KARAKTER MOLEKULER BEBERAPA SPESIES TRIPS
SUBORDO TEREBRANTIA DAN IDENTIFIKASI TRIPS
PADA BERINGIN, PALA, DAN SERUNI LAUT
(INSECTA: THYSANOPTERA)
NIA KURNIAWATY
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Karakter Molekuler beberapa Spesies Trips Subordo Terebrantia dan Identifikasi Trips pada Beringin, Pala, dan Seruni Laut (Insecta: Thysanoptera) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
iv
RINGKASAN
NIA KURNIAWATY. Karakter Molekuler beberapa Spesies Trips Subordo Terebrantia dan Identifikasi Trips pada Beringin, Pala, dan Seruni Laut (Insecta: Thysanoptera). Dibimbing oleh PURNAMA HIDAYAT dan AUNU RAUF.
Trips (Ordo Thysanoptera) terdiri dari dua subordo, yaitu Terebrantia dan Tubulifera. Trips anggota subordo Terebrantia telah banyak diketahui menjadi hama pada tanaman budidaya, namun trips subordo Tubulifera lebih sedikit diketahui jenis dan statusnya. Metode identifikasi spesies trips yang sering digunakan adalah dengan karakter morfologi dan karakter molekuler. Penggunaan karakter morfologi relatif lebih mudah dan memerlukan alat yang lebih sederhana dibandingkan dengan karakter molekuler. Karakter molekuler seperti sekuens
DNA Cytochrome Oxidase I mitokondria (mtCOI), merupakan karakter genetik
yang dapat digunakan untuk identifikasi spesies atau konfirmasi hasil identifikasi dengan karakter morfologi. Karakter molekuler juga sering digunakan untuk mempelajari kekerabatan serangga.
Penelitian ini bertujuan untuk mempelajari karakter molekuler beberapa spesies trips subordo Terebrantia yang menjadi hama tanaman dan mengidentifikasi spesies trips subordo Tubulifera yang ditemukan pada tanaman beringin (Ficus benjamina/ Moraceae), pala (Myristica fragrans/Myristicaceae), dan seruni laut (Wedelia biflora/Asteraceae). Pengambilan sampel dilakukan di Kabupaten Bandung, Bogor, Cianjur, Cirebon, dan Kuningan. Pengambilan sampel dilakukan langsung pada tanaman yang bergejala akibat serangan trips. Identifikasi menggunakan karakter morfologi dilakukan di Laboratorium Biosistematika Serangga, sedangkan identifikasi menggunakan karakter molekuler dilakukan di Laboratorium Virologi Departemen Proteksi Tanaman Institut Pertanian Bogor. Tahapan identifikasi karakter morfologi terdiri atas pembuatan preparat mikroskop dan pengamatan karakter eksternal seperti ruas antena, venasi sayap, tergit abdomen, dan seta oseli. Identifikasi dengan karakter molekuler terdiri dari ekstraksi DNA dengan metode CTAB, amplifikasi, sekuensing, dan analisis DNA. Analisis DNA dilakukan berdasarkan homologi sekuen DNA, analisis jarak genetik, dan filogeni.
Hasil penelitian menunjukkan terdapat lima trips subordo Terebrantia yang ditemukan banyak menyerang tanaman hortikultura diantaranya:
Ceratothripoides brunneus, Megalurothrips usitatus, Thrips alliorum, T.
hawaiiensis,dan T. parvispinus. Karakter molekuler berupa sekuens DNA mtCOI
spesies tersebut berturut-turut adalah: 693, 692, 678, 690, dan 668 pb (pasang basa). Nilai jarak genetik nukleotida antar spesies sekitar 0.00-0.401, dan jarak genetik asam amino antar spesies sebesar 0.00-0.268. Hasil identifikasi secara morfologi spesies trips subordo Tubulifera pada tanaman beringin adalah
Gynaikothrips uzeli, pada tanaman pala adalah Pseudophilothrips ichini, dan pada
sedangkan jarak genetik berdasarkan sekuens asam amino antar subordo sebesar 0.230-0.653.
Jarak genetik yang dihasilkan diilustrasikan melalui pohon filogeni. Filogeni berdasarkan karakter morfologi, sekuens nukleotida, dan sekuens asam amino DNA mtCOI menunjukkan spesies-spesies dari kedua subordo terpisah. Penelitian ini menunjukkan bahwa identifikasi dengan karakter molekuler menggunakan sekuens DNA mtCOI mengkonfirmasi dan mendukung hasil identifikasi dengan karakter morfologi.
vi
SUMMARY
NIA KURNIAWATY. Molecular Characters of some Thrips Species belong to Terebrantia and Identification of Thrips on Weeping fig, Nutmeg, and Wedelia (Insecta: Thysanoptera). Supervised by PURNAMA HIDAYAT and AUNU RAUF.
Thrips (Order Thysanoptera) consists of two suborders, namely Terebrantia and Tubulifera. Thrips suborder Terebrantia have been widely known to be pests on cultivated plants, but it is less information known on thrips belong to suborder Tubulifera. Thrips identification methods often use morphological characters and molecular characters. The use of morphological character method is relatively easy and requires simpler tools than the molecular character method. Molecular characters, such DNA sequences of mitochondrial Cytochrome Oxidase I (mtCOI), can be used for species identification or confirmation of the identification using morphological characters. Molecular character is also often used to study insect phylogeny.
The aims of this research were to study the molecular characters of thrips species belong to suborder Terebrantia using mtCOI DNA sequences and to identify the species of thrips suborder Tubulifera from weeping fig (Ficus
benjamina/Moraceae), nutmeg (Myristica fragrans/Myristicaceae), and wedelia
(Wedelia biflora/Asteraceae). Samples were collected from symptomatic thrips
attack plants in districts of Bandung, Bogor, Cianjur, Cirebon, and Kuningan. Morphological identification was conducted at the Insect Biosystematics Laboratory, while molecular identification was done at the Plant Virology Laboratory of the Department of Plant Protection, Bogor Agricultural University. Morphological identification process consists of preparation of slide microscope and observation of morphological characters (antennae, wing venation, abdominal tergite, and ocelli). Molecular identification process consisted of: DNA extraction using CTAB methods, DNA amplification, DNA sequencing, and analysis. DNA analysis was done by DNA sequence homology, genetic distance, and phylogeny construction.
There were five species of thrips suborder Terebrantia frequently found in cultivated crops. They were Ceratothripoides brunneus, Megalurothrips usitatus,
Thrips alliorum, T. hawaiiensis, and T. parvispinus. The length of their mtCOI
DNA sequences were: 693, 692,678, 690, and 668 bp respectively. The range of genetic distance DNA sequences was 0.00 to 0.41, while the range of genetic distance of amino acid sequences was 0.00 to 0.268. Thrips species of suborder Tubulifera found on weeping fig was Gynaikothrips uzeli, on nutmeg was
Pseudophilothrips ichini, and on wedelia was Haplothrips ganglbaueri. The
The thrips identification using molecular characters confirmed the results of identification using morphological characters.
viii
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KARAKTER MOLEKULER BEBERAPA SPESIES TRIPS
SUBORDO TEREBRANTIA DAN IDENTIFIKASI TRIPS
PADA BERINGIN, PALA, DAN SERUNI LAUT
(INSECTA: THYSANOPTERA)
NIA KURNIAWATY
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Entomologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
x
xii
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul tesis ialah “Karakter Molekuler beberapa Spesies Trips Subordo Terebrantia dan Identifikasi Trips pada Beringin, Pala, dan Seruni Laut (Insecta: Thysanoptera)” sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada program studi Entomologi, Sekolah Pascasarjana, Institut Pertanian Bogor. Penelitian ini dilaksanakan di Laboratorium Biosistematika Serangga, dan Laboratorium Virologi Tumbuhan Departemen Proteksi Tanaman sejak bulan September 2014-Agustus 2015.
Penulis mengucapkan banyak terima kasih kepada semua pihak yang telah membantu penulis dalam penyelesaian tugas akhir ini. Ungkapan terima kasih penulis sampaikan kepada Dr Ir Purnama Hidayat, MSc dan Prof Dr Ir Aunu Rauf, MSc selaku dosen pembimbing yang telah memberikan bimbingan, arahan, dan saran yang sangat bermanfaat sejak awal penelitian hingga akhir penyusunan tesis ini. Terima kasih juga penulis sampaikan pada Direktorat Jenderal Pendidikan Tinggi yang telah memberikan beasiswa kepada penulis melaui program beasiswa Fresh Graduate 2013 dan Dana Hibah Penelitian BOPTN No. 319/IT3.41.2/L2/SPK/2014.
Penghargaan dan terima kasih penulis sampaikan khususnya kepada Ayahanda Nasrudin, Ibunda Suciati, adinda Ramadhani, dan Muhammad Soni yang telah memberikan dukungan moral maupun materiil, kasih sayang dan doa restu kepada penulis. Terima kasih juga penulis sampaikan kepada Prof Dr Ir Sri Hendrastuti Hidayat, MSc selaku Kepala Laboratorium Virologi yang telah mengizinkan penulis bekerja di Laboratorium selama penelitian. Terima kasih kepada Ibu Sari Nurulita, MSi yang telah membantu penulis bidang molekuler, membuat database, membantu dalam mengolah data, dan diskusi yang sangat berharga bagi penulis. Rekan-rekan Pascasarjana Entomologi 2012, 2013, 2014, dan 2015, rekan-rekan di Laboratorium Biosistematika Serangga, Ibu Aisyah dan Mbak Atiek serta rekan-rekan lain yang tidak bisa disebutkan satu persatu
Semoga hasil penelitian ini dapat bermanfaat sebagai informasi dalam bidang pertanian dan dapat digunakan sebagai acuan untuk penelitian selanjutnya. Penulis menyadari masih terdapat kekurangan. Kritik dan saran sangat diharapkan oleh penulis untuk perbaikan kegiatan selanjutnya.
Bogor, Agustus 2016
DAFTAR ISI
Identifikasi Trips berdasarkan Karakter Morfologi 5
Identifikasi Menggunakan Karakter Molekuler 6
III KARAKTER MOLEKULER TRIPS SUBORDO TEREBRANTIA 8
Abstrak 8
IV IDENTIFIKASI TRIPS SUBORDO TUBULIFERA PADA TANAMAN
BERINGIN, PALA, DAN SERUNI LAUT 31
xiv
DAFTAR TABEL
3.1 Tanaman inang dan lokasi ditemukannya C. brunneus dan M.
usitatus 13
3.2 BLAST-N DNA mtCOI C. brunneus dan M. usitatus penelitian
dengan sampel dari GeneBank 15
3.3 Jarak genetik sekuens nukleotida DNA mtCOI C. brunneus dan M.
usitatus pada penelitian dengan sampel dari GeneBank 16
3.4 Jarak genetik asam amino DNA mtCOI C. brunneus dan M.
usitatus penelitian dengan sampel dari GeneBank 16
3.5 Posisi nukleotida sekuens DNA mtCOI C. brunneus dan M.
usitatus dengan database GenBank yang menunjukkan variasi 17
3.6 Posisi sekuens asam amino DNA mtCOI C. brunneus dan M.
usitatus dengan database GenBank yang menunjukkan variasi 18
3.7 Inang dan lokasi ditemukannya T. alliorum, T. hawaiiensis, dan T.
parvispinus 19
3.8 BLAST-N DNA mtCOI T. alliorum, T. hawaiiensis, dan T.
parvispinus 21
3.9 Jarak genetik sekuens DNA mtCOI T. alliorum, T. hawaiiensis, dan
T. parvispinus 22
3.10 Jarak genetik sekuens asam amino DNA mtCOI T. alliorum, T.
hawaiiensis, dan T. parvispinus 22
3.11 Variasi sekuens nukleotida DNA mtCOI T. alliorum, T.
hawaiiensis, dan T. parvispinus 23
3.12 Variasi asam amino sekuens DNA mtCOI T. alliorum, T.
hawaiiensis, dan T. parvispinus 24
4.1 Tanaman inang dan spesies trips subordo Tubulifera yang
ditemukan pada penelitian 35
4.2 BLAST DNA gen COI pada tiga spesies trips menggunakan
program BLAST-N (www.ncbi.nlm.nih.gov) 38
4.3 Jarak genetik sekuens nukleotida DNA mtCOI tiga spesies trips
subordo Tubulifera 38
4.4 Jarak genetik asam sekuens amino DNA mtCOI anggota subordo
Tubulifera 38
4.5 Variasi susunan nukleotida DNA mtCOI tiga spesies trips subordo
Tubulifera 40
4.6 Variasi sekuens asam amino DNA mtCOI tiga spesies trips subordo
Tubulifera 40
4. 7 Jarak genetik sekuens nukleotida DNA mtCOI spesies trips
subordo Tubulifera dan Terebrantia 45
4. 8 Jarak genetik sekuens asam amino DNA mtCOI spesies trips
DAFTAR GAMBAR
2.1 Posisi DNA mtCOI T. imaginis (Shao dan Barker 2003) 7 3.1 Karakter morfologi C. brunneus (a) antena 8 ruas, (b) Metanotum
tanpa companiform sinsila, (C) sayap depan berwarna coklat dengan venasi lengkap, (d) Seta oseli I berbaris secara vertikal, (e) deretan microtrichia yang lengkap pada tergit abdomen ruas 13 3.2 Karakter morfologi M. usitatus (a) antena 8 ruas, (b) Metanotum
tanpa companiform sinsila, (C) sayap depan berwarna gelap terang, (d) seta oseli III muncul pada garis segitiga oseli, (e) deretan
microtrichia yang VIII hanya sebagian, (f) seta S1 diatas 14 3.3 Hasil visualisasi DNA C. brunneus dan M. usitatus menggunakan
primer universal (M) Marker 1 kb (Thermo Scientific, US), (1)
Kontrol positif (C. brunneus), (2) C. brunneus, dan (3) M. usitatus 15 3.4 Filogeni DNA mtCOI C. brunneus dan M. usitatus menggunakan
software Mega dengan pendekatanUPGMA bootstrap 1000x (a)
berdasarkan sekuens nukleotida, (b) sekuens asam amino 16 3.5 Kepala dan antena (a) T. alliorum, (b) T. hawaiiensis, (c) T.
parvispinus 20
3.6 Metanotum (a) T. alliorum, (b) T. hawaiiensis, (c) T. parvispinus 20 3.7 Sayap (a) T. alliorum, (b) T. hawaiiensis, (c) T. parvispinus 20 3.8 Hasil visualisasi DNA genus Thrips menggunakan primer universal
(1) Marker (1kb Thermo Scientific, US), (2) Kontrol negatif, (3) Kontrol positif (T. parvispinus), (4) T. alliorum, (5) T. hawaiiensis,
dan (6) T. parvispinus 21
3.9 Filogeni DNA mtCOI genus Thrips menggunakan software Mega dengan pendekatan UPGMA (a) berdasarkan sekuens nukleotida,
(b) berdasarkan sekuens asam amino 22
4.1 Imago, (a) G. uzeli, (b) H. ganglbaueri, (c) P. ichini 36 4.2 Sayap depan, (a) G. uzeli, (b) H. ganglbaueri, (c) P. ichini 36 4.3 Metanotum, (a) G. uzeli, (b) H. ganglbaueri, (c) P. ichini 36 4.4 Pronotum, (a) G. uzeli, (b) H. ganglbaueri, (c) P. ichini 36 4.5 Ujung abdomen, (a) G. uzeli, (b) H. ganglbaueri, (c) P. ichini 37 4.6 Hasil visualisasi DNA mtCOI trips subordo Tubulifera
menggunakan primer universal (M) Marker 1 kb (Thermo Scientific, US), (1) Kontrol positif (G. uzeli), (2) G.uzeli, (3) H.
ganglbaueri, (4) P. ichini 37
4. 7 Filogenetik trips subordo Tubulifera menggunakan sekuens DNA
mtCOI dengan metode UPGMA, (a) nukleotida (b) asam amino 39 4. 8 Filogeni berdasarkan karakter morfologi menggunakan program
NTSys ver 21 dengan pendekatan UPGMA 43
4. 9 Filogeni berdasarkan sekuens nukleotida DNA mtCOI spesies trips
subordo Tubulifera dan Terebrantia 43
4. 10 Filogeni berdasarkan sekuens asam amino DNA mtCOI spesies
xvi
DAFTAR LAMPIRAN
1 Koordinat GPS dan ketinggian lokasi pengambilan sampel 63 2 Sekuens DNA mtCOI sampel trips pada penelitian 64
3 Sekuens asam amino DNA mtCOI trips 67
4 Gejala serangan G. uzeli, H. ganglbaueri, dan P. ichini 68
5 Bunga Thunbergia 69
6 Matrix karakter morfologi delapan spesies trips subordo Tubulifera
dan Terebrantia 70
7 Jarak genetik trips subordo Terebrantia 71
I PENDAHULUAN
Latar Belakang
Trips (Insecta: Thysanoptera) merupakan serangga yang memiliki ukuran tubuh 1-4 mm, namun didaerah tropis dapat mencapai 14 mm. Selain sebagai serangga fitofag, trips juga diketahui berperan sebagai serangga predator. Beberapa spesies dilaporkan sebagai hama yang umum ditemukan pada tanaman pertanian (Reitz et al. 2011). Berdasarkan bentuk ujung abdomen, trips dibagi menjadi dua subordo yaitu Tubulifera dan Terebrantia. Trips yang berhasil diidentifikasi dan diketahui memiliki banyak spesies adalah famili Phlaeothripidae (Tubulifera) dan Thripidae (Terebrantia) (Mound. 2008). Populasi yang tinggi menyebabkan kehilangan hasil pada tanaman. Kehilangan hasil akibat serangan trips di India dan Malaysia dapat mencapai 30-80% (Fauziah dan Saharan; Sastrosiswojo 1991).
Sebagai langkah awal pemecahan permasalahan trips, diperlukan informasi dasar yang lengkap dan akurat terutama mengenai studi taksonominya (Moritz 1994). Beberapa metode dapat digunakan untuk identifikasi trips, mulai dari metode tradisional dengan karakter morfologi hingga metode modern dengan teknik molekuler, morfometrik, atau biokimia (Mehle dan Trdan 2012). Penelitian taksonomi trips menggunakan metode tradisional di Indonesia telah dilakukan oleh Sartiami dan Mound (2013), dimana trips yang banyak ditemukan berasosiasi diberbagai tanaman di Pulau Jawa termasuk kedalam subordo Terebrantia. Selain itu Subagyo (2014) menemukan tujuh belas trips famili Thripidae yang berasosiasi dengan tanaman hortikultura didaerah Cianjur, Lembang, dan Bogor. Di Indonesia lima spesies yang dilaporkan menjadi hama umum ditanaman hortikultura, yaitu: Frankliniella intonsa Trybom, Megalurothrips usitatatus
Bagnall, Thrips parvispinus Karny, Thrips hawaiiensis (Morgan), dan Thrips
palmi Karny. Spesies M. usitatus dan T. hawaiiensis diketahui banyak menyerang
tanaman kacang panjang, buncis, pare, dan kacang bindi (okra). Sementara itu, T.
parvispinus dilaporkan menjadi hama baru yang menyerang pepaya (Fauziah dan
Saharan 1991). Selain menjadi hama pada pertanaman, trips juga menjadi vektor virus penyebab penyakit tanaman. Salah satunya adalah Ceratothripoides
brunneus Bagnall yang banyak ditemukan diberbagai tanaman di Malaysia dan
menjadi vektor Tospovirus (Mound dan Nickle 2009).
Selain anggota dari subordo Terebrantia, beberapa spesies trips anggota Tubulifera juga dilaporkan menjadi hama pada pertanaman. Informasi mengenai keberadaan dan status anggota subordo Tubulifera di Indonesia masih sangat terbatas. Khalsoven (1981) melaporkan trips subordo Tubulifera genus
Haplothrips ditemukan di Indonesia, namun tidak dilengkapi dengan karakter
morfologinya. Spesies dari subordo Tubulifera yang pernah dilaporkan adalah
Gynaikothrips uzeli Zimmermanyang menyerang tanaman beringin (Moraceae) di
Florida. Selain itu, Pseudophilothrips ichini Hood dilaporkan sebagai spesies yang menjadi hama pada tanaman sejenis semak (Anacardiaceae) di Amerika (Held 2005; Mound et al. 2010).
2
struktur tertentu yang sulit dibedakan (Subagyo 2014). Selain itu, variasi karakter morfologi yang terjadi didalam spesies dapat menjadi faktor kesalahan dalam pengidentifikasian. Salah satu contoh variasi morfologi dalam spesies adalah seksual dimorfisme antara jantan dan betina. Perbedaan jenis kelamin pada umumnya akan membentuk perbedaan bentuk tubuh. Beberapa spesies jantan dan betina yang memiliki ukuran yang berbeda akan membentuk allomorfik (pertumbuhan dan perkembangan ukuran dan struktur anggota tubuh misalnya antena dan seta). Selain itu bentuk nimfa instar awal trips yang tidak bersayap akan sulit untuk dibedakan dengan imago trips yang tidak bersayap ataupun warna yang kurang tersklerotisasi (Mound dan Kibby 1998). Identifikasi secara morfologi juga sulit dilakukan pada kondisi sampel yang tidak utuh (Ubaidillah dan Sutrisno 2009). Keterbatasan pada kunci identifikasi morfologi juga menyulitkan proses identifikasi misalnya terbatasnya jumlah karakter dan gambar yang digunakan, serta kunci identifikasi hanya tersedia untuk fase imago (Brunner
et al. 2002; Mound dan Morris 2007).
Identifikasi terhadap trips dapat dilakukan dengan metode modern. Metode identifikasi modern yang umum digunakan adalah dengan teknik molekuler. Teknik ini dapat menunjukkan hasil yang cukup baik untuk identifikasi hingga tingkat spesies (Bayar et al. 2001; Moritz et al. 2001). Kombinasi identifikasi secara morfologi dan molekuler sangat diperlukan dalam identifikasi trips untuk mendapatkan hasil identifikasi yang akurat. Penggunaan penanda genetik juga dapat dijadikan metode alternatif ketika identifikasi morfologi tidak memungkinkan untuk dilakukan (Mehle dan Trdan 2012). Salah satu metode yang digunakan untuk variasi dan keragaman genetik spesies yaitu dengan menggunakan DNA mitokondria (mtDNA) (Chahyadi 2013). DNA mitokondria yang banyak digunakan untuk analisis genetik adalah Cytochrome c Oxidase
subunit I (COI) (Pratami 2013). sekuens DNA mtCOI mampu mengidentifikasi spesies trips dari genus Kladothrips, Oncothrips, Gynaikotrips, Frankliniella dan
Thrips (Crespi et al. 1998; Shao dan Barker 2003; Rugman-Jones et al. 2010;
Mehle dan Trdan 2012). Sekuens DNA mtCOI juga dapat digunakan sebagai karakter tambahan untuk konfirmasi identifikasi spesies dan data dalam penyusunan basis data kodebar DNA (DNA barcode database) (Hebert et al. 2003; Goldstein dan DeSalle 2010).
Penelitian ini merupakan studi lanjutan dari studi sebelumnya mengenai identifikasi trips berdasarkan karakter morfologi pada tanaman hortikultura yang ditemukan didaerah Cianjur, Lembang, dan Bogor oleh Subagyo (2014). Hasil studi tentang karakter molekuler trips ini diharapkan dapat membantu mengkonfirmasi hasil identifikasi secara morfologi dan diharapkan dapat melengkapi data hasil identifikasi spesies-spesies trips yang telah ada.
Perumusan Masalah
anggota subordo Terebrantia maupun Tubulifera, terutama pada spesies yang mudah ditemukan dan banyak dilaporkan menyerang pertanaman seperti:
Ceratothripoides brunneus, Megalurothrips usitatus, Thrips alliorum, T.
hawaiiensis, dan T. parvispinus maupun pada trips pada tanaman beringin, pala,
dan seruni laut.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari karakter molekuler spesies trips subordo Terebrantia menggunakan sekuens DNA mtCOI dan mengidentifikasi spesies trips subordo Tubulifera yang ditemukan pada tanaman beringin (Ficus
benjamina/ Moraceae), pala (Myristica fragrans/ Myristicaceae), dan seruni laut
(Wedelia biflora/ Asteraceae).
Manfaat Penelitian
II TINJAUAN PUSTAKA
Trips (Insecta: Thysanoptera)
Trips (Thysanoptera) adalah serangga yang memiliki ukuran tubuh sekitar 1-4mm (daerah tropika panjangnya hampir 14mm). Beberapa spesies ada yang tidak memiliki atau tanpa sayap. Tipe alat mulut meraut – menghisap, terdiri atas probosis dengan struktur tidak simetris, besar, konis, dan terletak bagian posterior pada ventral kepala. Selain itu terdapat stilet, satu mendibel dan lasiniae dari dua maksila. Antena yang dimiliki trips biasanya pendek, terdiri dari empat sampai sembilan ruas. Tipe metamorfosis peralihan antara sederhana dan sempurna. Morfologi antara serangga jantan dan betina hampir sama, tetapi biasanya serangga jantan memiliki ukuran tubuh yang lebih kecil dibanding dengan serangga betina. Serangga ini berperan sebagai pemakan tanaman, menyerang bunga dan daun, memakan spora-spora jamur, pemangsa arthropoda kecil dan vektor penyakit tumbuhan (Borror et al. 1989; Ritz et al. 2010).
Salah satu famili trips yang menjadi hama adalah Thripidae. Ciri khasnya adalah jumlah ruas antena 7-8 segmen dan memiliki struktur seperti garpu atau sederhana (Sartiami dan Mound 2013). Famili ini terdiri atas 3 subfamili yaitu Panchaetothripidae, Dendothripinae, dan Thripinae. Subfamili Thripinae memiliki sekitar 1 600 spesies dari 250 genus yang banyak berasosiasi dengan tanaman dan sebagian besar merupakan hama penting dan menjadi vektor virus pada tanaman. Beberapa spesies trips anggota subfamili Thripinae yang menjadi hama penting baik pada sayuran maupun bunga hias diantaranya Frankliniella intonsa, F.
occidentalis, Megalurothrips sp, T. aspinus, T. hawaiiensis, T. parvispinus, Thrips
tabaci Lindeman, Scirtothrips dorsalis Hood, Haplothrips floricola Priesner, dan
Thrips flavus Schrank (Bansidhi dan Poonchaisri 1991; Fung et al. 2002; Kirk
2002; Sartiami et al. 2011; Subagyo 2014). Spesies lain yang dilaporkan menjadi vektor virus diantaranya T. tabaci dan Frankliniela schultzei (Trybom) (Kirsten
et al. 2009; Westmore 2012).
Kerusakan yang ditimbulkan oleh trips menimbulkan kerugian secara langsung ataupun tidak langsung. F. intonsa dilaporkan merupakan hama penting pada tanaman krisan, dimana terdapat bercak kecoklatan pada bunga (Yusuf et al.
2010). Spesies ini juga menyerang tanaman lili, jeruk, kacang-kacangan, selada, bawang, dan jagung (Subagyo 2014). Spesies lain yang merugikan tanaman yaitu:
T. palmi, T. tabaci, dan M. usitatus. Spesies-spesies tersebut merupakan spesies
yang banyak merusak tanaman hortikultura di negara Philipina terutama pada tanaman terung, tomat dan kentang (Bernardo 1991). Selain kerugian langsung, terdapat kerugian tidak langsung yang ditimbulkan oleh trips, misalnya T.
parvispinus yang menyerang daun pepaya menyebabkan infeksi cendawan
Cladosporium oxysporum sehingga menyebabkan malformasi, bercak, dan lubang
pada daun. Selain itu T. tabaci juga dilaporkan dapat menjadi vektor beberapa virus tanaman, seperti: Cucumber Mosaic Virus, Potato Leaf Roll Virus, Tobacco
Mosaic Virus, Potato Virus, dan Pea Mosaic Virus (Sakimura 1946).
dari subordo Tubulifera pembentuk puru pada tanaman, misalnya Arrhenothrips
ramakrishnae Hood, Teucotothrips longus (Schmutz), Gynripsikothrips
flaviantennus Multon, Schedothrips orientalis Ananthakrishnan, Crotonothrips
dantahasta (Ramakrishna), Thilakothrips babuli Ramakrishna, dan Androthrips
flavipes Schmutz (Varadarasan dan Ananthakrishnan 1982). Anggota Subordo
Tubulifera yang lain yang pernah dilaporkan adalah Gynaikothrips uzeli yang menyerang tanaman beringin, G. ficorum yang menyerang tanaman salam (Myrtaceae), dan Haplothrips ganglbaueri menyerang gulma Echinochloa
crusgalli (Poaceae) (Ananthakrishnan dan Thangavilu 1976), sedangkan
Pseudophilothrips ichini dilaporkan menyerang tanaman lada (Held et al. 2005;
Mound et al. 2010).
Identifikasi Trips berdasarkan Karakter Morfologi
Karakter yang Digunakan dalam Identifikasi Morfologi Trips
Identifikasi yang dilakukan dengan menggunakan karakter morfologi disebut juga identifikasi tradisional. Antena dan oseli merupakan karakter yang umum dilihat di bagian kepala trips (Mound dan Kibby 1998; Mound 2006). Bagian pada toraks yang dapat dijadikan karakter untuk identifikasi adalah pronotum, mesonotum, metanotum, tungkai, dan sayap depan. Rangkaian seta pada pronotum sering digunakan untuk menentukan spesies dari suatu genus, sedangkan pola retikulasi sklerit pada metanotum dapat digunakan untuk membedakan spesies tertentu, karena pola ini bervariasi dan cenderung khas. Sayap pada bagian toraks merupakan karakter yang paling sering digunakan untuk identifikasi dan karakter yang sering dilihat adalah jumlah seta pada venasi sayap depan serta ukuran seta terminal dan seta sub-terminal pada clavus (Mound dan Kibby 1998).
Karakter pada abdomen yang umumnya dilihat adalah pola retikulasi pada abdomen dan keberadaan microtrichia pada bagian tergit abdomen, selain itu keberadaan stenidia pada tergit abdomen ruas VIII dan microtrichia atau comb pada garis bagian belakang tergit abdomen ruas VIII dapat digunakan untuk membedakan spesies satu dengan yang lainnya secara spesifik. Karakter lainnya adalah setiap sternit abdomen mempunyai serangkaian seta pada tepi bawah, umumnya berjumlah tiga pasang pada beberapa spesies, tetapi pada genus Thrips
sternit juga mempunyai serangkaian seta diskal (Mound 2006).
Kelebihan dan Kekurangan Identifikasi Morfologi.
Identifikasi secara morfologi memiliki beberapa keuntungan diantaranya lebih cepat, mudah dan murah jika dibandingkan dengan metode molekular (Hoy 2003). Namun identifikasi trips secara morfologi memiliki keterbatasan diantaranya:
6
2. Bentuk nimfa instar awal trips yang tidak bersayap akan sulit untuk dibedakan dengan imago trips yang tidak bersayap ataupun warna yang kurang tersklerotisasi
3. Ukuran trips yang sangat kecil sehingga karakter kunci sulit untuk dilihat dengan jelas.
4. Identifikasi secara morfologi sulit dilakukan pada kondisi sampel yang tidak utuh.
5. Pada beberapa kasus, beberapa spesies secara morfologi tampak serupa atau hanya berbeda pada detail struktur tertentu yang sulit dibedakan
Identifikasi Menggunakan Karakter Molekular
Mitokondria Cytochrome Oxidase I
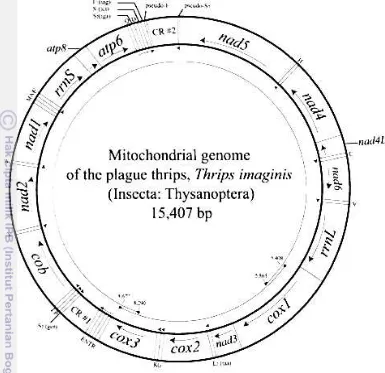
Gen penyusun DNA mitokondria pada hewan terdiri atas 37 gen untuk mengkode pembentukan sub unit rRNA kecil hingga besar, 13 protein, dan 22 tRNA sehingga menjadi tempat yang konservatif dalam suatu filum. Hal ini juga berlaku dalam dunia serangga, sehingga taksonomi dengan metode molekular dapat dilakukan berdasarkan sistematika pendekatan filogenetik (Hoy 2003). Analisis mitokondria pada trips mencatat pada T. imaginis dari 15 407 panjang total DNA mitokondria terdiri dari 4 daerah yang memiliki fungsi yang berbeda yaitu: (1) gen pengkode protein, (2) gen tRNA dan gen Pseudo-tRNA, (3) gen rRNA, dan (4) bagian yang tidak mengkode apapun. Posisi DNA mtCOI berada pada daerah yang mengkode protein, dimana daerah ini kaya akan kandungan A dan T (Shao dan Baker 2003).
Menurut Ubaidillah dan Sutrisno (2009) DNA mtCOI dipilih menjadi gen yang digunakan untuk barkoding, karena memiliki sifat-sifat yang memenuhi persyaratan untuk digunakan dalam menentukan identitas sebuah spesies. Gen mtCOI memiliki ukuran yang relatif pendek dan stabil (tidak mudah mengalami perubahan bila dibandingkan dengan gen mitokondria sejenis). Selain itu gen mtCOI sangat mudah untuk dilakukan pengurutannya dibandingkan dengan gen-gen yang berasal dari inti.
Kelebihan dan Kekurangan Identifikasi Molekuler
Pada trips bagian yang diamplifikasi pada proses PCR (Polymerase Chain
Reaction) adalah DNA mitokondria. Penggunaan DNA mtCOI sebagai penanda
berpotensi untuk diuji secara teoritis hanya dengan menggunakan jumlah nukleotida yang terbatas.
Gambar 2.1 Posisi DNA mtCOI T. imaginis (Shao dan Barker 2003)
III KARAKTER MOLEKULER TRIPS SUBORDO
TEREBRANTIA
ABSTRAK
Trips merupakan serangga yang sebagian besar anggotanya berperan sebagai hama maupun vektor penyakit tanaman hortikultura terutama sayuran. Kerusakan yang ditimbulkan oleh trips dapat menyebabkan 30-50% kehilangan hasil. Genus Thrips Linnaeus adalah genus kedua terbesar dari ordo Thysanoptera dan banyak dilaporkan menjadi hama pada banyak tanaman inang. Selain anggota dari genus Thrips, Ceratothripoides dan Megalurothrips dilaporkan sebagai hama penting pada tanaman tomat dan tanaman kacang-kacangan di Asia dan Afrika. C.
brunneus dan M. usitatus juga dilaporkan sebagai vektor Tomatto Spot Wilt Virus
(TSWV) atau penyakit layu pada tomat.
Tujuan dari penelitian ini adalah mempelajari sekuens DNA mtCOI sebagai karakter molekuler spesies trips dari subordo Terebrantia yaitu: genus
Ceratothripoides (C. brunneus), Megalurothrips (M. usitatus), dan Thrips (T.
alliorum, T. hawaiiensis dan T. parvispinus). Pengambilan sampel lakukan di
Kabupaten Bandung, Bogor, Cianjur, Cirebon, dan Kuningan. Pengambilan sampel dilakukan secara langsung pada bunga atau daun tanaman yang mengalami gejala akibat serangan trips. Identifikasi secara morfologi terdiri dari pembuatan preparat slide dan identifikasi menggunakan kunci identifikasi morfologi. Identifikasi menggunakan karakter molekuler terdiri atas empat tahapan yaitu ekstraksi DNA total, amplifikasi menggunakan PCR, sekuensing, dan analisis .
Program PCR berhasil mengamplifikasi DNA mtCOI C. brunneus, M.
usitatus, T. alliorum, T. hawaiiensis, dan T. parvispinus pada ±710 pb. Data
sekuens DNA mtCOI kelima spesies tersebut berturut-turut adalah 693, 692, 678, 690, dan 668 pb. Nilai jarak genetik nukleotida antar spesies sekitar 0.00-0.401, dan jarak genetik asam amino antar spesies sebesar 0.00-0.268. Sekuens DNA mtCOI kelima spesies dari subordo Terebrantia ini mengkonfirmasi hasil identifikasi morfologi.
Kata Kunci: COI, hama, hortikultura, mitokondria, sekuens
ABSTRACT
Thrips are insect that most of species are pests and vectors of diseases on horticultural crops, especially on vegetable plants. Thrips can caused 30-50% yield loss. Thrips Linnaeus is the second largest genus of the order Thysanoptera and most of them are pests on many host plant. Not only this genus,
Ceratothripoides and Megalurothrips also reported as important pest on tomatto
and nuts in Asia and Africa. Ceratothripoides brunneus and Megalurothrips
usitatus also been reported as Tomatto Spot Wilt Virus (TSWV) vectors.
Megalurothrips (M. usitatus), and Thrips (T. alliorum, T. hawaiiensis dan T.
parvispinus). Samples were collected from symptomatic thrips attack plants in
districts of Bandung, Bogor, Cianjur, and Cirebon. Morphological identification process consists of preparation of preparat slide and observation by using morphological identification keys. Molecular identification process consists of : DNA extraction using CTAB methods, DNA amplification, DNA sequencing, and analysis. There were five species of thrips suborder Terebrantia frequently found in cultivated crops they were C. brunneus, M. usitatus, T. alliorum, T.
hawaiiensis, and T. parvispinus. The length of their mtCOI DNA sequences were:
693, 692,678, 690, and 668 bp respectively. The range of genetic distance DNA sequences was 0.00 to 0.401, while the range of genetic distance of amino acid sequences was 0.00 to 0.268. MtCOI DNA sequences data confirmed morphological results data from five species Terebrantia suborder.
Keywords: COI, horticulture, mitochondrial, pest, sequence
PENDAHULUAN
Trips merupakan serangga yang sebagian besar anggotanya berperan sebagai hama maupun vektor penyakit tanaman hortikultura terutama tanaman sayuran (Oktaviany et al. 2013). Peranan trips sebagai hama pada tanaman disebabkan oleh aktivitas makan yang dilakukan. Kerusakan yang ditimbulkan berupa bintik putih pada bunga atau daun, sehingga mengganggu proses fotosintesis pada tanaman (Subagyo 2014). Indonesia belum memiliki data mengenai kerugian yang ditimbulkan oleh trips, namun di India serangan trips dapat menyebabkan kehilangan hasil sebesar 30-50% pada tanaman cabai, sedangkan di Malaysia dapat mencapai 80% pada tanaman krisan (Fauziah dan Saharan 1991; Subagyo 2014; Sastrosiswojo 1991).
Genus Thrips Linnaeus adalah genus kedua terbesar dari ordo Thysanoptera yang terdiri dari 286 spesies (Mound 2012). Sebagian besar anggota dari genus ini hidup pada bunga dan sebagian pada bagian daun (Mound dan Marullo 1996). Beberapa anggota dari genus ini merupakan hama penting pada tanaman di dunia seperti Thrips angusticeps Uzel, Thrips flavus Schrank, Thrips hawaiiensis
(Morgan), Thrips meridionalis Priesner, dan Thrips tabaci Lindeman (Moritz 1994). T. hawaiiensis dilaporkan banyak menyerang tanaman pare dan okra, sedangkan T. parvispinus merupakan hama baru yang menyerang pepaya di Malaysia (Fauziah dan Saharan 1991). Di Indonesia T. parvispinus, T.
hawaiiensis, dan T. palmi tercatat sebagai hama umum yang ditemukan pada
pertanaman hortikultura (Subagyo 2014). Genus Thrips yang tersebar luas dan memiliki spesies yang banyak menyebabkan tersedianya beberapa kunci identifikasi di beberapa negara seperti Malaysia pada tanaman sayuran (Mound dan Azidah 2009), Indonesia pada tanaman hortikultura (Sartiami dan Mound 2013; Subagyo 2014), serta Iran pada tanaman hortikultura dan palawija (Mirab-balou et al. 2012).
10
Mound 2013). Trips memberikan dampak langsung terhadap kerusakan tanaman pada bagian daun dan bunga, trips juga menjadi vektor virus penting pada tanaman. T. hawaiiensis juga dilaporkan menjadi hama penting di Taiwan dan Malaysia. Spesies ini merupakan hama umum yang ditemukan di Taiwan yang menyerang bunga tanaman pertanian dan bunga potong. T. hawaiiensis
menyebabkan permukaan buah menjadi kasar, pucat, dan menurunkan kualitas buah. Terdapat 21 tanaman yang diserang diantaranya: pisang, jeruk, anggur, mangga, jambu air, krisan, gladiol, mawar, cabai, teh dan tanaman kacang-kacangan (Chang 1991). Spesies T. parvispinus merupakan hama serius pada cabai. Kerusakan yang ditimbulkan terlihat pada daun/ tanaman muda dan akan terus menyebar selama perkembangan tanaman. Kehilangan hasil yang diakibatkan oleh serangan T. parvispinus dapat mencapai 22.8% pada tanaman cabai (Sastrosiswojo 1991). T. parvispinus juga menjadi hama pada tanaman tembakau di daerah Jawa Tengah dan Jawa Barat (Vos et al. 1991). Selain itu, T.
parvispinus juga dilaporkan menyerang tanaman bunga krisan. Serangan trips
dapat mencapai 11-52% tergantung varietas bunga, hal ini menurunkan kualitas bunga karena timbul bekas rautan dan bercak kehitaman pada mahkota bunga menjadi masalah yang cukup serius pada perdagangan internasional (Mehle dan Trdan 2012). Identifikasi yang cepat, tepat, dengan deskripsi yang jelas mengenai karakter morfologi terhadap trips sangat diperlukan untuk pengendalian hama trips. Identifikasi yang umum dilakukan di Indonesia adalah identifikasi menggunakan kunci identifikasi morfologi. Namun, terdapat beberapa kekurangan dalam penggunaan kunci identifikasi morfologi. Salah satunya adalah identifikasi morfologi dan tidak mudah untuk dilakukan terutama untuk genus dengan jumlah spesies yang banyak seperti dalam anggota genus Thrips (Hasmiwati et al. 2006). Kesulitan yang lain dalam pengidentifikasian terhadap trips adalah ukuran yang kecil, karakter-karakter yang sulit terlihat, serta variasi seksual dimorfisme dalam spesies. Seksual dimorfisme dapat menyebabkan kesalahan identifikasi (Mound dan Kibby 1998). Berdasarkan hal tersebut, perlu adanya karakter tambahan dalam proses identifikasi terhadap trips.
Karakter tambahan yang biasa digunakan dalam proses identifikasi trips adalah sekuens DNA mitokondria (Hoy 1994). Gen penyusun DNA mitokondria yang banyak digunakan untuk melihat variasi genetik spesies adalah
Mitochondria Cytochrome c Oxidase I (mtCOI). Gen ini digunakan karena
memiliki laju evolusi yang tinggi sehingga akan berbeda pada setiap spesies. DNA mtCOI juga penyandi protein yang memiliki variasi genetik yang tinggi (Ubaidillah dan Sutrisno 2009). Selain itu DNA mtCOI juga digunakan sebagai gen standar penanda molekuler (DNA barcode) untuk melihat karakter dan variasi genetik intraspesies dan interspesies (Hebert et al. 2003).
Penelitian ini bertujuan untuk mempelajari karakter molekuler berupa sekuens DNA mtCOI trips subordo Terebrantia yaitu: C. brunneus, M. usitatus, T.
alliorum, T. hawaiiensis, dan T. parvispinus, hal ini dikarenakan spesies-spesies
spesies tersebut. Manfaat penelitian ini berupa data sekuens DNA mtCOI yang digunakan sebagai tambahan karakter untuk melengkapi data morfologi dalam proses pengidentifikasian trips. Sekuens DNA mtCOI yang ada, diharapkan dapat memperkecil kesalahan identifikasi menggunakan karakter morfologi, sehingga hasil identifikasi menggunakan kedua metode ini menjadi lebih lengkap dan akurat.
METODE PENELITIAN
Sampel trips diambil dari Kabupaten Bandung, Bogor, Cianjur, Cirebon, dan Kuningan. Trips kemudian diidentifikasi berdasarkan karakter morfologi di Laboratorium Biosistematika Serangga. Ekstraksi dan amplifikasi DNA mtCOI trips di Laboratorium Virologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, IPB. Penelitian ini dilaksanakan pada bulan September 2014 sampai Agustus 2015.
Sampel trips diambil secara langsung pada bagian bunga dan daun yang bergejala akibat serangan trips. Sampel kemudian dimasukkan ke dalam tabung eppendorf berisi alkohol absolut, selanjutnya dimasukkan kedalam plastik klip yang diberi keterangan. Lokasi pengambilan sampel ditandai menggunakan GPS untuk mendapatkan koordinat. Trips yang sudah dikoleksi dari lapangan kemudian disortir. Sebagian disimpan untuk ekstraksi DNA trips sementara yang lain dibuat slide dengan metode Mound dan Kibby (1998). Identifikasi trips dilakukan di bawah mikroskop stereo OLYMPUS CX21FSI yang dilengkapi langsung dengan kamera Dino-eye. Identifikasi dilakukan menggunakan buku identifikasi Mound dan Kibby (1998), Sartiami dan Mound (2013) serta Subagyo (2014).
Ekstraksi DNA dan Amplifikasi dengan PCR.
Imago trips yang telah disimpan ke dalam alkohol absolut, kemudian diekstraksi dengan teknik molekuler untuk mendapatkan DNA total yang mengacu pada metode Goodwin et al. (1994) yang telah dimodifikasi. Individu imago trips dari masing-masing spesies dimasukkan ke dalam tabung eppendorf 1.5 mL, kemudian ditambahkan 100 μL bufer ekstraksi CTAB 2%. Selanjutnya ditambahkan 1 μL proteinase K, kemudian dihancurkan sampai halus menggunakan micropestle. Suspensi diinkubasi pada suhu 65 ⁰ C selama 3 menit. Larutan Chlorofoam: Isoamil alcohol (CI) (24:1) sebanyak 100 μL ditambahkan ke dalam suspensi dan vortek selama 3 menit. Suspensi tersebut kemudian disentrifugasi selama 15 menit dengan kecepatan 10 000 rpm sehingga dihasilkan supernatan. Supernatan yang telah diperoleh dipindah ke tabung eppendorf 1.5 mL yang baru sebanyak 60 μL. Larutan Isopropanol sebanyak 44 μL dan Sodium asetat 3 M (pH 5.2) sebanyak 6 μL ditambahkan ke dalam supernatan untuk proses presipitasi (pengendapan DNA) dan diinkubasi di lemari pendingin pada suhu -20 ⁰ C selama 3 jam atau semalam (overnight).
12
pelet DNA yang sudah bersih. Pelet DNA total trips disuspensikan kembali dengan larutan Tris-EDTA (TE) sebanyak 20 μL.
Amplifikasi dan Sekuensing DNA mtCOI Trips
Proses amplifikasi menggunakan mesin PCR Perkin Elmer 480 Thermocycler (Applied Biosystem, US). Amplifikasi fragmen DNA mtCOI trips menggunakan primer forward dan reverse dengan panjang amplikon sebesar ±710 pb. Primer yang digunakan untuk mengamplifikasi DNA mtCOI dari trips sepasang primer universal DNA mtCOI LCO 1490 (3'-GGTCAACAAATCATAA AGATATTGG-5') dan HCO 2198 (5'TAAACTTCAGGGTGACCAAAAAATCA -3') (Folmer et al. 1994). Total volume reaksi PCR yang digunakan adalah 25 μl yang terdiri atas 9.5 μl air destilata steril, 1 μl primer forward, 1 μl primer reverse, 1 μl DNA cetakan, dan 12.5 μl PCR Master Mix.
Program amplifikasi yang digunakan ialah: denaturasi inisiasi selama 5 menit pada 94 oC, kemudian dilanjutkan 35 siklus yang terdiri atas: denaturasi selama 1 menit pada 94 oC, penempelan primer selama 35 detik pada 52 oC, pemanjangan 72 oC selama 1 menit 30 detik, dan pemanjangan akhir 7 menit pada suhu 72 oC. Hasil PCR kemudian dielektroforesis menggunakan agarose 1% pada tegangan 50 volt selama 50 menit dan divisualisasi menggunakan UV transilluminator. Pita-pita yang tervisualisasi kemudian dianalisis ukuran fragmen DNA yang dibandingkan dengan marker 1 kb (Thermo Scientific, US)
Analisis Jarak Genetik dan Filogeni DNA mtCOI Trips
Identifikasi spesies lebih lanjut dilakukan analisis homologi basa nukleotida. Produk PCR disekuensing (proses ini dilakukan oleh perusahaan sekuensing). Selanjutnya sekuens DNA mtCOI trips diolah menggunakan perangkat lunak BioEdit 7.0.9 untuk dibandingkan dengan sekuen database dari GeneBank (www.ncbi.nlm.nih.gov). Analisis homologi dilakukan pada sekuen DNA mtCOI trips dengan data GeneBank. Program Basic Local Alignment Search
Tool-Nucleotide (BLAST-N) (www.ncbi.nlm.nih.gov /blast/) digunakan untuk analisis
homologi. Sekuen nukleotida DNA mtCOI trips di-alignment dengan data sekuen nukleotida DNA mtCOI trips yang diperoleh dari GeneBank. Data sekuen nukleotida untuk DNA mtCOI trips yang diperoleh dari GeneBank dibentuk matriks nukleotida dilakukan dengan bantuan perangkat lunak Clustal x (1.83). Konstruksi pohon filogeni dilakukan antar DNA mtCOI trips. Proses analisis filogeni tersebut menggunakan metode Unweighted Pair Group Method using
Arithmetic (UPGMA) dengan bootstrap 1000 kali pada program Molecular
Evolutionary Genetic Analisis (MEGA) 6.
HASIL
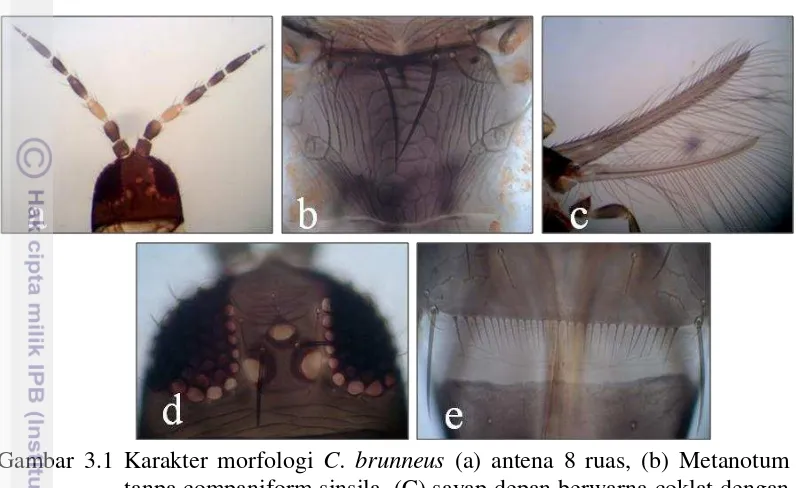
Karakter Morfologi Ceratothripoides brunneus dan Megalurothrips usitatus Berdasarkan hasil identifikasi menggunakan kunci identifikasi morfologi Subagyo (2014) dan Sartiami dan Mound (2013) spesies yang ditemukan merupakan Ceratothripoides brunneus Bagnall dan Megalurothrips usitatus
metanotum tanpa companiform sinsila dan memiliki sepasang seta tepat pada garis atas metanotum (Gambar 3.1 b), (3) sayap berwarna coklat dengan venasi yang lengkap (Gambar 3.1 c), (4) terdapat 3 pasang seta oseli dan seta oseli I berbaris secara vertikal (Gambar 3.1 d), (5) memiliki deretan microtrichia yang lengkap pada tergit abdomen ruas VII (Gambar 3.1 e).
Gambar 3.1 Karakter morfologi C. brunneus (a) antena 8 ruas, (b) Metanotum tanpa companiform sinsila, (C) sayap depan berwarna coklat dengan venasi lengkap, (d) Seta oseli I berbaris secara vertikal, (e) deretan microtrichia yang lengkap pada tergit abdomen ruas
Tabel 3.1 Tanaman inang dan lokasi ditemukannya C. brunneus dan M. usitatus
Spesies Tanaman inang Lokasi
Desa/ Kecamatan/ Kabupaten
C. brunneus Thunbergia sp* Cibodas/ Cibodas/ Cianjur
Mawar Taman Bunga Nusantara/ Cipanas/ Cianjur
M. usitatus Kacang panjang Sunyaragi/ Kesambi/ Cirebon
Buncis * Cikareumbi/Lembang/ Bandung Sukajadi/ Tamansari/ Bogor Cikarawang/ Dramaga/ Bogor Ciareteun/ Leuwiliang/ Bogor Teh Sukajadi/ Tamansari/ Bogor Bawang daun Cipanas/ Cipanas/ Cianjur
Keterangan: (*) menunjukkan sampel yang dilakukan identifikasi secara molekuler
14
(Gambar 3.2 d), (5) terdapat microtrichia pada tergit abdomen betina ruas VIII tetapi hanya sebagian (Gambar 3.2 e), dan (6) seta S1 berada diatas garis abdomen ruas ke VII (Gambar 3.2 f).
Gambar 3.2 Karakter morfologi M. usitatus (a) antena 8 ruas, (b) Metanotum tanpa companiform sinsila, (C) sayap depan berwarna gelap terang, (d) seta oseli III muncul pada garis segitiga oseli, (e) deretan microtrichia yang VIII hanya sebagian, (f) seta S1 diatas garis abdomen ruas ke VII
Analisis Karakter Molekuler C. brunneus dan M. usitatus
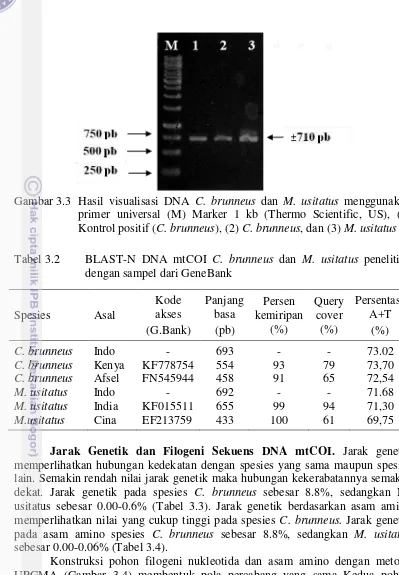
Primer forward dan reverse mampu mengamplifikasi DNA mtCOI dengan baik, terlihat pita DNA 1 sampai 3 merupakan hasil amplifikasi DNA mtCOI trips. Pita DNA ke-2 berasal dari DNA total C. brunneus yang ditemukan pada tanaman Thunbergia sp (Cibodas) (Lampiran 5), pita DNA ke-3 berasal dari DNA total DNA M. usitatus pada inang buncis (Bandung). Pita yang terlihat sesuai dengan target yang diinginkan yaitu berukuran sekitar ±710 pb (Gambar 3.3).
Analisis homologi sekuens DNA mtCOI. Sekuens DNA mtCOI C.
brunneus dan M. usitatus mempunyai panjang basa nukleotida berturut-turut
sebesar 693 dan 692 pb. Berdasarkan hasil BLAST susunan nukleotida sampel C.
brunneus yang ditemukan di Indonesia memiliki persentase kemiripan 93%
dengan spesies yang sama yang berasal dari Kenya dan 91% dengan spesies yang berasal dari Afrika Selatan. M. usitatus menunjukkan persentase kemiripan sebesar 99% dengan spesies yang sama yang berasal dari India dan 100% dengan spesies yang berasal dari Cina (Tabel 3.2). Trips yang telah diidentifikasi secara morfologi dan berdasarkan hasil BLAST-N merujuk pada spesies yang sama yaitu
Gambar 3.3 Hasil visualisasi DNA C. brunneus dan M. usitatus menggunakan memperlihatkan hubungan kedekatan dengan spesies yang sama maupun spesies lain. Semakin rendah nilai jarak genetik maka hubungan kekerabatannya semakin dekat. Jarak genetik pada spesies C. brunneus sebesar 8.8%, sedangkan M. usitatus sebesar 0.00-0.6% (Tabel 3.3). Jarak genetik berdasarkan asam amino memperlihatkan nilai yang cukup tinggi pada spesies C. brunneus. Jarak genetik pada asam amino spesies C. brunneus sebesar 8.8%, sedangkan M. usitatus
sebesar 0.00-0.06% (Tabel 3.4).
16
Tabel 3.3 Jarak genetik sekuens nukleotida DNA mtCOI C. brunneus dan M.
usitatus pada penelitian dengan sampel dari GeneBank
Keterangan: CbI= C. brunneus Indonesia, CbA= C. brunneus Afrika Selatan, CbK= C. brunneus
Kenya, MuI= M. usitatus Indonesia, MuD= M. usitatus India, MuC= M. usitatus Cina, Gu= Gynaikothrips uzeli
Tabel 3.4 Jarak genetik asam amino DNA mtCOI C. brunneus dan M. usitatus
penelitian dengan sampel dari GeneBank
Keterangan: CbI= C. brunneus Indonesia, CbA= C. brunneus Afrika Selatan, CbK= C. brunneus
Kenya, MuI= M. usitatus Indonesia, MuD= M. usitatus India, MuC= M. usitatus Cina, Gu= Gynaikothrips uzeli
(a)
(b)
Gambar 3.4 Filogeni DNA mtCOI C. brunneus dan M. usitatus menggunakan software Mega dengan pendekatanUPGMA bootstrap 1000x (a) berdasarkan sekuens nukleotida, (b) sekuens asam amino
C.brunneus_Kny_KF778754
Spesies CbI CbA CbK MuI MuD MuC Gu
CbI ID
CbA 0.092 ID
CbK 0.092 0.000 ID
MuI 0.319 0.321 0.321 ID
MuD 0.324 0.326 0.326 0.006 ID
MuC 0.319 0.321 0.321 0.000 0.006 ID
Gu 0.586 0.628 0.628 0.615 0.627 0.615 ID
Spesies CbI CbA CbK MuI MuD MuC Gu
CbI ID
CbA 0.088 ID
CbK 0.088 0.000 ID
MuI 0.258 0.258 0.358 ID
MuD 0.262 0.262 0.262 0.006 ID
MuC 0.258 0.258 0.258 0.000 0.006 ID
18
Tabel 3.6 Posisi sekuens asam amino DNA mtCOI C. brunneus dan M. usitatus dengan database GeneBank yang menunjukkan variasi
Spesies
Asam amino ke-
1 2 5 8 9
22 17 18 20 21 23 29 46 47 48 51 65 68 72 75 77 78 79 82 84 85 88 89 96 112 115 116
C.brunneus I Y I I R L I I I R S T I R Y I L T F I L P K G Q V P N A I V
C.brunneus_Kny_KF778754 . . M M S . M M . S . . . S . M I . . M . . N . . . H . . M .
C.brunneus_Afsel_FN545944 - - . . . I . . . N . . . H . . . .
M.usitatus A F L . . I T L L G A G T S L L . V L L K T . R S T T L G M L
M.usitatus_Chn_EF213759 - - L M S I T L L G A G T S L L . V L L K T . S S T T L G M L
Karakter Morfologi beberapa Spesies Genus Thrips
Berdasarkan hasil identifikasi menggunakan kunci identifikasi morfologi Subagyo (2014), dan Sartiami dan Mound (2013) terdapat 3 spesies yang termasuk dalam subordo Terebrantia genus Thrips. Ketiga spesies tersebut diantaranya: Thrips alliorum (Priesner), Thrips hawaiiensis (Morgan), dan Thrips
parvispinus (Karny). Trips tersebut temukan di Kabupaten Bandung, Bogor,
Cianjur, dan Kuningan pada inang yang berbeda (Tabel 3.7).
Tabel 3.7 Inang dan lokasi ditemukannya T. alliorum, T. hawaiiensis, dan
T. parvispinus
Keterangan : (*) menunjukkan sampel yang dilakukan identifikasi secara molekuler (**) TBN = Taman Bunga Nusantara
Karakter morfologi dari T. alliorum: antena terdiri dari 7 ruas (Gambar 3.5 a), metanotum tanpa companiform sensila (Gambar 3.6 a), dan venasi sayap depan tidak lengkap (Gambar 3.7 a). T. hawaiiensis antena terdiri dari 7 ruas (Gambar venasi pertama dan kedua sayap depan lengkap (Gambar 3.7 c).
Spesies Tanaman inang Lokasi
Desa/ Kecamatan/ Kabupaten
Thrips alliorum Bawang daun* Cipanas/ Cipanas/ Cianjur
T. hawaiiensis Pepaya * Cisantana/ Cigugur/ Kuningan
Cikarawang/ Dramaga/ Bogor Bengkoang Cilimus/ Cilimus/ Kuningan Mawar Balithi/ Cipanas/ Cianjur
Sukajadi/ Tamansari/ Bogor
T. parvispinus Cabai Cisantana/ Cigugur/ Kuningan
Tomat Cisantana/ Cigugur/ Kuningan Terung Cisantana/ Cigugur/ Kuningan Buncis Cisantana/ Cigugur/ Kuningan
Ciputri/ Lembang/ Bandung barat Sukajadi/ Tamansari/ Bogor Ciareteun/ Leuwiliang/ Bogor Kacang panjang Sukajadi/ Tamansari/ Bogor Mentimun* Ciputri/ Lembang/ Bandung
Marga mekar/ Pangalengan/ Bandung Wortel Cipanas/ Cipanas/ Cianjur
Kailan Cipanas/ Cipanas/ Cianjur Terung pipit Sukajadi/ Tamansari/ Bogor Mawar TBN**/ Cipanas/ Cianjur
20
Gambar 3.5 Kepala dan antena (a) T. alliorum, (b) T. hawaiiensis, (c) T. parvispinus
Gambar 3.6 Metanotum (a) T. alliorum, (b) T. hawaiiensis, (c) T. parvispinus
Gambar 3.7 Sayap (a) T. alliorum, (b) T. hawaiiensis, (c) T. parvispinus
Analisis Kekerabatan Spesies Genus Thrips dengan Teknik Molekuler
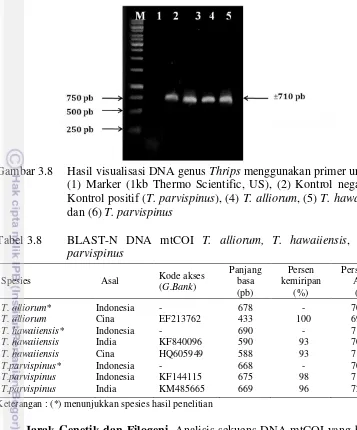
Primer forward dan reverse mampu mengamplifikasi DNA mtCOI dengan baik, terlihat pita DNA 2 sampai 5 merupakan hasil amplifikasi DNA mtCOI dari trips. Pita DNA 1 merupakan kontrol negatif yang tidak diberikan DNA cetakan, pita 2 berasal dari DNA total T. parvispinus yang telah diekstraksi dan diamplifikasi sebelumnya. Pita 3-5 merupakan hasil amplifikasi DNA mtCOI dari
T. alliorum, T. hawaiiensis dan T. parvispinus. Pita yang terlihat sesuai dengan
target yang diinginkan yaitu berukuran sekitar 710 pb (Gambar 3.8).
Analisis Homologi Sekuens DNA mtCOI. Trips yang telah diidentifikasi secara morfologi kemudian dianaliis sekuensnya. Hasil BLAST-N merujuk pada spesies yang sama yaitu T. alliorum, T. hawaiiensis,dan T. parvispinus. Ketiga sekuens DNA mtCOI trips tersebut mempunyai nilai homologi dengan trips dari GeneBank 93-100% (Tabel 3.8).
Gambar 3.8 Hasil visualisasi DNA genus Thrips menggunakan primer universal
T.parvispinus India KM485665 669 96 72.20
Keterangan : (*) menunjukkan spesies hasil penelitian
Jarak Genetik dan Filogeni. Analisis sekuens DNA mtCOI yang berhasil disejajarkan terlihat adanya perbedaan dibeberapa titik pada urutan basa nukleotida pada masing-masing spesies. Hal tersebut merupakan variasi susunan pada masing-masing sampel. Jarak genetik interspesies T. alliorum sebesar 0.0%,
T. hawaiiensis sebesar 6.5-7.1%, dan T. parvispinus sebesar 2.4-3.8% (Tabel 3.9).
Ketiga sepesies memiliki 397 daerah conserve dengan 100 nukleotida yang bervariasi sehingga variasi nukleotida pada trips sebesar 25.18% (Tabel 3.11), sedangkan jarak genetik asam amino yang berhasil disejajarkan dari sekitar 121 asam amino (aa) sekitar 49 aa mengalami perubahan atau variasi yang terjadi sekitar 40.1% (Tabel 3.12), dan jarak genik antar genus sebesar 0.00-0.041(Lampiran 7)
Hasil konstruksi filogeni berdasarkan sekuens DNA mtCOI memperlihatkan pengelompokkan yang terpisah antar spesies. Tidak ada perbedaan antara pohon filogeni yang dibuat berdasarkan sekuens nukleotida (Gambar 3.9 a) maupun filogeni berdasarkan urutan asam amino (Gambar 3.9 b). Nilai kepercayaan dari kedua filogeni tersebut berkisar antara 53-100%. Filogeni menunjukkan sinyal yang baik dimana ketiga spesies trips terpisah sesuai dengan spesies.
22
Tabel 3.9 Jarak genetik sekuens DNA mtCOI T. alliorum, T. hawaiiensis, dan
T. parvispinus Keterangan :(*) menunjukkan spesies dari subordo yang berbeda digunakan sebagai pembanding
(out-grup) Keterangan : (*) menunjukkan spesies dari subordo yang berbeda digunakan sebagai pembanding
24
Tabel 3.12 Variasi asam amino sekuens DNA mtCOI T. alliorum, T. hawaiiensis, dan T. parvispinus
Spesies 8 9 18 22 25 45 47 75 76 77 79 81 82 83 84 Asam amino ke- 85 86 87 88 89 90 91 92 93 95 96 97 98 99 100 101 102
T.alliorum I R S I R S I I N L R K D L S A E K I S L F V W V I L T A I L L
T.alliorum_Cina_EF213762 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
T.hawaiiensis . . T . I . N . . . A . K I . T . . . R . . . . . . . . . . . .
T.hawaiiensis_Ind_KF840096 M S T M M . N . . . A . K I . T . . . . . . . . . M . . . . . .
T.hawaiiensis_Chn_HQ605949 . . T . I . N . W F S . N . N R K N W I I C L I Y . N S N S T S
T.parvispinus . . I . I A . L . . N E N I P I . . T . . . . . . F . . . . . .
T.parvispinus_Ina_KF144115 . . I . I A . L . . N E N I P I . . T . . . . . . F . . . . . .
T.parvispinus_Ind_KM485665 M S I M M A M L . . N E N M P M . . T . . . . . . F . . . . . .
Spesies Asam amino ke-
103 104 106 109 110 111 112 113 114 115 116 117 118 119 120 121 122
T.alliorum L L L V L A G A I T I L L T D R N
T.alliorum_Cina_EF213762 . . . . . . . . . . . . . . . . .
T.hawaiiensis . . . . . . . . . . . . . . . . .
T.hawaiiensis_Ind_KF840096 . . . . . . . . . . M . . . . . .
T.hawaiiensis_Chn_HQ605949 F I T S F S R S Y Y N T F N R I E
T.parvispinus . . . . . . . . . . . . . . . . .
T.parvispinus_Ina_KF144115 . . . . . . . . . . . . . . . . .
PEMBAHASAN
Hasil penelitian menunjukkan trips yang banyak ditemukan pada tanaman hortikultura adalah Ceratothripoides brunneus Bagnall, Thrips alliorum
(Priesner), Thrips hawaiiensis (Morgan), dan Thrips parvispinus (Karny). M.
usitatus merupakan spesies yang banyak dilaporkan menyerang tanaman
hortikultura di Asia dan Afrika. Genus Ceratothripoides pertama kali perkenalkan oleh Bagnall untuk satu spesies yang ditemukan dari Ghana berdasarkan bentuk antena dari spesies betina. Sejak tahun 1918, lebih dari empat belas spesies yang dimasukkan ke dalam genus ini, enam diantaranya merupakan spesies sinonim (Mound dan Nickle 2009). C. brunneus banyak ditemukan pada berbagai tanaman di Afrika (Sevgan et al. 2009). Pada penelitian ini spesies C. brunneus ditemukan pada tanaman mawar dan Thunbergia didaerah Cianjur dan Cipanas, sedangkan Subagyo (2014) menemukan spesies ini pada tanaman terung di daerah Bogor. C.
brunneus juga dilaporkan menjadi hama baru yang ditemukan di daerah dataran
rendah Belanda dan Puerto rico (Sevgan et al. 2009). Selain itu, spesies ini banyak ditemukan di Malaysia seperti daerah Serdang, Selangor, Gua Musang, dan Kelantan dengan inang dari genus yang beragam, seperti: Asystasia, Hibiscus, Impatiens, Ocimum, Orthosiphon, Rhodomyrtus, Salvia, Solanum, Thunbergia,
dan Tabernaemontana (Mound dan Azidah 2009).
M. usitatus merupakan spesies yang berasal dari Hawaii (Reynaud et al.
2008). Spesies M. usitatus umum ditemukan di Asia seperti Indonesia, Taiwan, dan Malaysia. Tanaman inang dari M. usitatus bervariasi seperti tanaman dari genus Vigna dan Sesbania serta pada beberapa tanaman lain (Mound dan Azidah 2009). M. usitatus dilaporkan menyerang tanaman Fabaceae dan Solanaceae seperti: kacang kedelai, kacang merah, kacang hijau, kacang tanah, kacang polong, kacang panjang, kacang tunggak, puri, dan bengkoang di Taiwan serta Malaysia (Chang 1991; Fauziah dan Saharan 1991). Bahkan di Taiwan M.
usitatus ditemukan berasosiasi dengan 28 jenis tanaman (Chang 1991). M.
usitatus merupakan hama penting pada tanaman kacang hijau pada fase vegetatif.
Pada tingkat serangan yang parah dapat menyebabkan kehilangan hasil sebesar 13-64% (Indiati 2012). Menurut laporan terbaru, M. usitatus merupakan satu dari tiga spesies yang menjadi vektor tospovirus pada pollen bunga matahari
(Helianthus annuus) dan Marigold (Tagetes patula ) (Rao 2015).
T. alliorum, T. hawaiiensis, dan T. parvispinus merupakan trips yang
banyak dilaporkan menjadi hama serius dipertanaman dengan sebaran dan inang yang luas. T. alliorum pada penelitian ini ditemukan pada tanaman bawang di daerah Cipanas. Hal ini sama seperti laporan Sartiami dan Mound (2013) menemukan spesies T. alliorum pada tanaman bawang di daerah Cianjur, sedangkan Subagyo (2014) tidak berhasil menemukan spesies T. alliorum. Selain
T. alliorum, T. hawaiiensis juga dilaporkan sebagai hama umum di Taiwan.
Spesies ini menyerang bunga pada tanaman pertanian dan bunga potong. Selain itu, T. hawaiiensis juga menyerang menyebabkan kerusakan pada 22 jenis tanaman, termasuk pada bunga pisang. Daerah persebaran spesies ini antara lain: Asia tenggara, sebagian Afrika, Austrasia, dan Kepulauan Pasifik (Chang 1991; Mound dan Kibby 1998).
Pada penelitian ini, dari ketiga spesies yang ditemukan T. parvispinus