(Scylla serrata
Forsskal) PADA BERBAGAI SALINIT AS MEDIA
DAN EVALUASINYA PADA SALINITAS OPTIMUM DENGAN
KADAR PROTEIN PAKAN BERBEDA
MUHAMMAD YUSRI
KARIM
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Kinerja Pertumbuhan Kepiting Bakau Betina (Scylla serrata ForsskaI) pada Berbagai Sarinitas Media dan Evaluasinya pada Salinitas Optimwn dengan Kadar Protein Pakan Berbeda adalah karya saya sendiri dan belwn diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
iエsARjセセセ@
er 2005.MUHAMMAD YUSR! KARIM. Kinerja Pertumbuhan Kepiting Bakau Betina (Scylla serrala Forsskall pada Berbagai Salinitas Media dan Evaluasinya pada Salinitas Optimum dengan Kadar Protein Pakan Berbeda. Dibimbing olch BAMBANG KIRANADI, RlDWAN AFFANDI, WASMEN MANALU dan DEWI APR! ASTUTI.
Salinitas merupakan salab satu faktor lingkungan penting yang berpengaruh pada kelangsungan hidup dan pertumbuhan kepiting bakau. Penelitian ini bertujuan mengkaji kinelja pertumbuhan kepiting bakau yang dipelihara pada berbagai salinitas dan menentukan salinitas optimum yang menghasilkan pertumbuhan kepiting bakau yang maksimal. Penelitian dilakaanakan di Balai Budidaya Air Payau (BBAP) Takalar, Sulawesi Selatan mulai bulan Oktober 2003 sampai Februari 2005.
Penelitian dilaksanakan dalam 4 tabapan percobaan yaitu (I) pengaruh salinitas pada osmoregulasi kepiting bakau, (2) pengaruh salinitas pada metabolisme kepiting bakau, (3) pengaruh salinitas pada kelangsungan hidup dan pertumbuhan kepiting bakau, dan (4) pengaruh kadar protein pakan pada kelangsungan hidup dan pertumbuhan kepiting bakau. Penelitian dirancang dengan pola nmcangan acak lengkap dengan 4 perlakuan salinitas (5, 15, 25, dan ppt) dan 3 ulangan Salinitas terbail< yang diperoleh diaplil<asikan pada percobaan penggunaan kadar protein pakan 30, 35, 40, dan 45%. Data dianalisis dengan menggunakan sidik ragam dan dilanjutkan dengan uji respon. Uji Tukey digunakan untuk membandingkan perbedaan antarperlakuan.
Salinitas media berpengaruh pada osmoregulasi. metabolisme. tingkat kelangsungan hidup, pertumbuhan, dan molting kepiting bakau. Salinitas 25 ppt menghasilkan tingkat kerja osmotik kepiting bakau paling rendah. meningkatkan derajat kelangsungan hidup, laju pertumbuhan bobot spesifik harian, pertumbuhan lebar karapas, produksi biomassa, retensi Hーイッエ・ゥセ@ lemak, energi. kalsiurn dan fosfor), dan mempersingkat periode waktu antarmolting. Laju metabolisme basal tertinggi dibasilkan pada media bersalinitas 5 ppt, sedangkan laju metabolisme kenyang dan rutin tertinggi dihasilkan pada media bersalinitas 25 ppt. Laju metabolisme kepiting bakau menurun dengan peningkatan bobot hadan kepiting bakau.
MUHAMMAD YUSRI KARIM. Growth Perfonnance of Female Mud Crab (Scylla serrata Forsskal) at Various Medium Salinity and Evaluation its in Optimum Salinity at Various of Feeding Protein Levels. Under the supervision of BAMBANG KIRANADI, RIDWAN AFFANDI, WASMEN MANALU, and DEWI APR! ASTUTI.
Salinity is one of environmental important factors
that
affect the survival and growth rate of mud crab. The objective of this experiment was to study the growth performance of mud crab cultured at different salinities and to detennine the optimum salinity which produce maximum growth. The experiment was carried out at the Center of Brackishwater Aquaculture Development, Takalar, South Sulawesi from October 2003 until February 2005.The experiment consisted of four parts (1) the effect of salinity on osmoregulation of mud crab, (2) the effect of salinity on the metabolism of mud crab, (3) the effect of salinity on the survival and growth rate of mud crab, and (4) the effect of protein level on the survival and growth rate of mud crab.
Complete randomized design with 4 treatments (salinity with 5, 15,25, and 35 ppt) and 3 replications was done in this study. The best salinity was applied for the treatments of protein level, 30, 35, 40, and 45%. Analysis of data using variance analysis and continue with response test. Tukey test was used to compare the different of mean among the treatments.
Medium salinity affected the osmoregulation, metabolism, survival rate, groMh, and moulting. Salinity 25 ppt produced low osmotic regulation, increased survival rate, spesific growth rate, carapace width, biomass production, retension (protein, lipid, energy, calcium, and phosphor), and shorter molting time period. The highest basal metabolic rate was found at salinity 5 ppt, while feeding and routine metabolic rate was at salinity 25 ppt. Metabolic rate of mud crab became lower with the increase of the body weight.
(Scylla serrata Forsskal) PADA BERBAGAI SALINITAS MEDIA
DAN EVALUASINY A PADA SALINITAS OPTIMUM DENGAN
KADAR PROTEIN PAKAN BERBEDA
MUHAMMAD YUSRI KARIM
Disertasi
sebagai salah satu syarat untuk memperoleb gelar Doktorpada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Forsskal) pada Berbagai Salinitas Medi. dan Ev.luasiny.
Nam. NIM
pad. Salinitas Optimum dengan Kadar Protein Pakan Berbeda : Muhammad Yusri Karim
: G426010031
Disetujui
Komisi Pembimbing
Mu.; ...
c.Y;
Dr. Bambang Kiranadi. M.Sc. Ketua
Prof Dr. Jr. Wasmen Manalu Anggota
Ketua Program Sludi Biologi
Dr. Ir. Dedy Duryadi. DEA
Tanggal Ujian : 30 Agustus 2005
andi,PEA. Anggota
Dr. Ir. Hi. Dew; ADri Astuti, M.S. Anggota
Diketahui
afiida Manuwoto, M.Sc.
Puji syukur penulis panjalkan ke hadirat Allah SWT alas segaJa karunia-Nya serungga karya iJmiah ini berhasil diseJesaikan. Tema yang dipilih daJam penelitian yang diJaksanakan sejak bulan Oktober 2003 sampai Februari 2005 iaJah kepiting bakau, dengan judul Kinerja perturnbuJum Kepiting Bakau (Scylla serrata Forsskal) pada Berbagai Salinitas Media dan Evaluasinya pada Salinitas
Optimum dengan Kadar Protein Pakan Berbeda.
Selesainya karya ilmiah ini tidak Jepas dari hantuan dan dorongan dari
berbagai pihak. Melalui prakata ini penulis mengucapkan terima kasih kepada:
1 Rektor Universitas Hasanuddin dan Dekan Fakultas !lmu Kelautan dan
Perikanao, Universitas Hasanuddin, Makassar yang telah memberikan
kesempatan mengikuti pendidikan Program Doktor pada SekoJah Pascasarjana Institut Pertanian Bogor.
2 Rektor Institut Pertaruan Bogor dan Dekan Sekolah Pascasarjana, Institut
Pertanian Bogor yang berkenan menerima penulis melanjutkan pendidikan
Program Doktor.
3 Departemen Pendidikan Nasional Repuhlik Indonesia dalam hal ini Direktorat
Jenderal Pendidikan Tinggi yang telah membiayai penulis selama mengikuti
pendidikan Program Doktor di Institut Pertanian Bogor lewat BPPS.
4 Bapak Dr. Bambang Kiranadi, MSc., Dr. Ir. H. Ridwan Affandi, DEA, Prof. Dr. Ir. Wasmen Manalu, dan Jbu Dr. Ir. Hj. Dewi Apri Astuti, MS. selaku Komisi Pembimbing atas segala petunjuk, saran
dan
bimbingannya.5 Bapak Ir. H. Haruna HamaJ seJaku KepaJa BaJai Budidaya Air Payau Takalar beserta staf yang telah memberikan izin
dan
fasilitas selama penelitian.6 Saudara Umarrifai S.Pi., AkmaJ S.Pi., Andi Ahmad Tahir S.Pi., Ir. Usman, M Si., Mansyur S.Pi, Ghurdi, dan H. Hasanuddin yang teJah membantu selama penelitian.
mengikuti pendidikan di IPH.
8 Khusus kepada kedua orang tuaku Drs. KH. Abd. Karim (almarhum) dan Hj. Sitti Badriah, kedua mettuaku Sutarm. Sury.permana dan Mien Rukminawati. isteriku
Ir.
Nita Rukminasari MP, kedua anakku Ratu SyamsiahNil. Kencan. dan Muluumn.d Mubarak Dimas Aditya Kusuma terciota, serta selurub keluarga .Ias segal. pengorbanan, dukungan, bantuan, pengertian dan doa yang selalu menyertai penulis selama mengikuti pendidikan di Institut
Pertanian Bogor.
Semoga segala harrtuan yang diberikan mendapat pahala yang berlipat
ganda dari Allah SWT.
Akhir kata semoga karya ilmiah ini bennanfaat.
Penulis dilahirkan di Pangkep, Sulawesi Selatan pada tanggal 8 Januari 1955 dari pasangan Drs. KH. Abd. Karim (almarhum) dan Hj. Sitti Badriah. Pendidikan satjana ditempuh di Jurusan Perikanan., Fakultas Peternakan dan
Perikanan UNHAS, lulus pada tabun 1989. Pada tabun 1996, penulis diterima di Program studi !lrnu perairan pada Program PascasaIjana IPB dan menammatkannya pada tabun 1998. Kesempatan untuk melanjutkan ke program doktor di Program Studi Biologi pada perguruan tinggi yang sarna diperoleh pada tabun 2001. Beasiswa BPPS diperoleh dari Dirjen Dikti, Depattemen Pendidikan Nasional Republik Indonesia.
Penulis bekerja sebagai staf pengajar pada Yayasan Pendidikan Arfanita Pangkep tabun 1989 sampai 1990. Sejak tabun 1991 sampai sekarang bekeIj. sebagai staf pengajar pada Fakultas !lrnu Kelautan dan Perikanan UNHAS Makassar. Menikah dengan Ir. Nita Rukminasari, MP. pada tabun 1994 dan dikarunia dua orang anak bemama Ratu Syamsiah Nila Kencana dan Muhammad
Halaman DAFTAR TABEL ... XII
DAFTARGAMBAR ... xiii
DAFT AR LAMPIRAN ... xv
PENDAHULUAN Latar Belakang ... ... .... ... . ... ... ... ... .... ... ... ... .... ... I Pendekatan Pemecaban Masalab .... ... .... ... .... .... ... .... ... .... ... .... ... ... 2
Tujuan dan Kegunaan ... ... 4
Hipotesis ... ... 4
TINJAUAN PUSTAKA Sistematika dan Ciri Morfologi Kepiting Bakau ... 5
Sa1initas dan Osmoregulasi .. ... ... ... .... ... .... ... ... ... .... ... ... .... 6
Kebutuhan Pakan Kepiting Bakau ... .... .... ... ... .... .... ... .... .... ... ... 9
Kebutuban Protein dan Energi Kepiting Bakau .... .... .... ... ... ... .... ... to Pertombuban dan Molting Kepiling Bakau ... 12
Pengaruh Salinitas pada Efisiensi Pemanfaatan Pakan dan Pertombuban . 14 Metabolisme .... ... .... .... ... ... ... .... ... .... ... .... .... ... ... .... ... .... .... 15
Fisika Kimia Air ... 17
Subu ... 17
Keasaman (pH) ... .... ... ... ... ... ... .... ... .... ... ... ... ... .... .... 18
Oksigen Terlarut ... ... 19
Amonia ... 20
Nitrit ... 21
BAHAN DAN METODE PENELlTIAN Waktu dan Tempat ... 23
Bahan Penelitian . ... .... .... ... ... ... ... .... .... ... .... .... ... ... .... ... 23
Hewan Uji ... ... ... ... ... ... ... ... .... ... ... ... .... .... ... ... 23
Sumber Air dan Media Pemeliharaan .... ... ... ... 24
Metode Penelitian ... 26
Pengaruh Salinilas pada OsmoreguJasi Kepiting Bakan ... 26
Tingkat KeIja Osmotik ... 27
Glukosa, Protein, Asam Amino. dan Amonia Hemolimfe ... 28
Konsentrasi Ion-ion Media dan Hemolimfe ... ... ... ... .... .... 28
Konsumsi Oksigen Insang ... 28
Pengaruh Salinitas pada Metabolisme Kepitiog Bakau ... 28
Pengaruh Salinitas pada Tingkat Kelangsungan Hidup dan Pertombuban Kepiting Bakau ... 30
Tingkat Kelangsungan Hidup ... 33
Konsumsi Pakan ... 33
Laju Pertumbuban Bobot Spesifik Harian ... 33
Pertombuban Lebar Karapas ... 33
Produksi Biomassa ... 34
Retensi Nutrien ... 34
Efisiensi Pemanfaatan Pakan ... 35
Neraca
Energi...
35Fisika Kimia Air ... 36
Pengaruh Kadar Protein Pakan pada Tingkat Kelangsungan Hidup dan Pertumbuhan Kepiting Bak.u pada S.linitas Optimum ... 36
Analisis Data.. ... .... ... .... ... ... ... ... ... ... ... ... ... 37
HASIL DAN PEMBAHASAN Pengaruh Salinitas pada Osmoregulasi Kepiting Bakau .. ... 38
Osmolaritas dan Tingkat KeIja Osmotik ... 38
Glukosa. Protein, Asam Amino,
dan
Amoma Hemolimfe ... 42Konsumsi Oksigen Insang ... 46
Pengaruh Salinitas pada Metabolisme Kepiting Bakau . .... ... ... ... 48
Pengaruh Salinitas pada Tingkat Kelangsungan Hidup dan Pertumbuhan Kepiting Bakau ... ... ... ... ... ... ... .... ... ... ... 56
Tingkat Kelangsungan Hidup ... ... ... .... ... .... .... ... ... .... 56
Pertumbuhan ... 58
Produksi Biomassa ... 65
Fenomena Molting ... ... 67
Neraca Energi ... " .... ... ... ... ... ... ... ... 70
Pengaruh Kadar Protein Pakan pada Tingkat Kelangsungan Hidup dan Pertumbuhan Kepiting Bakau pada Salinitas Optimum ... 76
Tingkat Kelangsungan Hidup ... 76
Pertumbuhan ... ... ... ... 76
Produksi Biomassa ... , ... 82
Fenomena Molting ... 82
Fisika Kimia Air ... ... 83
Pembabasan Umum ... 84
SIMPULAN DAN SARAN Simpulan ... ... ... ... ... ... ... ... ... ... 86
Saran ... 86
DAFTARPUSTAKA ... 87
LAMPlRAN ... ... ... .... ... ... 97
Halaman
1 Parameter fisika kimia air yang diukur dan metode/alat yang digunakan ... 36 2 Osmolaritas hcmolimfe (OH), osmolaritas media (OM), dan tingkat
kerja osmotik (TKO) kepiting bakau pada berbagai salinitas media ... 38 3 Konsentrasi ion-ion hemolimfe kepiting bakau pada berbagai salinitas
media ... 39 4 Glukosa (GH), protein (PH), total asam amino (AAH), dan arnonia
hemolimfe (AH) kepiting bakau pada berbagai salinitas media ... 42 5 Konsumsi oksigen insang kepiting bakau pada berbagai salinitas media. .... 46 6 Konsumsi oksigen basal, kenyang, dan rutin (mg O,/g/jarn) kepiting
bakau pada berbagai salinitas dan bobot tubuh... ... ... ... ... ... .... ... 48 7 Laju metabolisme basal, kenyang, rutiI1, dan SDA (kJ/g/hari) kepiting
bakau pada berbagai salinitas dan bobot tubuh ... ... ... .... ... .... ... .... ... 53 8 Tingkat kelangsungan hidup (TKH), konsumsi paksn (KP), laju
pertumbuhan bobot spesifik harian (LPBH), pertumbuhan lebar karapas (PLK), produksi biomassa (PB), dan efisiensi pernanfaatan paksn (EPP) kepiting bakau yang dipelihara pada berbagai salinitas media ... ... 56 9 Persenlase retensi protein (RPr), lemak (RL), kalsium (RCa), dan fosfor
(RP) kepiting bakau yang dipelihara pada berbagai salinitas media .... .... .... 63 \0 Komposisi tubuh kepiting bakau yang dipelihara pada rerbagai salinitas
media ... 63 11 Frekuensi molting (F), periode waktu antarmolting pertama (Mo-I), kedua
(MI-'), konsentrasi kalsium, dan fosfor eksuvia kepiting bakau pada berbagai salinitas media ... 67 12 Konsumsi energi (KE), energi metabolis (EM), Laju metabolisme (LM),
SDA, dan retensi energi (RE) kepiting bakau pada berbagai salinitas
media ... 70 13 Tingkat kelangsungan bidup (TKH), konsumsi pakan (KP), laju
pertumbuhan bobot spesifik harian (LPBH), pertumbuhari lebar karapas (PLK), produksi biomassa (PB), dan efisiensi pemanfaatan paksn (EPP) kepiting bakau yang diberi pakan dengan ksdar protein berbeda .... ... ... 76 14 Persentase retensi protein (RPr), lemak (RL), energi (RE), kalsium (RCa),
dan fosfor (RP) kepiting bakau yang diberi paksn dengan kadar protein berbeda ... _... 79 15 Frekuensi molting (F), periode waktu antarmolting pertama (Mo-I), kedua
(MI-'), konsentrasi kalsium, dan fosfor eksuvia kepiting bakau yang diberi pakan dengan kadar protein berbeda.. ... ... ... ... ... ... ... ... ... ... 83
HalsmaD
I Skema pendekatan pemecahan masalah dan reneana evaluasi pengaruh salinitas pada kinerja pertumbuhan kepiting bakau betina dan evaluasinya
pada salinitas optimum dengan kadar protein pakan berbeda .... ... 3
2 Kepiting bakau (s. .errata Forsskal) ... 5
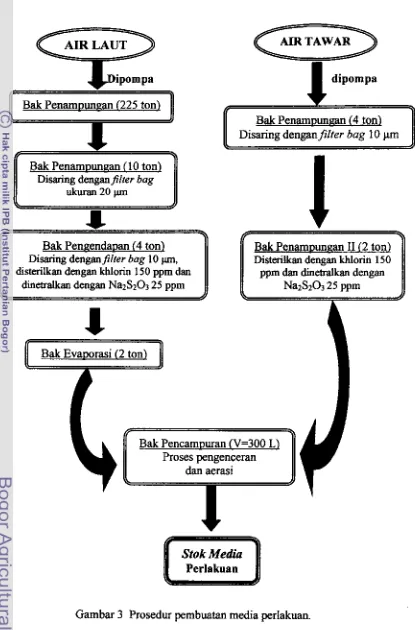
3 Prosedur pembuatan media perlakuan ... ... ... ... ... ... .... .... .... .... .... 25
4 Tala letak satuan percobaan setelah pengacakan ... ... ... .... .... .... .... ... 27
5 Wadab percobaan pengukuran metabolisme ... 29
6 Wadah percobaan pemelibataan kepiting bakau ... 31
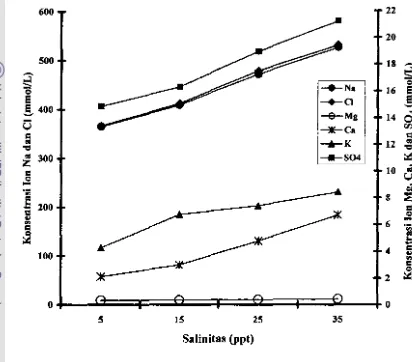
7 HubWlgan salinitas media dengan konsentrasi ion Na +,
cr,
Mi+, Ca2+, K+, dansol-
hemolimfe kepiting bakau ... ... .... ... ... .... .... ... ... ... .... .... 408 Hubungan salinitas media dengan tingkat kerja osmotik kepiting bakau .... 42
9 Hubungan salinitas media dengan konsentrasi glukosa (a) dan protein (b) hemolimfe kepiting bakau ... ... ... .... ... ... ... .... ... .... .... ... .... .... ... 44
10 Hubungan salinitas media dengan konsentrasi amonia hemolimfe kepiting bakau ... 46
II Hubungan salinitas media dengan konsurnsi oksigen insang kepiting bakau ... 48
12 Hubungan salinitas media dengan konswnsi oksigen pada kondisi basal (a), kenyang (b), dan rutin (e) kepiting bakau bobot 20,40,60, dan80g ... 51
13 Tingkat konsurnsi oksigen kenyang kepiting bakau pada bobot 20 (a), 40 (b),
e
(60), dan 80 g (d) pada berbagai tingkat salinitas selama 24 jam sesudah makan ... ... .... ... ... ... ... .... .... ... ... .... ... .... .... ... ... ... 5214 Hubungan salinitas media dengan SDA kepiting bakau pada bobot 20, 40, 60, dan 80
g...
5615 Hubungan salinitas media dengan tingkat kelangsungan hidup kepiting bakau ... 58
16 Hubungan salinitas media dengan konsumsi pakan (a) dan efisiensi pemanfaatan pakan (b) kepiting bakau .. ... ... ... .... ... .... .... ... ... ... .... ... 61
17 Hubungan salinitas media dengan laju pertumbuhan bobot spesifik batian (a) dan pertumbuhan lebar karapas (b) kepiting bakau ... 62
18 Hubungan salinitas media dengan retensi protein (a) dan kalsiurn (b) kepiting bakau ... ... ... ... ... ... .... ... ... ... ... ... ... .... ... 65
19 Hubungan salinitas media dengan produksi biomassa kepiting bakau ... 66
dan periode waktu antarmolting kedua (b) kepiting bakau... .... ... ... ... 69 21 Hubungan salinitas media dengan konsumsi energi (a). energi
metabolis (b), laju metabolisme basal (e), dan laju metabolisme
kenyang (d) kepiting bakau ... 74 22 Hubungan salinitas media dengan SDA (a),laju metabolisme rutin (b),
dan retensi energi (e) kepiting bakau ... 75 23 Hubungan kadar protein pakan dengan laju pertumbuban bobot spesifik
harian kepiting bakau... 78 24 Hubungan kadar protein pakan dengan persentase retensi protein (a),
kalsium (b), dan fosfor (e) kepiting bakau ... 81
Halaman
I Komposisi bahan pakan percobaan.. ... .... ... ... ... .... ... ... ... ... ... ... .... ... 98
2 Komposisi vitamin mix pakan percobaan... 98
3 Komposisi mineral mix pakan percobaan ... ... .... .... .... ... ... ... ... 99
4 Hasil anaiisis proksimat bahan pakan percobaan .... ... ... ... ... ... ... ... 99
5 Prosedur pembuatan pakan percobaan ... ... ... .... .... .... ... ... ... ... ... ... 100
6 Hasil analisis proksimat pakan percobaan ... ... ... .... .... ... .... ... ... ... ... 10 I 7 Prosedur pengukuran osmolaritas hemolimfe kepiting bakan .... .... ... ... 102
8 Prosedur pengukuran osmolaritas media percobaan .... .... ... ... ... 103
9 Prosedur pengukuran glukosa hemolimfe kepiting bakau ... 104
10 Prosedur pengukuran amonia hemolimfe kepiting bakau... ... ... ... 104
II Prosedur pengukuran ion-ion.... ... ... ... ... ... ... .... ... ... ... .... ... ... ... 105
12 Prosedur anaiisis proksimat kadar protein ... 106
13 Prosedur anaiisis proksimat kadar lemak ... 107
14 Prosedur analisis proksimat kadar energi ... " ... ". 108 15 Prosedur analisis proksimat kadar kalsium ... 109
16 Prosedur analisis proksimat kadar fosfor .... ... .... .... .... .... .... .... ... ... ... 110
17 Prosedur pengukuran konsumsi oksigen kepiting bakau .. .... ... ... ... III 18 KhIorinitas dan osmolaritas media pada berbagai salinitas ... ... ... ... 112
19 Konsentrasi ion-ion media pada berbagai salinitas ... ', ... '... .... 113
20 Analisis ragam pengaruh salinitas pada tingkat keIja osmotik kepiting bakau ... 123
21 Analisis ragam pengaruh salinitas pada konsentrasi Na+ hemolimfe kepiting bakau ... 113
22 Analisis ragam pengaruh salinitas pada konsentrasi
cr
hemolimfe kepiting bakau ... 11323 Analisis ragam pengaruh salinitas pada konsentrasi Mi+ hemolimfe kepiting bakau.. ... .... ... ... ... ... .... .... ... .... .... .... ... ... .... ... ... 114
24 Analisis ragam pengaruh salinitas pada konsentrasi
ea
2+ hemolimfe kepiting bakau. ... ... ... ... ... ... .... ... .... .... .... .... ... ... .... ... ... ... 11425 Analisis ragam pengaruh salinitas pada konsentrasi K+ hemolimfe kepiting bakau ... 114
kepiting bakau ... 114 27 Analisis mgam pengarub salinitas pada glukosa hemolimfe kepiting
bakau ... 115 28 Analisis·mgam pengarub sai;nitas pada protein hemolimfe kepiting
bakau ... 115 29 Analisis mgam pengaruh salinitas pada konsentmsi total asam amino
hemolimfe kepiting bakau ... 115 30 Analisis mgam pengarub salinitas pada amonia hemolimfe kepitiog
bakau ... 115 31 Analisis mgam pengarub salinitas pada konsumsi oksigen insang (tanpa
digitalis blok ATPase) kepiting bakau ... 116 32 Analisis mgam pengarub salinitas pada konsumsi oksigen insang
(dengan digitalis blok ATPase) kepiting bakau ... 116 33 Analisis mgam pengarub salinitas pada kemapuan pompa ion kepiting
bakau ... 116 34 Analisis ragam pengarub salinitas pada konsumsi oksigen basal kepiting
bakau bobot 20 g ... 116 35 Analisis mgam pengarub salinitas pada konsumsi oksigen basal kepiting
bakau bobot 40 g ... 117 36 Analisis ragam pengaruh salinitas pada konsumsi oksigen basal kepiting
bakau bobot 60 g ... 117 37 Analisis mgam pengaruh salinitas pada konsumsi oksigen basal kepiting
bakau bobot 80 g ... 117 38 Analisis ragam pengaruh salinitas pada konsumsi oksigen kenyang
kepiting bakau bobot 20 g ... 117 39 Analisis ragam pengaruh salinitas pada konsumsi oksigen kenyang
kepiting bakau bobot 40 g ... 118 40 Analisis ragam pengaruh salinitas pada konsumsi oksigen kenyang
kepiting bakau bobot 60 g ... 118 41 Analisis ragam pengarub salinitas pada konsumsi oksigen kenyang
kepiting bakau bobo! 80 g ... 118 42 Analisis mgam pengarub salinitas pada konsumsi oksigen rutin kepiting
bakau bobot 20 g ... 118 43 Analisis ragam pengarub salinitas pada konsumsi oksigen rutin kepiting
bakau bobot 40 g ... 119 44 Analisis ragam pengaruh salinitas pada konsumsi oksigen rutin kepiting
bakau bobot 60 g ... 119
bakau bobot 80 g ... 119 46 Analisis ragam pengaruh saIinitas pada l.ju metabolisme basal kepiting
bakau bobot 20 g ... 119 47 Analisis ragam pengaruh salinitas pada l.ju metabolisme basal kepiting
bak.u bobot 40 g ... 120 48 Analisis ragam pengaruh saIinitas pada l.ju metabolisme basal kepiting
baksu bobot 60 g ... 120 49 Analisis ragam pengaruh saIinitas pada l.ju metabolisme basal kepiting
bak.u bobot 80 g ... 120 50 Analisis ragam pengaruh saIinitas pada l.ju metabolisme kenyang
kepiting bak.u bobot 20 g ... 120 51 Analisis ragam pengaruh s.linitas pada laju metabolisme kenyang
kepiting bakau bobot 40 g ... 121 52 Analisis ragam pengaruh saIinitas pad. l.ju metabolisme kenyang
kepiting baksu bobot 60 g ... 121 53 Analisis ragam pengaruh saIinitas pada l.ju metabolisme kenyang
kepiting bakau bobot 80 g ... 121 54 Analisis ragam pengaruh saIinitas pada l.ju metabolisme rutin kepiting
bakau bobot 20 g ... 121 55 Analisis r.gam pengaruh salinitas pada l.ju metabolisme rutin kepiting
baksu bobot 40 g ... 122 56 Analisis ragam pengaruh salinitas pada l.ju metabolisme rutin kepiting
baksu bobot 60 g ... 122 57 Analisis ragam pengaruh salinitas pada laju metabolisme rutin kepiting
baksu bobot 80 g ... 122 58 Analisis ragam pengaruh salinitas pada SDA kepiting baksu bobot 20 g ... 122 59 Analisis ragam pengaruh saIinitas pada SDA kepiting baksu bobot 40 g ... 123 60 An.lisis ragam pengaruh salinitas pada SDA kepiting bakau bobot 60 g ... 123 61 Analisis ragam pengaruh salinitas pada SDA kepiting baksu bobot 80 g ... 123 62 Analisis ragam pengaruh saIinitas pada tingkat kelangsungan bidup
kepiting baksu ... 123 63 Analisis ragam pengaruh saIinitas pada I.ju pertumbuban bobot spesifik
harian kepiting baksu ... 124 64 Analisis ragam pengaruh saIinitas pada pertumbuban lebar karapas
kepiting bak.u ... 124. 65 analisis ragam pengaruh s.linitas pada konsumsi pakan kepiting baksu .... 124
kepiting bakau ... 124
67 Analisis mgam pengaruh salinitas pada kandungan protein daging kepiting bakau ... 125
68 Analisis mgam pengaruh salini"", pada kandungan lemak daging kepiting bakau ... 125
69 Analisis mgam pengaruh salinitas pada kandungan energi daging kepiting pertama kepiting bakau ... 125
70 Analisis ragam pengaruh salinitas pada retensi protein kepiting bakau ... 125
71 Analisis ragam pengaruh salinitas pada retensi lemak kepiting bakau ... 126
72 Analisis ragam pengaruh salinitas pada retensi kalsium kepiting bakau ... 126
73 Analisis ragam pengaruh salinitas pada retensi fosfor kepiting bakau ... 126
74 Analisis ragam pengaruh salinitas pada produksi biomassa kepiting bakau ... 126
75 Analisis ragam pengaruh salinitas pada frekuensi molting kepiting bakau .. 127
76 Analisis ragam pengaruh salinitas pada periode waktu antarmolting pertama kepiting bakau ... 127
77 Analisis ragam pengarub salinitas pada periode waktu antarmolting kedua kepiting bakau ... 127
78 Analisis ragam pengaruh salinitas pada kadar kalsium eksuvia kepiting bakau setiap molting ... 127
79 Analisis ragam pengarub salinitas pada kadar fosfor eksuvia kepiting bakau setiap molting ... 128
80 Analisis ragam pengaruh salinitas pada konsumsi energi kepiting bakau ... 128
81 Analisis ragam pengarub salinitas pada energi metabolis kepiting bakau ... 128
82 Analisis ragam pengaruh salinitas pada I.ju metabolisme basal kepiting bakau ... 128
83 Analisis mgam pengarub salinitas pada laju metabolisme kenyang kepiting bakau ... 129
84 Analisis mgam pengaruh salinitas pada SDA kepiting bakau ... 129
85 Analisis mgam pengaruh salinitas pada laju metabolisme rutin kepiting bakau ... 129
86 Analisis ragam pengaruh salinitas pada retensi energi kepiting bakau ... 129
87 Analisis ragam pengaruh salinitas pada rasio energi metabolis--konsurnsi energi kepiting bakau ... 130
88 Analisis ragam pengaruh salinitas pada rasio retensi energi-konsumsi energi kepiting bakau ... 130
kepiting bakau ... 130 90 Analisis ragam pengaruh salinitas pada rasio laju metabolisme
rutin-konsumsi energi kepiting bakau. ... 130 91 Analisis ragam pengarub kadar protein pakan tingkat ke1angsungan
bidup kepiting hakau ... 131 92 Analisis ragam pengarub kadar protein pakan pada laju pertumbuban
bobot spesifIk barian kepiting bakau ... 131 93 Analisis ragam pengarub kadar protein pakan pada pertumbuban lebar
kanspas kepiting bakau ... 131 94 Analisis ragam pengarub kadar protein pakan pada konsumsi pakan
kepitingbakau ... 131 95 Analisis ragam pengarub kadar protein pakan pada efisiensi pernanfaatan
pakan kepiting bakau ... 132 96 Analisis ragam pengarub kadar protein pakan pada retensi protein
kepiting bakau ... 132 97 Analisis ragam pengarub kadar protein pakan pada retensi lemak
kepitingbakau ... 132 98 Analisis ragam pengarub kadar protein pakan pada retensi energi
kepiting bakau ... 132 99 Analisis ragam pengarub kadar protein pakan pada retensi kalsium
kepiting bakau ... 133 100 Analisis ragam pengarub kadar protein pakan pada retensi fosfor
kepiting bakau ... 133 101 Analisis ragam pengarub kadar protein pakan pada produksi biomassa
kepiting bakau ... 133 102 Analisis ragam pengarub kadar protein pakan pada frekuensi molting
k .. epltmg bak au ... 133 103 Analisis ragam pengarub kadar protein pakan pada periode waktu
anlannolting pertama kepiting bakau ... 134 104 Analisis ragam pengarub kadar protein pakan pada periode waktu
antar molting kedua kepiting bakau ... 134 105 Analisis ragam pengarub kadar protein pakan pada konsentrasi kalsium
eksuvia kepiting bakau setiap molting ... 134 106 Analisis ragam pengarub kadar protein pakan pada konsentrasi fosfor
eksuvia kepiting bakau setiap molting ... 134
Latar Belakang
Kepiting bakau (&y/la serrato Forsskal) merupakan salah s.tu organisme estuari. bernilai ekonomis tinggi (Millamen. dan Quinito 2000; Fr.tini dan Vannini 2002; Trino dan Rodriguez 2002). Jenis kepiting ini telah dikenal b.ik eli pasaran dalam negeri maupilll luar negeri karena rasa dagingnya yang lezat dan bemil'; gizi tinggi yakni mengandung berl>agai nutrien penting seperti mineral dan asam lemak 0)-3 (Catacutan 2002).
Selama ini permintaan konsumen akan kepiting bakau sebagian besar
dipenuhi dan basil penangkapan eli alam yang sifatoy. fluktuatif. Di sisi lain,
meningkatnya pennintaan konsumen terutama di pasar internasional menuntut produksi kepiting bakau yang berkesinambungan yang hany. dapat dipenuhi
melalui upaya buelidaya secara intensif. Keberhasilan budidaya kepiting bakau
elitunjukkan oleh pertumbuhan yang pes.t dalam waktu singkat dan tingkat kelangliungan hidup yang tinggi. Pertumbuhan hany. dapat terjadi apabil.
terdapat kelebihan energi, setelah energi yang elikonsumsi dikurangi dengan
kebutuhan energi untuk hidup pokok. Perubahan konelisi lingkungan, terutam.
salinitas, akan berpengaruh pada besaran energi yang dikonsumsi, dan besamya
energi yang digunakan untuk osmoregulasi. Dengan dernikian, perubahan
lingkungan akan mempengaruhi pertumbuhan.
S.linitas merupakan s.lah satu faktor lingkungan yang berpeng.ruh penting
pada kelangliungan hidup, konsumsi pakan, metabolisme, dan pertumbuhan organisme akuatik (Kumlu ef of. 2001; Rowe 2002; Villareal ef al. 2003; Huynh
dan Fotedar 2004; Z""hati. dan Kak.ti 2004) karena merupakan masking factor
yang dapat rnemodifikasi peubah fisika dan kimia air menjadi satu kesatuan
pengaruh yang berdampak osmotik pada osmoregulasi dan bioenergetik organisme akuatik (Gilles dan Pequeux 1983; Ferraris et of. 1986). Dalam hal ini, salinitas akan berpengaruh pada pengaturan ion-ion internal, yang seeara langsung
memerlukan energi untuk transport
aktif
ion-ion guna mempertahankan·kondisi salinitas medium mampu mendukung proses·proses fisiologis seeara
maksimal. Dengan demikian, untuk menghasilkan pertumbuhan kepiting bakau yang pesat dan tingkat kelangsungan hidup yang tinggi diperlukan salinitas media pemeliharaan yang mampu mengoptimalkan konsumsi pakan
dan
meminimalkan pembelanjaan energi untuk osmoregulasi.Pendekatan Pemecaban Masalah
Pertumbuhan kepiting bakau berkaitan dengan efisiensi pemanfaatan pakan khususnya energi pakan. Salah satu faktor penentu efisiensi tersebut adalah
kualitas media, yaitu salinitas. Meskipun kepiting bakau termasuk organisme akuatik yaog bersifat eurihaline, yaitu mampu beradaptasi pada media dengan rentang salinitas lebar yakni I sarnpai 42 ppt (Chen dan Chia 1997), kisaran salinitas yang optimun untuk tumbub lebih sempit jika dibandingkan dengan kisaran untuk mempertahankan kehidupannya.
Pads dasamya pertumbuhan kepiting bakau bergantung pads energi yang tersedia, bagaimana energi tersebut digunakan di dalarn tubub, dan pertumbuhan hanya akan terjadi apabila terdapat kelebihan energi setelab kebutuban energi minimalnya (untuk hidup pokok) terpenubi. Kepiting bakau akan kehilangan energi sebagai akihat dari penggunaan energi uotuk: osmoregulasi. Tingkat
pemanfaatan pakan (energi) untuk pertumbuhan sangat bergantung pads daya dukung lingkungannya. Dalarn rentang kualitas air yang layak dan kebutuhan pakan bagi kehidupan kepiting bakau terpenubi, salinitas media akan menentukan pertumbuhan yang selanjutnya akan menentukan tingkat efisiensi pemanfaatan
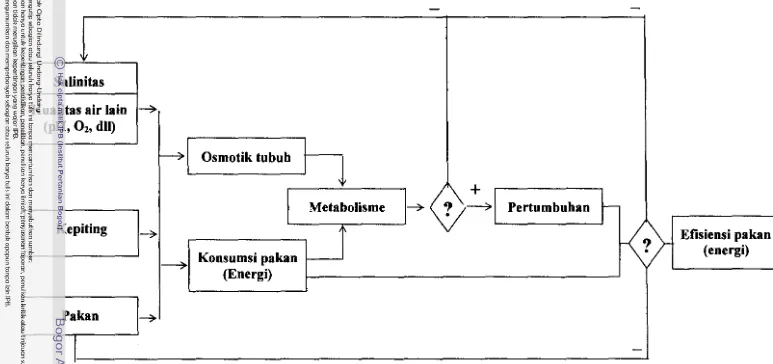
pakan (Garnbar 1).
Dati permasalahan di atas dapat dinyatakan bahwa untuk menghasilkan pertumbuhan kepiting bakau yang maksimal diperlukan media pemeliharaan dengan salinitas yang optimun sehingga mampu meningkatkan efisiensi
pemanfaatan pakan (energi). Oleh karena tingkat salinitas optimun untuk pertumbuban kepiting bakau belum dapat ditentukan, perlu dilakukan penelitian dengan Iingkup kajian sebagai berikut :
1 Osmoregulasi. yaitu kemampuan kepiting untuk: melakukan adaptasi sehingga
Kualitas air lain
セ@[
(pH, 0"
dll)
セi@
Osmotik tubuh
I
1
+
Metabolisme
---?
?
- - 7Pertumbuhan
I-•
Kepiting
I---?
W
Efisiensi pakan
Konsumsi pakan
- - 7(Energi)
Pakan
---?
I
-Gambar 1 Skema pendekatan pemecahan masal.h dan rencana ev.luasi pengaruh salinitas p.d. kioerj. pertumbuban kepiting bakau dan evaluasinya pada salinitas optimum dengan kadar protein pakan berbeda.
[image:22.830.5.778.84.448.2]2 Metabolisme, yaitu kemampuan kepiting untuk. memetabolisme energi pada
salinitas tertentu.
3 Pertumbuhan, yaitu kemampuan kepiting untuk tumbub pada salinitas tertentu. 4 Efisiensi pemanfaatan pakan, yaitu kemampuan kepiting mengkonsumsi pakan secara maksimun pada salinitas tertentu
dan
memanfaatkannya untuk tumbuh .. Tujuan dan Kegunaan
Berdasarkan rumusan masalah yang diuraikan di atas. penelitian ini bertujuan:
I Mengetabui pola osmoregulasi dan metabolisme kepiting bakau yang dipelihara pada berbagai salinitas.
2 Mengevaluasi kineIja pertumbuban kepiting bakau yang dipelibara pada berbagai salinitas dan menentukan salinitas optimum yang mengbasilkan pertumbuban yang maksimal.
3 Mengevaluasi kineIja pertumbuban kepiting bakau yang diberi pakan dengan kadar protein berbeda pada salinitas optimun.
Hasil penelitian ini diharapkan dapat memberi sumbangan informasi
pengetabuan mengenai peran salinitas pada tingkat kelangsungan hidup dan pertumbuban kepiting bakau
Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah sebagai berikut :
1 Perbedaan salinitas media akan mempengaruhi perbedaan kineIja
pertmnbuhan kepiting bakau.
Sistematika daD Ciri Morfologis Kcpiting Bakau
Kepiting bakau (S<yl/a serrata Forsskal) tergolong ke dalam filum Arhtropoda, subfilum Mandibulata, kelas Crustacea, subkelas Malacostraca, sen
Eumalacostraca, superordo Eucarida. erdo d・」。ーッ、。 セ@ subordo Raplantia. seksi
Brachiura, subseksi Branchyrhyncha, famili Portunidae, marga Scylla (Motoh 1977; Keenan el al. 1998).
Ciri-ciri morfologis kepiting bakau adaJah karapasnya berukuran lebib lebar
dari pada panjaog, paojang karapas kurang lebih dua pertig. dari lebamya. Sisi antero-lateral berduri sembiJan buah dengan ukuran yang harnpir sarna besar. Oi
sntaea sepasang matanya terdapat eDam buab duri. sedangkan di bagian kanan dan kirinya masing-rnasing mempunyai sembilan buah duri . Pada dahi terdapat empat
buah gigi tumpul, tidak lermasuk ruaog mata sebelah dalarn yang berukuran kurang lebih sarna. Mcmpunyai sepasang sapit, riga pasang kaki jalan dan sepas80g kalci rcoang. Pasangan sapit mempunyai bagian propodus yang menggembung dengan permukaan licin
dan w..uan
yang cukup besar dibanding kakj-kaki yang lain dan berfungsi untuk memegang. Pasangan kaki terakhir (periopod V) berebentuk pipih pada ruas terakhir dan berfungsi sebagai alat renang (Motoh 1977; Kuntiyo el al. 1994; Keenan el al. I 998)(G.mbar 2). [image:24.608.85.546.54.780.2]Ukuran ruas-ruas abdomen dapa! digunakan untuk membedakan antara kepiting janlan dan belina. Ruas abdomen kepiling janlan sempit, sementara pada betina lebih besar. Perut kepiting jantan berbentuk segitiga meruncing. sementara belina berbentuk segitiga melebar. Perbedaan lain adalab pleopod (kaki renang) yang terletak di bawah abdomen. Pada kepiting janlan pleopod tersebut berfungsi sebagai alat kopulasi, sedangkan pada betina berfungsi sebagai tempat pelekatan/ penempelan telur (Motoh 1977).
Salinitas dan Osmoregulasi
Salinitas merupakan konsentrasi total dari semua ion yang larot dalam air,
dan dinyatakan dalam bagian perseribu (ppt) yang setara dengan gram per liter (Boyd 1990). Sifat osmolik air berasa1 dari seluruh elektrolit yang lamt dalam air tersebut. Semakin tinggi salinitas, konsentrasi elektrolit makin besar, sehingga
tekanan osmotiknya makin linggi (McConnaughey dan Zottoli 1983). Air laut mengandung 6 elemen terbesar, yaitu
cr,
Na+, Mg'+, Ca'+, K, dan SO," (Iebih dari 90% dari garam total yang terlamt) ditambab elemen yang jurnlahnya kecil (unsur mikro) seperti Br, Sr'+, dan B+. Ion-ion yang dominan dalam menentukan tekanan osmolik (osmolaritas) air laut adalab Na+ (450 mM) dancr
(560 mM),dengan porsi 30.61 dan 55.04 persen dari total konsentrasi ion-ion terlarut
(McConnaughey dan Zottoli 1983; Nybakken 1990; Boeuf dan Payan 2001; Mananes e( 01. 2002).
Salinitas dapat mempengaruhi aktivitas fisioiogis organisrne akuatik karena
pengaruh osmotiknya (Gilles dan Pequeux 1983; Ferraris e( 01. 1986). Ditinjau
dari aspek ekofisiologi, organisme akualik dapat dibagi menjadi dua kategori sehubungan dengan mekanisme faalinya dalam mcnghadapi osmolaritas media
(salinitas), yaitu osmokonfonner dan osmoregulator. Osmokonformer adalah
orgarusme yang secara osmotik labil karena tidak mempunyai kemampuan
mengatur kandungan garam serta osmolaritas cairan intemalnya. Oleh sebab itu,
osmolaritas cairan tubuhnya selalu berubah sesuai dengan kondisi osmolaritas
media bidupnya. Osmoregulator adalah organisme yang mempunyai mekanisme
faali untuk menjaga kemantapan linglrungan intemalnya dengan cara mengatur
Fanner 1983; Nybakken 1990). Sesuai dengan rentang salinitas yang masih dapat
ditolerir yaitu I sampai 42 ppt (Chen dan Chia 1997), kepiting bakau termasuk organisme akuatik tipe osmoregulator. Kemampuan osmoregulasinya sangat
bergantnng pada tingkat salimtas medianya
Osmoregulasi merupakan mekanisme adaptasi lingkungan yang penting
bagi organisme akuatik, khusnsnya krustasea, nntuk menjaga kemantapan
lingkungan intemalnya dengan cara mengatur keseimbangan osmotik antara
cairan intrasel dengan cairan ekstraselnya (Mantel dan Fanner 1983; Lignot el al. 1999; Huynh dan Fotedar 2004). Dalam proses ini diperlukan transpor aktif terutama pompa Na·K.ATPase. karena ion-ion Na+ dan K+ merupakan ion-ion
yang dominan. Hal ini dilakukan nntuk mempertahankan gradien osmotik antara
cairan tubnh dan cairan medinm agar proses fisiologis dalam tubnh berjalan normal (Towle el al. 2001; Mananes el al. 2002). Aktivitas Na+fI(+-ATPase lebih tinggi pada insang bagian posterior karena insang bagian posterior berhnhnngan
dengan fungsi osmoregulasi utama, sedangkan insang anterior sebagian besar berfungsi nntuk respirasi (Thurman 2003; Palacios el al. 2004). Osmoregulasi pada organisme akuatik bergantung pada transport aktif ion-ion melalui pompa elektrogenik pada membran selluler (Alvares el al. 2004).
Osmoregulasi dapat teIjadi lewat dua mekanisrne yaitu dengan
mempertahankan kemantapan osmolaritas cairan ekstrasel tanpa barus mendekati isoosmotik pada salinitas media, dan menjaga kemantapan osmolaritas cairan intrasel agar tetap isoosmotik dengan cairan ekstraselnya Kedua mekanisme
tersebut dilakukan dengan cam mengatur volume air di dalam cairan ekstrasel
serta mengatur pertukaran ion antara cairan intrasel dan ekstrasel (Gilles dan Pequeux 1983). Faktor-faktor yang mempengarnhi tekanan osmotik intraselluler
akan mempengaruhi metabolisme asam amino dan selanjutnya mempengaruhi
komposisi protein pada kondisi stres osmotik. Dalam hal lain, osmoregulasi
ekstraselluler, dengan kata lain, berhubungan dengan penggunaan energi nntuk
transpor ion aktif, meliputi degradasi bahan-bahan yang kaya energi seperti
lemak. Mekanisme ini akan menghasilkan respon perubahan biokimia pada.
Komponen yang terlibat dalam osmoregulasi adalab (I) kendali honnona! (organ X/kelenjar sinus dan organ perikardia), dan (2) protein pada membran sel, yang berperan sebagai sistem pompe ion, pengemban (karier) dan biokatalisator (enzim, Na-K ATP ase), serta energi (ATP) untuk transport aktif. Adapun organ-organ tubuh yang ikut berperan sebagai tempat berlangsungnya osmoregulasi adalab insang, saluran pencernaan, integumen (kulit) dan organ ekskresi pada kelenjar antena (Mantel dan Farmer 1983; Wilder ef al. 2000; Mananes ef al.
2002).
Pada kondisi lingkungan yang hipertonik, cairan tubuh kepiting bersifat hipoosmotik terbadap media hidupnya. Oleh sebab itu, air dari cairan tubuh cenderung untuk bergerak ke luar secarn osmosis (Mantel dan Farmer 1983). Dalam kondisi tersebut kepiting akan berusaha mempertahankan osmolaritas
cairan tubuh agar cairan internal
tidak
keluar dari selnya serta mencegah agarcairan urin tidak lebih pekat dan hemolimfenya. Untuk keperluan itu, kepiting mengekstrak H20 dati med.ianya, dengan cara minum air atau memasukkan air
lewat insang dan kulit (pada saat ganti kulit). Di dalarn saluran pencemaan, air dan ion terlarut itu diabsorbsi. Kelebihan ion, terutama Na +
dan
cr
yang diamhiloleh hemolimfe akan dikeluarkan oleh insang melalui sel-sel epitei (salt secreting epithelium), sehingga diperoieh air bebas ion untuk pembentukan urin dan keseimbangan osmotik cairan tubuh kepiting. Pengaturan keseimbangan ion
tersebut dilakukan dengan cara transport aktif yang memerlukan sejurnJah energi
yang berasal dan ATP (Adenosin tri-fosfat) (Mantel dan Farmer 1983; Verslycke dan Janssen 2002).
Pada kondisi lingkungan yang hipotonik, cairan tubuh kepiting bersifat
hiperosmotik terhadap media eksternalnya. Pada kondisi tersebut, air dari media
ekstemal cenderung untuk menembus masuk ke dalam bagian tubuh yang berlapis
tipis, antara lain insang, usus dan kulit (pada saa! ganti kUlit). Ion-ion cenderung berdifusi ke luar tubuh dan cairan internal akan terancarn kekurangan ion melalui
ekskresi. Untuk mengatasi bal itu, kepiting akan berusaba mempertahankan kernantapan osmolaritas cairan tubuh dengan mekanisme regulasi ィゥー・イッウュッエゥセ@
ekskresi (kelenjar antena). Dalam hal ini alat ekskresi berfungsi sebagai "pompa air" sehingga kelebiban volume air di dalarn cairan ekstrasel dapat dikeluarkan melalui urin yang hipoosmotik (GiUes dau pequeux 1983; Mantel dan Farmer 1983). Rainbow dan Black (2001) mengemukakan babwa permeahilitas krustasea dekapoda berhubungan dengao salinitas. Adaptasi secara fisiologis dalam permeabilitas pada salinitas rendab mengurangi pengambilan dau pengeluaran air, kehilangao garam-garam dau pembelanjaan energi dibutuhkan untuk pengambilan ion pacta salinitas rendah.
KebutubaD Pakan Kepiting Bakau
Informasi mengenai pakan kepiting bakau di alam telab dilaporkan oleh Lavina (1980), Kuntiyo e/ 01. (1994) dau Catacutan (2002). Kepiting bakau dewasa termasuk jenis hewan pemakan segala dan bangkai (omnivorous scavenger). Pada saat larva, kepiting bakau memakan plankton, dau pada saat juvenil menyukai detritus sedaugkan kepiting dewasa menyukai ikan dau moluska terutarna kekerangan. Penggunaan pakan buatan pada kepiting bakau telab dilakukan (Sheen dau Wu 1999; Sheen 2000; MiUarnena dan Quinito 2000; Catacutan 2002) meskipun masih sebatas penelitian.
Kepiting bakau membutuhkan pakan untuk mempertahankan eksistensi hidup serta pertumbubannya, dau akan bertumbuh dengao baik jika pakan yang tersedia mengandung seroua unsur-UDsur nutrien yang dibutuhkan.
Gutierrez-Yunita dau Montes (2001) mengemukakan bahwa komposisi nutrisi pakan esensial akan menentukan pertumbuhan dan efisiensi pemanfaatan pakan
organisme.
Seeara umum kebutuhan nutrien kepiting bakau meliputi protein,
karbohidrat, lemak, vitamin, dan mineraL Protein merupakan komponen pakan
terpenting dalam sel makhluk hidup yang berfungsi untuk membentuk jaringan tubuh, mernperbaiki jaringao tubuh yang rusak, merupakan komponen enzim dalam tubuh dau sumber energi untuk keperluan metabolisme. Karbohidrat, selain untuk memenuhi kebutuhan energi dan persediaan makanan di dalam tubuh, juga .
berfungsi untuk sintesis khitin pada kulit, polimerisasi khitin
dan
pembentukanuntuk pertumbuhan, yang berfungsi untuk pemeliharaan struktur dan integritas membran sel dalam bentuk fosfolipid
dan
sebagai sumher energi. Vitaminmerupakan senyawa organik yang dibutubkan meskipun dalam jurnlab yang sedikit, tetapi mempunyai peraoan penting dalam proses fisiologtsnya. Mineral
mempuny.i peranan dalam pembentukan eksoskeleton, mempertahankan tingkat koloidal cairan tubuh dan mengatur beberapa sifat fisik sistern koloidal seperti viskositas, difusi. tekanan osmosis, struktur jaringan, pengiriman impuls syaraf,
kontraksi otot, mengatur keseimbangan asam basa dan sebagai komponen atau
aktifator enzim (NRC 1993; Anwar dan Piliang 1992; Hernandez et al. 2001; Pr.toomchat et al. 2002; S.tpathy et al. 2003).
Manajemen pemberian pakan pada budiday. kepiting b.kau merupakan faktor penentu keberhasilan. Menurut Jlyas et al. (1987), terdapat 5 prinsip yang perlu dipertimhangkan dalam memilih pakan yang tepat, yaitu : (1) kuantitas, (2) kualitas (nilai nutrisi dan sanitasi), (3) hentuk dan ukuran, (4) day. tarik, dan (5) ketabanan (stabilitas) di dalam .ir (media). Tujuan akhir .p1ilcasi pakan adalab untuk mendapalkan tingkat kelangsungan hidup yang tinggi, I.ju perturnbuhan yang pesat dan bi.ya tetjangkau.
Kebutuhan Protein dan Eoergi Kepiting Bakau
Keberadaan protein dalam pak.n sang.t penting, baik dalam kualitas m.upun kuantitasny.. Sel.in digunakan sebagai baban untuk perturnbuhan, protein juga penting untuk poduksi enzim. dan lain-lain (Hernandez et al. 2001;
Kim dan Lall 2001; S.tp.thy et al. 2003). Perturnbuhan digambarkan seb.gai peningkatan protein tubuh dan protein merupakan komponen paling mahal dalam
formuiasi pakan (Kim dan Lall 2001).
Jika kebutuhan protein dari pakan tidak tercukupi, akan teqadi penunman drastis dan penghentian perturnbuhan .tau kehilangan bobot tubuh karen. hewan akan menarik kembali protein dari beberapa jaringan untuk mempertabankan fungsi jaringan yang lebih penting. Seb.likny. jik. pasokan protein berlebih m.k. energi yang digunakan untuk proses deaminasi asam-asam amino akan meningkat
sehingga mengurangi energi untuk perturnbuh.n vital (NRC 1993; Hernandez et
dengan pemasukan protein dan ketersediaan sumber energi nonprotein, yaitu
lemak. Pemasukan energi nonprotein memperlihatkan penghematan protein dari
katabolisme untuk penyediaan energi dan meningkatkan pemanfaatan protein
UDtuk pertumbuhan, suatu proses yang dikenal dengan protein sparing effect
(Satpathy ef af. 2003).
Energi adalah suatu kapasitas untuk melakukan kerja yang diperlukan dalam
semua fase metabolisme tubuh. Kebutuhan energi dipengaruhi oleh beberapa
faktor antara lain spesies, wnur, ukuran, aktivitas, dan jenis pakan. Energi dibutuhkan untuk kontraksi (gerak) dan untuk aktivitas proses metabolisme. Hampir 60% energi pakan yang dikonsumsi organisme digunakan untuk memelihara tubuh dan selebihnya digunakan untuk pertumbuhan. Kebutuhan energi kepiting harus dipenuhi melalui pemberian pakan seperti protein, lema!<,
dan karbohidrat yang merupakan sumber energi. Kandungan energi dan ukuran partikel pakan penting karena akan menentukan laju pertumbuhan dan deposisi energi (Halver 1989; Kim dan Lall 2001). Hasil penelitian Cataeutan (2002) memperlihatkan bahwa perturnbuhan kepiting bakau yang haik dihasilkan pada
kepiting yang diberi pakan dengan kadar energi 14.7 sampai 15.7 MJ/kg.
Pertumbuhan sangat dipengarubi oleh imbangan protein dan energi dalam pakan Pakan yang mengandung protein tinggi belurn tentu dapat mempercepat
pertumbuhan apahila kandungan energinya rendah. Kebutuhan energi untuk
pemeliharaan dan kegiatan tubuh harus dipenuhi terlebih dahulu, sebelurn energi
pakan tersebut digunakan untuk pertumbuhan. Protein dalam pakan akan
digunakan untuk menghasilkan energi jika kandungan energi pakan tak seimbang
dengan kandungan proteinnya (Lovell 1989). Taboada ef af. (1998) dan Rosas
ef af. (200 1) mengemukakan bahwa pakan dengan rasio protein per energi optimum menggambarkan titik keseimbangan antara jumlah energi yang
dibutuhkan untuk: metabolisme basal dan pertumbuhan. Keseimbangan antara
kebutuhan protein dan energi untuk pertumbuhan .dalah salah satu kunei untuk mendapatkan pakan yang sesuai. Satpathy ef af. (2003) mengemukakan bahwa pakan dengan rasio protein per energi optimun akan menghasilkan pertumbuhan
Penambahan nutrien sebagai penghasil energi dapat menurunkan
penggunaan protein sebagai sumber energi (protein sparing effect) sehingga dapat
meningkatkan fungsi protein dalam menunjang pertumbuban. Halver (1989)
mengemukakan bahwa pakan yang kekurangan energi akan rnenyebabkan' sebagian besar protein pakan digunakan sebagai sumber energi untuk keperluan
metabolisme. Sebaliknya jika kandungan energi pakan terlalu tinggi dapat menyebabkan pakan yang dimakan berkurang dan penerimaan nutrien lainnya
termasuk protein yang diperlukan untuk pertumbubanjuga berkurang.
Pet1umbuhan dan Molting Kepiting Bakau
Pertumbuban pada kepiting bakau merupakan pertambahan bobot badan dan lebar karapas yang terjarli secara berkala setelab terjadi pergantian kulit atau
molting (Sheen dan Wu 1999; Mayrand ef al. 2000; Catacutan 2002). Molting
bagi krustasea merupakan periode kritis yang menggambarkan kondisi fisiologis
dari proses pergantian kulit lama (eksoskeleton) (Gimenez ef al. 2001; Dooley
ef al. 2002). Molting dipengaruhi oleh faktnr ekatemal seperti salinitas,
tempemtur, dan faktor internal tennasuk status nutrisi dan ablasi mata (koo et al.
2005).
Peningkatan ukuran tubuh dan biomassa teIjadi setelah molting (Hoang
ef al. 2003) yakni selama periode antara postmolt dan premolt karena proses
fisiologis intensif dan biokimia terjadi selama periode tersebut (Mayrand et al.
2000; Godbout ef al. 2002), sedangkan pada periode postmolt terjadi peningkatan
pengambilan air (Dooley ef al. 2002). Pratoomchat ef al. (2002) mengemukakan
bahwa perubahan osmolalitas hemolimfe S. serrafa selama siklus molting
mungkin berkorelasi dengan elemen-elemen minor seperti kalsium, kalium,
tembaga, dan mangan atau tingkat protein dan karbohidmt. Kemungkinan
konsentrasi paling tinggi bahan organik ini terjadi selama premolt. Kalsium adalah
ion ulama yang diangknt dan disimpan dalam kutikula sebagai kalsium karbonat,
sedangkan tembaga memainkan peranan penting sebagai karier oksigen dalam
hemolimfe pada S. serrata.
pascaganti kulit (postmolt), periode waktu antarganti kulit (intennolt). Pada persiapan ganti kulit pertama, epidennis menyebar dari lapisan membran
dan
epikutikula barn, dan mobilisasi glikogen pada jaringan epidennal. Pada persiapan ganti kulit kedua, endokutikula barn mulai disekresi, duri barn keluar dari jaringan lipatan dactylus, dan lapisan memhran lama mengalami degenerasi menjadi
lapisan gelatin. Pada persiapan ganti kuht ketiga, periode resorhsi eksoskeleton
tua. Pada persiapan ganti kulit ォ・・ュー。セ@ resorpsi lengkap, eksoskeleton tua retal< sepanjang garis epimeraI dan penyerapan air dimulai. Pacia ekdisis, kepiting
melepas eksoskeleton tua dan penyempan air dipercepat. Pada periode ganti kulit pertarna, eksoskeleton tereliri alas membmn yang sangat lunak, kaki belum dapat menyanggah bobot badan, kepiting tidak akuf, penyerapan air kontinyu, dan mineralisasi eksoskeleton dimulai. Pada periode ganti kulit kedua, eksoskeleton temsa keras, kaki sudah dapat menyanggab bobot badan, kandungan air dalam tubuh konstan sekitar 86% dan mulai teIjadi penumpukan dan mineralisasi pada endokutikula. Pada peri ode pascaganti kulit pertarna, eksoskeleton umumnya tidak cacat. Pada periode pascaganti kulit kedua, bagian-bagian eksoskeleton kaku, dan kepiting mulai meneari pakan. Pada periode waktu antannolting pertarna, karapas hampir seluruhnya kaku kecuali branchiostegites, stemites, carpus dan merus dari kaki jalan dan periode pertumbuhan jaringan. Pada periode waktu antannolting kedua, kampas seluruhnya kaku, bmnchiostegites, stemites dan kaki jalan hampir kaku dan retak jika ditekuk, dan jaringan terus tumbuh. Pada periode waktu antannolting ketiga, seluruh eksoskeleton kaku tetapi mineralisasi pada endokutikula terns berlangsung, lapisan memhran bagian dalam sampai akhir tidak lengkap, pertumbuhan sempurna dan lapisan membmn melekat pada eksoskeleton.
Molting merupakan salah satu faktor penting yang mempengaruhi
konsentrasi total protein dan glukosa hemolimfe beberapa spesies krustasea.
Osmolalitas hemolimfe krustasea sering bervariasi pada
tingkat
siklus moltingyang berbeda. Hal ini mungkin merupakan bagian dari refleksi yang berhubungan dengan pergerakan ion-ion natrium, khlorida, kaisium, magnesium, kalium, dan
Pertumbuhan krustasea dib.tasi oleh keberadaao eksoskeleton yang keras, konsekuensiny. eksoskeleton tersebut secara berkala horus diganti oleh yang b.ru untuk tumbub. P.da sa.t kepiting ganti kulit, seb.gian bobot hilang seb.gai eksuvi.. Kehilangan bobot pada seti.p ganti kulit ini mengakibatkan model pertumbuban kepiting bak.u bersifat diskontinyu karen. hany. terjadi setelah ganti kulit. Ketik. molting, kepiting meningkatkan pengambilan air untuk mengembangkan eksoskeleton baru
dan
menambah ukuran sehelumeksoskeletonny. mengeras (Styrish.ve ef at. 2004).
Selama masa pertumbuhan menjadi dewasa, kepiting bakau akan mengalami
beberapa leaH molting yaitu berkisar 17 sampai 20 kali. Hal ini terjadi karena
rangka luar yang membungkus tubuhny. tidak dapat membesar, sehingga perlu dibuang dan diganti dengan yang lebih besar. Seti.p periode (fase intermolt) pertumbuban kepiting dapat mencapai 20 sampai 30% dari ukuran sernul. (Kuntiyo ef at. 1994). Warner (1977) mengemukakan bahwa pada kepiting yang masih kecil penambaban bobot dapat menc.pai 400 persen. Secara keseluruhan, penambahan hobot pada setiap molting berkisar 3 sampai 44%. Menurut Lavina
(1980), pertumbuhan kepiting bak.u sejak dari telur sampai dewasa dengan lebar karapas sekitar 114.2 mm mernerlukan waktu minimal 369 ban.
Pengaruh Salinitas pada EflSiensi Pemanfaatan Pakan dan Pertumbuhan
Pengaruh s.linitas pada efisiensi pemanfaatan pakan dan pertumbuban dapat terjadi baik secara langsung maupun secara tidak langsWlg. Pada kebanyakan
organisme laut tipe osmoregulator-eurihaline, pengaruh langsung salinitas media
adalah lewat efek osmotiknya pada osmoreguiasi dan kemampuan pencemaan serta absorpsi sari pakan, sedangkan secara tidak: langsung salinitas
mempengaruhi organisme akuatik melalui perubahan kualitas air seperti pH dan
oksigen terlarut (Gilles dan Pequeux 1983; Ferraris ef at. 1986).
Penelitian tentang hubungan .ntara salinitas dan pemanfaatan pakan telah dilakukan oleh beberapa peneliti. Dil.porkan bahw. salinitas telah terbukti mempengaruhi tingkat konsumsi, kecemaan, dan efisiensi pakan pada berbagai jenis
ikan
laut, antara lain Tricnectes maculatus, MugU cephalus, Salmo gairdneribila media ekstemaJ sedikit hipotonik di bawah rentang isoosmotik organisme
akuatik eurihaline (Ferraris el 01. 1986). Akan tetapi, pada krustasea khususnya kepiting bakau, fenomena tersebut belum diketahui.
Pada dasamya pertumbuhan kepiting bakau bergantung pada energi yang tersedia, bagaimana energi itll dipergunakan di dalam tuhuh dan secara teoritis
hanya akan teIjadi bila kebutuhan minimumnya (untuk hidup pokok) terpenuhi. Kepiting bakau memperoleh energi dari pakan yang dikonsumsi dan kehilangan energi sebagai akibat metabolisme. tennasuk: untuk keperluan osmoreguJasi.
Elisiensi pemanfaatan pakan (energi) untuk pertumbuhan sangat bergantung pada daya dukung lingkungannya. Pertumbuhan akan elisien bila hewan itu hidup pada media yang tidak jauh dari titik isoosmotik (Ferraris ef 01. 1986).
Salinitas merupakan salah satu faktor lingkungan ahiotik penting yang mempengaruhi pertumbuhan oragnisme akuatik. Dilaporlcan bahwa kisaran salinitas yang optimum untuk pertumbuhan krustasea berbeda pada setiap sepesies, antara lain IS sampai 25 ppt untuk Penaeus monodon, 20 sampai 30 ppt untuk P. chinensis, 30 ppt untuk P. escelentus, 34 ppt untuk P. merguiensis, 30
ppt uotuk Metapenaeus monoceros
dan
untuk P. vanname; di atas 30 ppt (Kumlu el 01. 200 I; Huynh dan F otedar 2004).Dalam kaitannya dengan osmoregulasi, Jobling (1994) menjelaskan bahwa pembelanjaan energi uotuk: osmoregulasi dapat ditekan apabila organisme
dipelihara pada media yang isoosmotik sehingga pemanfaatan pakan menjadi efisien serta pertumbuhan dapat meningkat Pertumbuhan kepiting bakau secara
internal selain dipengaruhi oleh kelancaran proses ganti kulit, juga oleh tingkat
keIja osmotik (osmoregulasi).
Metabolisme
Metabolisme adalah segala proses reaksi kimia yang teIjadi di dalam tubuh
organisme yang meliputi anabolisme
dan
katabolisme. Dalam proses iniorganisme mendapat, mengubah dan memakai senyawa kimia dari sekitamya
untuk mempertahankan kelangslUlgan hidup. Anabolisme dibedakan dari
katabolisme dalam beberapa hal. Anabolisme merupakan proses sintesis molekul
proses penguraian molekul besar menjadi kecil. Anabolisme adalah proses yang membutuhkan energi sedangkan katabolisme melepaskan energi. Anabolisme
merupakan reaksi reduksi, sedangkan katabolisme adalah reaksi oksidasi. Hasil akhir anabolisme seringkali merupakan senyawa pemula untuk. proses katabolisme (Wirahadikusumah 1985).
Perubahan salinitas media dapat mempengaruhi taju metabolisme dan tingkah laku kepiting bakau. Peningkatan laju metabolisme ditandai dengan peningkatan taju konsumsi oksigen dan penurunan aktivitas meneari pakan.
Selain itu, terjadi perubahan pola respirasi sebagai bentuk respons penyesuaian metabolik terhadap tekanan osmotik dan pengaturan keseimbangan kornposisi ion terhadap carran ekstraselluler (Vemberg 1983). Hubungan antara salinitas dan
proses fisiologis krustasea dapa! menjadi variabel yang ekstrim (Rowe 2002). Verslycke dan Janssen (2002) mengemukakan bahwa ketika organisme ditempatkan pada medium yang lebih tinggi atau tekanan osmotik rendah, proses yang membutuhkan energi terjadi untuk memelihara tekanan osmotik internal agar konstan. Glukosa menghasilkan energi dengan cepat dalam bentuk A TP melalui proses glikolisis dan fosforilasi oksidatif.
Semua energi yang diperlukan untuk anabolisme dan katabolisme berasal dari hasil proses oksidasi zat-zat makanan dalam seL Tingkat metabolisme ini dapat ditaksir secara tidak langsung yaitu dengan mengukur tingkat konsumsi oksigen yang dipergunakan dalam proses oksidasi. Penentuan tingkat konsumsi oksigen merupakan suatu teknik pengujian yang biasa digunakan untuk menaksir laju metabolisme. Konswnsi oksigen dapat digunakan sebagai pendekatan tingkat metabolisme yang praktis karena jumlah panas yang dihasilkan untuk setiap liter oksigen yang digunakan dalam metabolisme hampir konstan terlepas dari apa yang teroksidasi (protein, karbohidrat, atau lemak) dan dapat dikonversikan ke dalam nilai energi. Taksiran metabolisme ini juga penting dalam kajian bioenergetika karena merupakan suatu bagian yang penting dari persamaan neraca energi (Schmidt-Nielsen 1990; Lemos dan Phan 2001).
Tingkat konsumsi oksigen dipengaruhi oleh dua faktor yaitu faktor ekstemal
Faktor internal adalah spesies, stadia, aktivitas, jenis kelamin. reproduksi. dan
molting. Jika organisme berada pada medium yang tekanan parsial oksigennya lebih rendah dari lingkungan (ambienl), maka untuk mencukupi kebutuhan
oksigennya dilakukan pemompaan air yang lebih besar melalui peningkatan
frekuensi pergerakan operkulum. Peningkatan CO, lebib besar pengaruhnya pada
peningkatan gerakan operkulum dalam respirasi dibandingkan dengan kandungan oksigen (Kumlu el af. 2001; Verslycke dan Janssen 2002; Villareal af af. 2003).
Adaptasi lingkungan untuk hidup, termasuk kisaran salinitas yang lebar melalui efisiensi osmoregulasi dan fisioiogi respirasi (hyper-hypo-osmoregulator).
Mekanisme adaptasi ini adalah mengkonsurnsi energi. Perubahan metabolisme
energi secara umum, aican rnempengaruhi karakteristik kebidupan organisme
selanjutnya seperti pertumbuban dan reproduksi (Verslycke dan Janssen 2002).
Fisika Kimia Air
Suhu
Suhu merupakan salah satu faktor abiotik penting yang mempengaruhi
aktivitas, oafsu makan, konsumsi oksigen, iaju metabolisme. kelangsungan hidup,
pertumbuhan, dan molting krustasea (Kumlu el af. 200 I; Whiteley ef af. 2001; Hoang ef al. 2003; Villareal el al. 2003; Zacharia dan Kakati 2004; Xiangli el al.
2004; Kumlu dan Kir 2005).
Hubungan antara laj u pertumbuban kepiting dan suhu telah dilapnrkan oleh beberapa peneliti bahwa laju pertumbuhan proporsional dengan suhu air media.
Boeuf dan Payan (200 I) mengemukakan bahwa suhu dan salinitas adalah faklor
yang secara langsung menentukan peningkatan atau penurunan pertumbuhan.
Pengaruh utama suhu adalah meningkatkan laju pergesekan intermolekular dan
laju reaksi-reaksi kimia (Reiber dan Birchard 1993).
Di antara faktor-faktor lingkungan, suhu merupakan faktor yang paling
berpengaruh pada pertumbuhan dan molting (Hoang ef al. 2003). Perairan yang mempunyai suhu tinggi cenderung akan meningkatkan pertumbuhan dan memperpendek masa interval molting krustasea (Hoang et al. 2003; Xiangli et af.
2004). Fenomena ini tetjadi pada Penaeus semicufalus yang pertumbuhan dan
rendab (Kumlu dan Kir 2005). Menurut Kuntiyo el al. (1994) subu yang optimun untuk pertumbuhan kepiting bakau adalab 26 sampai 32 "C.
Ke.saman (pH)
Boyd (1990) mengemukakan babwa pH yang didefinisikan sebagai logaritma negatif dan konsentrasi ion hidrogen
(If'),
merupakan indikator keasaman serta kebasaan air. Nilai pH ini penting untuk dipertimbangkan, karena dapat mernpengaruhi proses dan kecepatan reaksi kimia di dalam air serta reaksibiokomia di dalam tubuh kepiting bakau. Wang ef al. (2002) mengemukakan babwa pada pH rendah dan tinggi teIjadi peningkatan penggunaan energi atau penurunan produksi energi dan penahananlpeoekanan metabolisme energi aerohik.
Jika orgamsme dipelihara pada pH rendab maka jumlab mukus pada permukaan insang akan meningkat. Peningkatan mukus tersebut akan mengganggu pertukaran gas pada saat respirasi dan pertukaran ion melalui insang. Keasaman rendah akan mengganggu keseimbangan asam-basa darah dan
menurunkan konsentrasi NaCI dalam darah yang pada akhimya akan mengacaukan metabolisme tubuh organisme. Efek lebih Ian jut adalah kerusakan
insang sehingga proses respirasi dan keseimbangan asam-basa dalam darah
terganggu (Boyd 1990), penurunan laju metaholisme, dan dapat mempengaruhi potensi toksin seperti logam-logam berat (Wang el at. 2002).
Jika
perairan bersifat asam (pH rendah), organisme (ikan dan krustasea)dapat mengalami kelambatan pertumbuhan dan merusak pengaluran ion (Wang
ef al. 2002), daya racun nitrit .kan meningkat (Chen dan Cheng 2000), sedangkan
pada pH tinggi daya racun amonia menjadi meningkat (Wang el aI. 2002).
Chen dan Chen (2003) merangkum beberapa basil penelitian tentang toksitas akut pH pada krustasea. Dijelaskan bahwa pH rendab memperlambat pertumbuhan Penaeus monodon. mengganggu pengaturan ion
pada
crayfish dantiger prawn, ketidakseirnbangan asam basa pada crayfish dan udang air tawar. Menurut Kuntiyo el al. (1994) dan Christensen el al. (2005), agar pertumbuhan maksimal, kepiting bakau sebaiknya dibudidayakan pada media dengan pH antara
Oksigen Terlarut
Oksigen ter1arut merupakan salah satu faktor hngkungan yang sangat esensial yang mempengaruhi proses fisioiogis organisme akuatik (Warner 1977;
Cheng ef al. 2003). Secara umum, kandungan oksigen terlarut rendah
«
3 ppm) akan menyebahkan nafsu makan organisme dan tingkat pernanfaatannya rendah, berpengaruh pada tingkah 1aku dan proses fisio1ogis seperti tingkat ke1angsungan hidup. pemafasan, sirkulasi, makan, metabolisme, molting,dan
pertwnbuhankrustasea. Bila kondisi ini berlanjut untuk waktu yang relatif lama konsumsi
pakan akan berhenti dan akibatnya pertumbuban menjaeli terhenti (Boyd 1990; Cheng ef al. 2003).
Walaupun kepiting bakuu dapat hidup pada konsentrasi oksigen terlarut yang rendah (Warner 1977), konelisi tersebut sangat berbahaya karena dapat
menyebabkan stress bahkan kematian. Penurunan ketersediaan oksigen
menyebabkan ketidakmampuan organisme untuk mendukung kebutuhan energi
tinggi bagi organisme untuk makan dengan baik (Jobhng 1994).
Sehubungan dengan variabel hngkungan, subu dan salinitas berpengaruh pada konsentrasi oksigen terlarut dalam air. Peningkatan suhu dapat menyebabkan
kadar oksigen terlarut menjadi rendah. Demikian pula haloya dengan salinitas,
kelarutan oksigen akan rendah apabila tingkat salinitas tinggi atau sebaliknya
(Riley dan Chester 1981).
Kebutuban oksigen terlarut untuk tiap jenis organisme air herbeda,
bergantung pada jenis yang mentolerir fluktuasi (naik-turunnya) oksigen. Pada
umumnya semua organisme yang dibudidayakan (kepiting. udang, ikan) tidak
mampu mentolerir perubahan fluktuasi oksigen yang ekstrim (mendadak). Oleh sebab itu, untuk mengbasilkan pertumbuban kepiting bakuu yang dibudidayakan secara maksimal. kandungan oksigen terlarut harus selalu dipertahankan dalam
kondisi optimun. Untuk budidaya kepiting bakuu agar pertumbubannya baik maks
kandungan oksigen sebaiknya lebih besar dari 3 ppm (Kuntiyo ef al. 1994;