bunga, kulit, kanker batang, tangkai
buah, buah muda (cherelle), buah dan tangkai daun. Peran masing-masing sumber
inokulum tersebut berbeda antar daerah atau negara. Umumnya tanah dan akar berperan sebagai sumber inokulum primer yang memberikan inokulum infektif pada awal musim hujan untuk mulainya epidemi busuk buah; sedangkan buah dan bagian kanopi yang sakit berperan sebagai sumber inokulum sekunder dan berhubungan langsung dengan kehilangan hasil (Pereira, 1995).
Epidemi penyakit busuk buah kakao terjadi akibat penyebaran vertikal
(dalam satu pohon) dan horizontal (antar pohon) inokulum P. palmivora.
Penyebaran vertikal terjadi akibat kontak langsung antara buah sakit dan buah sehat, penyebaran inokulum oleh tetesan air hujan dari buah sakit ke buah sehat di bawahnya, bantuan serangga vektor, dan percikan air hujan dari tanah ke buah di sekitar pangkal batang. Penyebaran horizontal dapat terjadi dengan bantuan serangga, kontak antar pohon, angin (Muller, 1974). Penyebaran horizontal lebih lambat dibandingkan dengan penyebaran vertikal (Gambar 3).
Gambar 3. (A) Infeksi P. palmivora pada buah di pohon dan (B) serangan P. palmivora
pada bibit.
Penyakit busuk buah sukar dikendalikan karena epidemiologi penyakit ini kompleks dan belum dapat diungkapkan secara tuntas (Gregory & Maddisson, 1981 cit. Tey, 1991), pembuangan sumber inokulum primer yang terdapat di pohon (buah sakit dan kanker batang) maupun di tanah (serasah dan kulit buah) tidak menyebabkan penundaan terjadinya epidemi pada musim hujan. Hal ini
menunjukkan adanya sumber inokulum lain yang memperbesar deposit (pool)
inokulum primer (Dennis & Konam, 1994).
Perkembangan busuk buah dipengaruhi oleh kelembapan udara.
Kelembaban udara 80-95% selama 2-4 jam mendukung infeksi spora kembara P.
palmivora. Ada interaksi antara curah hujan, keragaan (performance) tanaman dan penyakit. Busuk buah berhubungan langsung dengan jumlah buah di pohon dan curah hujan, namun jumlah buah berbanding terbalik dengan curah hujan (Thorold, 1975).
Menurut Purwantara (1990); Purwantara & Pawirosoemardjo (1992) kebasahan permukaan buah dan kelembaban udara berperan langsung terhadap
infeksi P. palmivora pada buah kakao. Peranan curah hujan terjadi secara tidak
langsung melalui terjadinya kebasahan permukaan buah dan meningkatnya kelembaban udara. Pengaruh suhu terhadap perkembangan infeksi terjadi secara tidak langsung, melalui pengaruhnya pada kelembaban udara dan kebasahan buah.
16
D. Pengendalian Penyakit Busuk Buah
Pengendalian kultur teknik merupakan bagian penting dalam pengelolaan penyakit busuk buah. Cara ini meliputi pembuangan gulma dan epifit, pemangkasan, pengaturan jarak tanaman dan manipulasi naungan. Cara ini dapat memperbaiki sirkulasi udara dan mengurangi kelembaban tajuk dan membatasi insiden busuk buah (Akrofi & Opuku, 2000). Pengurangan sumber inokulum dapat dilakukan dengan membuang kulit buah yang tersebar di tanah, buah kering (mummifiedpod) dan buah sakit pada pohon (Muller, 1974).
Pengendalian serangga vektor, seperti Drosophila dengan insektisida dapat
mengurangi kerugian akibat busuk buah (Muller, 1974). Kumbang (Coleoptera:
scolytidae dan nitidulidae) mempunyai peranan cukup penting dan penyebaran
inokulum P. palmivora di Papua New Guinea, sehingga mempunyai implikasi
penting dalam strategi pengendalian penyakit (Konam et al., 2000).
Penyemprotan fungisida merupakan cara pengendalian busuk buah kakao yang penting sejalan dengan intensifikasi pengusahaan tanaman ini. Umumnya
dipergunakan fungisida tembaga seperti bubur Bordeaux, tembaga-oksida,
tembaga-oksiklorida, dan tembaga-hidroksida (Thorold, 1975). Berdasarkan pengalaman di banyak negara selama 25 tahun, Gorenz (1974) menyatakan penyemprotan fungisida kontak sering kali memberikan hasil yang tidak konsisten
dan tidak menguntungkan. Keefektivfan fungisida kontak tergantung pada meratanya deposit bahan tersebut pada permukaan buah. Keadaan ini sukar dicapai karena adanya lapisan lilin pada permukaan buah, bentuk dan letak buah di pohon (Gorenz, 1974) dan pertumbuhan buah yang cepat (Thorold, 1975). Oleh karena itu diperlukan fungisida sistemik yang dapat memberikan perlindungan pada tanaman.
Fungisida sistemik yang efektif mengendalikan busuk buah dan kanker batang kakao dan memberikan perlindungan lama pada buah adalah fosfonat
(Brown et al., 1997; Pereira, 1995). Fungisida ini efektif terhadap jenis jamur
dalam bangsa Peronosporales. Dosis anjuran 2,5 – 5,0 gram bahan aktif per liter air (Schwin, 1983). Sebagai fungisida sistemik, fosfonat diformulasi dalam bentuk
fosetyl-Al dan kalium fosfonat. Fosfonat dapat menurunkan virulensi P.
inang (Akrofi & Opoku, 2000). Fosfonat mempunyai residual activity selama sepuluh bulan sehingga melindungi pohon dan buah selama satu musim
(Anderson et al., 1989 cit. Akrofi & Opoku, 2000). Namun injeksi fosfonat
menyebabkan gejala terbakar (scorching) jaringan internal batang dan kulit batang menjadi retak. Sampai sekarang teknologi ini tidak dapat ditransfer ke petani di Indonesia, meskipun telah diterapkan secara luas di Papua Nugini (Akrofi & Opoku, 2000).
Penanaman varietas tahan merupakan cara pengendalian penyakit yang paling bermanfaat karena cara ini ramah lingkungan (Akrofi & Opoku, 2000). Varietas dengan tingkat ketahanan tertentu yang lebih mudah ditemukan di antara bahan tanam yang ada atau yang dihasilkan melalui hibridisasi merupakan cara terbaik untuk mengatasi busuk buah kakao (Muller, 1974).
Menurut Muller (1974), ketahanan kakao terhadap penyakit busuk buah
dibedakan atas ketahanan sejati (true resistance) dan ketahanan semu (false
resistance) atau disease avoidance. Ketahanan pertama merupakan hasil dari karakteristik anatomi, fisiologi dan biokimia, sedangkan ketahanan kedua hasil dari karakteristik fenologi pohon sehingga terhindar dari infeksi P. palmivora.
Penggunaan bahan tanam tahan dapat memperlambat perkembangan epidemi penyakit (Campbell & Madden, 1990). Berdasarkan epidemilogi,
ketahanan tanaman dapat bekerja dengan cara berikut: a) reduksi jumlah infeksi, b) reduksi laju perluasan bercak, c) reduksi sporulasi patogen, d) memperpanjang masa inkubasi, dan e) reduksi deposisi spora (Berger, 1977).
E.Mekanisme Ketahanan
18
resistance) yang didasarkan pada satu atau lebih komponen ketahanan yang dapat atau tidak dapat berkorelasi satu sama lain.
Bahan tanam tahan terhadap penyakit ini merupakan pemecahan masalah tersebut untuk jangka panjang. Simmonds (1994) menyatakan bahwa ketahanan
buah kakao terhadap P. palmivora diperkirakan lebih bersifat horizontal daripada
vertikal. Menurut Agrios (1997) ketahanan tanaman dapat bersifat pasif (terbentuk tanpa rangsangan dari patogen) atau aktif (ekspresinya diimbas oleh serangan patogen), melibatkan mekanisme struktural dan biokimia. Duniway (1983)
menyatakan bahwa ketahanan tanaman terhadap Phytophthora spp. meliputi
ketahanan struktural, penghalang struktural terimbas, reaksi hipersensitif, dan produksi senyawa antimikrobia.
Ketahanan buah kakao terhadap P. palmivora merupakan sistem
multikomponen yang terekspresi dalam dua tahap, dinyatakan sebagai ketahanan prapenetrasi dan pascapenetrasi. Ketahanan prapenetrasi berhubungan dengan faktor morfologi yang mempengaruhi perkembangan prapenetrasi dan penetrasi patogen, dan menentukan jumlah bercak yang terjadi. Ketahanan pasca penetrasi berhubungan dengan mekanisme biokimia yang dapat mempengaruhi luasnya
jaringan yang diserang patogen (Irwaro et al., 1995). Fry (1982) menyatakan
bahwa walaupun patogen berhasil mempenetrasi jaringan inang, sering kali
perkembangan selanjutnya terhambat.
F. Mekanisme Ketahanan Struktural
Mekanisme ketahanan struktural dapat berupa sifat morfologi dan anatomi. Menurut Fry (1982) walaupun sering kali mekanisme ketahanan bekerja setelah jaringan terpenetrasi, karakteristik struktural dapat mempengaruhi ketahanan inang. Fulton (1989) memperkirakan morfologi buah kakao berpengaruh pada
disposisi dan penyebaran efektif inokulum P. palmivora. Permukaan buah kakao
dapat menjadi inkubator mikro yang baik bagi pertumbuhan spora P. palmivora.
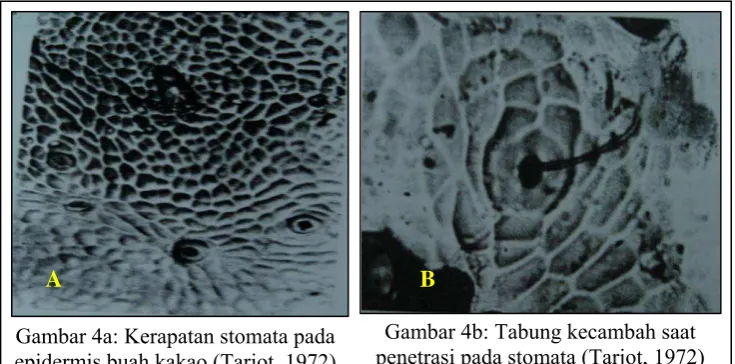
Penelitian Tarjot (1972) menunjukkan bahwa jumlah mulut kulit dan rambut-rambut pada epidermis tidak berkorelasi dengan ketahanan buah kakao
terhadap P. palmivora. Patogen ini selalu dapat melakukan penetrasi ke dalam
jaringan buah rentan maupun tahan. Diperkirakan ketahanan terhadap patogen ini terletak pada beberapa lapisan sel parenkima di bawah epidermis (Gambar 4).
Phillips-Mora (1999) menyatakan bahwa hubungan antara jumlah, panjang, lebar, panjang x lebar dan panjang/lebar mulut kulit (stomata) tidak dapat
menjelaskan ketahanan kultivar kakao terhadap P. palmivora, meskipun ada
perbedaan nyata antar kultivar. Kultivar tahan (P 7) dan moderat (UF 668) mempunyai jumlah mulut kulit terbanyak, sebaliknya CATIE 1000 (tahan) dan P 12 (rentan) mempunyai jumlah mulut kulit yang lebih sedikit.
Flores (1989 cit. Enriquez & Soria, 1999) yang mengkaji hubungan Monilia roreri dan T. cacao menunjukkan tidak ada perbedaan anatomi eksternal antara
buah kakao tahan dan rentan. Hasil penelitian Iwaro et al. (1997) menunjukkan
adanya korelasi nyata antara ketahanan penetrasi (jumlah bercak) dengan kerapatan mulut kulit dan panjang pori. Ketahanan ini tidak berkorelasi dengan lapisan lilin pada permukaan epidermis, ketebalan, kekerasan, dan kandungan lengas perikarp. Ciri morfologi buah tidak berkorelasi dengan ketahanan pasca penetrasi, ini menunjukkan kemungkinan peran mekanisme biokimiawi.
Enriquez & Soria (1999) menunjukkan bahwa setiap buah kakao yang tahan
terhadap M. roreri mempunyai cellular arrangement parenkim sub epidermis
[image:44.595.129.497.205.387.2]Gambar 4a: Kerapatan stomata pada epidermis buah kakao (Tarjot, 1972)
Gambar 4b: Tabung