Pendugaan Komponen Ragam dan Parameter Genetik
Beberapa Sifat Produksi Puyuh Pada Seleksi Jangka Panjang
(Genetic Parameter and Variance Component Estimation of
Production Traits in Japanese Quail Following Long-Term Selection)
Hamdan*
*)Staf Pengajar Jurusan Peternakan, Fakultas Pertanian, Universitas Sumatera Utara, Jl. Prof. Dr. A. Sofyan No. 3 Medan 20155
Abstract: Estimation of additive variances and parameter genetics were obtained from 38.578 records of two lines of quail over 21 generation selection from 1993 to 2001 in Merbiz Research Station University of Martin Luther, Germany. The purposes of this study were 1) to estimate variances component and genetic parameters 2) to examine stability of genetic parameters from generation to generation following long-term selection for production traits in Japanese quail. The first line of quail was selected on high body weight and small egg weight while the second line was selected only on hight egg weight. Variance component and genetic parameters were estimated using Univariate and Multivariate Animal Model REML procedure using VCE program. The averages of body weight at age of 42 days (BB42), body weight at age of 200 days (BB200), egg weight at the first 11 weeks (BT11), and the last 12 weeks (BT12) in both quail lines increased following 21st generation selection, however the number of eggs laid between 42 days and 200 days of age (JT200) decreased. Additive variance of selected traits still remained following 21st generation selection. Realized heritability estimated for the traits of JT200 and low-egg weight were low i.e. 0,09 to 0,12 and 0,01 to 0,09 respectively , while the trait of low-body weight (line 2) was low to moderate i.e. 0,04 to 0,37, heritability of high-egg weight was moderate i.e. 0,35 to 0,42. Whereas, heritability of high-body weight (line 1) was moderate to high i.e. 0,32 to 0,67.
Key words: Japanese quail, long-term selection, additive variance, genetic parameters, heritability.
Abstrak: Pendugaan komponen ragam dan parameter genetik diperoleh dari 38.578 catatan dari dua galur puyuh hasil seleksi selama 21 generasi mulai 1993 sampai 2001 di Stasiun Penelitian Martin Luther Universitas Merbiz, Jerman. Tujuan penelitian ini adalah 1) untuk menduga komponen ragam dan parameter genetik 2) untuk menguji stabilitas parameter genetik dari generasi ke generasi hasil seleksi jangka panjang beberapa sifat produksi puyuh. galur puyuh pertama diseleksi atas bobot badan besar dan bobot telur yang kecil, sementara galur kedua diseleksi hanya berdasarkan sifat bobot telur yang besar. Komponen ragam dan parameter genetik diduga dengan menggunakan Univariate dan Multivariate Animal Model REML menggunakan VCE program. Rata-rata bobot badan umur 42 hari (BB42), bobot badan umur 200 hari (BB200), bobot telur 11 minggu pertama produksi (BT11), dan 12 minggu terakhir produksi (BT12) pada kedua galur puyuh meningkat sejalan dengan seleksi selama 21 generasi, sementara jumlah telur mulai dari umur 42 hari sampai 200 hari (JT200) menurun. Keragaman aditif sifat yang diseleksi masih tetap bertahan sampai 21 generasi seleksi. Sifat JT200 dan bobot telur ringan, nilai heritabilitasnya rendah, yakni berturut-turut antara 0,09 sampai 0,12 dan 0,01 sampai 0,09, sementara bobot badan ringan (galur 2) heritabilitasnya rendah sampai sedang yakni 0,04 sampai 0,37, bobot telur yang besar heritabilitasnya sedang yakni antara 0,35 sampai 0,42. Sementara, heritabilitas sifat bobot badan yang tinggi (galur 1), sedang sampai tinggi yakni antara 0,32 sampai 0,67.
Pendahuluan
Sejak abad ke-11, puyuh (Coturnix coturnix japonica) dikenal sebagai penghasil daging dan telur, tetapi ia tidak pernah sepopuler ayam karena ukuran tubuhnya yang kecil. Walaupun demikian karena ukuran tubuhnya yang kecil, mudah dipelihara pada kandang pembibitan, prolifik, interval generasi singkat, keragaman genetik dan produktivitas tinggi, serta kemiripan antara puyuh dengan beberapa spesies unggas lainnya untuk beberapa parameter genetik menjadikan puyuh sebagai model hewan percobaan dalam penelitian seleksi unggas, khususnya untuk penelitian seleksi jangka panjang (Maeda et.al., 1997).
Seleksi jangka panjang telah dilakukan untuk menduga parameter genetik bobot badan 4 minggu pada puyuh dengan pemberian pakan yang berbeda untuk melihat batas seleksi. Walaupun keragaman genetik masih bertahan sampai 97 generasi seleksi, namun hasil dugaan menunjukkan adanya kehilangan keragaman genetik aditif sejalan dengan seleksi (Mark, 1996).
Menurut Nestor et.al. (1996), dengan dilakukan seleksi selama lebih dari 30 generasi pada dua galur puyuh yang berbeda, respon bobot badan dan bobot telur meningkat pada galur yang memiliki bobot badan besar ataupun pada galur yang memiliki bobot badan kecil. Aggrey et.al.
(2003), juga telah melakukan penelitian seleksi jangka panjang bobot badan 4 minggu pada dua galur puyuh yang berbeda menghasilkan respon yang tidak simetris pada galur (line) yang memiliki bobot badan besar dan kecil. Hasil seleksi jangka panjang untuk peningkatan bobot badan 4 minggu pada puyuh dipengaruhi oleh perbedaan jenis kelamin
Komponen ragam merupakan faktor penting dalam pendugaan parameter genetik diantaranya heritabilitas, korelasi genetik (Van der Werf dan Boer, 1990). Juga digunakan untuk perancanaan program pemuliaan serta interpretasi mekanisme genetik sifat-sifat kuantitatif (Henderson, 1986). Heritabilitas dan ragam aditif dapat berubah setiap waktu karena beberapa alasan, salah satunya adalah seleksi. Melalui seleksi secara langsung, heritabilitas dan ragam aditif suatu sifat akan menurun (Bulmer, 1971).
Secara teori, nilai heritabilitas berkisar antara nol sampai dengan satu,
tetapi dalam praktiknya jarang dijumpai nilai-nilai ekstrim tersebut (Warwick dan Legates, 1979). Kategori besar kecilnya nilai heritabilitas, dikemukakan oleh Cole (1966) yaitu: h2 <0,20 rendah; 0,20 – 0,40 sedang dan >0,40 tinggi. Sedangkan menurut Preston dan Willis (1974), heritabilitas rendah <0,25; 0,25 – 0,50 sedang dan >0,50 tinggi.
Heritabilitas dapat menduga peningkatan kemajuan genetik yang mungkin diperoleh bila dilakukan seleksi sifat tertentu. Jika heritabilitas suatu sifat memiliki nilai tinggi, berarti performans atau penampilan individu lebih banyak dipengaruhi oleh faktor genetik dibanding dengan faktor lingkungan dan seleksi berdasarkan individu efektif. Heritabilitas yang tinggi juga menandakan aksigen aditif penting untuk sifat tersebut dan sebaliknya jika heritabilitas rendah, maka mungkin aksigen seperti lewat dominant (over dominance), dominan dan epistasis lebih penting (Lasley, 1978).
Nilai heritabilitas suatu sifat akan bervariasi antarpopulasi. Perbedaan variasi tersebut dapat disebabkan oleh perbedaan faktor genetik (ragam genetik), perbedaan lingkungan (ragam lingkungan), metode, dan jumlah cuplikan data yang digunakan (Falconer dan Mackay, 1989). Selain itu juga dipengaruhi oleh waktu generasi seleksi (Marks, 1985). Marks (1996), melaporkan bahwa nilai heritabilitas bobot badan puyuh hasil seleksi jangka panjang dan jangka pendek menunjukkan nilai yang berbeda, selain itu perbedaan nilai heritabilitas juga dipengaruhi oleh galur yang berbeda (galur bobot badan berat dan ringan) dan lingkungan (pakan).
Warwick et.al. (1983), menyatakan heritabilitas akan menentukan perubahan pada sifat yang diseleksi (respon seleksi), korelasi genetik akan mempengaruhi perubahan genetik sifat lain yang tidak diseleksi (respon terkorelasi). Makin tinggi korelasi genetik, makin besar perubahan yang terjadi pada sifat yang berkorelasi. Korelasi genetik dapat berubah dalam populasi yang sama selama beberapa generasi apabila ada seleksi yang intensif. Nilai pendugaan korelasi genetik hanya berlaku pada populasi di mana nilai tersebut diestimasi dan pada kurun waktu tertentu pula.
dilakukan. Nestor et.al. (2000) melaporkan bahwa bobot badan umur 16 minggu pada kalkun berkorelasi negatif terhadap jumlah telur. Francesh et.al. (1997) memperoleh nilai korelasi genetik antara jumlah telur dengan bobot telur pada dua galur ayam layer masing-masing sebesar –0,22 dan – 0,21. Pada puyuh, korelasi genetik bobot badan pada umur empat minggu dengan bobot badan umur enam minggu pada jantan diperoleh 0,62 dan betina sebesar 0,60 (Kuswahyuni, 1989). Sementara Saatci
et.al.(2003) mendapatkan nilai korelasi genetik bobot badan pada umur empat minggu dengan bobot badan umur enam minggu pada puyuh sebesar 0.76.
Penelitian ini bertujuan untuk menduga nilai komponen ragam dan parameter genetik serta mempelajari stabilitas parameter genetik beberapa sifat produksi puyuh dari generasi ke generasi hasil seleksi jangka panjang
Materi dan Metode Penelitian
Waktu dan Tempat
Penelitian ini dilakukan selama 5 bulan dilakukan di Laboratorium Pemuliaan dan genetika Ternak Fakultas Peternakan Institut Pertanian Bogor, mulai bulan Desember 2003 sampai April 2004.
Materi
Materi penelitian berasal dari catatan produksi puyuh dua galur hasil seleksi selama 21 generasi mulai dari tahun 1993 sampai 2001. Dari generasi 1 sampai 2, puyuh dipelihara di kandang penelitian di Leipzig, kemudian dari generasi 3 sampai 21 ditempatkan di Stasiun Percobaan Merbiz, Universitas Martin Luther, Jerman.
Metode
Puyuh dipelihara pada kandang beterai tiga lantai dalam satu kandang. Pakan yang digunakan selama penelitian ada tiga jenis yakni 1) deuka- Putenstarter-P1, kandungan protein kasar 29.80% diberikan pada umur 1-4 minggu; 2) deuka-Landkornendmast, kandungan protein kasar 21.50% diberikan pada umur 5-6 minggu; dan 3) deuka-all-mash-LC, kandungan protein kasar 17.00% diberikan pada umur 7 minggu sampai selesai. Dari generasi 1 sampai 10 puyuh terdiri atas 110 pasang tetua dan pada generasi 11 sampai 21 jumlah pasangan tetua dikurangi menjadi 80 pasang.
Sifat yang dianalisis dikelompokkan ke dalam dua kelompok: 1) sifat yang diukur dari betina saja, dan 2) sifat bobot badan yang diamati pada jantan dan betina. Pengamatan dilakukan pada sifat bobot badan umur 42 hari pada betina (BB42B) dan jantan (BB42J), jumlah telur yang diproduksi dari umur 42 hari sampai 200 hari (JT200), rataan bobot telur pada 11 minggu pertama (BT11) dan rataan bobot telur pada 12 minggu terakhir selama periode bertelur (BT12), bobot badan betina (BB200B) dan jantan (BB200J) pada umur 200 hari.
Galur pertama diseleksi berdasarkan bobot badan yang tinggi dan bobot telur yang kecil, sementara galur kedua diseleksi hanya berdasarkan pada bobot telur yang besar. Pada galur pertama seleksi didasarkan pada indeks sifat dari JT200, BT11 dan BT12, dengan persamaan indeks:
I = Bobot Badan – (11 x Bobot Telur)
Pada galur kedua kriteria seleksi hanya didasarkan pada BT11 dan BT12. Setiap generasi diseleksi dengan intensitas seleksi sebesar 50%. Ternak yang terseleksi dikawinkan dengan perbandingan 1: 1 serta menghindarkan perkawinan saudara kandung dan saudara tiri.
Analisis Data
1. Data deskriptif sifat produksi meliputi nilai rataan, standar deviasi, dan koefesien variasi menggunakan program paket SAS 6.12
2. Analisis Efek Tetap
Efek tetap diuji perbedaannya menggunakan general linear model
dengan SAS 6.12. Model analisis ragamnya sebagai berikut:
ijk k j i
ijk
G
K
S
E
Y
=
µ
+
+
+
+
Untuk sifat pada jantan dan betina.
ij j i
ij
G
K
E
Y
=
µ
+
+
+
Untuk sifat pada betina saja
Yijk = Sifat yang diakibatkan pengaruh generasi ke – i, kandang ke-j, jenis kelamin ke-k dan galat
Yij = Sifat yang diakibatkan pengaruh generasi ke – i, kandang ke-j dan sisaan
µ = Rataan
3. Pendugaan Komponen Ragam, Nilai Heritabilitas, dan Korelasi Genetik
Dihitung Dengan Menggunakan
Univariate dan Multivariate Animal Model Restricted Maximum Likelihood (REML) dengan program paket yang digunakan adalah Variance Component Estimation (VCE) 4.2 (Groeneveld, 1998).
Model matematik yang digunakan sebagai berikut:
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
ny
y
y
2 1 =⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
nX
X
X
0
0
0
0
0
0
2 .⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
nb
b
b
2 +⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
nZ
Z
Z
0
0
0
0
0
0
2 .⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
nu
u
u
2 1 +⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
ne
e
e
2 1 di mana:y = vektor untuk pengamatan (n x 1) b = vektor untuk efek tetap (p x 1) a = vektor untuk efek random (q x 1) X = desain matrik yang berhubungan dengan efek tetap (n x p)
Z = desain matrik yang berhubungan dengan efek random (n x q)
Model ragam-peragamnya adalah:
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
=
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
2 e 2 e 2 a 2 a 2 e 2 a addI
σ
0
I
σ
0
A
σ
σ
AZ'
I
σ
ZA
σ
V
e
a
y
V
di mana:V(y) = ragam fenotip V(a) = ragam aditif V(e) = ragam lingkungan
A = matrik yang berhubungan dengan ragam aditif ternak
I = matrik identitas
Z = desain matrik yang berhubungan dengan efek random
Maka, nilai heritabilitas dapat dihitung dengan persamaan:
V(y)
V(a)
h
2=
=2 e 2 a 2 a
I
σ
Z
ZA
σ
A
σ
+
′
(Quass, 1988).Hasil dan Pembahasan
Keragaan Sifat Produksi
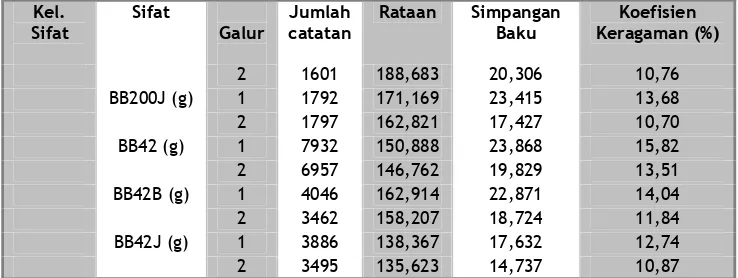
Keragaman sifat produksi pada puyuh selama 21 generasi seleksi sangat bervariasi. Keragaman tertinggi terdapat pada sifat JT200 dengan nilai koefisien keragaman (KK) fenotip untuk galur 1 dan 2 masing-masing sebesar 17,80% dan 21,26%, sedangkan keragaman paling rendah adalah pada BT11 dan BT12 untuk masing-masing galur 1 dan 2 adalah 7,89% dan 8,47%; 8,13% dan 9,24%. Seperti disajikan pada Tabel 1.
Tabel 1.
Nilai rataan, standar deviasi, dan KK sifat produksi puyuh hasil seleksi selama 21 generasi. Kel. Sifat Sifat Galur Jumlah catatan Rataan Simpangan Baku Koefisien Keragaman (%) 1 JT200 (butir)
1 1715 126,670 22,550 17,80
2 1671 120,299 25,575 21,26
BT11 (g) 1 1710 10,280 0,811 7,89
2 1724 11,459 0,931 8,13
BT12 (g) 1 1678 11,253 0,953 8,47
2 1654 12,571 1,161 9,24
BB200B (g) 1 1641 187,771 22,578 12,02
2 1596 188,677 20,267 10,74
BB42B (g) 1 1725 169,314 22,281 13,16
2 1724 160,289 17,436 10,88
2 BB200 (g) 1 3453 179,219 24,512 13,68
2 3398 175,002 22,832 13,05
Kel. Sifat
Sifat
Galur
Jumlah catatan
Rataan Simpangan Baku
Koefisien Keragaman (%)
2 1601 188,683 20,306 10,76
BB200J (g) 1 1792 171,169 23,415 13,68
2 1797 162,821 17,427 10,70
BB42 (g) 1 7932 150,888 23,868 15,82
2 6957 146,762 19,829 13,51
BB42B (g) 1 4046 162,914 22,871 14,04
2 3462 158,207 18,724 11,84
BB42J (g) 1 3886 138,367 17,632 12,74
2 3495 135,623 14,737 10,87
Dari Tabel 1 di atas dapat dilihat bahwa keragaman sifat produksi pada puyuh selama 21 generasi seleksi sangat bervariasi. Keragaman tertinggi terdapat pada sifat jumlah telur, sedangkan keragaman paling rendah adalah pada sifat bobot telur 11 minggu pertama dan 12 minggu terakhir.
Selama 21 generasi seleksi, keragaman sifat produksi yang diseleksi pada puyuh mengalami penurunan dari generasi ke generasi seleksi, kecuali pada sifat JT200 masih menunjukkan fluktuasi yang cukup tinggi dan bahkan cenderung
terjadi peningkatan. Hal ini disebabkan karena sifat jumlah telur tidak menjadi kriteria seleksi, sedangkan bobot telur merupakan kriteria seleksi pada kedua galur. Juga menunjukkan bahwa seleksi memberikan pengaruh terhadap penurunan keragaman dari sifat yang diseleksi. Seleksi bobot telur yang kecil lebih menunjukkan tingkat keragaman fenotip yang lebih kecil dibanding dengan bobot telur yang besar. Ini menunjukkan bahwa seleksi negatif lebih cepat mencapai keseragaman.
0 5 10 15 20 25
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Generasi
KK
(%
)
JT200 BT11 BT12 BB200 BB42
Gambar 1.
0 5 10 15 20 25 30
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Generasi
KK
(%
)
JT200 BT11 BT12 BB200 BB42
Gambar 2.
Koefisien keragaman sifat produksi pada puyuh galur 2 selama 21 generasi seleksi
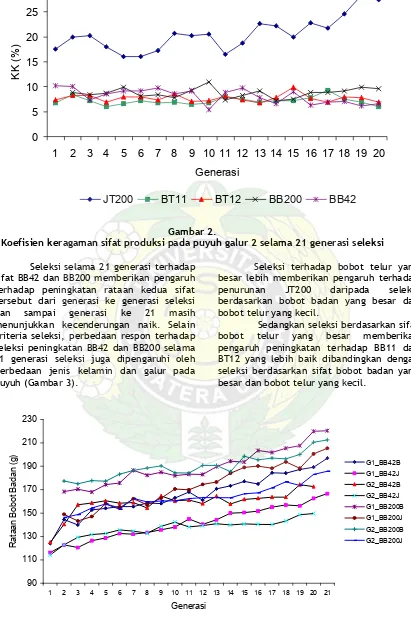
Seleksi selama 21 generasi terhadap sifat BB42 dan BB200 memberikan pengaruh terhadap peningkatan rataan kedua sifat tersebut dari generasi ke generasi seleksi dan sampai generasi ke 21 masih menunjukkan kecenderungan naik. Selain kriteria seleksi, perbedaan respon terhadap seleksi peningkatan BB42 dan BB200 selama 21 generasi seleksi juga dipengaruhi oleh perbedaan jenis kelamin dan galur pada puyuh(Gambar 3).
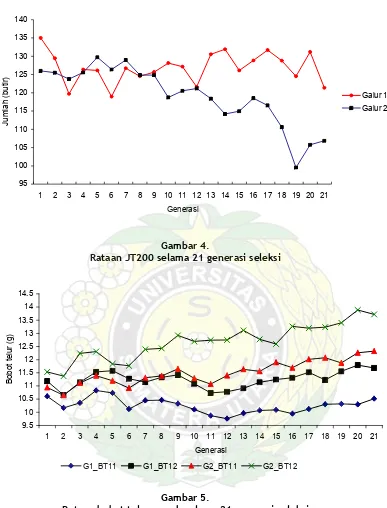
Seleksi terhadap bobot telur yang besar lebih memberikan pengaruh terhadap penurunan JT200 daripada seleksi berdasarkan bobot badan yang besar dan bobot telur yang kecil.
Sedangkan seleksi berdasarkan sifat bobot telur yang besar memberikan pengaruh peningkatan terhadap BB11 dan BT12 yang lebih baik dibandingkan dengan seleksi berdasarkan sifat bobot badan yang besar dan bobot telur yang kecil.
90 110 130 150 170 190 210 230
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Generasi
R
a
ta
a
n
B
o
b
o
t
B
a
d
a
n
(
g
)
G1_BB42B G1_BB42J G2_BB42B G2_BB42J G1_BB200B G1_BB200J G2_BB200B G2_BB200J
Rataan BB42 dan BB200 puyuh selama 21 generasi seleksi
95 100 105 110 115 120 125 130 135 140
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Generasi
Ju
ml
a
h
(b
u
ti
r)
Galur 1 Galur 2
Gambar 4.
Rataan JT200 selama 21 generasi seleksi
9.5 10 10.5 11 11.5 12 12.5 13 13.5 14 14.5
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Generasi
B
o
b
o
t
te
lu
r
(g
)
G1_BT11 G1_BT12 G2_BT11 G2_BT12
Gambar 5.
Rataan bobot telur puyuh selama 21 generasi seleksi
Parameter Genetik
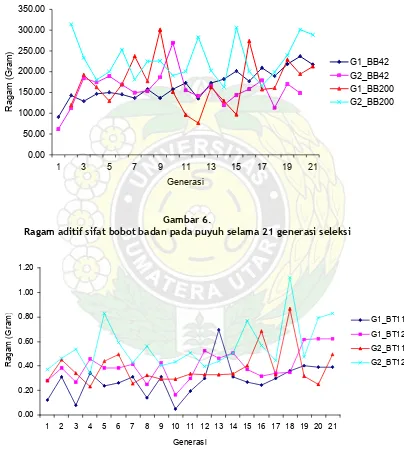
Keragaman Aditif. Selama 21 generasi seleksi, ragam aditif sifat-sifat yang diseleksi pada kedua galur puyuh masih tetap bertahan. Bahkan terlihat masih menunjukkan adanya peningkatan. Untuk sifat bobot badan, baik pada sifat bobot badan umur 42 hari maupun bobot badan umur 200 hari pada kedua galur masih menunjukkan ragam aditif yang masih bertahan sampai 21 generasi seleksi. Bahkan untuk sifat bobot badan puyuh galur 1,
kecenderungan ragam aditif terlihat meningkat, sementara pada galur 2 arahnya lebih mendatar, walau nilai ragam antargenerasi lebih berfluktuasi. Hal ini disebabkan karena sifat bobot badan merukan kriteria seleksi pada galur 1. Seperti terlihat pada gambar 6.
pengaruh menurunkan keragaman. Keragaman aditif BT11 dan BT12 pada kedua galur puyuh masih bertahan sampai 21 generasi seleksi. Seperti terlihat pada gambar 7.
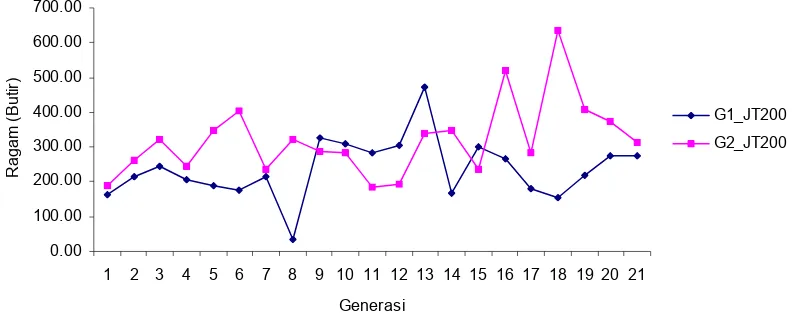
Keragaman JT200 pada galur 1 dan galur 2 juga masih tetap bertahan sampai 21 generasi seleksi. Bahkan pada galur 2, keragam aditif cenderung meningkat,
sedangkan pada galur 1 kecenderungan arahnya relatif lebih mendatar. Hal ini dikarenakan pada galur 2 seleksi hanya didasarkan pada sifat bobot telur, sedangkan pada galur 1 seleksi juga didasarkan atas indeks jumlah telur. Seperti ditunjukkan pada gambar 8.
0.00 50.00 100.00 150.00 200.00 250.00 300.00 350.00
1 3 5 7 9 11 13 15 17 19 21
Generasi
R
a
g
a
m
(G
ra
m
)
G1_BB42 G2_BB42 G1_BB200 G2_BB200
Gambar 6.
Ragam aditif sifat bobot badan pada puyuh selama 21 generasi seleksi
0.00 0.20 0.40 0.60 0.80 1.00 1.20
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Generasi
R
a
g
a
m
(
G
ra
m
)
G1_BT11
G1_BT12 G2_BT11 G2_BT12
Gambar 7.
0.00 100.00 200.00 300.00 400.00 500.00 600.00 700.00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Generasi
R
a
g
a
m
(
B
u
tir
)
G1_JT200 G2_JT200
Gambar 8.
Ragam aditif sifat jumlah telur pada puyuh selama 21 generasi seleksi
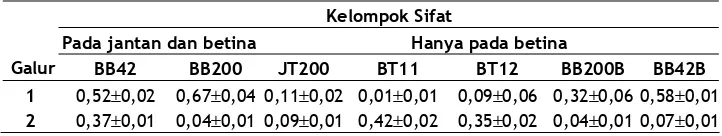
Heritabilitas. Nilai heritabilitas BB42 pada galur 1 dan 2 masing-masing berkisar antara 0,52 – 0,58 dan 0,07 – 0,37. Nilai heritabilitas BB200 pada galur 1 dan 2 masing-masing berkisar antara 0,32 – 0,67 dan 0,04 – 0,07. Nilai heritabilitas BT11 pada galur 1 dan 2 masing-masing 0,01 dan 0,42. Nilai heritabilitas BT12 pada galur 1 dan 2 masing-masing 0,09 dan 0,35. Sedangkan nilai heritabilitas JT200 pada galur 1 dan 2 masing-masing 0,12 dan 0,09, seperti terlihat pada Tabel 2.
Nilai heritabilitas BB42 dan BB200 pada puyuh galur 1 lebih tinggi dibanding dengan galur 2. Namun untuk sifat bobot telur, nilai heritabilitas yang diperoleh pada puyuh galur 1 lebih kecil daripada nilai heritabilitas bobot telur pada galur 2. Sedangkan sifat jumlah telur nilai heritabilitas yang diperoleh pada galur 1 sedikit lebih tinggi dibanding nilai heritabilitas sifat yang sama pada galur 2. Dari nilai heritabilitas yang diperoleh di antara sifat produksi pada puyuh menunjukkan bahwa perbedaan kriteria seleksi memberikan pengaruh terhadap nilai heritabilitas sifat tersebut.
Nilai heritabilitas total jumlah telur yang diperoleh dalam penelitian ini sebesar 0,11± 0,02 pada galur 1 dan 0,09±0,01 pada galur 2, lebih kecil jika dibandingkan
dengan yang diperoleh Schuler et.al. (1998) dengan nilai 0,233±0,035. Menurut Moritsu
et.al. (1997), nilai heritabilitas untuk produksi telur berdasarkan hasil penelitian dalam periode yang berbeda bervariasi antara 0,09 sampai 0,51 dengan nilai rataan 0,25. Sedangkan nilai heritabilitas berdasarkan seleksi selama lima generasi diperoleh nilai untuk galur tingi dan rendah masing-masing 0,06 dan 0,35. Hal ini disebabkan karena jumlah telur dalam penelitian ini bukan merupakan kriteria seleksi.
Korelasi genetik. Korelasi genetik antara sifat yang diukur juga bervariasi di antara kedua galur puyuh. Korelasi genetik antara JT200 dengan BT11, BT12, BB42, dan BB200 pada galur 1 bernilai negatif, seperti yang disajikan pada Tabel 2.
Tabel 2.
Nilai heritabilitas±SE sifat produksi pada puyuh selama 21 generasi seleksi
Kelompok Sifat
Pada jantan dan betina Hanya pada betina
Galur BB42 BB200 JT200 BT11 BT12 BB200B BB42B
1 0,52±0,02 0,67±0,04 0,11±0,02 0,01±0,01 0,09±0,06 0,32±0,06 0,58±0,01
2 0,37±0,01 0,04±0,01 0,09±0,01 0,42±0,02 0,35±0,02 0,04±0,01 0,07±0,01
Tabel 3.
Korelasi genetik sifat produksi puyuh betina hasil seleksi selama 21 generasi
Galur BT11 BT12 BB42 BB200
JT200 -0,13 (0,05)
-0,10 (0,05) -0,02 (-0,01) -0,06 (0,04)
BT11 0,78 (0,78) 0,08 (0,18) 0,03 (0,27)
BT12 0,05 (0,18) 0,07 (0,29)
1 (2)
BB42 0,40 (0,38)
Kesimpulan dan Saran
Kesimpulan
1. Seleksi jangka panjang memberikan pengaruh terhadap peningkatan rataan produksi puyuh pada sifat yang diseleksi dan terkorelasi positif, tetapi memberikan pengaruh penurunan produksi terhadap sifat yang terkorelasi negatif.
2. Keragaman sifat produksi yang diseleksi cenderung mengalami penurunan, namun keragaman aditif masih tetap bertahan dari generasi ke generasi selama 21 generasi seleksi-seleksi.
3. Nilai heritabilitas yang diperoleh dalam penelitian ini untuk sifat jumlah telur dan bobot telur ringan tergolong kecil dengan nilai h2 masing-masing antara 0,09 – 0,12 dan 0,01 - 0,09, bobot badan ringan nilai h2 tergolong kecil sampai sedang yakni antara 0,04 – 0,37 dan h2 sifat bobot telur yang berat tergolong sedang berkisar antara 0,35 – 0,42. Nilai h2 bobot badan berat tergolong sedang sampai tinggi berkisar antara 0,32 – 0,67.
Saran
1. Intensitas seleksi yang kecil dan sistem perkawinan 1: 1 dengan menghindarkan silang dalam (inbreeding) perlu dipertimbangkan dalam melakukan
seleksi jangka panjang untuk tetap mempertahankan ragam aditif.
2. Penelitian seleksi untuk pendugaan parameter genetik pada unggas dapat menggunakan puyuh sebagai pembanding, mengingat nilai heritabilitas untuk beberapa sifat memiliki kemiripan. Namun perlu dipertimbangkan penggunakan metode seleksi dan sistem perkawinan yang sama.
Ucapan Terima Kasih
Ucapan terima kasih saya sampaikan kepada pihak Stasiun Penelitian Martin Luther Universitas Merbiz, Jerman yang telah memberikan kesempatan kepada saya untuk menggunakan data seleksi puyuh selama 21 generasi dalam penelitian ini.
Daftar Pustaka
Aggrey S.E., Ankra-Badug. A., Marks H.L. 2003. Effect of Long-Term Divergent Selection on Growth Characteristics in Japanese Quail. Poultry Sci., 82: 538-542.
Arthur J. A. and Abplanalp H. 1975. Linear Estimates of Heritability and Genetic.
Correlation for Egg Production, Body Weight, Conformation, and Egg Weight of Turkey. Poultry Sci.,
54:11-23.
Cole H.H. 1966. Introduction to Livestock Production. W.H. San Francisco: Freeman and Company.
Falconer D.S., Mackay T.F.C. 1989.
Introduction to Quantitative Genetics. New York: Longman Inc.
Francesh A., Estany J., Alfonso L., Iglesias M. 1997. Genetic Parameters for Egg Number, Egg Weight, and Eggshell
Color in Three Catalan Poultry Breeds.
Poultry Sci., 76:1627–1631.
Groeneveld E. 1998. VCE 4 User’s Guide and Reference Manual Version 1.1. Institute of Animal Husbandry and Animal Behavior. Germany: Federal Agricultural Research Centre.
Henderson C.R. 1986. Recent Development In Variance and Covariance
Estimation. J. Anim. Sci., 63: 208-216.
Kuswahyuni I.S. 1989. Respon Seleksi Jangka Pendek Bobot Badan Umur Empat Minggu Terhadap Penampilan Produksi dan Reproduksi Burung Puyuh. [Disertasi].FPS, IPB. Bogor.
Lasley L.J. 1978. Genetics of Livestock Improvement. 3 rd Edition. New Delhi: Prentice-Hall of India Private Limited.
Maeda Y., Minvielle F., and Okamoto S. 1997. Changes of Protein Polymorphism in Selection Program for Egg Production in Japanese Quail, (Coturnix coturnix
japonica). Japanese Poultry Science, 34: 263-272.
Marks H. L. 1985. Direct and Correlated Responses to Selection forgrowth.. Dalam Poultry Genetics and Breeding. Hill W.G., Manson J.M., Hewit D., ed. Longmangroup Limited, Harlow UK. Hal. 47-57
Marks H. L. 1996. Long-Term Selection for Body Weight in Japanese Quail
Under Different Environment.
Poultry Sci,. 75: 1198-1203.
Mielenz N., Kovac M., Groeneveld E., Preisinger R., Schmutz M., Schuler L. 2002. Genetic Evaluation of Egg Production Traits Based on Additive and Dominance
Models in Laying Hens. 11th European Poultry Conference 6-10 September 2002. Bremen.
Moritsu Y., Nestor K.E., Noble D.O., Antony N.B., Bacon W.L. 1997. Divergent Selection for Body Weight and Yolk Precursor in Coturnix coturnix japonica. 12. Heterosis in Reciprocal Crosses Between Divergently Selected Lines. Polutry Sci., 76: 437-444.
Nestor K.E., Anderson J.W., Patterson R.A. 2000. Genetics of Growth and Reproduction in Turkey. 14 Changes Ingenetic Parameter Over Thirty generations of Selection for Increased Body Weight. Poultry Sci.,
79:445-452.
Nestor K.E., Bacon W.L., Anthony N.B., Noble D.O. 1996. Divergent Selection for Body Weight and Yolk Precursor in Coturnix coturnix japonica. 10 Response to Selection Over Thirty Generations. Poultry Sci., 75: 303-310.
Preston T.R., Willis M.B. 1974. Intensive Beef Production. Second Ed. New York: Pergamon Press Inc.
Quass R.L. 1988. Additive Genetics Model Withgroups and Relationships.
J.Dairy Sci., 71:1338.
Saatci M., Dewi I.A.P., Aksoy A.R., Kirmizibayrak T. 2003. Aplication of REML Procedure to Estimate the Genetic Parameter of Weekly Liveweights in One-to-One Sir and Dam Pedigree Recorded Quail.
J.Anim.Breed.Genet, 120: 23-28.
SAS. 1996. SAS User’s Guide: Statistics. Cary, NC USA: SAS Institute Inc.
Performance Traits of Japanese Quails. Proc.6th WCGALP, Univ. New England, Armidale, Australia. 26:101-104.
Van der Werf J.H.J., de Boer I.J.M. 1990. Estimation of Additive Genetic Variance When Base Populations are Selected. J. Anim. Sci., 68: 3124 - 3132.
Warwick E.J., Legates J.E. 1979. Breeding and Improvement of Farm Animal.
New York: Mc-Grow-Hill Book Company.
Warwick E.J., Maria Astuti J., Hardjosubroto W. 1983. Pemuliaan Ternak.