KAJIAN GENETIK TERHADAP TNDUK DOMBA
PRIANGAN PERlDl DlTlNJAU D A M ASPEK
KUANTITATIF DAN MOLEKULER
Oleh
Bess
TiesnamurtiDiseriasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi nlau Ternak
PROGRAM PASCASARJANA
lCNSTITUT
PERTANIAN
BOGOR
KAJIAN GENETIK TERHADAP INDUK DOMBA
PRfANGAN PERTDT DITmJAU DART ASPEK
KUANTITATTF DAN
MOLEKIJLERABSTRAK
BESS TIESNAMURTI. Kajian Genetik terhadap Induk Domba kiangan Peridi ditinjau dari Aspek Kuantitatif dan Molekuler. Dibimbing oleh Harimurti Martojo sebagai ketua, Adi Sudono, Sri Soepraptini hhmjoer, Muladno dan Subandriyo masing-masing sebagai anggota.
Pengamatan terhadap produksi susu domba Priangan mempergunakan 74, 62 dan 47 ekor induk selama tiga periode beranak dilakukan den- metode penimbangan anak sebelum dan sesudah menyusu pada induk, dimana anak dipisahkan dari induk selama enam jam dimulai pada pukul 06:00-12:OO. Pengukwan produksi susu dilakukan selama maksimal 1 5 menit dan mencegah anak untuk tidak minum dan membuang urine. Pengukuran dilakukan setiap minggu, dimulai pada hari ketujuh setelah kelahiran sampai ternak disapih pada umur 90 hari, selama tiga periode beranak, dari 1997 - 1999. Estimasi produksi susu satu hari diperoleh dengan mengalikan produksi susu enam jam dengan angka empat, sedangkan produksi susu miyguan dihitung den- mengalikan produksi susu harian dengan angka tujuh, demikian pula total produksi susu didapat dengan menambahkan estimasi produksi susu mingguan.
Tampilan reproduksi induk selama pengamatan menunjukkan persentase beranak 96,3% dengan rataan jumlah anak sekelahiran dan kematian anak prasapih berturut-turut 1,98+0,94 dan 20,2%.
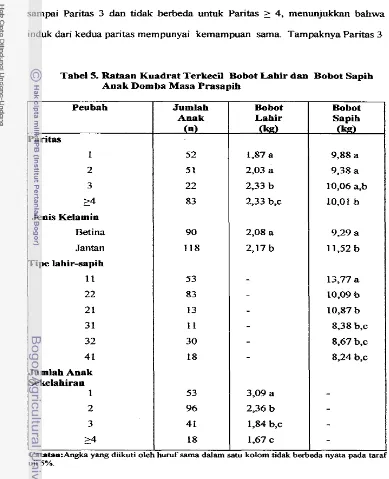
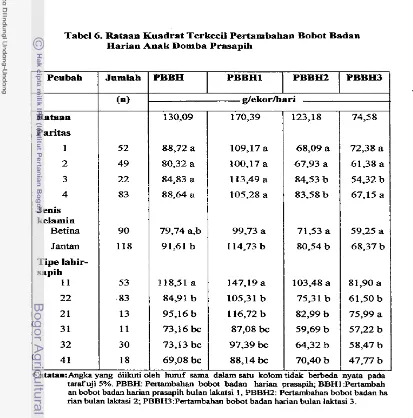
Rataan pertumbuhan an& domba prasapih 130,9+41,8 g/ekorihari dengan pengar& nyata (p<O,05) paritas, jenis kelamin anak
dzLn
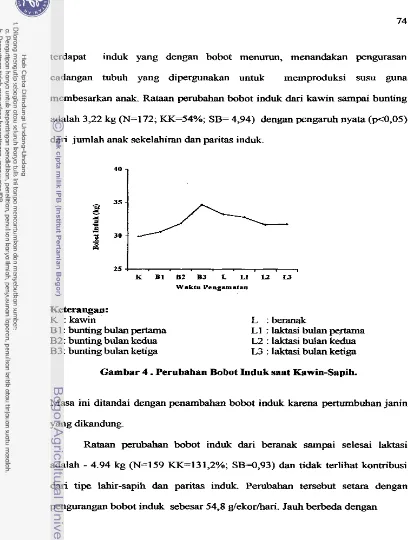
tipe lahir- sapih. Rataan bobot sapih anak 10,62kg, nyata ( ~ 0 , 0 5 ) dipengaruhi paritas, jenis kelamin anak dan tipe lahir-sapih anak.Rataan perubahan bobot induk selama kawin-menyapih yaitu 3,25 kg, nyata(pd,05) dipengaruhi paritas. Selama masa kawin-beranak, rataan perubahan bobot induk (3,22 kg) nyata (p<0,05) dipengaruhi j d a h anak sekelahiran dan paritas, sedangkan perubahan bobot induk s e h a masa laktasi adalah -4,94 kg.
Hasii penelitian mendapatkan rataan produksi susu harian sebesar 5 19,5 dekorhari setara dengan 43,9 kg/Iaktasi, dipengaruhi secara nyata ( F 0 , 0 5 ) oleh paritas dan jumlah anak sekelahiran. Produksi susu tertinggi secara nyata (p<0,05) diperoleh dari induk dengan paritas 3, dan menurun untuk paritas sesudahnya.
dipengaruhi secara nyata (p<0,05) oleh j d a b anak sekelahiran dan paritas. Analisi ragam nilai b yang mencerminkan laju produksi susu awal kurva taktasi dipengaruhi secara nyata ( ~ 4 , 0 5 1 hanya oleh paritas induk, dernikian pula analisis ragam dari laju produksi susu d i a a r kurva laktasi (nilai c) dipengaruhi secara nyata ( ~ 0 , 0 5 ) oleh paritas.
Berdasarkan konstanta yang &pat diketahui, dilakukan estimasi terhadap sifat-sifat laktasi secara individual, yaitu dalam ha1 persistensi {panjangnya waMu puncak produksi dicapai)
.
dengan rataan 22,523 hari. Demikian pula dengan produksi puncak setiap laktasi individu, dapat diestimasi dengan mempergunakan konstanta tersebut dan didapatkan rataan sebesar 708,41 gJekor/hari. Estimasi waktu dicapainya produksi puncak mendapatkan rataan 3,s minggu, sedangkan estimasi total produksi mempergunakan konstanta tersebut mendapatkan rataan 12,7kg/laktasi.Perhitungan nilai rivitabilitas dan heritabilitas total ~roduksi susu dari data y a n g diperoleha mendapatkan angka sebesar 0;61-+0,10 dan 0,25+0,24.sedangka.n pendugaan heritabilitas dan ripitabilitas jumlah anak
GENETIC EVACUATION OF PROLIFIC PRTANGAN SHEEP USLNG QUANTITATIVE AND MOLECULER ASPECTS
ABSTRACT
BESS TIESN-TI. Genetic evaluation of prolific Priangan sheep using quantitative and moleculer aspects, under supervision of Harimurti Martojo,
Adi Sudono, Sri Soepraptini Mansjoer, Muladno and Subandriyo.
This study was planned to investigate the variability of milk production in Priangan sheep using 74, 62 and 47 ewes for 3 consecutive lambing periods, between 1997- 1999, using method of weighing lambs before and shortly after suckling to the dam, where lambs were separated for 6 hours between 06:OOam-12:OOpm. These measurement were made for only 15 minutes to avoid the lambs to drink and urinate. This study was started on day 7 after lambing, at weekly intervals until lambs were weaned from the dam at 90 days of age. The estimation of daily milk production was calculated by multiplying 6 hours estimation with 4, whereas weekly milk production was estimated by multiplying daily estimation with 7, finally total mikk production was tabulated by sumation of weekly milk production.
The reproduction pertormance of ewes during observation showed that the lambing percentage was 96.3% with average litter size of 1.7850.94
and average preweaning mortality of 20.2%.
The average preweaning growth rate of Iambs was 130.9&41.8g/head/day with significant ( ~ 0 . 0 5 ) contribution of parity, sex and type of birth and rearing. The average individual birth weight was 2.39kg with significant ( ~ 0 . 0 5 ) contribution of parity and sex. The average individual weaning weight was 10.62 kg, with significant contribution ( ~ 0 . 0 5 ) of parity, sex and type of birth.
The average ewe body weight changes during mating-weaning and mating-lambing were 3.25 and 3.22kg, respectively, with significant effect ( ~ 0 . 0 5 ) of parity. Ewe body weight changing during lambing-weaning was -4.94kg with significant (F0.05) contribution of parity and litter size.
The result showed that daily milk production was 519.5 &headday equivalent to 43.9 kgflactation, with significant effect ( ~ 0 . 0 5 ) of parity and litter size. The highest milk production significantly affected ( F 0 . 0 5 ) from ewes of the third parity and decline at the following parities.
curve was affected significantly ( ~ 0 . 0 5 ) by dam parity. Finally, the analysis of variation from c parameter w h c h is the final characteristics of lactat~on curve was significantly affected (FO.05) by dam parity.
According to the regression constants found from the equation, several estimation was conducted to Investigate the characteristics of lactation curve ~ndrv~dually, such as persistency which was reflected as the duration of maximum milk production that could be achieved, was 22.58
days The maximum milk production was estimated to be 708.41 g/head/day, whereas the time interval where maximum production reached was estimated to be at 3.5 weeks, and finally total milk production estimation using the parameters was found to be 12.7 kg/lactation.
The repeatability and heritability estimate for total milk production was 0.61 %0.10 and 0.259.24, respectively. The repeatability and heritability estimates of litter size were 0.2320.12 and 0 . 9 7 9 . 3 2 , respectively. The phenotipic correlation among total milk production and total weaning weight was 0.29, respectively, whereas the phenotipic correlation between total milk production and ewe body weight changing during lambing-weaning was -0.49.
Judul : Kajian Genetik terhadap Induk Domba Priangan Peridi Ditinjau dari Aspek Kuantitatif dan Molekuler
Nama : Bess Tiesnamurti
NIM : 96.5036 Program Studi : Ilmu Ternak
Menyetujui,
1. Komisi Pembimbing
Ketua
Anggota
Dr. Tr. Muladno MSc Anggota
-
Dr. Ir. Subandrivo MSc Anggota
Mengetahui,
2. Ketua Program Studi Ilmu Ternak
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa disertasi yang kxjudul :
'KAJIAN GENETIK
TERHADAP
~ U DOMBA PRk4NGAN KPERTDT DITTNJAIJ DART ASPEK KUANTTTATlF DAN
adalah benar rnerupakan hasil karya saya sendiri dan bdum pemah
dipublikasikan. Semua sumber data d m infonnasi yang digunakan telah dinyatakan secara jelas dan diperiksa kebenarannya.
Promovenda adalah putra kedua dari empat bersaudara Kolonel (purn) Soewasono DSOG dan ibu Mardjuni, yang dilahirkan di Yogyakarta, 24 Mei 1957.
Dalam perjalanan hidupnya, lulus sekolah dasar Susteran Putri tahun 1969 di Singkawang, Kabupaten Sambas. Demikian pula SMP Susteran diselesaikan pada tahun 1971 di Pontianak. SMA Loyola 11 Sedes Sapientiae, ditamatkan pada tahun 1975 di Semarang. Memulai lculiah di Fakultas Peternakan U~versitas Diponegoro pada tahun 1976, dan meraih gelar Insinyur pada tahun 1982. Gelar Magister of Science didapatkan dari Universitas ~alipomia, Davis, USA pada tahun 1987 dengan sponsor World Bank (NAR
II).
Promovenda memulai Program Pascasatjana IPB pada bulan September 19% dengan mendapatkan bea siswadari
Proyek ARM I1 di Program Studi Ilmu Ternak.Memulai karier iirniahnya sebagai asisten dosen Ilmu Makanan Ternak pada tahun 1979 di Fakultas Peternakan Universitas Diponegoro. Selanjutnya menapaki jenjang keilmuan di bidang Pemuliaan Ternak Ruminansia KeciI pada tahun 1982 di Lembaga Penelitian Peternakan, Bogor, yang kemudian menjadi Balai Penelitian Ternak. Jenjang fingsional Assisten Peneliti Muda didapatkan pada tahun 1990, selanjutnya naik ke Ajun Peneliti Muda pada tahun 1992 dan meningkat ke jenjang Peneliti Muda pada tahun 1994.
Promovenda menikah dengan kolega sekantor, Lr. Armiadi Semali MSG pada
tanggal 7 April 1989. Kebahagian hidup disempumakan dengan kelahiran putra/putri tercinta yang selalu menjadi sumber semangat, Adhiwienanto Semali dan Nisrina Yuliamurti.
Bogor, April 2002
PRAKATA
Fuji syukur penulis panjatkan kepada Allah SWT, atas segala karunia Nya sehingga karya ilmiah ini berhasil diselesaikan. Topik penelitian yang dipilih dikerjakan pada Lnuun waktu penelitian 1997-1999 adalah mengetahui produktivitas produksi susu domba Pringan peridi ditinjau dari aspek kuantitatif clan molekuler.
Sebagai bagian dari perjalanan hidup, pengungkapan secara tertulis bagi sementara orang merupakan proses yang cukup sulit. Penelitian merupakan suatu rangkaian dari proses yang terdiri atas beberapa urutan, yang dimulai dari "apa". diikuti dengan "mengapa" yang kemudian dilanjutlcan dengan "bagaimana" yang akhirnya ditutup dengan "karena". Mengaktualisasikan serangkaian kata tanya tersebut menjadi sesuatu yang dapat berwujud, merupakan tahap yang memerlukan kegigihan, ketabahan, perjuangan dan sikap kepasrahan terhadap Sang Pencipta yang pada akhirnya memberikan hasil, diibaratkan sebagai penggodokan didalarn kawah
candradirnuka. Pada akhirnya rangkaian proses ini &pat terlewati dm membecikan suatu persembahan kecil bagi bidang keilmuan, khususnya Pemuliaan Ternak Domba di Indonesia.
AkhiruI kata, penulis berharap agar disertasi ini &pat bermanfaat bagi perkembangan kemajuan ilmu peternaksn, khususnya pemuliaan ternak ruminansia kecil.
Bogor, April 2002
UCAPAN TERIMA KASIH
Dengan selesainya karya ilmiah ini, tidak lepas dari bantuan moral dan
material semua pihak yang secara langsung maupun tidak langsung memberikan kontribusinya.
Pertama-tam% penulis mengucapkan rasa terima kasih kepada pimpinan instansi Balai Penelitian Ternak dan Proyek ARM
II
yang memberikan kesempatan untulf mendapatkan tambahan ilmu pengetahuan dan memperluas wawasan ilmiah dengan mengirimkan untuk betajar di Institut Pertanian Bogor.Ucapan terima kasih juga penulis tujukan kepada bapak Prof. Dr. Harimurti Martojo selaku ketua Komisi Pembimbing, yang memberha bimbingan tidak hanya untuk komponen ilmiah, tetapi juga kepada pemanf'aatan untuk mengisi hidup.
Selanjutnya ungkapan terima kasih penulis tujukan kepada Prof Dr. A& Sudono, yang dengan penuh ketelitian dan ketekunan memberikan masukkan untuk penyempumaan karya ilmiah ini.
Terima kasih penulis ungkapkan kepada Ibu Dr. Sri Soepraptini Mansjoer
.
yang memberikan nuansa keilmiahan guna melengkapi kesempurnaan penulisan disertasi ini. Dernikian pula rasa terima kasih kepada Dr. Subandriyo, yang membagipengalaman ilmiah statist* dan analisis data guna memberikan interpretasi penuh makna bagi penelitian ini. Ungkapan terima kasii penulis tujukan kepada Dr. Muladno yang memberikan masukan tentang tee-tehniik pemanfkatan teknologi DNA bagi penelitian pemuliaan guna melengkapi kesempurnaan tulisan ini. Terima kasih penuIis sampaikan pula kepada kotega sejawat, Dr. Ismeth Inouw, yang banyak memberikan masukan untuk pelaksanaan penelitian di kandang pemuliaan domba, guna melengkapi informasi yang dibutuhkan untuk data dasar domba peridi dalarn program payung penelitian Balai Penelitian Ternak. Akhimya, penulis mengucapkan terima kasih kepada penyandang dana APBN 1997-1999, yang banyak mencmahkan dana guna menghidupi kelompok domba diamati.
Tidak pula dapat dilupakan peran serta rekan-rekan peneliti di program Ruminansia Kecil serta para pendukung pelaksanaan penelitian di stasiun Pemuliaan Domba yang merupakan garis depan dari penelitian ini, sdr Kusma Sopandi, Zaenuddin, Atik, Akik, Achmad, Junaedi serta Zulqoyah Laila dari laboratorium Genetik yang banyak membantu melengkapi inhmasi teknik DNA.
DAFTAR IS1
Halaman DAFTARTASEL ... vi
DAFTAR GAMBAR ... vii DAFTAR LAMPIRAN ... viii
TINJAUAN PUSTAKA ... Asal usul Domba di Indonesia
...
Potensi Genetik Domba LokaI...
Sifat Peridi pa& Domba Priangan...
Faktor yang Mempengaruhi Produksi dan Kualitas Susu Domba....
Bangsa Domba ...
...
Jumlah Anak Sekelahiran...
Umur IndukMutu Pakan
...
Hubungan antara Produksi Susu d m Sifat Pertumbuham AnakPnlsapih
...
Parameter Genetik Sifat Produksi...
Mengenali Keragaman Sifat Produksi Susu secara Molekuler...
Pananda DNA dalam Pemetaan S i a t Produksi Tern&...
MATERI DAN METODE...
Lokasi dan Waktu Penelitian...
Materi...
... Metode
...
Definisi IstilahPengamatan Produksi Susu Domba
...
...
Pemeli haraanTernak...
Pemilihan Ternak dan Sistem Perkawinan...
Pengukuran Produksi SusuTampilan Anak Prasapih ... Tampilan Induk ... Deteksi Ragam Genetik Sifitt Produksi Susu dengan Penanda DNA
...
Ekstraksi DNA ... ... Amplifikasi Penanda h4ikrosatelit
... Prosedur Pewarnaan Perak Nitrat
Analisis Data ... Data ...
...
Faktor Koreksi
Faktor Koreksi Anak Prasapih ... Faktor Koreksi Bobot Induk ...
Periode Kawin-bunting
...
Periode Beranak.1aktasi ...... Tampilan Anak Prasapih
...
Tampilan IndukProduksi Susu ...
Kurva Produksi Susu Induk
...
Parameter Genetik Sifat Produksi Susu...
Keeratan Hubungan Penanda Mikrosatelit dengan Produksi Susu ......
HASIL DAN PEMBAHASAN
... Keragaan Reproduksi Induk
Tampilan Anak Prasapih ... Perubahan Bobot Induk
...
... Produksi Susu
Parameter Genetik Sifat Produksi Susu ... Ripitabilitas
. . .
... Hentabditas ......
KorelasiSifat Produksi Susu dengan Bobot IndukMengenali Ragam Produksi Susu Domba Peridi dengan Penanda Mihosatelit ...
Kondisi Optimal Amplifikasi DNA dengan Penanda ... Milcrosatelit
Keeratan Hubungan Sifat Produksi Susu dengan Penanda
Mikrosatelit ...
...
DAFTAR TABEL
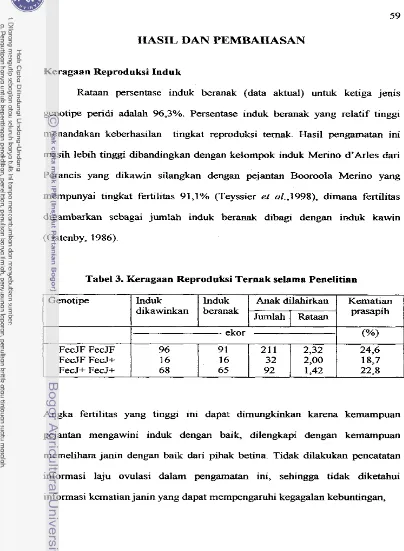
Kelasifikasi Tipe Lahir dan Sapih Anak Domba ... Penanda DNA yang Dipergunakan ... Keragaan Reproduksi Ternak selama Penelitian
...
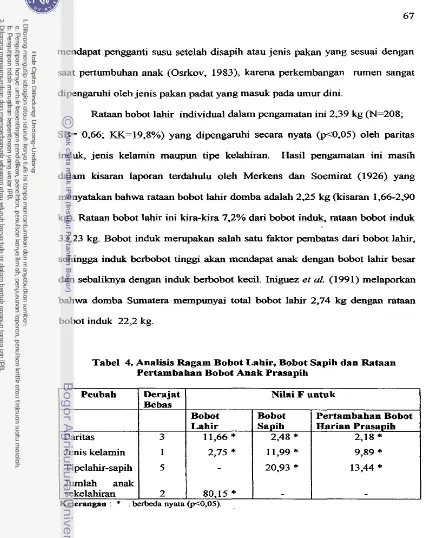
Analisis Ragam Bobot Lahir. Bobot Sapih clan Rataan ... Pertambahan Bobot Anak Prasapih
Rataan Kuadrat Terkecil Bobot Lahir dan Bobot Sapih
Anak Domba masa Prasapih ... Rataan Kuadrat Terkecil Pertambahan Bobot Harian Anak Domba Prasapih ... Analisis Ragam Bobot Kawin, Bobot Beranak dan Perubahan Bobot Induk selama Bunting.Laktasi ... Rataan Kuadrat Terkecil Bobot Kawin, 3obot Beranak dan Perubahan Bobot Kawin-Sapih ... A d i s i s Ragarn Produksi Susu Domba
...
Rataan Kuadrat Terkecil Produksi Susu ... Rataan Parameter Kurva Laktasi pada Domba Priangan .....
Analisis Ragam Konstanta Regresi
Rataan Ken- Komponen Regresi Sesuai dengan
...
Paritas dan Jumlah Anak SekelahiranRataan Produksi Puncak, Persistensi dan Waktu ... Mencapai Produksi Pun&
DAFTAR
GAMBAR
vii
Halaman
Persentase Jumlah Anak Sekelahiran . . .
. . .
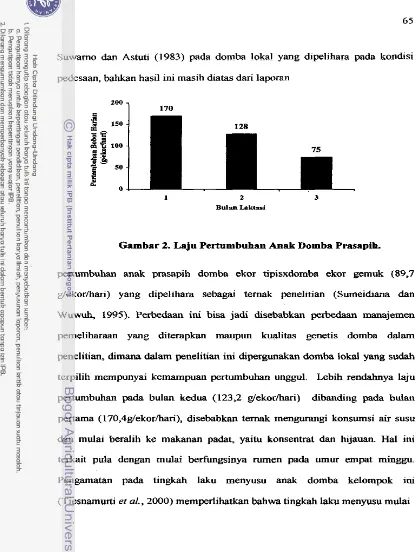
. . . 64Laju Perturnbuhan Anak Domba Prasapih
. . .
. . ..
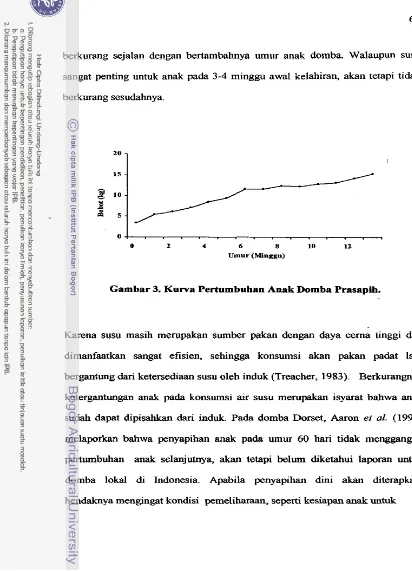
6 5Kurva Perturnbuhan Anak Dornba Prasapih
.
. ..
. . ..
.. . . .
66Perubahan Bobot Induk Saat Kawin-sapih . . .
.
.. . .
. . .,
68Produksi Susu Harian Selarna Laktasi (Aktual vs
Terkoreksi) ... 84
Kurva Laktasi lnduk Domba Menurut Paritas.
. . .
89Kurva Laktasi Induk Menurut Jumlah Anak Sekelahiian.. 90
Hasil Arnplifikasi DNA dengan Penanda OarAE101..
. .
..
. 97Hasil Arnplifikasi DNA dengan Penanda OarJMP08..
. . . .
. 98Runutan Hasil Amplifikasi DNA dengan Penanda
DAFTAR
LAMPIRAN
Halaman
1 Penghitungan Konsentrasi DNA dengan Spektrofotometer
...
1 172 Komposisi Beberapa Reagen dalam Pengerjaan DNA.. ... 1 19
3 Genotipe Domba yang Teramplifikasi dengan Penanda
Mikrosatelit OarAElOl dan JMP08 ... 120
4 Jumlah Perkawinan dan Kelahiran Induk dalam Penelitian
... 121
5 Catatan Jumlah Anak Sekelahiran dan genotipe Domba
PENDAHULXJAN
Hampir seluruh petemakan domba di Indonesia dipelihara secara tradisional dengan pemilikan terbatas, dimana temak domba mempunyai peran penting sebagai penghasil daging, dan sebagai sumber tabungan keluarga yang dapat diuangkan setiap saat, serta sebagai s h b e r pup& organik maupun nilai ekonomis yang ditimbullcan dalam semua aktivitas ini. Tanpa menggunakan tehologi canggih dan dengan input minimal, pekmakan domba dapat bertahan dari tahun ke tahun den= penyebaran mekuas hampir & seluruh propinsi. Dengan hanya pemberian pakan berupa rumput lapang, leylminosa maupun sisa
hasil pertanian, ternak dornba dapat b e r t h
dan
berkernbang biak sepanjang tahun Halini disebabkan o1eh kemampuan genetis domba lokal yang me.mang sudah teruji dan beradaptasi den- baik pada kondisi Iingkungan setempat.Guna menyiapkan pangan hewani dari ternak domba pada masa mendatang, perlu dipersiapkan bibit
temak
dengan tingkat keragaman tinggi darisemua aspek/parameter produksi dan reproduksi. Hal ini diperlukan untuk mengantisipasi pentbatmn kondisi lingkungan yang memerlukan adaptasi ternak
Beberapa parameter gene& penting yang patut dipertimban- daiam
pemilihan temak tersebut adalah sifat adaptif pada kon&si pemeliharaan setempat (lembab panas untuk di Indonesia), kelnampuan berreproduksi s e p j a n g tahun, efislensi produksi tinggi terhadap pakan kualitas pakan rendah dan daya tahan tub& terhadap serangan penyakit
dengan tampilan anak masa prasapih telah dilakukan. Selain bobot lahir dan
agresivitas
anak
dalam menyusu, pruduksi susu induk sangat penting dalam menentukan daya hidup clan pertumbuhan anak prasapih, karena susu merupakan sumber kehidupan utama anak domba. Apabila anak domba mempunyai tampilan bagus pada masa prasapih, &pat diperlarakan bahwa tampilan produksi pada masa mendatang akan bagus pula, karena bobot sapih anak domba erat kaitannya dengan tampilan pada fase hidup selanjutnya..P+oduksi susu domba akan berbeda sesuai dengan jenis domba yang
dipelihara, bangsa domba, jumlah anak sekelrthimn, pengalaman induk dalarn beranak dan menejemen pemeliharaan. Domba tipe perah akan mempunyai total produksi susu dan waktu pemerahan relatif lebih panjang dibanding tipe non
perah, demikian pula diantara domba tip pedaging terdapat ragam produksi susu yang culcup luas. Produktivitas induk domba dapat dilihat dari selang berat&, jumlah an& diiahirkan dan total bobot anak saat disapih a a m kurun waktu tertentu. Faktor genetik dan lingkungan yang mendukung tempat pemeliharaan krnak menentukan tingkat produktivitas domba. Pertumbuhan anak domba sampai disapih menentukan tingkat produktivitas induk
dan
ragam pertumbuhan anak domba pada awal kehidupan sangat ditentukan oleh produksi susu induk.Produksi susu merupakan salah satu sifat kuantitatif yang dipengaruhi
sehingga pengenalannya secara individu relatif s d i t dilakukan dan sangat
bergantung atas ketersediaan mutu pakan yang diberikan dan iingkungan yang kondusif untuk pemeliharaan. Sejauh ini belurn diketahui sifat alamiah dari gen- gen yang mengatur ragam genet& ploduksi susu dan k l u m diketahui gen yang memberikan respon paling menonjol. Seleksi ternak, secara konvensional merupakan upaya untuk mengumpulkan sifat genetis unggul dan mengeluarkan si%t yang tidak dikehendaki, akan tetapi cara ini diketahui makan waktu yang relatif lama, disebabkan panjangnya masa interval genwasi ternak.
Domba Priangan adalah salah satu domba pen& yang sudah beradaptasi
baik dengan kondisi lingkungan Indonesia yang tropis basah. Produktivitas induk domba ini relatif tinggi dilihat dari kemampuan beranak sepanjang tahun, jumlah anak dilahirkan, kemampuan menyapih anak dan daya
tahan
tub& terhadap serangan parasit internal. Domba ini kebanyakan dipelihara secara tradisional didaet-ah Jawa Barat, khususnya di Kabupaten Garut. Pemeliharaan domba di seantero Indonesia diakui atau tidak memberikan peran positif terhadap pendapatan pemilik serta membangkitkan kegiatan ekonomi, sosial dan budaya. Salah satu dari keunggulan genetik domba tersebut antara lain daya tahan tinggi terhadap serangan parasit internal. Selain keunggulan tersebut
Lebih jauh, domba dengan jumlah anak berbeda mempunyai ragam genetik total bobot sapih (sebagai ekspresi kapasitas produksi susu) dari induk dengan jumlah anak berbeda. Oleh karena itu deteksi ragam produksi susu induk merupakan salah satu faktor penting dalam usaha pemeliharaan domba untuk mengetahui kemampuan produktitvitas optimal.
Penelitian ini dimaksudkan untuk mengukur kemampuan genetis produksi susu domba Priangan peridi dan mengenali ragam genetik produksi susu dengan memanfmtkan penanda mikrosatelit DNA.
Manfaat Peneiitian
1. Tnforrnasi tentang kapasitas produksi susu domba Priangan Peridi 2. Korelas~ antara produksi susudengan bobot anak prasapih 3. Sasaran penelitian dalam jangka panjang adalah mempergunakan
hasil penelitian ini sebagai alat bantu seleksi terhadap induk dengan produksi susu bagus pada umur dini
Hipotesis
1. Tingkat keragaman produksi susu induk dengan jumlah an&
sekelahiran berbeda dapat dideteksi dan kemudian &pat dibedakan
secara molekuier melalui penanda mikrosatelit
2.Terdapat korelasi antara keragaman lokus penanda DNA e o s a t e l i t
TLNJAUAN PUSTAKA
Asal usul Domba di Indonesia
Domba yang kini banyak dipelihara mempunyai taksonomi sebagai berikut (Piper clan Ruvinsky, 1997):
Farnili : Bovidae Sub-famili : Caprinae Genera : Ovis
G ~ P : tipe ekor, tipe penutup tub&
Pada awal sebelum terja& proses domestikasi, domba masih hidup liar di
pegunungan. Perbunran hanya dilakukan untuk mendapatkan daging guna pemenuhan hidup sesaat. Pemeliharaan ternak dimulai ketika manusia merasa perlu mempunyai cadangan daging setiap saat diperlukan, sehingga dimuhilah proses domestikasi.
Bangsa domba yang dipeIihara sekarang ini adalah domba tipe perah, pedaging dan pnghasil wol. Pada umumnya domba perah sudah diseleksi secara
khusus sebagai produsen air susu, sedangkm domba pedaging dipelihara hanya untuk memanf- pertumbuhan yang tinggi. Walaupun pada awalnya semua
domba dipelihara hanya untuk produksi daging, dalam proses evolusi terjadi seleksi antar individu terhadap induk yang mempunyai sifat produksi susu tinggi dan akhimya terbentuklah domba @ur penghasil air susu
akan tetapi dengan adanya relief domba di candi Borobudur (circa 800 SM), menandakan bahwa domba sudah dikenal masyarakat sekitamya pada saat itu (Ryder, 1983). Domestikasi domba sudah dimulai pada 2,5 juta tahun yang l a l y pada akhir masa Mesolitik yaitu sekitar 10.00040.000 tahun SM. Domba yang sekarang menyebar di seluruh dunia ini sesungguhnya berasal dari daerah pegunungan Asia Tengah, dimana sebagian menyebar ke arah Barat dan Seiatan
sehingga dikenal sebagai kelompok urzal dan yang laimya menyebar ke Timur dan
Utara yang dikenal sebagai kelompok argali. Terdapat tiga macam dornba berdasarkan asalnyst (bagian Bamt clan Selatan Asia), yaitu 0vz.v muvimon, Ovis ummon dan Ovis orientcrlis. Sebelum terjadi pemisahan daratan antara kepulauan Indonesia dan jazirah Melayu, maka domba yang ada di kawasan tersebut boleh ja& menyebar dari kawasan Asia Tengah (sekarang daerah Tibef Mongolia), kemudian ke daerah Kamboja, Thailand, Malaysia dan kawasan Bamt Indonesia seperti
Sumatera yang pada saat itu masih bersatu dengan Malaysia. Hal tersebut terbuicti
dari jenis domba yang dijumpai di kawasan tersebut adalah dari jenis ekor tipis dengan penutup tubuh bentpa rambut.
Pada masa kolonial Belanda, berbagai importasi ternak dilakukan oleh pemeriniah Hindia Belanda, diantamnya adalah kambing dan domba, terutama ke pulau Jawa sebagai pusat pemerintahan pada saat itu dan Sumatera Barat dengan tujuan untuk meningkatkan kuaiitas domba lokal yang ada (Merkens dan Soemirat,
membawa domba ekor gemuk ke propinsi Sulawesi Selatan dan pulau Madura memberikan kontribusi pada keragaman jenis ternak domba yang ada. Demikian pula setelah masa kemerdekaan, dapat dilihat dari banyaknya imprtasi jenis domba pada
masa Orde Baru dengan tujuan llbma meningkatkan produktivitas ternak domba lokal. Bisa disebutkan antara lain domba yang berasal dari daerah bermusim empat sepem Merino, Suffolk, Dorset, Texel (Natasasmita et UI., 1979) maupun domba dari daerah tropis dengan penutup tubuh berupa rambut, seperti domba St. Croix dan Barbados Slackbelly (Subandriyo etal., 1998).
Potensi Genetik Domba Lokal
Di propinsi J a w Barat terdapat domba Priangan yang merupakan persilangan
antara domba lokal, domba Merino dan domba Kaapstaadt, yang dilakukan pada rnasa pemerintahan kolonial Belanda (Merkens dan Soemirat, 1926). Domba Priangan ini dilaporkan mempunyai kemarnpuan beranak banyak dengan perhmbuhan relatif bagus -n, 1980). Pengamatan lebih jauh memperlihatkan
bahwa sifat beranak banyak secara genetis diatur ad~tif oleh gen major FecJF
(Bradford et al., 1991). Segregasi gen ~ec.T~ dalam suatu popdasi a+n
mengelompokkan ternak ke dalam t i p galur laju kesuburan, yaitu
1) F e d F F e d F : induk domba mempunyai kemampuan beranak 3 4 ; 2) FecJF F&: induk domba dengan kemampuan laju ovulasi atau anak
3) F e d + FecJt: induk dengan rataan laju ovulasi dan jumlah an& 11,7. Jumlah anak domba dilahirkan yang terdapat dalam kondisi perneliharaan
pedesaan di Kecamatan Sukawargi, Garut memperlihatkan rataan 1,92+0,35 ckor
(Sell et al., 1983) dengan &saran jumlah anak sekelahiran antara 1,5-3,O; dengan jumlah anak sekelahiran tertinggi ditemukan pada k e l h r a n temak bulan September- Oktober, sedangkan terendah dijumpai pada kelahiran bulan Jul i. Pengamatan di
Rumah Pemotongan Hewan Bogor, memperlihatkan bahwa jumlah anak sekelahiran terendah ( 1 3 dijumpai pada pemotongan Man September-Oktober, sedangkan pa& bulan lainnya mempunyai rataan jumlah anak sekelahiran 1,7-1.9 (Obst et a[.,
1 980). Keadaan tersebut menandakan bahwa segregasi gen yang bertanggung jawab
terhadap sifat laju kesuburrtn ternyata menyebar cukup luas pada peternakan rakyat, dengan fiekuensi sekitar 60-75 O h (Mason, 1980; Obst et al., 1980). Segregasi gen
utama (major gene) F ~ C J ~ dalanl suatu populasi ditandai antara lain oleh angka heritabilitas dan ripitabilitas yang relatif tinggi dibandingkan pertritungan sifat sempa pada populasi normal (tanpa gen utama). Bradford ef al. (1986) melaporkan
pendugaan ripitabilitas laju ovulasi dan jumfah anak sekelahiran pada kelompok domba lokal ekor tipis berturut-turut adalah 0,80 dan 0,35 untuk domba yang berasal
dari daerah Jawa Barat, Jawa Tengah dan Jawa Timur. Data penelitian untuk
0,53 (Bradford et al., 1991). Untuk kelompok domba yang sama, Tnounu (1996) mendapatkan dugaan ripitabilitas dan heritabilitas laju ovutasi 0,42+-0,03 dan 0,84+0,20; sedangkan dugaan ripitabilitas dan heritabilitas jumlah anak sekelahiran adalah 0,+0,03 dan 0,81+0,19. Dibandingkan dengan sifat serupa tanpa gen utama, maka pendugaan angka heritabilitas jumlah anak sekelahiran domba Merino yaitu 0,12 (Tosh dan Kemp, 1994), &gkan A1-Shorepy dan Notter (1994) mendapatkan dugam heritabilitas jumlah anak sekelahiran domba Dorset O,11 dan 0.2 untuk domba SuEolk (Hanrahan dan Quirke, 1985).
Keunggulan lain dornba lokal Indonesia antara lain daya tahan tubuh terhadap
serangan parasit. Walau belum dipahami mekanisme kerja gen ini, akan tetapi diketahui bahwa domba ekor tipis Jawa dan Sumatera yang digembalakan di perkebunan karet Sumatera Utara mempunyai daya tahan tinggi terhadap serangan parasit internal (Romjali et al., 1998). Keunggulan tersebut &pat dirnanfaatkan secara optimal untuk meningkatkan produktivitas domba, sehingga petani tradisional Indonesia yang jarang menggunakan pengobatan modem- karena harga ti&
terjangkau- dapat mernanfaatkan potensi tersebut ditambah dengan menggunakan ramuan tradisional (Murdiati, 1991). Pengobatan modem, sejauh ini mengandalkan pada obat yang diproduksi oleh industri f m a s i hewan, akan tetapi lama kelamaan
Jambre, 1996) dan residu antibiotika tertinggal, baik &lam feses maupun &lam bagian tub& ternak yang tidak dapat hilang begitu saja. Ini menirnbulkan dampak pencemaran luas pada lingkungaan sekitar. Selain dengan memanfaatkan penggembalan berotasi (rotational grazing),
usaha
petemak sekarang beralih dengan menggunakan cara biologis, yaitu mengenali individu tahan terhadap parasit internal, karena biaya yang dikeluarkan dari strategi ini Iebih menguntungkan dibanding dengan mencari pengobatan baru. Cara lain adalah perkawinan silang antarindividu resisten dan rentan terhadap parasit, sehingga terbentuk keturunan heterozigot clanrelatif tahan terhadap serangan parasit tersebut (Sivarajasingam, 1995). Bahkan suatu proyek kerjasama penelitian antara Indonesia dan Australia diciptakan untuk mengetahui mekanisme kerja gen tersebut, menunjukkan keinginan kuat gum
mempelajari segregasi gen tersebut. Dalam masa mendatang, diharapkan pemanfaatan teknologi DNA dapat dilakukan untuk mengenali individu resisten pada usia dini. Bahkan dengan mengenali kode genetik d h i gen penyandi pada temak
resisten, diharapkan d a p t dilakukan terupi gen terhadap ternak yang rentan penyalcit tersebut (Sivarajasingam, 1995).
Tndonesia yang terletak di daerah equator merupakan aset positif bagi pengembangan ternak domba, karena aktivitas reproduksi tidak dipengaruhi musim,
sehingga secara teoritis domba di daerah ini &pat dikawinkan sepanjang tahun. Dalam suatu survei dan pemantauan selama 10 bulan terhadap petemak di daerah
tahun dengan mayoritas kelahiran tinggi pada bulan Juli, Agustus, September dan
Januari, dimana sebenamya petemak mengin- agar induk dapat beranak pada saat ketersediaan pakan berlimpah yaitu pada awal dan selarna musim huian. Hal ini membuka peluang untuk mengatur perkawinan temak sedemikian mpa, sehingga kelahiran dapat disesuaikan dengan kecukupan pakan guna menunjang rnasa laktasi dan pertumbuhan anak sampai saat sapih (Bell et ucrl., 1983).
Untuk pemeliharaan di Indonesia, potensi perhmbuhan dim kualitas karkas temak I O U temyata Lebih baik dibanhngkan temak hasil persilangan antara domba lokal dan domba impor dari daerah bennusim empat. Dalam kondisi pedesaan
dimana pemberian pakan hanya mengandalkan pada rumput saja, rataan pertumbuhan domba muda adalah 25 g/ekor/hari (Prasetyo, 1992), sedangkan perneliharaan dengan tam- pakan konsentrat mendapatkan rataan pertumbuhan berkisar antara 60,O-91,s g/ekor/hari - (Haryanto, 2000). Natasasmita et al. (1 979) melaporkan bahwa secara kuantitas, tampilan anak domba Priangan murni (8,95 kg)
kalah dibandingkan dengan persilangan antara domba Priangan x Suffolk (9,99 kg) sampai umur 120 hari. Bahkan respons pemberian pakan konsentrat sebanyak 200g/ekor/hari menunjukkan bahwa prestasi pertumbuhan anak domba persilangan (69,5 g/ekor/hi), lebih baik dibanding dengan anak domba Priangan mumi (54,s
dilaporkan, bahwa persentase karkas domba lokal mempunyai rataan sekitar 40-55% dan bobot karkas domba betina 46,4% lebih tinggi dibanding ternak jantan (Arnsar et al., 1984). Dengan materi penelitian yang sama, Duliaman et al. (1984) dan Sugana et a/. (1984) menunjukkan ba.hwa luas otot urat daging mata rusuk
ternak betina lebih lebar dibanding dengan tern& jantan (kisaran 7,X-11,5 cm2 vs 5,9-11,s cmz ). Lebih lanjut, Sitepu et al. (1984) yang mengadakan pengamatan terhadap domba lokal di sekitar Bogor dan persilangannya dengan domba Yorkshire, Polled Dorset dan Suffolk menyatakan bahwa domba lokal lebih unggul dibanding dengan persilangan dalam hal lebih sedikit deposisi lemak dihasilkan dibandingkan dengan domba persilangarz Dari komposisi karkas, terlihat bahwa kandungan daging pada domba lokal (52,2%) temyata lebih tinggi
dibanding domba persilangan (50,8%). Bahkan deposisi lemak pada seluruh tubuh menunjukkan bahwa domba lokal (46,6%) mempunyai deposisi lebih sedikit dibanding dengan domba persilangan (57,9%). Terlihat peluang potensi genetis domba lokal dibandingkan domba impor dalam ha1 perlemakan, dan menghasiikan persentase daging lebih tinggi.
Pada m u m n y a tujuan pemeliharaan domba di Indonesia dhanfhatkan sebagai penghasil daging, sejauh ini belum diketahui pemeliharaan domba dengan manfaat lain, apakah itu sebagai tipe perah, penghasil wol, penghasil rambut, atau
alasan irnportasi temak domba pa& masa kolonial adalah untuk memenuhi kebutuhan akan pupuk kandang bagi perkebunan kopi di Jawa Timur (Merkens dan Soemirat, 1926). Informasi tersebut mengartikan, bahwa pada masa itu pemeliharaan domba mempunyai peran tambahan sebagai sumber pupuk organik.
Sifat Peridi pada Domba Priangan
Sebagai sifat dengan nilai ekonomis tinggi, peridi domba diatur secara kuantitatif oleh banyak gen (Sourdon, 1997) dan peneliti berbondong-bondong untuk menyingkap latar belakang dibalik kemampuan istimewa tersebut. Domba peridi akan menghasilkan j umlah anak per satuan waktu lebih tinggi dibanding den- non peridi, tetapi sifat peridi menimbdkan kekhawatiran akan tingkat kematian yang tinggi apabila tidak diikuti dengan tata laksana yang baik, yang pada akhirnya akan mengurangi produktivitas induk. Si&t peridi tidak hanya diatur secara genetik, tetapi
juga oleh pengaruh pakan, tatal aksana dan lingkungan. Secara u m u m domba dapat dikelompokkan sebagai peridi, kalau k d u k mempunyai paling tidak r&ian jumlah anak sekelahiran 1,75 yang didapat dari minimal tiga M i kelahiran, dengan pofensi menghasilkan rataan anak dilahirkan adalah 2,OO. Beberapa jenis segregasi domba peridi diketahui antara lain yaitu yang diatur oleh gen major, diatur secara kuantitatif
Sifat peridi yang diatur oleh gen major misalnya dijumpai pada heberapa bangsa domba yang mempunyai pola serupa dengan hanya perbeciaan jumlah sel telur diovulasikan, yaitu domba Priangan (pengaruh aditif) dan Booroola Merino (pengaruh dominan) dengan kemarnpuan ovulasi sel telur berturut-turut sebanyak 1-5 (Bradford et al., 1986) dan 1 -1 1 (Piper dan Bindon, 1 985). Ada pula sifat peridi gen major yang diperkiralcan terdapat pada Kromosom X pada domba Romney di Selandia Baru, dengan rataan laju ovulasi 3,08+0,10 dibandingkan dengan rataan laiu
ovulasi domba Rornney kontrol 1,655,11. dimana ada seekor induk yang mempunyai jumlah anak 33 ekor untuk 1 1 kali kelahiran (Davis er al., 1991 ). Pada kelompok domba ini, gen Inverdale akan meningkatkan Iaju ovulasi sebesar 1,O buah dan jumlah anak sekelahiran sebanyak 0,6 ekor.
Selain itu ada pula domba Thoka di Islandia yang kemungkinan mempunyai kerja gen single dengan rataan laju ovulasi 2,i 8 dan 1.59 berturut-brut untuk domba yang mewarisi gen single dan domba normal, dimana satu duplikat gen mempunyai perbeciaan 1,21 buah ova atau 0,64 anakflcelahiran (Eythorsdottir er al., 1991). Demikian pula domba Olkuska di Polandia yang diperkirakan mempunyai segregasi
gen single dengan pengaruh besar terhadap peridi, terlihat dari rataan laju ovulasi 2,83+0,12 (Ivlartyniuk dan Radomska, 1991). Domba Cambridge dan Belcare dari
Sifat peridi yang diatur secara poligenik, menyebar secara luas dari daratan Eropa, Cina dan Afrika antara lain yaitu domba Finn yang brasal dari Finlandia W i j a l a , 1996), domba Romanov yang banyak terdapat di Inggeris (Fahmy, 1996), domba Chios dari Yunant {Hatzirninaglou et a/., 1996), domba D'man dari Maroko (Boujenane, 1996), domba Barbados Blackbelly dari kepulauan Karihia (Rastogi,
1995) clan domba Hu dan Han dari daratan Cina (Feng et u!., 1996).
Menarik untuk disimak, bahwa kejadian peridi pada domba dapat disebabkan antara lain akibat seleksi induk terhadap sifat yang hiperprolifk untuk memenuhi keinginan kelornpok industri perbibitan, yang biasanya dimulai dengan seleksi didalam suatu kelompok kecil untuk meningkatkan j d a h anak sekelahiran atau adanya perkawinan silang sehingga terlihat kontribusi bangsa domba yang memang peridi membentuk kelompok tersebut (Hanrahan, L991), misal pada kelompok domba Cambridge mempunyai populasi dasar domba Finn (terkenal dengan sifat kesuburan tinggi) dengan carnpuran ban* domba Lleyn, Suffoik dan Border Leicester.
Dalam penelitiannya, Davis et al. (2001) melaporkan tipe mutasi sifat peridi pada delapan bangsa domba di dunia, dan mendapatkan bahwa domba Booroola Merino, Priangan clan Garole mempunyai tipe mutasi yang sama (yaitu FecB,
disebabkan oleh mutasi pada reseptor BMPIB(ALK6)), akan tetapi tidak wtuk kelompok domba Olkuska, Thoka, Belclare, Cambridge, Woodlands dan Lacauna,
bahwa kejadian peridi pada beberapa bangsa domba di dunia dapat disebabkan oleh
t i p mutasi berbeda.
Fakior yang Mempengaruhi Produksi dan Kualitas Susu Domba
Bangsa Domba
Produksi susu rnerupakan sifat kuantitatif yang dipengaruhi oleh banyak gen yang masing-masing memberi efek secara bersamaan -pun lokasi dari gen-gen tersebut belum semua dikebhui. Selama ini produksi susu seekor temak tidak dapat diketahui secara pasti, yang diketahui hanyalah perkiraan produksi susu saja. Secara fisiologis, sifat ini merupakan kerjasama antara sistem hormonal, rangsangan mekanis dan ketersediaan prekursor susu untuk dapat te jadinya pembentukan susu (Hafez, 1 980).
Domba tipe perah di Eropa, Laxtra Blond-face clan Lautra Black-f~ce msing- masing mempunyai rataan produksi susu sebanyak 93,9 1 clan 93,2 1 (Gabina et ul..
Chios); 103,0+. 4,l kg Chios x (Awassi-Chios); 109,6-+3,0 kg (Chios x Awassi) dan 109,7+.3,9 kg Chios x (Chios-Awassi). Pengamatan tersebut dimulai hari ke 28
setelah kelahiran anak dan dihentikan apabila produksi susu mencapai 0,2 kg/han. Dari pengamatan tersebut, lama Iaktasi domba Chios dan persilangan berkisar 102-
114 hari. Faktor linghngan seperti tahun pengamatan, musim beranak, kelompok temak yang berbda dan umur laktasi merupakan sumber keragaman yang memberikan pengaruh nyata terhadap produksi selama 90 hari. Gootwine et al. (1995) melaporkan bahwa rataan produksi susu domba Awassi 506 1 untuk 205 hari laktasi, sedangkan persilangan antara Awassi x Booroola Merino d a n silang balik antara Booroola-Me~o x Awassi-BooroolaMerino mempunyai produksi susu 48
dan 63% lebih rendah d i b d n g k a n dengan produksi susu domba Awassi. Disimpulkan bahwa persilangan dengan domba 3ooroola
oofat
meningkatkan jumlah anak dilahi rkan tanpa mernpengaruhi produksi susu.Sakul dan Boylan (1992) melakukan estimasi produksi susu domba pedaging
laktasi), dengan kandungan protein, lemak, laktose dan bahan kering tanpa lemak berturut-turut 6,32; 7,16; 5,27 dan 12,68%. Korelasi antara sekali pengukuran produksi susu dengan total produksi susu adalah 0,64, sedangkan korelasi antara sekaii pengllkuran produksi susu sesudah sapih dengan total produksi susu 0.79. Dalam pengamatan ini, &pat dilihat bahwa pertumbuhan anak bukan merupakan suatu indikator produksi susu, akm tetapi justru satu kali pengukuran produksi susu pada masa laktasi dini mempunyai korelasi yang kuat dengan total produksi susu. Pengarnatan pada domba pedaging SardifS) dan D'man(D) selama I 1 minggu pemerahan memperlihatkan bahwa kapasitas produksi susu ternak dengan prkawinan silang balk DxDS adalah 88,9 kg, sedangkan induk domba bangsa Sardi dan D'man mempunyai produksi susu sebanyak 68,5 dan 81,2 kg (Boujenane dan Lairini ,1992).
Dalam penelitiannya, Jandal (1996) melaporkan kandungan lemak, bahan kering tanpa lemak dan laktose susu domba twrturut-turut adalah 6,72; 10,33 clan 3,7%, sedangkan kandungan susu domba menurut Fadel et al. (1989) untuk komponen lemak, protein, laktose dan bahan kering berturut-turut adalah 83.8; 41,s; 48,4 dan 18,923 g/kg susu segar. Lebih lanjut Boujenane dan Lairini (1992) melaporkan kandungan protein susu mempunyai an* tertinggi untuk ban- domba Dman (8,58 %) dan terendah pa& bangsa persilangan D x DS (6,43 %).
,1959). Pengamatan Sudjatmogo (1998) memperlihatkan rataan produksi susu domba lokal per hari adalah 292 cc dengan total produksi selama 12 rninggu addah
26,5 1, dengan kadar lemak, protein dan laktosa masing-masing 9,18; 2,95 dan 4,78%. Diperlihatkan bahwa produksi susu domba lokal yang diberi dua jenis pakan yang berbeda kualitasnya rnempunyai rataan 250 ccihari (Sitorus et al., 1985). Tiesnamurti dan Inounu (1988) melaporkan potensi produksi susu seminggu s e m i pa& domba lokal Priangan yang dimdai pada minggu ke empat setelah kelahiran anak berkisar 200-329 g/ekor/hari. Terdapat perbedaan lapasitas produksi susu antara domba tipe pedaging dan tipe perah, bahkan antar tipe pedaging. Diantara domba tipe pedagmg maka domba dari daerah tropis seperti Indonesia mempunyai produksi susu jauh dibawahnya. Hal ini dapat dimunglankan karena pehedaan
potensi g e n e penghasil susu temak tersebut.
Jumlah Anak Sekelahiran
laju sintesa air susu. Pada awal laktasi (hari ke ernpat dan 14) induk dengan anak kembar dua mempunyai produksi susu lebih banyak (3.108 dan 4.176 m l / e k o r M ) dibanhngkan dengan anak tunggal (2.136 dan 2.664 mllekorhui). Demikian pula rataan produksi susu induk pada bari ke-50 laktasi untuk induk beranak tunggd dan
kembar dua adalah 1.260 dan 1.584 mllekorhari (Malher clan Anesti, 1994).
Beberapa hasil penelitian produksi susu pada domba peridi memperlihatkan rataan produksi susu harian berturut-turut 1,03; 1,233; 2,33 dan 2,67 kg untuk induk dengan anak 1,2,3 dan 4 ekor pada domba Finn (Antila, 1976). Erokhin et al. (1984) melaporkan rataan produksi susu induk domba Finn di Rusia beranak tunggai, kembar dua dan tiga berturut-turut adalah 0,98, 1,58 dan 2,08 kg/ekor/hari dimana
rataan produksi susu untuk Paritas 1, 2 dan 3 berturut-turut adalah 0,99, 1,84 dan 2-27 kg/ekor/hari. Domba Romanov menampilkan produksi susu puncak pa& hari ke 15-20 laktasi untuk induk berurnur 6-7 tahun dengan rataan berkisar 2,30-2,58 kg/ekor/hari (Kovnerev dan Smirnov, 1988). Dornba Cambridge dengan kerja gen major untuk peridi mempunyai rataan produksi susu dari 50 ml-2,2 1 pada saat kelahuan (Owen, 1996).
Pengamatan pada kelompok domba lokal, diketahui bahwa rataan produksi susu induk dengan anak tunggal dan kembar adalah 212,5 dan 263,s g/ekor/hari (Tiesnamurti dan Inouny 1988) tidak jauh dengan yang dilakukan oleh Sitorus et d.
luteurn 7,l buah) iebih tinggi dibanding dengan induk nirsuperovuiasi (rataan korpus Iuteum 2,8 buah), berturut-turut 486,7 dan 323,21 &ekor/hari. Boujenane dan Lairini
(1992) mendapatkan rataan total produksi susu b e r b a p jenis persilangan domba
Sardi dan D'man dengan pengaruh nyata anak tunggal dan kembar dua berturut-turut 74,9 dan 81.0 kg untuk masa laktasi 10 minggu, demikian pula rataan produksi susu harian domba perah Churra beranak tunggal dan kembar dua berturut-turut 893,1&7,1 dan 932,0+9,7 ml (Gonzalo et al., 1994). Akan tetapi Godfiey et aZ. (1997) tidak
mendapatkan perbedaan nyata produksi susu untuk induk beranak h g g a l dan
kembar dua, yaitu 8,9+0,9 dan 8,9+0,6 kg11 0 hari laktasi pada domba tipe rambut Barbados Blackbelly, St Croix dan Flroida White.
Umur lnduk
Umur induk secara langsung sangat berpengaruh terhadap produksi susu. Induk primipam mempunyai produksi susu lebih sedikit dibandingkan dengan induk yang sudah mempunyai pengalaman beranak Hal
ini
disebabkan karena betina yang baru pertarna kali beranak, belum mempunyai kecukupan perkembangan sel-sel kelenjar ambing yang mendukung produksi susu. Mavrogevis (1996) melaporkan bahwa umur induk domba Chios d m Awassi serta persilangamya secam nyata mempengaruhi total produksi susy rnencapai pun& saat induk laktasi ketigapengamatan tersebut, laktasi ternak secara tidak langsung terkait dengan umur temak. Kremer et ul. (1996) mendapatkan bahtva urnur temak secara nyata mempengaruhi total produksi susu domba Corriedale dan dicapai pada saat induk berumur enam
tahun, yaitu 83,74 1/100 hari Laktasi. Rataan produksi susu domba perah Churra dipengaruhi oleh umur temak, dirnana produksi susu pada paritas pertama tidak sebanyak pada paritas k d u a , yaitu 895,029,3 dan 93 I,O-+8,5 mUekormati (Gonzalo et al., 1994). Akan tetapi, Bencini dan Purvis (1990) melaporkan bahwa pengamatan pada domba Merino dan Meridales (persilangan antara Merino dan Border Leicester, Dorset dan Poll Dorset tidak mendapatkan pengaruh umur terhadap produksi susu dirnana dipergunakan induk dengan umur 2 , 4 dan 6 tahun, yaitu 1,23+0,4 Vekor/hari.
Mutu Pakan
Pakan berrnutu yang diberikan induk pada akhir kebuntingan (6 minggu sebelum kelahiran) d m selama masa laktasi berpengaruh nyaia terfiadap produksi susu induk, dimana Robinson et aZ. (1974) menyatakrtn bafiwa pemberian ko~l~entrat dengan kandungan protein kasar sebesar 10,3; 13,6 dan 16,9% dan enersi metabolis
10 MJ menghasilkan produksi susu 2,4; 2,9 dan 3,l kghari untuk domba Cheviot. Demikian pula pengamatan pada kelompok induk domba Merino (Bencini dan Purvis, 1990) yang memperoleh pakan t a m b b luprn 1 kg/ekor/hari dimufai
hanya mendapat pakan dari padang penggembalaan (1,25+0,06 Vekorhari). Sudjatrnogo (1998) memberikan dua jenis pakan berbeda kandungan protein kasar
(12 dan 15%) dan enersi metabolis 17,4 dan 16,3 kkal pada domba lokal rnendapatkan rataan produksi susu 269,6 dan 323,l g/ekor/hari. Lebih jauh Louca et a1. (1 974) melaporkan bahwa pemberian pakan berkualitas tinggi pada periode akhir kebuntingm yang dilanjutkan dengan pakan b u t u tinggi pada awal laktasi, akan
meningkatkan jumlah susu yang dihasilkan.
Hubungan antara Produksi Susu dan Sifat Pertumbuhan Anak Pra-sap*
Susu menjamin pertumbuhan anak selama masa laktasi, karena pada masa itu sistem pencemaan anak belum sesuai dengan jenis pakan berserat. Pada umur berapa anak siap untuk menerima -put dan pakan berserat lainnya, sangat bergantung pada perkembangan sistem pencemaan. Sebagai patokan, antara umur 5-
6 minggu an& sudah mulai makan sedilut konsentrat dan derlaunan muda. Anak
kembar domba Targhee (Ramsey ef ai.. 1994) mengkonsumsi hijauan iebih banyak &banding anak tunggal (4,s vs 3,4 % bobat b a d a d h i ) . Hal ini inengekspresikan bahwa sebetulnya anak membutuhkan konsumsi susu yang lebih banyak, akan tetapi
kembar lebih banyak diawal laktasi, akan tetapi sebetulnya jumlah susu yang tersedia kurang mencukupi untuk kebutuhan anak kembar pa& akhir laktasi dibandingkan kebutuhan untuk anak tunggal. Terdapat korelasi antara produksi susu induk dengan bobot badan anak, sebesar 0,53 dan 0,57 pada umur laktasi 89 hari
untuk anak tunggal dan kembar dua, ha1 ini mengisyaratkan bahwa hubungan antara
anak tunggal lebih s&t &banding dengan anak kembar dua. Pengamatan lain terhadap domba Cheviot mobinson et al.., 1974) dengan anak kernbar dua memperlihatkan b a h w hubungan antara produksi susu induk dengan pertumbuhan anak prasapih menunjukkan bahwa untuk setiap gram pertumbuhan badan anak dibutuhkan 5,3 g air susu induk
Parameter Genetik Sifat Produksi
Sebagai siFat produksi dengan daya pewarisan rendah, beberapa has11 penelitian menunjukkan angka heritabilitas produksi susu bervariasi antara 0,14,34
(van Vleck et a/., 1993; Falconer. 1986). Pada sapi perah Friesian Holstein Jepang, didapatkan heritabilitas produksi susu berkisar antara 0,26-0,34 (Suzula dan van Vleck, f994), sedangkan nitai heritabilitas produksi susu dan bahan kering tanpa lemak bemtur-turut adalah 0,26 dan 0 27. Perhitungan nilai heritabilitas pada domba
perah Manchega rnendapatkan nilai dugaan total produksi susu 120 hari sebesar
panjang laktasi pada domba perah Churra adalah 0.01 59.01 3.
Secara tmritis, hetitabilitas dan korelasi genetik terletak pacia kisaran -1 sampai I, akan tetapi perhitugan korelasi genetik pada beberapa sifat produksi tertentu mernperoleh angka dugaan diluar &saran itu. Dzakuma et al. (1978) mendapatkan perhitungan heritabilitas bobot anak umur 70 hari domba Hampshire
adalah -0,09-t0,06 dihitung dengan rnetode puterml h r - s i b (saudara tiri sebapak) dengan j d a h data 1.328 catatan. Sebagai konsekuensi dari pendugaan nilai heritabilitas yang negatif, terdapat perhitungan korelasi genetik negatif (-3,2212,J) seperti dilaporkan oleh Vogt er al. (1967) untuk korelasi genetik antara bobot lahir
dan bobot 120 hari pada persilangan domba Hampshire x Rambouillet dengan metode perhitungan saudara kandung (Fullsib) rnenggumkan 534 catatan. Pendugaan korelasi genetik bobot lahir dan bobot 120 hari sebesar 1,07 terdapat pada dornba Columbia dengan menggunakan 516 catatan saudara tin sebapak (Ercanback
d m
Price, 1972). B